
INTERNATIONAL PROGRAMME ON CHEMICAL SAFETY
ENVIRONMENTAL HEALTH CRITERIA 149
CARBENDAZIM
This report contains the collective views of an international group of
experts and does not necessarily represent the decisions or the stated
policy of the United Nations Environment Programme, the International
Labour Organisation, or the World Health Organization.
Published under the joint sponsorship of
the United Nations Environment Programme,
the International Labour Organisation,
and the World Health Organization
First draft prepared by IPCS staff, using texts made available
by Dr L.W. Hershberger and Dr G.T. Arce, Wilmington, Delaware, USA
World Health Orgnization
Geneva, 1993
The International Programme on Chemical Safety (IPCS) is a
joint venture of the United Nations Environment Programme, the
International Labour Organisation, and the World Health
Organization. The main objective of the IPCS is to carry out and
disseminate evaluations of the effects of chemicals on human health
and the quality of the environment. Supporting activities include
the development of epidemiological, experimental laboratory, and
risk-assessment methods that could produce internationally
comparable results, and the development of manpower in the field of
toxicology. Other activities carried out by the IPCS include the
development of know-how for coping with chemical accidents,
coordination of laboratory testing and epidemiological studies, and
promotion of research on the mechanisms of the biological action of
chemicals.
WHO Library Cataloguing in Publication Data
Carbendazim.
(Environmental health criteria ; 149)
1.Benzimidazoles - adverse effects 2.Benzimidazoles - toxicity
3.Fungicides, Industrial - adverse effects 4.Fungicides, Industrial
-toxicity I.Series
ISBN 92 4 157149 7 (NLM Classification: WA 240)
ISSN 0250-8634
The World Health Organization welcomes requests for permission
to reproduce or translate its publications, in part or in full.
Applications and enquiries should be addressed to the Office of
Publications, World Health Organization, Geneva, Switzerland, which
will be glad to provide the latest information on any changes made
to the text, plans for new editions, and reprints and translations
already available.
(c) World Health Organization 1993
Publications of the World Health Organization enjoy copyright
protection in accordance with the provisions of Protocol 2 of the
Universal Copyright Convention. All rights reserved.
The designations employed and the presentation of the material
in this publication do not imply the expression of any opinion
whatsoever on the part of the Secretariat of the World Health
Organization concerning the legal status of any country, territory,
city or area or of its authorities, or concerning the delimitation
of its frontiers or boundaries.
The mention of specific companies or of certain manufacturers'
products does not imply that they are endorsed or recommended by the
World Health Organization in preference to others of a similar
nature that are not mentioned. Errors and omissions excepted, the
names of proprietary products are distinguished by initial capital
letters.
CONTENTS
ENVIRONMENTAL HEALTH CRITERIA FOR CARBENDAZIM
1. SUMMARY AND CONCLUSIONS
1.1. Summary
1.1.1. Identity, physical and chemical properties, and
analytical methods
1.1.2. Sources of human and environmental exposure
1.1.3. Environmental transport, distribution and
transformation
1.1.4. Environmental levels and human exposure
1.1.5. Kinetics and metabolism
1.1.6. Effects on laboratory mammals and in vitro
test systems
1.1.7. Effects on humans
1.1.8. Effects on other organisms in the laboratory
and field
1.2. Conclusions
2. IDENTITY, PHYSICAL AND CHEMICAL PROPERTIES, AND ANALYTICAL
METHODS
2.1. Identity
2.1.1. Primary constituent
2.1.2. Technical product
2.2. Physical and chemical properties
2.3. Analytical methods
3. SOURCES OF HUMAN AND ENVIRONMENTAL EXPOSURE
3.1. Natural occurrence
3.2. Anthropogenic sources
3.2.1. Uses
4. ENVIRONMENTAL TRANSPORT, DISTRIBUTION AND TRANSFORMATION
4.1. Transport and distribution between media
4.1.1. Air
4.1.2. Water
4.1.3. Soil
4.1.4. Leaching
4.1.5. Crop uptake
4.2. Transformation
4.2.1. Soil biodegradation
4.2.2. Abiotic degradation
4.2.3. Bioaccumulation
5. ENVIRONMENTAL LEVELS AND HUMAN EXPOSURE
5.1. Environmental levels
5.1.1. Air, water and soil
5.1.2. Food and feed
5.1.3. Terrestrial and aquatic organisms
5.2. General population exposure
5.2.1. Sweden
5.2.2. The Netherlands
5.2.3. National maximum residue limits
5.3. Occupational exposure during manufacture,
formulation, or use
5.3.1. Exposure during manufacture
5.3.2. Exposure during use
6. KINETICS AND METABOLISM
6.1. Absorption
6.2. Distribution and accumulation
6.3. Metabolic transformation
6.4. Elimination and excretion
6.5. Reaction with body components
7. EFFECTS ON LABORATORY MAMMALS AND IN VITRO TEST SYSTEMS
7.1. Single exposure
7.2. Short-term exposure
7.2.1. Gavage
7.2.2. Feeding
7.2.2.1 Rat
7.2.2.2 Dog
7.2.3. Dermal
7.3. Skin and eye irritation; sensitization
7.3.1. Dermal
7.3.2. Eye
7.3.3. Sensitization
7.4. Long-term exposure
7.4.1. Rat
7.4.2. Dog
7.4.3. Mouse
7.5. Reproduction, embryotoxicity and teratogenicity
7.5.1. Reproduction
7.5.2. Embryotoxicity and teratogenicity
7.6. Mutagenicity and related end-points
7.7. Carcinogenicity
7.8. Neurotoxicity
7.9. Toxicity of contaminants
7.10. Mechanisms of toxicity - mode of action
8. EFFECTS ON HUMANS
8.1. General population exposure
8.2. Occupational exposure
9. EFFECTS ON OTHER ORGANISMS IN THE LABORATORY AND FIELD
9.1. Microorganisms
9.2. Aquatic organisms
9.3. Terrestrial organisms
9.4. Population and ecosystem effects
10. EVALUATION OF HUMAN HEALTH RISKS AND EFFECTS ON ENVIRONMENT 95
10.1. Evaluation of human health risks
10.2. Evaluation of effects on the environment
10.3. Conclusions
11. FURTHER RESEARCH
12. PREVIOUS EVALUATIONS BY INTERNATIONAL BODIES
REFERENCES
RESUME ET CONCLUSIONS
RESUMEN Y CONCLUSIONES
WHO TASK GROUP ON ENVIRONMENTAL HEALTH CRITERIA FOR BENOMYL AND
CARBENDAZIM
Members
Dr G. Burin, Office of Pesticide Programmes, US Environmental
Protection Agency, Washington, D.C., USA
Dr R. Cooper, Reproductive Toxicology Branch, US Environmental
Protection Agency, Research Triangle Park, North Carolina, USA
Dr I. Desi, Department of Public Health, Albert Szent-Györgyi
University Medical School, Szeged, Hungary
Dr S. Dobson, Institute of Terrestrial Ecology, Monks Wood, Abbots
Ripton, Huntingdon, United Kingdom
Dr A. Helweg, Department for Pesticide Analysis and Ecotoxicology,
Danish Research Service for Plant and Soil Science,
Flakkebjerg, Slagelse, Denmark
Dr M. Lotti, Institute of Occupational Medicine, University of
Padua, Padua, Italy (Chairman)
Dr K. Maita, Toxicology Division, Institute of Environmental
Toxicology, Kodaira-Shi, Tokyo, Japan
Dr F. Matsumura, Department of Environmental Toxicology, Institute
of Toxicology and Environmental Health, University of
California, Davis, California, USA
Dr T.K. Pandita, Microbiology and Cell Biology Laboratory, Indian
Institute of Science, Bangalore, Indiaa
Dr C. Sonich-Mullin, Environmental Criteria and Assessment Office,
US Environmental Protection Agency, Cincinnati, Ohio, USA
Dr P.P. Yao, Institute of Occupational Medicine, Chinese Academy of
Preventive Medicine, Beijing, China
Secretariat
Dr B.H. Chen, International Programme on Chemical Safety, World
Health Organization, Geneva, Switzerland ( Secretary)
Dr L.W. Hershberger, Dupont Agricultural Products, Walker's Mill,
Barley Mill Plaza, Wilmington, Delaware, USA
Mr P. Howe, Institute of Terrestrial Ecology, Monks Wood, Abbots
Ripton, Huntington, United Kingdom
a Invited but unable to attend the meeting
NOTE TO READERS OF THE CRITERIA MONOGRAPHS
Every effort has been made to present information in the
criteria monographs as accurately as possible without unduly
delaying their publication. In the interest of all users of the
Environmental Health Criteria monographs, readers are kindly
requested to communicate any errors that may have occurred to the
Director of the International Programme on Chemical Safety, World
Health Organization, Geneva, Switzerland, in order that they may be
included in corrigenda.
* * *
A detailed data profile and a legal file can be obtained from
the International Register of Potentially Toxic Chemicals, Palais
des Nations, 1211 Geneva 10, Switzerland (Telephone No. 7988400 or
7985850).
ENVIRONMENTAL HEALTH CRITERIA FOR CARBENDAZIM
A WHO Task Group on Environmental Health Criteria for Benomyl
and Carbendazim, sponsored by the US Environmental Protection
Agency, met in Cincinnati, USA, from 14 to 19 September 1992. On
behalf of the host agency, Dr T. Harvey opened the meeting and
welcomed the participants. Dr B.H. Chen of the International
Programme on Chemical Safety (IPCS) welcomed the participants on
behalf of the Director, IPCS, and the three IPCS cooperating
organizations (UNEP/ILO/WHO). The Task Group reviewed and revised
the draft criteria monograph and made an evaluation of the risks for
human health and the environment from exposure to carbendazim.
The first draft of this monograph was prepared by the staff of
IPCS, using texts made available by Dr L.W. Hershberger and Dr G.T.
Arce, Wilmington, Delaware, USA. The second draft was prepared by Dr
L.W. Hershberger and Dr B.H. Chen incorporating comments received
following the circulation of the first draft to the IPCS Contact
Points for Environmental Health Criteria monographs. Dr M. Lotti
(Institute of Occupational Medicine, University of Padua, Italy)
made a considerable contribution to the preparation of the final
text. Dr B.H. Chen and Dr P.G. Jenkins, both members of the IPCS
Central Unit, were responsible for the overall scientific content
and technical editing, respectively.
The efforts of all who helped in the preparation and
finalization of the monograph are gratefully acknowledged.
Financial support for the meeting was provided by the US
Environmental Protection Agency, Cincinnati, USA.
ABBREVIATIONS
a.i. active ingredient
BSP Bromosulfophthalein
HPLC high-performance liquid chromatography
Koc Distribution coefficient between pesticide adsorbed
to soil organic carbon and pesticide in solution
Kom Distribution coefficient between pesticide adsorbed
to soil organic matter and pesticide in solution
NOEC no-observed-effect concentration
NOEL no-observed-effect level
2-AB 2-aminobenzimidazole
5-HBC methyl (5-hydroxy-1H-benzimidazol-2-yl)-carbamate
1. SUMMARY AND CONCLUSIONS
1.1 Summary
1.1.1 Identity, physical and chemical properties, and analytical
methods
Carbendazim, a white crystalline solid, is a systemic fungicide
belonging to the benzimidazole family. It melts at approximately 250
°C and has a vapour pressure of < 1 x 10-7 Pa (< 1 x 10-9
mbar) at 20 °C. Carbendazim is essentially insoluble in water (8
mg/litre solubility) at pH 7 and 20 °C. It is stable under normal
storage conditions.
Residual and environmental analyses are performed by extraction
with an organic solvent and the extract is purified by a
liquid-liquid partitioning procedure. Measurement of residues may be
determined by HPLC or immunoassay.
1.1.2 Sources of human and environmental exposure
Carbendazim is the most widely used member of the benzimidazole
family of fungicides. It is formulated as an aqueous dispersion,
aqueous suspension, flowable water-dispersible granule and a
wettable powder.
1.1.3 Environmental transport, distribution and transformation
Benomyl is rapidly converted to carbendazim in the environment,
with half-lives of 2 and 19 h in water and in soil, respectively.
Data from studies on both benomyl and carbendazim are therefore
relevant for the evaluation of environmental effects.
Carbendazim is decomposed in the environment with half-lives of
6 to 12 months on bare soil, 3 to 6 months on turf, and half-lives
in water of 2 and 25 months under aerobic and anaerobic conditions,
respectively. Carbendazim is mainly decomposed by microorganisms;
2-aminobenzimidazole (2-AB) is the major degradation product and is
further decomposed by microbial activity. When phenyl-14C-labelled
benomyl was decomposed, only 9% of the 14C label was evolved in
CO2 during 1 year of incubation, the remaining 14C being
recovered mainly as carbendazim and bound residues. The fate of a
possible degradation product (1,2-diaminobenzene) may shed further
light on the degradation pathway of benzimidazole fungicides in the
environment.
Field and column studies have shown that carbendazim remains in
the soil surface layer. No determination of carbendazim adsorption
in soil is available, but it is likely to be as strongly adsorbed to
soil as benomyl (Koc values ranking from 1000 to 3600). Log Kow
values for benomyl and carbendazim are 1.36 and 1.49, respectively.
A risk of leaching was not apparent when this was evaluated in
a screening model based on adsorption and persistence data. This
statement is supported by analysis of well water in the USA, where
carbendazim has not been found in any of 212 wells (limit of
detection not available). Surface run-off of benomyl and carbendazim
is expected to consist only of fungicide adsorbed to soil particles,
and the fungicides are likely to be strongly adsorbed to sediments
in the aqueous environment.
Carbendazim is hydrolysed to 2-AB. This is also the primary
metabolite in soil and plants.
In animal systems, carbendazim is metabolized to (5-hydroxy-
1H-benzimidazol-2-yl)-carbamate (5-HBC) and other polar metabolites,
which are rapidly excreted. Carbendazim has not been observed to
accumulate in any biological system.
1.1.4 Environmental levels and human exposure
There appear to be no environmental monitoring data for
carbendazim. However, the following can be summarized from
environmental fate studies.
Due to the fact that they are stable for several weeks on plant
material, benomyl and carbendazim may become accessible to organisms
feeding on leaf litter. Soil and sediments may contain residues of
carbendazim for up to 3 years. However, the strong adsorption of
carbendazim to soil and sediment particles reduces the exposure for
terrestrial and aquatic organisms.
Primary exposure for the general human population will be from
residues of benomyl and carbendazim on food crops. Dietary exposure
analysis in the USA (combined benomyl and carbendazim) and the
Netherlands (carbendazim) estimated the expected mean intake to be
about one-tenth of the recommended Acceptable Daily Intake (ADI) of
0.02 mg/kg body weight for benomyl and 0.01 mg/kg body weight for
carbendazim.
Occupational exposure during manufacture is below the Threshold
Limit Value established for benomyl. Agricultural workers engaged in
pesticide mixing and loading or in re-entering benomyl-treated
fields are expected to be exposed to dermal contact of a few mg
benomyl per h. This type of exposure could be reduced by the use of
protective devices. Furthermore, since dermal absorption is expected
to be low, the probability of systemic toxicity of benomyl through
this route is very low.
1.1.5 Kinetics and metabolism
Carbendazim is well absorbed (80-85%) after oral exposure but
much less so by dermal exposure. Absorbed carbendazim is metabolized
into many compounds within the organism. The main metabolites are
5-HBC and 5,6-HOBC-N-oxides. Minor metabolites are 5,6-DHBC-S and
5,6-DHBC-G.
The tissue distribution of carbendazim showed no
bioconcentration. In the rat, the highest concentration after oral
carbendazim administration (< 1% of the dose) occurred in the
liver. It was distributed as carbendazim in the mitochondria, 5-HBC
in the cytostol, and 2-AB in the microsomes. Carbendazim and its
metabolites were also found in the kidney of hens and cows; but no
significant levels were detected in other tissues. After carbendazim
was fed to lactating cows, small amounts of 5-HBC and 4-HBC were
found in the milk.
Carbendazim is excreted in the urine and faeces within 72 h
after oral dosing in rats.
In rats and mice, high doses of carbendazim, both in the diet
and by gavage, affect certain liver microsomal enzymes. Styrene-7,8-
hydrolase and epoxide hydrolase were induced whereas 7-
hydroxycoumarin O-deethylase activity was found to be reduced.
Cytosolic glutathione S-transferase activity was also induced.
1.1.6 Effects on laboratory mammals and in vitro test systems
1.1.6.1 Single exposure
Carbendazim has low acute toxicity. The LD50 values range
from > 2000 to 15 000 mg/kg in a wide variety of test animals and
routes of administration. However, significant adverse reproductive
effects have been noted following a single exposure (see section
1.1.6.5).
1.1.6.2 Short-term exposure
Dietary administration of carbendazim for up to 90 days
produced slight effects on liver weight in female rats exposed to
360 mg/kg body weight per day. In a 90-day gavage study in the rat,
the NOEL was 16 mg/kg per day based on hepatotoxicity. Short-term
feeding studies on dogs were not adequate for establishing a NOEL. A
10-day dermal study in the rabbit revealed no systemic toxicity at
the only dose tested (200 mg/kg).
1.1.6.3 Skin and eye irritation and sensitization
Application to the skin of the rabbit and guinea-pig produced
no irritation or skin sensitization. Application to the eyes of
rabbits produced moderate or mild conjunctival irritation.
1.1.6.4 Long-term exposure
Male and female rats fed 2500 mg/kg diet showed reduced
erythrocyte count and haemoglobin and haematocrit values. No
liver-related toxicity was noted. Male rats fed 2500 mg/kg diet or
more presented a marginal increase in diffuse testicular atrophy and
prostatitis. The NOEL in the rat is 500 mg/kg diet. Elevated serum
cholesterol and alkaline phosphatase activity and other indications
of hepatotoxicity were observed in dogs fed a diet containing 500 mg
carbendazim/kg for 1 year or longer. The NOEL in the dog is 300
mg/kg diet.
Male and female mice fed 5000 mg/kg diet showed increased
absolute liver weight. There was also significant centrilobular
hypertrophy, necrosis and swelling of the liver in male mice fed
1500 mg/kg diet.
1.1.6.5 Reproduction, embryotoxicity and teratogenicity
Carbendazim was without adverse effects on reproduction when it
was fed to rats in a three-generation reproduction study at levels
up to and including 500 mg/kg diet. Male fertility was depressed in
rats when carbendazim (200 mg/kg per day) was administered by gavage
for 85 days. A dose of 50 mg/kg body weight per day in this study
caused a significant decrease in epididymal sperm count.
Following a single oral dose to rats, histological examination
revealed early (0-2 days) disruption of spermatogenesis with
occlusion of efferent ducts and increased testicular weights at 100
mg/kg body weight. No effect was observed at 50 mg/kg in this single
dose study. These effects persisted until day 70 in rats treated
with 400 mg/kg.
Carbendazim caused an increase in malformations and anomalies
in rats when administered at daily dose levels greater than 10 mg/kg
on days 7-16 of gestation. There was a slightly decreased rate of
implantation in rabbits administered 20 and 125 mg/kg per day on
days 7-19 of gestation and an increased incidence of resorption at
125 mg/kg per day. Maternal toxicity was observed at 20 mg/kg per
day and 125 mg/kg per day in the rat and rabbit, respectively.
In addition to decreased pregnancy rate and increased early
resorptions in the rat, there were significant reductions in fetal
weights at 20 and 90 mg/kg per day and a significant increase in
fetal malformations at 90 mg/kg per day. These consisted primarily
of hydrocephaly, microphthalmia, anophthalmia, malformed scapulea
and axial skeletal malformations (vertebral, rib and sternebral
fusions, exencephaly, hemivertebrae and rib hyperplasia). However,
in the rabbit there were no significant malformations.
1.1.6.6 Mutagenicity and related end-points
Assays in mammalian and non-mammalian systems in vitro and
in vivo and in somatic cells as well as in germ cells show that
carbendazim does not interact with DNA, induce point mutation or
cause germ cell mutation.
Carbendazim does, however, cause numerical chromosome
aberrations (aneuploidy and/or polyploidy) in experimental systems,
both in vitro and in vivo.
1.1.6.7 Carcinogenicity
Benomyl and carbendazim feeding resulted in an increase in the
incidence of hepatocellular tumours in CD-1 and SPF Swiss mice.
A carcinogenicity study of carbendazim using CD-1 mice showed a
statistically significant dose-related increase in the incidence of
hepatocellular neoplasia in females. There was also a statistically
significant increase in the mid-dose (1500 mg/kg diet) males, but
not in the high-dose males because of a high mortality rate. A
carcinogenicity study of carbendazim in a genetically related mouse
strain, SPF mice (Swiss random strain) at doses of 0, 150, 300 and
1000 mg/kg diet (increased to 5000 mg/kg during the study) showed an
increase in the incidence of combined hepatocellular adenomas and
carcinomas. A study carried out in NMRKf mice at dose levels of 0,
50, 150, 300 and 1000 mg/kg diet of carbendazim (increased to 5000
mg/kg during the study) showed no carcinogenic effects. Benomyl or
carbendazim caused liver tumours in two strains of mice (CD-1 and
SPF), both of which have a high spontaneous rate of liver tumours.
In contrast, carbendazim is not carcinogenic in NMRKf mice, which
have a low spontaneous rate of such tumours.
Carcinogenicity studies of both benomyl and carbendazim in rats
were negative.
1.1.6.8 Mechanism of toxicity - mode of action
The biological effects of benomyl and carbendazim result from
their interaction with cell microtubules. These structures are
involved in vital functions such as cell division, which is
inhibited by benomyl and carbendazim. Benomyl and carbendazim
toxicities in mammals are linked to microtubular dysfunction.
Benomyl and carbendazim, as well as other benzimidazole
compounds, display species-selective toxicity. This selectivity is,
at least in part, explained by the different binding of benomyl and
carbendazim to tubulins of target and non-target species.
1.1.7 Effects on humans
No adverse effects on human health have been reported.
1.1.8 Effects on other organisms in the laboratory and field
Carbendazim has little effect on soil microbial activity at
recommended application rates. Some adverse effects have been
reported for groups of fungi.
The 72-h EC50, based on total growth, for the green alga
Selenastrum capricornutum was calculated to be 1.3 mg/litre; the
NOEC was 0.5 mg/litre. The toxicity of carbendazim to aquatic
invertebrates and fish varies widely, with 96-h LC50 values
ranging from 0.007 mg/litre for the channel catfish to 5.5 mg/litre
for the bluegill sunfish. In a 21-day test on Daphnia magna the
onset of reproduction was significantly delayed at 0.025 mg/litre;
the NOEC was 0.013 mg/litre.
Carbendazim is toxic to earthworms in laboratory experiments at
realistic exposure concentrations and from recommended use in the
field. It is "relatively non-toxic" to honey-bees and of low
toxicity to birds.
1.2 Conclusions
Benomyl causes dermal sensitization in humans. Benomyl and
carbendazim represent a very low risk for acute poisoning in humans.
Given the current exposures and the low rate of dermal absorption of
benomyl and carbendazim, it is unlikely that they would cause
systemic toxicity effects either in the general population or in
occupationally exposed subjects. These conclusions are drawn from
animal data and the limited human data, but these extrapolations are
supported by the understanding of the mode of action of carbendazim
and benomyl in both target and non-target species.
Further elucidation of the mechanism of toxicity of carbendazim
and benomyl in mammals will perhaps enable a better definition of
no-observed-effect levels. Binding studies on tubulins of target
cells (testis and embryonic tissues) will facilitate inter-species
comparisons.
Carbendazim is strongly adsorbed to soil organic matter and
persists in soil for up to 3 years. It also persists on leaf
surfaces and, therefore, in leaf litter. Earthworms have been shown
to be adversely affected (population and reproductive effects) at
recommended application rates. There is no information on other soil
or litter arthropods that would be similarly exposed.
The high toxicity to aquatic organisms in laboratory tests is
unlikely to be seen in the field because of the low bioavailability
of sediment-bound residues of carbendazim. However, no information
is available on sediment-living species, which would receive the
highest exposure.
2. IDENTITY, PHYSICAL AND CHEMICAL PROPERTIES, AND ANALYTICAL METHODS
2.1 Identity
2.1.1 Primary constituent
Common name: Carbendazim (BSI, ISO)
Chemical structure:
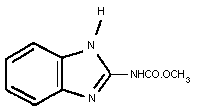 Empirical formula: C9H9N3O2
Relative molecular mass: 191.2
CAS chemical name: Methyl (1H-benzimidazol-2-yl)carbamate
IUPAC chemical name: Methyl benzimidazole-2-ylcarbamate
CAS Registry number: 10605-21-7
Synonyms: carbendazol (ZMAF), methyl-2-benzimidazole
carbamate (MBC, MCB, BCM, BMC)
2.1.2 Technical product
Major trade names:
Carbendazim, Delsene, Bavistin, Corbel, Konker, Bendazim,
Derosal, Kombat, Kemdazin, Carbendor, Hoe 017411, Cekudazim,
Equitdazin, Aimcozim (Some of these are formulations with other
pesticides.)
Purity: > 98% (FAO specifications)
Impurities: 2,3-diaminophenazine (DAP), 2-amino-3-
hydroxyphenazine (HAP)
2.2 Physical and chemical properties
Table 1. Some physical and chemical properties of carbendazim
Physical state Crystalline solid
Colour White
Odour Negligible
Table 1 (contd).
Melting point/boiling
point/flash point Melts at -250 °C
Explosion limits LEL = 0.13 g/litre in air
Vapour pressure < 1 x 10-7 Pa (< 1 x 10-9 mbar) at 20 °C
Density 0.27 g/cm3 (loose); 0.62 g/cm3 (packed)
Log n-octanol/water 1.49
partition coefficient
Solubility in water pH 4 28 mg/litre
(at 20 °C) pH 7 8 mg/litre
pH 8 7 mg/litre
Solubility in organic solvents
Hexane 0.5 mg/litre
Benzene 36 mg/litre
Dichlorom ethane 68 mg/litre
Ethanol 300 mg/litre
Dimethylformamide 5000 mg/litre
Acetone 300 mg/litre
Chloroform 100 mg/litre
Henry's constant 1.02 x 10-9 atm-m3/mol at 20 °C
2.3 Analytical methods
Methods for determining carbendazim and its by-product residues
in plant and animal tissue and in soil involve isolation of the
residue by extraction with an organic solvent and purification of
the extract by a liquid-liquid partitioning procedure. Measurement
of the residues may be determined by procedures using high-speed
cation exchange liquid chromatography, reversed phase HPLC, and
immunoassay. Recoveries of carbendazim and 2-aminobenzimidazole
(2-AB) from various types of soils average 88 and 71%, respectively.
The lower limit of sensitivity of the method is 0.05 ppm for each of
these components. The recoveries and sensitivities in the case of
plant tissues are similar. Table 2 outlines various methods for
soil, water, plants and animal tissue.
Table 2. Analytical methods for carbendazim
Analytical method Medium Detection limit Comments Reference
Strong cation exchange/HPLC soil 0.05 mg/kg acidic methanol extraction converts Kirkland et al. (1973)
residual benomyl to carbendazim
Strong cation exchange/HPLC plant 0.05 mg/kg acidic methanol extraction converts Kirkland et al. (1973)
residual benomyl to carbendazim
Strong cation exchange/HPLC animal 0.01 mg/kg (milk) acidic aqueous hydrolysis followed Kirkland (1973)
0.05 mg/kg by organic extraction converts
(tissue) benomyl to carbendazim and frees
metabolites from conjugates
Reversed phase HPLC water 9.0 x 10-6 g/litre on-line HPLC with preconcentration; Marvin et al. (1991)
benomyl and carbendazim
determined separately
Reversed phase blueberries 0.03 mg/kg acidic methanol extraction converts Bushway et al. (1991)
HPLC/fluorescence detection residual benomyl to carbendazim
Radioimmunoassay plant 0.05-1.0 mg/kg ethyl acetate extraction converts Newsome & Shields
(dependent on crop) residual benomyl to carbendazim (1981)
Enzyme-linked immunosorbent plant 0.50 mg/kg ethyl acetate extraction converts Newsome & Collins
assay (ELISA) residual benomyl to carbendazim (1987)
3. SOURCES OF HUMAN AND ENVIRONMENTAL EXPOSURE
3.1 Natural occurrence
Carbendazim does not occur naturally.
3.2 Anthropogenic sources
3.2.1 Uses
Carbendazim is a fungicide in its own right as well being as
the main metabolite of other fungicides such as benomyl and
thiophanate-methyl.
Carbendazim is used to control a wide range of fungi, including
Ascomycetes, Fungi Imperfecti and numerous Basidiomycetes, which
result in plant diseases such as: leaf spots, blotches and blights;
fruit spots and rots; sooty molds; scabs; bulb, corn and tuber
decays; blossom blights; powdery mildews; certain rusts; and common
soilborne crown and root rots. It is used on cereals, cotton,
grapes, bananas and other fruit, ornamentals, plantation crops,
sugar beet, soybeans, tobacco, turf, vegetables, mushrooms, and many
other crops under most climatic conditions worldwide. Registered
carbendazim usage specifies rates from 0.2 to 2.0 kg a.i./ha and
applications from once per year to spray intervals ranging from 7 to
14 days (FAO/WHO, 1985b; 1988b).
A key limitation to the use of carbendazim and other
benzimidazoles is the development of fungal resistance. Resistance
management can be achieved by using carbendazim in combination as a
tank mix or alternately with other non-benzimidazole fungicides
(Delp, 1980; Staub & Sozzi, 1984).
Carbendazim is formulated as an aqueous dispersion, aqueous
suspension, flowable water dispersible granules and a wettable
powder.
In 1991, the estimated worldwide sales of benomyl was US$ 290
million. This was about 50% of the worldwide market for
benzimidazole products. Carbendazim (20%) and thiophanatemethyl
(20%) account for most of the rest of the benzimidazole market
(County NatWest Woodmac, 1992).
4. ENVIRONMENTAL TRANSPORT, DISTRIBUTION AND TRANSFORMATION
4.1 Transport and distribution between media
4.1.1 Air
Carbendazim has a vapour pressure of < 1 x 10-7 Pa (< 1 x
10-9 mbar) at 20 °C, an aqueous solubility of 8 mg/litre at 20 °C
and pH 7, and a Henry's constant of approx. 1.02 x 10-9
atm-m3/mol at 20 °C. It is essentially non-volatile from water
surfaces.
4.1.2 Water
Anaerobic aquatic degradation studies of [phenyl(U)-14C]-
benomyl in pond water and sediment showed that more than 98% of the
carbendazim residues partitioned into the sediment after 7 days. The
half-life of carbendazim was 743 days. After one year 36% of the
applied radioactivity was bound to the sediment (Arthur et al.,
1989a).
An aerobic aquatic degradation study of [phenyl(U)-14C]-
benomyl in pond water and sediment showed that carbendazim had a
half-life of 61 days under nonsterile conditions. After 30 days, 22%
of the applied radioactivity was bound to the sediments and < 1% of
the applied radioactivity was evolved as carbon dioxide (Arthur et
al., 1989b).
4.1.3 Soil
In greenhouse studies to determine run-off and leaching of
[2-14C]-carbendazim on soil, a container of Keyport silt loam was
treated with labelled carbendazim, at a rate of 11 kg a.i./ha, by
spraying the upper one-third (0.093 m2) of the plot and was
allowed to stand for 24 h. Artificial rain was then applied (3.75 cm
the first day after treatment and 2.5 cm on the third and seventh
days). All water that ran off or leached through the soil was
collected and analysed for total 14C. Soil in the plot was divided
into layers for analysis, air dried and analysed separately for
total 14C. After each of the three rain applications, 0.05-0.39%
of the applied 14C was found in run-off water; < 0.01% was found
in the leach water after the first two artificial rains and 0.19%
after the third one. Soil analyses showed that 90.6% of the applied
activity remained in the treated area and 93.1% in the top 10 cm of
soil (Rhodes & Long, 1983).
4.1.4 Leaching
To evaluate the risk of pollution of ground and drainage water,
screening models based on adsorption and persistence can be used,
together with existing analyses of groundwater samples. Gustafson
(1989) proposed the use of the equation GUS = log T´ (4 - log
Koc); GUS values < 1.8 = "improbable leachers", GUS values of
1.8-2.8 = "transition" and GUS values > 2.8 = "probable leachers".
For benomyl, Kom values of 550, 620, 2100 and 1100 (mean 1093)
were found in four different soils (Priester, 1985). A Kom of 1093
is equal to a Koc of 1857 since Koc = Kom x 1.7. The half-life
of 320 days given by Marsh & Arthur (1989) seems in good agreement
with field half-lives of 6 to 12 months (Baude et al., 1974).
When the calculation of the GUS value is based on a Koc of
1857 and a T0.5 of 320 days, a value of 1.83 is obtained.
According to this value, benomyl/carbendazim lies between the
"improbable leachers" and "transition", and, therefore, would not be
expected to occur in ground water. The adsorption of benomyl and of
carbendazim is expected to be of the same order of magnitude since
the Kow values are almost identical (log Kow = 1.49 and 1.36 for
carbendazim and benomyl, respectively). In groundwater studies in
the USA (Parsons & Witt, 1988), benomyl was not found in any of 495
wells tested and carbendazim not in any of 212 wells (detection
limit not reported).
In an EEC survey (Fielding, 1992), the presence of carbendazim
in groundwater in the Netherlands and in Italy was investigated.
Carbendazim was found in one of two samples from the Netherlands
(0.1 µg/litre), and the level was above 0.1 µg/litre in 23 of 70
samples in Italy. Detection of the non-polar DDT and lindane in many
wells in the Italian study may indicate macropore transport or
artifacts such as direct pollution of wells.
4.1.5 Crop uptake
Various greenhouse and outdoor tests with carbendazim indicate
that it remains on plant surfaces as the major component of the
total residue (Baude et al., 1973).
A greenhouse crop-rotation study was undertaken by application
of [2-14C]carbendazim to a loamy sand soil, followed by aging
periods of 30, 120 or 145 days. The crops studied were beets, barley
and cabbage. Radioactivity did not accumulate in these crops grown
to maturity in a loamy sand soil treated 30 days earlier (1.1 kg
a.i./ha) or 120 to 145 days earlier (3.4 kg a.i./ha). Accumulation
factors, calculated as the ratio of radioactivity in the crop to
that in the corresponding soil, were very low in beet foliage (0.04)
and beet roots (0.03), low in cabbage and barley grain (0.2) and
ranged from 0.9 to 1.2 in barley straw (Rhodes, 1987).
Alfalfa, soybean and ryegrass, which were grown in 0.028 m3
(1 cu ft) containers in a greenhouse in soil treated with an 80:20
mixture of carbendazim and 2-aminobenzimidazole (2-AB), contained
small but detectable residues of both compounds. Both 14C-labelled
and non-labelled mixtures were applied at the rate of 2.2 kg/ha
uniformly incorporated in the 0-10 cm layer of soil. In the 14C
studies, alfalfa contained total 14C residues equivalent to
0.13-0.30 mg/kg of carbendazim/2-AB. Soybean plants contained
0.32-0.53 mg/kg and ryegrass (20-183 days after planting) contained
0.09-0.19 mg/kg. Each plant contained approximately equal amounts of
carbendazim, 2-AB and a polar unknown fraction. Alfalfa from the
non-labelled series contained 0.05 and 0.08 mg/kg, respectively, of
carbendazim and 2-AB at the first cutting and < 0.05 mg/kg of each
compound at the second and third cuttings. Soybean plants contained
< 0.1 mg 2-AB/kg and 0.59 mg carbendazim/kg. Ryegrass from six
cuttings (20-149 days after planting) contained 0.08-0.48 mg
carbendazim/kg and < 0.05 mg 2-AB/kg. All data were calculated on a
fresh weight basis (Rhodes et al., 1983).
4.2 Transformation
4.2.1 Soil biodegradation
Aerobic degradation studies of labelled [2-14C]-2-AB, the
primary degradation product of carbendazim in soil, showed that
14C evolution increased exponentially from 1 to 22 °C, reached a
maximum at 22 °C, remained almost constant up to 35 °C, then became
almost zero at 40 °C, when the soil water content was 100% of field
capacity. At 25 °C, 14C evolution increased exponentially with an
increase in the field capacity of water from 28 to 94%. These and
other results indicate the presence of organisms that are able to
decompose 2-AB (Helweg, 1979).
Laboratory studies on two types of soil under anaerobic
conditions using [2-14C]-carbendazim showed only a small amount of
2-AB (< 0.1%) and no other degradation products (< 0.05%).
Re-incorporation of 14C into soil humus was indicated by
fractionation studies, which showed that the unextracted 14C
residue was widely distributed in various organic soil components
(Han, 1983b).
The persistence of carbendazim was monitored in nonsterile and
sterile Keyport silt loam soil after it was treated with
[phenyl(U)-14C]-benomyl at a concentration of approximately 7.0
mg/kg. Distilled water was added to each sample until it reached 75%
of its moisture-holding capacity at 0.33 bar. The soils were
incubated in the dark at approximately 25 °C. The nonsterile flasks
were sampled after 0.1, 0.2, 1, 3, 7, 14, 30, 60, 120, 270 and 365
days, while samples of sterilized soil were taken after 14, 30, 120,
270 and 365 days. Carbendazim had a half-life of 320 days under
nonsterile aerobic conditions (Marsh & Arthur, 1989). This is in
close agreement with reported half-lives of 6-12 months for
benzimidazoles applied to bare soil (Baude et al., 1974).
After 365 days of incubation, 9% of the 14C was evolved as
14CO2, 34% could still be recovered as carbendazim, and 36% was
not extractable. The total recovery of 14C was 88%. In the
sterilized soil, the half-life of carbendazim was approximately 1000
days (Marsh & Arthur, 1989).
When the degradation of [2-14C]-carbendazim in soil (20
mg/kg) was determined, 33% of added 14C was evolved as 14CO2
during 270 days. Identical or even faster 14C evolution was
observed from 2-14C-labelled 2-AB (Helweg, 1977). The relatively
low 14C evolution from phenyl-14C-labelled benomyl/carbendazim
may be caused by the formation of strongly adsorbed degradation
products or compounds that are readily incorporated in soil organic
matter. Thus, most of the remaining radioactivity was accounted for
in the organic fraction of the soil.
To elucidate the reason for the low 14C evolution from
phenyl-14C-labelled fungicide, the fate of a possible degradation
product, 1,2-diaminobenzene, needs to be determined.
In a study on the effect of different factors on the
degradation of carbendazim in soil, carbendazim was found to remain
in the soil for 120 days. There was 15-29% greater persistence in
sterilized soil than nonsterilized ones. Aspergillus niger tiegh.,
Penicillum chrysogenum Thom., Mucor sp and 2 bacteria ( Bacillus
spp.) alone or in combination degraded the fungicide faster.
Bacteria were most efficient, and there was faster degradation
during the first 20 days. High temperature, acidic pH and higher
moisture level in soil in the presence of microbes all let to faster
degradation of the fungicide. Soil pre-treatment with microbes
resulted in rapid degradation (Gupta & Sharma, 1989).
Soils collected from various fields, which had a history of
carbendazim application, showed increased carbendazim degradation
rates. Low initial doses of carbendazim sufficed to condition soil
with no history of carbendazim application to rapid degradation.
Previous application of the fungicide was not the only means of
inducing the phenomenon. When soil with a history of
carbendazim-treatment was mixed with untreated soil, the ability to
accelerate degradation was observed in the entire soil volume. This
capacity was maintained in soil for over 2 years without
intermediate carbendazim application (Yarden et al., 1987).
Bean plants grown to maturity in Delaware, USA, contained less
than 0.1 mg/kg total 14C residue in the edible beans following two
foliar applications of 1 kg a.i./ha of [2-14C]-carbendazim (as
Delsene 50% WP) at 25% and 50% bloom. Total 14C residues in the
bean foliage decreased from about 5 mg/kg one week after the second
spraying to 0.2 mg/kg three weeks later. The total 14C residue in
edible beans was less than 0.1 mg/kg one week after the second
spraying. Of the total 14C in the edible beans and foliage, 89-95%
was intact free carbendazim and 2-8% was free 2-AB. An additional
1-3% of the 14C was found as ß-glycosidic conjugates of
carbendazim and 2-AB (Han, 1983a).
Chiba & Veres (1981) applied benomyl to apple trees as Benlate
50% WP at a rate of 1.7 kg/ha. Three successive applications were
made in 1977 and a single spray was applied in 1979. Between 3 and 7
days after application there was a marked reduction of about 50% in
benomyl residues from an initial level of about 110 mg/kg. This fall
in benomyl was accompanied by a doubling in the level of carbendazim
residues over the same period due to benomyl degradation to
carbendazim. Within 46 days of the single appli cation in 1979,
benomyl residues fell to 0.63 mg/kg foliage and carbendazim was
present at 1.2 mg/kg. Following the three sprayings in 1977 (at 0,
13 and 27 days after the initial application), residue levels were
2.6 and 17.1 mg/kg foliage for benomyl and carbendazim 83 days after
the first spraying. Both experiments showed an exponential fall in
benomyl residues but the rate of decline was much slower in the case
of the more persistent metabolite.
4.2.2 Abiotic degradation
The hydrolytic stability of carbendazim at pH 5, 7, and 9 and a
nominal temperature of 22, 50 and 70 °C was studied at intervals up
to 30 days. Elevation of temperature and pH increased carbendazim
degradation. The half-life calculated for the degradation of
carbendazim at pH 5 and at 22, 50 and 70 °C was 457, 108, and 29
days, respectively. At pH 7 and at 50 and 70 °C the half-life was 43
and 12 days (no appreciable decline at 22 °C). The half-life at pH 9
and at 22, 50, and 70 °C was 22, 1.4 and 0.3 days, respectively
(Purser, 1987).
Carbendazim was exposed to sunlight for 30 h (as a residue on
silica gel G) and less then 10% was lost after exposure.
Photo-oxidation of the benzene ring of carbendazim was the
predominant reaction with some guanine, carbomethoxyguanine, and
carbomethoxyurea detected. When carbendazim was applied to the
leaves of corn plants and exposed to sunlight for 18 h, no
photolysis products were detected in the extracts of plants (Fleeker
& Lacy, 1977).
4.2.3 Bioaccumulation
Bluegill sunfish ( Lepomis macrochirus) were exposed to
radiolabelled carbendazim concentrations of 0.018 or 0.17 mg/litre
for 4 weeks in a dynamic study designed to measure the
bioaccumulation of 14C residues in edible tissue, viscera,
remaining carcass and whole fish. A two-week depuration phase
followed the exposure phase. Results were similar at the two
exposure concentrations, the peak whole fish bioconcentration
factors (BCFs) being 27 and 23 at the low and high exposure levels,
respectively. The radioactivity was concentrated more in the viscera
than in other tissues, the peak viscera BCFs being 460 and 380 for
the low and high exposure levels, respectively. Very little
bioconcentration occurred in the muscle tissue (BCF = < 4) or the
remaining carcass (BCF = < 7). During the 14-day depuration phase,
> 94% of the peak level of radioactivity was lost from the whole
fish, viscera and muscle. The rate of loss from the carcass tissue
was lower (77% and 82% loss for the low and high exposure levels,
respectively) (Hutton et al., 1984).
When rainbow trout ( Oncorhynchus mykiss), channel catfish
( Ictalurus punctatus) and bluegill sunfish ( Lepomis macrochirus)
were injected intraperitoneally with carbendazim, branchial and
biliary excretion were the major pathways for the elimination
(Palawski & Knowles, 1986). In a separate experiment, the three fish
species were exposed to 45 µg carbendazim/litre for 96 h, except in
the case of catfish, which were exposed for 48 h. This was followed
by a 96-h depuration phase. Rainbow trout had the highest uptake
rate constant (1.78 per h) and bioconcentration factor (159) of the
three species. Much less carbendazim was accumulated by channel
catfish than by the other two species, but this residue level (0.44
µg/g) appeared to be lethal after 48 h of exposure. The elimination
rate constant and the biological half-life of carbendazim were
similar for rainbow trout and bluegill sunfish. However, the
elimination rate constant was greater and the biological half-life
shorter in channel catfish (13 h) than in the other two species.
5. ENVIRONMENTAL LEVELS AND HUMAN EXPOSURE
5.1 Environmental levels
5.1.1 Air, water and soil
The environmental levels in air, water and soil are discussed
in detail in chapter 4.
5.1.2 Food and feed
Levels of carbendazim in food and feed are indicated in section
5.2.
5.1.3 Terrestrial and aquatic organisms
Carbendazim levels in terrestrial and aquatic organisms are
discussed in detail in chapters 4 and 6.
5.2 General population exposure
The principal exposure of the general population to carbendazim
is through dietary exposure. It was recommended by the Joint FAO/WHO
Meeting on Pesticide Residues (JMPR) (FAO/WHO, 1985a) that all
maximum residue limits (MRLs) for benomyl, thiophanate-methyl and
carbendazim be listed as carbendazim (Tables 3 and 4).
5.2.1 Sweden
Monitoring data from Sweden are shown in Table 5 (FAO/WHO,
1988b). No further analysis to determine dietary intake was
performed.
5.2.2 The Netherlands
Over a period of two years (June 1976 to July 1978), "market
basket" samples for 16- to 18-year-old males, including 126
different food items, were purchased every two months. This age
group was chosen by the authors under the assumption that they had
the greatest food consumption. The food was prepared for eating
(including cooking) and combined in 12 different commodity groups,
and the concentrations of 78 different chemical pesticides were
determined. Using concentrations found in the total diet samples,
the daily intakes were calculated. For this sub-population, the
maximum intake of carbendazim was 0.6 mg/day (calculated using the
detection limit as the concentration of the non-detectable residues)
and the average dietary intake was 0.05 mg/day. The recommended ADI
is 0-0.01 mg/kg body weight (FAO/WHO, 1985a), corresponding to 0.6
mg/day for a 60-kg adult. Therefore, the maximum intake is at the
recommended ADI, whereas the average intake is 12 times below the
recommended ADI (de Vos et al., 1984).
Table 3. Benomyl/carbendazim/thiophanate-methyl residues in food in Swedena
Samples Swedish/imported No. of samples Samples with residues Residue level Median value
>0.20 mg/kg (mg/kg) (mg/kg)
1986
Pineapples imported 3 1 0.69
Grapes imported 20 3 0.17-0.35 0.26
Strawberries imported 7 1 0.29
Mangoes imported 17 4 0.20-1.82 0.70
Papayas imported 5 2 0.25-0.45
Pears Swedish 17 3 0.32-0.62 0.43
imported 45 7 0.20-0.45 0.34
Apples Swedish 78 17 0.20-0.72 0.40
imported 91 30 0.21-0.74 0.39
1987
Grapes imported 28 3 0.52-0.87 0.60
Strawberries imported 7 0.23
Mangoes imported 14 5 0.29-1.30 0.66
Papayas imported 4 2 0.86-1.14
Table 3 (contd).
Samples Swedish/imported No. of samples Samples with residues Residue level Median value
>0.20 mg/kg (mg/kg) (mg/kg)
Pears Swedish 14 1 0.52
imported 62 13 0.21-0.45 0.29
Apples Swedish 61 25 0.20-1.17 0.45
imported 94 12 0.21-0.82 0.36
a From: FAO/WHO (1988b)
Table 4. National Maximum Residue Limits (mg/kg) for certain commoditiesa
banana cereal cherries citrus bean cucumber peach pome fruit strawberries grapes
Australia 1 0.05 5 10 3 3 5 5 6 2
Austria 0.2 0.5 7 1 0.5 2 1.5 3
Belgium 2 0.5 2 2 0.5 2 5 5 2
Brazil 1 0.5 10 10 2 0.5 10 5 5 10
Bulgaria 0.5 10 5 5 10
Canada 5 10 1 0.5 10 5 5 5
Denmark 2 0.1 2 5 2 2 2 2 5 5
France 1 1.5 6
Finland 0.2 1 2 0.5 0.5 1 1 1
Germany 0.2 0.5 2 7 1 0.5 2 2 3
Hungary 2 1
Israel 10 10 10 5 10
Italy 0.5 0.5 1 1
Mexico 10 2 11 5 7 5 10
Netherlands 3 0.1 3 4 3 3 3 3 3 3
New Zealand 5 1 5 5 2 2 5 5 5 5
Spain (guidelines) 1 0.5 5 7 2 2 5 5 1 5
Table 4 (contd).
banana cereal cherries citrus bean cucumber peach pome fruit strawberries grapes
Switzerland 1 0.2 3 7 0.2 0.1 3 3 3 3
United Kingdom
(proposed) 1 0.5 10 0.5 10 5 5 10
USA 1 0.2 15 10 2 1 15 7 5 10
USSR 1 0.5 10 10 2 0.5 10 5 5 5
Yugoslavia 0.1 7 0.5 0.1 2 0.5 2
a From: FAO/WHO (1988a)
Table 5. Proposed Maximum Residue Limits for carbendazim from any
sourcea
Commodity MRL (mg/kg) Applicationb
Apricot 10c B,C
Asparagus 0.1d B,T
Avocado 0.5 B
Banana 1c B,C,T
Barley straw and fodder, dry 2 B
Bean fodder 50 C
Beans, dry 2 B
Berries and other small fruit 5 B,C,T
Brussel sprouts 0.5 B
Broad bean 2 T
Carrot 5c C,T
Cattle meat 0.1d B
Celery 2 B,C
Cereal grains 0.5 B,C,T
Cherries 10c B,C,T
Citrus fruits 10c B,C,T
Coffee beans 0.1d C
Common beane 2 C
Cucumber 0.5 B,C,T
Eggs (poultry) 0.1d B,T
Egg plant 0.5 C
Gherkin 2 C,T
Hops, dry 50 C
Lettuce, head 5 B,C,T
Mango 2 B
Melons, except watermelons 2c B,C
Milk 0.1d B
Mushrooms 1 B,C,T
Nectarine 2 B
Onion, bulb 2 C,T
Peach 10c B,C,T
Peanut 0.1d B,C
Peanut fodder 5 B,C
Peppers 5 C
Pineapple 20c B
Plums (including prunes) 2c B,C,T
Pome fruit 5c B,C,T
Potato 3c,f B,C
Poultry meat 0.1d B,T
Rape seed 0.05d C
Rice straw and fodder, dry 15 B,C,T
Sheep meat 0.1d B
Soya bean, dry 0.2 C
Soya bean fodder 0.1d C
Table 5 (contd).
Commodity MRL (mg/kg) Applicationb
Squash, summer 0.5 B
Sugar beet 0.1d B,C,T
Sugar beet leaves on tops 10 B,C,T
Swedeg 0.1d C
Sweet potato 1 B
Taro 0.1d B
Tomato 5 B,C,T
Tree nuts 0.1d B
Wheat straw and fodder, dry 5 B
Winter squash 0.5 B
a From: FAO/WHO (1988b)
b B = benomyl; C = carbendazim; T = thiophanate-methyl
c MRL based on post-harvest use
d At or about the limit of detection
e JMPR recommended 2 mg/kg for dry, dwarf, lima and snap beans.
These are all covered by "VP 0526, Common bean" and "VP 0071,
Beans, dry" in the new classification
f washed before analysis
g Described as rutabagas in 1983 recommendation
5.2.3 National maximum residue limits
National MRLs for certain commodities are listed in Table 4
(FAO/WHO, 1988a).
A complete list of MRLs for carbendazim, including new
proposals and an indication of the source of the data (application
of benomyl, carbendazim, or thiophanate-methyl) on which the MRL is
based, is given in Table 5 (FAO/WHO, 1988b).
5.3 Occupational exposure during manufacture, formulation, or use
5.3.1 Exposure during manufacture
Levels of carbendazim, monitored as worker inhalation exposure,
in a major manufacturing facility (Du Pont) were reviewed from 1986
to 1989. The average level of carbendazim was less than 0.3 mg/m3.
Table 6 lists established inhalation exposure limits for benomyl and
carbendazim.
� 5.3.2 Exposure during use
Although no studies have been performed with carbendazim,
potential dermal and respiratory exposure to benomyl wettable powder
formulation in actual-use situations was determined for tank loading
and mixing for aerial application, re-entry into treated crops, and
home use (garden, ornamental and greenhouse). For crop treatments,
approximately 17 kg benomyl (formulation) was handled per cycle.
Maximum exposure occurred in the loading and mixing operation for
aerial application, where dermal exposure was 26 mg benomyl per
mixing cycle, primarily to hands and forearms (90%), and respiratory
exposure averaged 0.08 mg benomyl. Re-entry data revealed dermal and
respiratory exposures of 5.9 mg/h and < 0.002 mg/h, respectively.
Home-use situations (application of 7 to 8 litres benomyl in
hand-held compressed air sprayers) produced exposures of 1 mg and
0.003 mg per application cycle for the dermal and respiratory
routes, respectively (Everhart & Holt 1982).
Table 6. Established inhalation exposure limitsa
Country and agency Compound TWAb STELc
(mg/m3) (mg/m3)
Australia benomyl 10 -
Belgium benomyl 10 -
Denmark benomyl 5 -
Finland benomyl 10 30
France benomyl 10 -
Switzerland benomyl 10 -
United Kingdom benomyl 10 15
USA: ACGIHd benomyl 10 -
USA: NIOSHe/OSHAf benomyl 10 -
(inhalable dust)
USA: NIOSH/OSHA benomyl 5 -
(respirable dust)
USSR carbendazim - 0.1
a From: ILO (1991)
b Time-weighted average
c Short-term exposure limit
d American Conference of Governmental Industrial Hygienists
e National Institute of Occupational Safety and Health
f Occupational Safety and Health Administration
6. KINETICS AND METABOLISM
Carbendazim is extensively metabolized by animals as described
in detail in section 6.3. Metabolite names and structures are given
in Table 7, Fig. 1 and Fig. 2.
6.1 Absorption
Male albino rats were orally administered a single
14C-carbendazim dose of 12 mg/kg as a solution in diethyl
glycol-ethanol. Based on urinary excretion of 14C-carbendazim and
its metabolites, 5-HBC and 2-AB, the absorption was determined to be
about 85% (Krechniak & Klosowska, 1986).
In a study by Monson (1990), young male and female
Sprague-Dawley rats were administered a single dose via gavage with
14C-radiolabelled [phenyl(U)-14C]-carbendazim (94% pure,
suspended in corn oil) at either a low (50 mg/kg) or a high (1000
mg/kg) dose, and excretion of the radiolabel was monitored every 12
h for 72 h. Most of the radioactivity was excreted by 72 h. The
percentages of originally administered radioactivity (i.e.
14C-carbendazim equivalent) recovered were: a) in urine, 61.7
(male), 53.8 (female) for low dose, 41.4 (male) and 40.7 (female)
for high dose; and b) in faeces, 24.4 (male), 33.2 (female) for low
dose and 61.9 (male) and 69.5 (female) for high dose. The author
concluded that: a) > 98% of the administered dose was recovered in
urine and faeces in all test groups; and b) the absorption
efficiency of carbendazim was approximately 80% of the actual
administered dose for all dose levels, based on the level of
radioactivity appearing in urine and the sum of all metabolites
appearing in faeces as the result of hepatic metabolism (Monson,
1990).
6.2 Distribution and accumulation
In a rat gavage study, [phenyl(U)-14C]-carbendazim was
administered to Sprague-Dawley rats (five rats per sex per group)
using three dosing regimes: a single oral dose of 50 mg/kg; a single
oral dose of 50 mg/kg following pre-conditioning gavage of
non-labelled carbendazim (50 mg/kg) for 14 days; and a single oral
dose of 1000 mg/kg. All rats were sacrificed 72 h after the last
dose. Tissue distribution data showed lack of bioconcentration of
radiolabelled compound. The highest concentrations of radio labelled
tissue residues (less than 1% of the dose) were detected in the
residual carcass and liver (Monson, 1990). This study is discussed
in detail in sections 6.3 and 6.4.
Table 7. Carbendazim and its metabolites in animalsa
Code name Chemical name
Carbendazim (MBC) methyl (1-H-benzimidazol-2-yl)carbamate
5-HBC methyl (5-hydroxy-1H-benzimidazol-2-yl)
carbamate
4-HBC methyl (4-hydroxy-1H-benzimidazol-2-yl)
carbamate
5-HBC-Sb 2-[(methoxycarbonyl)amino]-1H-benzimidazol-5-
yl hydrogen sulfate
5-HBC-Gc 2-[(methoxycarbonyl)amino]-1H-benzimidazol-
5-yl] ß-D-glucopyranosiduronic acid
MBC-4,5-epoxide
MBC-5,6-epoxide
MBC-4,5-dihydrodiol (4,5-dihydro-4,5-dihydroxy-1H-benzimidazol-
2-yl) carbamate
MBC-5,6-dihydrodiol (5,6-dihydro-5,6-dihydroxy-1H-benzimidazol-
2-yl) carbamate
MBC-4,5-diol
MBC-5,6-diol
5-OH-6-GS-MBCd S-[5,6-dihydro-5-hydroxy-2-(methoxycarbonyl
amino)-1H-benzimidazol-4-yl]glutathione
5-OH-4-GS-MBC S-[4,5-dihydro-5-hydroxy-2-(methoxycarbonyl
amino)-1H-benzimidazol-4-yl]glutathione
5,6-HOBC-N-oxide methyl (6-hydroxy-5-oxo-5H-benzimidazol-2-
yl)-carbamate-N-oxide
5,6-HOBC-N-oxide-G 2-[(methoxycarbonyl)amino]-6-oxo-6H-
benzimidazol-5-yl] ß-D-glucopyranosiduronic
acid-N-oxide
5,6-DHBC methyl (5,6-dihydroxy-1H-benzimidazol-2-yl)
carbamate
Table 7 (contd).
Code name Chemical name
5,6-DHBC-G 6-hydroxy-2-[(methoxycarbonyl)amino]-1H-
benzimidazol-5-yl] ß-D-glucopyranosiduronic
acid
5,6-DHBC-S 6-hydroxy-2-[(methoxycarbonyl)amino]-1H-
benzimidazol-5-yl 5-(hydrogen sulfate)
2-AB 2-aminobenzimidazole
2-AB dihydrodiol 2-amino-4,5-dihydro-4,5-dihydroxy-1H-
benzimidazol
5-HAB 5-hydroxy-2-aminobenzimidazole
a From: Krechniak & Klosowska (1986); Monson (1986, 1990)
b S = conjugate with sulfuric acid
c G = conjugate with glucuronic acid
d GS = conjugate with glutathione
In a study by Krechniak & Klosowska (1986), male albino rats
were administered by gavage a single dose of 12 mg carbendazim per
kg and the distribution of carbendazim and its metabolites among
subcellular liver fractions was determined 1.5 h after dosing. The
distribution was not uniform and was not dependent on the lipid
content of the fractions. The highest relative concentration of
unchanged carbendazim was in the mitochondria. The highest relative
concentration of 5-HBC was in the cytosol and that of 2-AB was in
the microsomes.
Ten hens were individually dosed for six consecutive days with
0.625 mg [2-14C]-carbendazim at a rate equivalent to 5 mg/kg in
the average total daily feed. An additional 10 hens were dosed daily
with 12.5 mg at a rate equivalent to 120 mg/kg in the average total
daily feed. The hens were sacrificed 24 h after the sixth dose and
muscle, kidney, liver and fat were sampled. Carbendazim was
extensively metabolized to 5-HBC and methyl (4,5-dihydro-4,5-
dihydroxy-1H-benzimidazol-2-yl) carbamate. The concentration of
radioactivity, calculated as mg carbendazim per kg, in the high-dose
hens was 2.63 (liver), 1.74 (kidney), 0.06 (thigh muscle), 0.05
(breast muscle), 0.03 (fat) and 0.63 (day-6 eggs). Approximately 73%
of the total radioactivity in the day-6 eggs was identified as 5-HBC
(0.26 mg/kg) and unchanged carbendazim (0.15 mg/kg) (Monson, 1986).
In another feeding study, groups of 20 hens were fed
carbendazim daily at levels of 0, 5, 15 and 100 mg/kg diet for 28
days. Eggs were collected daily and faeces once a week. After 28
days, 15 hens in each group were sacrificed and samples taken for
analysis. The remaining hens were fed a carbendazim-free diet for a
further week and then sacrificed. The majority of the material fed
was rapidly eliminated as parent compound or as 5-HBC. The only
residues found in any of the samples of blood, fat, liver, kidney,
tissue or eggs were in the eggs from the 100-mg/kg group. However,
the residues (up to 0.1 mg carbendazim/kg and 0.36 mg 5-HBC/kg each)
decreased to < 0.05 mg/kg within 4 days of withdrawal of treatment
(Eckert et al., 1985).
A lactating Holstein cow was dosed by capsule twice daily (483
mg [2-14C]-carbendazim each dose), equivalent to 50 mg/kg in the
average total daily feed, for five consecutive days. Samples of milk
were collected at each dosing. Approximately 17 h after the tenth
dose, the cow was sacrificed. Small amounts of radioactivity
corresponding to carbendazim or its metabolites were found in the
liver (2.62 mg/kg) and kidney (0.45 mg/kg), but no significant
amounts (< 0.09 mg/kg) were detected in other tissues or fat. Only
2.7% of the liver radioactivity was 5-HBC whereas 41% and 3% of the
kidney radioactivity corresponded to 5-HBC and 4-HBC, respectively.
14C residue levels in the milk averaged 0.25 mg/kg (calculated as
carbendazim) of which 0.11 mg/kg was 5-HBC and 0.05 mg/kg 4-HBC. No
carbendazim (< 0.01 mg/kg) was detected in the milk (Monson, 1985).
Twelve non-lactating female goats were administered a
feed-rate-equivalent [phenyl(U)-14C]-carbendazim dose of at least
50 mg/kg (range: 50-101 mg/kg) once a day for up to 30 days. A
plateau of 14C residues in the liver was achieved within 2 weeks
of dose initiation and was calculated to be 9.48 mg/kg of liver
(group mean of the total radiolabelled liver residues for goats
sacrificed 2, 3 and 4 weeks after initiation of dosing). The total
14C residue levels in the liver decreased to 5.17, 3.55 and 1.67
mg/kg at 1, 2 and 3 weeks, respectively, after discontinuing dosing.
Based on this data, the elimination half-life for the total 14C
residues from the liver was calculated to be approximately 9 days.
The half-life for removal of carbendazim from the general
circulation, based on 14C-carbendazim equivalent whole blood
levels, was approximately 10 h. The results of this study suggest
that levels of carbendazim-derived residues do not continue to
accumulate beyond 2 weeks when goats are exposed to a constant
feed-level of 50 mg carbendazim/kg. Furthermore, discontinuation of
exposure results in a clearing of residues from the liver (Johnson,
1988).
6.3 Metabolic transformation
In a rat gavage study, carbendazim was found to be extensively
metabolized. Three dosing regimes (five rats of each sex per group)
were used: a single oral dose of 50 mg/kg (low dose); a single oral
dose of 50 mg/kg following pre-conditioning gavage with
non-radiolabelled carbendazim of 50 mg/kg for 14 days
(pre-conditioned low dose); and a single oral dose of 1000 mg/kg
(high dose). The 48-h urine from the low-dose and the high-dose
rats, and the 14-day urine from the pre-conditioned low-dose group
were collected. The total recovery from urine was 61.5 and 61.7% of
given doses for the low-dose and pre-conditioned low-dose male
groups, 53.2 and 59.3% for the low-dose and pre-conditioned low-dose
female groups, and 39 and 41% for both male and female high-dose
groups, respectively. 5-HBC-S (21-43% of given dose) was identified
as the main metabolite, except in the case of the pre-conditioned
low-dose and the high-dose female rat groups (5.5-10%), while in all
female rat groups 5,6-HOBC-N-oxide-G (10-19%) was predominant. Both
5,6-DHBC-S and 5,6-DHBC-G were identified as minor metabolites.
In the same study, the faeces were collected at the same
periods as the urine. The total recovery from faeces was about 24%
for the low-dose and pre-conditioned low-dose male groups, 33-38%
for the low-dose and pre-conditioned low-dose female groups, and
higher (> 60%) for both male and female high-dose groups. Unchanged
carbendazim was about 10-15% of the given dose in the faeces of
high-dose rats (Monson, 1990). The proposed metabolic pathway for
carbendazim in rats is given in Fig. 1.
NMRI mice and Wistar rats of both sexes were given carbendazim,
via gavage, as a single dose of 3 and 300 mg/kg, respectively. Urine
was collected during the first 6 h, after which the animals were
killed. Almost all the metabolites in urine were conjugated with
sulfuric acid. Cleavage of these conjugates by ß-glucuronidase/
arylsulfatase released 5-HBC as the only metabolite extractable from
water. Mouse urine contained a greater amount of compounds that
remained polar after enzyme treatment than the corresponding urine
of rats. Analyses revealed no sex differences (Dorn et al., 1983).
In a further study, male albino rats were administered a single
intravenous dose of 12 mg carbendazim/kg as a solution in
diethylene-glycol. The composition of the measured radioactivity in
urine 12 h after dosing was 94% as 5-HBC, 3% as 2-AB, and 3% as
carbendazim (Krechniak & Klosowska, 1986).
In hens dosed with [2-14C]-carbendazim (5 and 120 mg/kg in
the daily feed), carbendazim was metabolized to 5-HBC, 4-HBC,
4,5-dihydrodiol-MBC and its sulfuric acid conjugate, and also to
2-AB (Monson, 1986). The proposed metabolic pathway in laying hens
is given in Fig. 2.
Empirical formula: C9H9N3O2
Relative molecular mass: 191.2
CAS chemical name: Methyl (1H-benzimidazol-2-yl)carbamate
IUPAC chemical name: Methyl benzimidazole-2-ylcarbamate
CAS Registry number: 10605-21-7
Synonyms: carbendazol (ZMAF), methyl-2-benzimidazole
carbamate (MBC, MCB, BCM, BMC)
2.1.2 Technical product
Major trade names:
Carbendazim, Delsene, Bavistin, Corbel, Konker, Bendazim,
Derosal, Kombat, Kemdazin, Carbendor, Hoe 017411, Cekudazim,
Equitdazin, Aimcozim (Some of these are formulations with other
pesticides.)
Purity: > 98% (FAO specifications)
Impurities: 2,3-diaminophenazine (DAP), 2-amino-3-
hydroxyphenazine (HAP)
2.2 Physical and chemical properties
Table 1. Some physical and chemical properties of carbendazim
Physical state Crystalline solid
Colour White
Odour Negligible
Table 1 (contd).
Melting point/boiling
point/flash point Melts at -250 °C
Explosion limits LEL = 0.13 g/litre in air
Vapour pressure < 1 x 10-7 Pa (< 1 x 10-9 mbar) at 20 °C
Density 0.27 g/cm3 (loose); 0.62 g/cm3 (packed)
Log n-octanol/water 1.49
partition coefficient
Solubility in water pH 4 28 mg/litre
(at 20 °C) pH 7 8 mg/litre
pH 8 7 mg/litre
Solubility in organic solvents
Hexane 0.5 mg/litre
Benzene 36 mg/litre
Dichlorom ethane 68 mg/litre
Ethanol 300 mg/litre
Dimethylformamide 5000 mg/litre
Acetone 300 mg/litre
Chloroform 100 mg/litre
Henry's constant 1.02 x 10-9 atm-m3/mol at 20 °C
2.3 Analytical methods
Methods for determining carbendazim and its by-product residues
in plant and animal tissue and in soil involve isolation of the
residue by extraction with an organic solvent and purification of
the extract by a liquid-liquid partitioning procedure. Measurement
of the residues may be determined by procedures using high-speed
cation exchange liquid chromatography, reversed phase HPLC, and
immunoassay. Recoveries of carbendazim and 2-aminobenzimidazole
(2-AB) from various types of soils average 88 and 71%, respectively.
The lower limit of sensitivity of the method is 0.05 ppm for each of
these components. The recoveries and sensitivities in the case of
plant tissues are similar. Table 2 outlines various methods for
soil, water, plants and animal tissue.
Table 2. Analytical methods for carbendazim
Analytical method Medium Detection limit Comments Reference
Strong cation exchange/HPLC soil 0.05 mg/kg acidic methanol extraction converts Kirkland et al. (1973)
residual benomyl to carbendazim
Strong cation exchange/HPLC plant 0.05 mg/kg acidic methanol extraction converts Kirkland et al. (1973)
residual benomyl to carbendazim
Strong cation exchange/HPLC animal 0.01 mg/kg (milk) acidic aqueous hydrolysis followed Kirkland (1973)
0.05 mg/kg by organic extraction converts
(tissue) benomyl to carbendazim and frees
metabolites from conjugates
Reversed phase HPLC water 9.0 x 10-6 g/litre on-line HPLC with preconcentration; Marvin et al. (1991)
benomyl and carbendazim
determined separately
Reversed phase blueberries 0.03 mg/kg acidic methanol extraction converts Bushway et al. (1991)
HPLC/fluorescence detection residual benomyl to carbendazim
Radioimmunoassay plant 0.05-1.0 mg/kg ethyl acetate extraction converts Newsome & Shields
(dependent on crop) residual benomyl to carbendazim (1981)
Enzyme-linked immunosorbent plant 0.50 mg/kg ethyl acetate extraction converts Newsome & Collins
assay (ELISA) residual benomyl to carbendazim (1987)
3. SOURCES OF HUMAN AND ENVIRONMENTAL EXPOSURE
3.1 Natural occurrence
Carbendazim does not occur naturally.
3.2 Anthropogenic sources
3.2.1 Uses
Carbendazim is a fungicide in its own right as well being as
the main metabolite of other fungicides such as benomyl and
thiophanate-methyl.
Carbendazim is used to control a wide range of fungi, including
Ascomycetes, Fungi Imperfecti and numerous Basidiomycetes, which
result in plant diseases such as: leaf spots, blotches and blights;
fruit spots and rots; sooty molds; scabs; bulb, corn and tuber
decays; blossom blights; powdery mildews; certain rusts; and common
soilborne crown and root rots. It is used on cereals, cotton,
grapes, bananas and other fruit, ornamentals, plantation crops,
sugar beet, soybeans, tobacco, turf, vegetables, mushrooms, and many
other crops under most climatic conditions worldwide. Registered
carbendazim usage specifies rates from 0.2 to 2.0 kg a.i./ha and
applications from once per year to spray intervals ranging from 7 to
14 days (FAO/WHO, 1985b; 1988b).
A key limitation to the use of carbendazim and other
benzimidazoles is the development of fungal resistance. Resistance
management can be achieved by using carbendazim in combination as a
tank mix or alternately with other non-benzimidazole fungicides
(Delp, 1980; Staub & Sozzi, 1984).
Carbendazim is formulated as an aqueous dispersion, aqueous
suspension, flowable water dispersible granules and a wettable
powder.
In 1991, the estimated worldwide sales of benomyl was US$ 290
million. This was about 50% of the worldwide market for
benzimidazole products. Carbendazim (20%) and thiophanatemethyl
(20%) account for most of the rest of the benzimidazole market
(County NatWest Woodmac, 1992).
4. ENVIRONMENTAL TRANSPORT, DISTRIBUTION AND TRANSFORMATION
4.1 Transport and distribution between media
4.1.1 Air
Carbendazim has a vapour pressure of < 1 x 10-7 Pa (< 1 x
10-9 mbar) at 20 °C, an aqueous solubility of 8 mg/litre at 20 °C
and pH 7, and a Henry's constant of approx. 1.02 x 10-9
atm-m3/mol at 20 °C. It is essentially non-volatile from water
surfaces.
4.1.2 Water
Anaerobic aquatic degradation studies of [phenyl(U)-14C]-
benomyl in pond water and sediment showed that more than 98% of the
carbendazim residues partitioned into the sediment after 7 days. The
half-life of carbendazim was 743 days. After one year 36% of the
applied radioactivity was bound to the sediment (Arthur et al.,
1989a).
An aerobic aquatic degradation study of [phenyl(U)-14C]-
benomyl in pond water and sediment showed that carbendazim had a
half-life of 61 days under nonsterile conditions. After 30 days, 22%
of the applied radioactivity was bound to the sediments and < 1% of
the applied radioactivity was evolved as carbon dioxide (Arthur et
al., 1989b).
4.1.3 Soil
In greenhouse studies to determine run-off and leaching of
[2-14C]-carbendazim on soil, a container of Keyport silt loam was
treated with labelled carbendazim, at a rate of 11 kg a.i./ha, by
spraying the upper one-third (0.093 m2) of the plot and was
allowed to stand for 24 h. Artificial rain was then applied (3.75 cm
the first day after treatment and 2.5 cm on the third and seventh
days). All water that ran off or leached through the soil was
collected and analysed for total 14C. Soil in the plot was divided
into layers for analysis, air dried and analysed separately for
total 14C. After each of the three rain applications, 0.05-0.39%
of the applied 14C was found in run-off water; < 0.01% was found
in the leach water after the first two artificial rains and 0.19%
after the third one. Soil analyses showed that 90.6% of the applied
activity remained in the treated area and 93.1% in the top 10 cm of
soil (Rhodes & Long, 1983).
4.1.4 Leaching
To evaluate the risk of pollution of ground and drainage water,
screening models based on adsorption and persistence can be used,
together with existing analyses of groundwater samples. Gustafson
(1989) proposed the use of the equation GUS = log T´ (4 - log
Koc); GUS values < 1.8 = "improbable leachers", GUS values of
1.8-2.8 = "transition" and GUS values > 2.8 = "probable leachers".
For benomyl, Kom values of 550, 620, 2100 and 1100 (mean 1093)
were found in four different soils (Priester, 1985). A Kom of 1093
is equal to a Koc of 1857 since Koc = Kom x 1.7. The half-life
of 320 days given by Marsh & Arthur (1989) seems in good agreement
with field half-lives of 6 to 12 months (Baude et al., 1974).
When the calculation of the GUS value is based on a Koc of
1857 and a T0.5 of 320 days, a value of 1.83 is obtained.
According to this value, benomyl/carbendazim lies between the
"improbable leachers" and "transition", and, therefore, would not be
expected to occur in ground water. The adsorption of benomyl and of
carbendazim is expected to be of the same order of magnitude since
the Kow values are almost identical (log Kow = 1.49 and 1.36 for
carbendazim and benomyl, respectively). In groundwater studies in
the USA (Parsons & Witt, 1988), benomyl was not found in any of 495
wells tested and carbendazim not in any of 212 wells (detection
limit not reported).
In an EEC survey (Fielding, 1992), the presence of carbendazim
in groundwater in the Netherlands and in Italy was investigated.
Carbendazim was found in one of two samples from the Netherlands
(0.1 µg/litre), and the level was above 0.1 µg/litre in 23 of 70
samples in Italy. Detection of the non-polar DDT and lindane in many
wells in the Italian study may indicate macropore transport or
artifacts such as direct pollution of wells.
4.1.5 Crop uptake
Various greenhouse and outdoor tests with carbendazim indicate
that it remains on plant surfaces as the major component of the
total residue (Baude et al., 1973).
A greenhouse crop-rotation study was undertaken by application
of [2-14C]carbendazim to a loamy sand soil, followed by aging
periods of 30, 120 or 145 days. The crops studied were beets, barley
and cabbage. Radioactivity did not accumulate in these crops grown
to maturity in a loamy sand soil treated 30 days earlier (1.1 kg
a.i./ha) or 120 to 145 days earlier (3.4 kg a.i./ha). Accumulation
factors, calculated as the ratio of radioactivity in the crop to
that in the corresponding soil, were very low in beet foliage (0.04)
and beet roots (0.03), low in cabbage and barley grain (0.2) and
ranged from 0.9 to 1.2 in barley straw (Rhodes, 1987).
Alfalfa, soybean and ryegrass, which were grown in 0.028 m3
(1 cu ft) containers in a greenhouse in soil treated with an 80:20
mixture of carbendazim and 2-aminobenzimidazole (2-AB), contained
small but detectable residues of both compounds. Both 14C-labelled
and non-labelled mixtures were applied at the rate of 2.2 kg/ha
uniformly incorporated in the 0-10 cm layer of soil. In the 14C
studies, alfalfa contained total 14C residues equivalent to
0.13-0.30 mg/kg of carbendazim/2-AB. Soybean plants contained
0.32-0.53 mg/kg and ryegrass (20-183 days after planting) contained
0.09-0.19 mg/kg. Each plant contained approximately equal amounts of
carbendazim, 2-AB and a polar unknown fraction. Alfalfa from the
non-labelled series contained 0.05 and 0.08 mg/kg, respectively, of
carbendazim and 2-AB at the first cutting and < 0.05 mg/kg of each
compound at the second and third cuttings. Soybean plants contained
< 0.1 mg 2-AB/kg and 0.59 mg carbendazim/kg. Ryegrass from six
cuttings (20-149 days after planting) contained 0.08-0.48 mg
carbendazim/kg and < 0.05 mg 2-AB/kg. All data were calculated on a
fresh weight basis (Rhodes et al., 1983).
4.2 Transformation
4.2.1 Soil biodegradation
Aerobic degradation studies of labelled [2-14C]-2-AB, the
primary degradation product of carbendazim in soil, showed that
14C evolution increased exponentially from 1 to 22 °C, reached a
maximum at 22 °C, remained almost constant up to 35 °C, then became
almost zero at 40 °C, when the soil water content was 100% of field
capacity. At 25 °C, 14C evolution increased exponentially with an
increase in the field capacity of water from 28 to 94%. These and
other results indicate the presence of organisms that are able to
decompose 2-AB (Helweg, 1979).
Laboratory studies on two types of soil under anaerobic
conditions using [2-14C]-carbendazim showed only a small amount of
2-AB (< 0.1%) and no other degradation products (< 0.05%).
Re-incorporation of 14C into soil humus was indicated by
fractionation studies, which showed that the unextracted 14C
residue was widely distributed in various organic soil components
(Han, 1983b).
The persistence of carbendazim was monitored in nonsterile and
sterile Keyport silt loam soil after it was treated with
[phenyl(U)-14C]-benomyl at a concentration of approximately 7.0
mg/kg. Distilled water was added to each sample until it reached 75%
of its moisture-holding capacity at 0.33 bar. The soils were
incubated in the dark at approximately 25 °C. The nonsterile flasks
were sampled after 0.1, 0.2, 1, 3, 7, 14, 30, 60, 120, 270 and 365
days, while samples of sterilized soil were taken after 14, 30, 120,
270 and 365 days. Carbendazim had a half-life of 320 days under
nonsterile aerobic conditions (Marsh & Arthur, 1989). This is in
close agreement with reported half-lives of 6-12 months for
benzimidazoles applied to bare soil (Baude et al., 1974).
After 365 days of incubation, 9% of the 14C was evolved as
14CO2, 34% could still be recovered as carbendazim, and 36% was
not extractable. The total recovery of 14C was 88%. In the
sterilized soil, the half-life of carbendazim was approximately 1000
days (Marsh & Arthur, 1989).
When the degradation of [2-14C]-carbendazim in soil (20
mg/kg) was determined, 33% of added 14C was evolved as 14CO2
during 270 days. Identical or even faster 14C evolution was
observed from 2-14C-labelled 2-AB (Helweg, 1977). The relatively
low 14C evolution from phenyl-14C-labelled benomyl/carbendazim
may be caused by the formation of strongly adsorbed degradation
products or compounds that are readily incorporated in soil organic
matter. Thus, most of the remaining radioactivity was accounted for
in the organic fraction of the soil.
To elucidate the reason for the low 14C evolution from
phenyl-14C-labelled fungicide, the fate of a possible degradation
product, 1,2-diaminobenzene, needs to be determined.
In a study on the effect of different factors on the
degradation of carbendazim in soil, carbendazim was found to remain
in the soil for 120 days. There was 15-29% greater persistence in
sterilized soil than nonsterilized ones. Aspergillus niger tiegh.,
Penicillum chrysogenum Thom., Mucor sp and 2 bacteria ( Bacillus
spp.) alone or in combination degraded the fungicide faster.
Bacteria were most efficient, and there was faster degradation
during the first 20 days. High temperature, acidic pH and higher
moisture level in soil in the presence of microbes all let to faster
degradation of the fungicide. Soil pre-treatment with microbes
resulted in rapid degradation (Gupta & Sharma, 1989).
Soils collected from various fields, which had a history of
carbendazim application, showed increased carbendazim degradation
rates. Low initial doses of carbendazim sufficed to condition soil
with no history of carbendazim application to rapid degradation.
Previous application of the fungicide was not the only means of
inducing the phenomenon. When soil with a history of
carbendazim-treatment was mixed with untreated soil, the ability to
accelerate degradation was observed in the entire soil volume. This
capacity was maintained in soil for over 2 years without
intermediate carbendazim application (Yarden et al., 1987).
Bean plants grown to maturity in Delaware, USA, contained less
than 0.1 mg/kg total 14C residue in the edible beans following two
foliar applications of 1 kg a.i./ha of [2-14C]-carbendazim (as
Delsene 50% WP) at 25% and 50% bloom. Total 14C residues in the
bean foliage decreased from about 5 mg/kg one week after the second
spraying to 0.2 mg/kg three weeks later. The total 14C residue in
edible beans was less than 0.1 mg/kg one week after the second
spraying. Of the total 14C in the edible beans and foliage, 89-95%
was intact free carbendazim and 2-8% was free 2-AB. An additional
1-3% of the 14C was found as ß-glycosidic conjugates of
carbendazim and 2-AB (Han, 1983a).
Chiba & Veres (1981) applied benomyl to apple trees as Benlate
50% WP at a rate of 1.7 kg/ha. Three successive applications were
made in 1977 and a single spray was applied in 1979. Between 3 and 7
days after application there was a marked reduction of about 50% in
benomyl residues from an initial level of about 110 mg/kg. This fall
in benomyl was accompanied by a doubling in the level of carbendazim
residues over the same period due to benomyl degradation to
carbendazim. Within 46 days of the single appli cation in 1979,
benomyl residues fell to 0.63 mg/kg foliage and carbendazim was
present at 1.2 mg/kg. Following the three sprayings in 1977 (at 0,
13 and 27 days after the initial application), residue levels were
2.6 and 17.1 mg/kg foliage for benomyl and carbendazim 83 days after
the first spraying. Both experiments showed an exponential fall in
benomyl residues but the rate of decline was much slower in the case
of the more persistent metabolite.
4.2.2 Abiotic degradation
The hydrolytic stability of carbendazim at pH 5, 7, and 9 and a
nominal temperature of 22, 50 and 70 °C was studied at intervals up
to 30 days. Elevation of temperature and pH increased carbendazim
degradation. The half-life calculated for the degradation of
carbendazim at pH 5 and at 22, 50 and 70 °C was 457, 108, and 29
days, respectively. At pH 7 and at 50 and 70 °C the half-life was 43
and 12 days (no appreciable decline at 22 °C). The half-life at pH 9
and at 22, 50, and 70 °C was 22, 1.4 and 0.3 days, respectively
(Purser, 1987).
Carbendazim was exposed to sunlight for 30 h (as a residue on
silica gel G) and less then 10% was lost after exposure.
Photo-oxidation of the benzene ring of carbendazim was the
predominant reaction with some guanine, carbomethoxyguanine, and
carbomethoxyurea detected. When carbendazim was applied to the
leaves of corn plants and exposed to sunlight for 18 h, no
photolysis products were detected in the extracts of plants (Fleeker
& Lacy, 1977).
4.2.3 Bioaccumulation
Bluegill sunfish ( Lepomis macrochirus) were exposed to
radiolabelled carbendazim concentrations of 0.018 or 0.17 mg/litre
for 4 weeks in a dynamic study designed to measure the
bioaccumulation of 14C residues in edible tissue, viscera,
remaining carcass and whole fish. A two-week depuration phase
followed the exposure phase. Results were similar at the two
exposure concentrations, the peak whole fish bioconcentration
factors (BCFs) being 27 and 23 at the low and high exposure levels,
respectively. The radioactivity was concentrated more in the viscera
than in other tissues, the peak viscera BCFs being 460 and 380 for
the low and high exposure levels, respectively. Very little
bioconcentration occurred in the muscle tissue (BCF = < 4) or the
remaining carcass (BCF = < 7). During the 14-day depuration phase,
> 94% of the peak level of radioactivity was lost from the whole
fish, viscera and muscle. The rate of loss from the carcass tissue
was lower (77% and 82% loss for the low and high exposure levels,
respectively) (Hutton et al., 1984).
When rainbow trout ( Oncorhynchus mykiss), channel catfish
( Ictalurus punctatus) and bluegill sunfish ( Lepomis macrochirus)
were injected intraperitoneally with carbendazim, branchial and
biliary excretion were the major pathways for the elimination
(Palawski & Knowles, 1986). In a separate experiment, the three fish
species were exposed to 45 µg carbendazim/litre for 96 h, except in
the case of catfish, which were exposed for 48 h. This was followed
by a 96-h depuration phase. Rainbow trout had the highest uptake
rate constant (1.78 per h) and bioconcentration factor (159) of the
three species. Much less carbendazim was accumulated by channel
catfish than by the other two species, but this residue level (0.44
µg/g) appeared to be lethal after 48 h of exposure. The elimination
rate constant and the biological half-life of carbendazim were
similar for rainbow trout and bluegill sunfish. However, the
elimination rate constant was greater and the biological half-life
shorter in channel catfish (13 h) than in the other two species.
5. ENVIRONMENTAL LEVELS AND HUMAN EXPOSURE
5.1 Environmental levels
5.1.1 Air, water and soil
The environmental levels in air, water and soil are discussed
in detail in chapter 4.
5.1.2 Food and feed
Levels of carbendazim in food and feed are indicated in section
5.2.
5.1.3 Terrestrial and aquatic organisms
Carbendazim levels in terrestrial and aquatic organisms are
discussed in detail in chapters 4 and 6.
5.2 General population exposure
The principal exposure of the general population to carbendazim
is through dietary exposure. It was recommended by the Joint FAO/WHO
Meeting on Pesticide Residues (JMPR) (FAO/WHO, 1985a) that all
maximum residue limits (MRLs) for benomyl, thiophanate-methyl and
carbendazim be listed as carbendazim (Tables 3 and 4).
5.2.1 Sweden
Monitoring data from Sweden are shown in Table 5 (FAO/WHO,
1988b). No further analysis to determine dietary intake was
performed.
5.2.2 The Netherlands
Over a period of two years (June 1976 to July 1978), "market
basket" samples for 16- to 18-year-old males, including 126
different food items, were purchased every two months. This age
group was chosen by the authors under the assumption that they had
the greatest food consumption. The food was prepared for eating
(including cooking) and combined in 12 different commodity groups,
and the concentrations of 78 different chemical pesticides were
determined. Using concentrations found in the total diet samples,
the daily intakes were calculated. For this sub-population, the
maximum intake of carbendazim was 0.6 mg/day (calculated using the
detection limit as the concentration of the non-detectable residues)
and the average dietary intake was 0.05 mg/day. The recommended ADI
is 0-0.01 mg/kg body weight (FAO/WHO, 1985a), corresponding to 0.6
mg/day for a 60-kg adult. Therefore, the maximum intake is at the
recommended ADI, whereas the average intake is 12 times below the
recommended ADI (de Vos et al., 1984).
Table 3. Benomyl/carbendazim/thiophanate-methyl residues in food in Swedena
Samples Swedish/imported No. of samples Samples with residues Residue level Median value
>0.20 mg/kg (mg/kg) (mg/kg)
1986
Pineapples imported 3 1 0.69
Grapes imported 20 3 0.17-0.35 0.26
Strawberries imported 7 1 0.29
Mangoes imported 17 4 0.20-1.82 0.70
Papayas imported 5 2 0.25-0.45
Pears Swedish 17 3 0.32-0.62 0.43
imported 45 7 0.20-0.45 0.34
Apples Swedish 78 17 0.20-0.72 0.40
imported 91 30 0.21-0.74 0.39
1987
Grapes imported 28 3 0.52-0.87 0.60
Strawberries imported 7 0.23
Mangoes imported 14 5 0.29-1.30 0.66
Papayas imported 4 2 0.86-1.14
Table 3 (contd).
Samples Swedish/imported No. of samples Samples with residues Residue level Median value
>0.20 mg/kg (mg/kg) (mg/kg)
Pears Swedish 14 1 0.52
imported 62 13 0.21-0.45 0.29
Apples Swedish 61 25 0.20-1.17 0.45
imported 94 12 0.21-0.82 0.36
a From: FAO/WHO (1988b)
Table 4. National Maximum Residue Limits (mg/kg) for certain commoditiesa
banana cereal cherries citrus bean cucumber peach pome fruit strawberries grapes
Australia 1 0.05 5 10 3 3 5 5 6 2
Austria 0.2 0.5 7 1 0.5 2 1.5 3
Belgium 2 0.5 2 2 0.5 2 5 5 2
Brazil 1 0.5 10 10 2 0.5 10 5 5 10
Bulgaria 0.5 10 5 5 10
Canada 5 10 1 0.5 10 5 5 5
Denmark 2 0.1 2 5 2 2 2 2 5 5
France 1 1.5 6
Finland 0.2 1 2 0.5 0.5 1 1 1
Germany 0.2 0.5 2 7 1 0.5 2 2 3
Hungary 2 1
Israel 10 10 10 5 10
Italy 0.5 0.5 1 1
Mexico 10 2 11 5 7 5 10
Netherlands 3 0.1 3 4 3 3 3 3 3 3
New Zealand 5 1 5 5 2 2 5 5 5 5
Spain (guidelines) 1 0.5 5 7 2 2 5 5 1 5
Table 4 (contd).
banana cereal cherries citrus bean cucumber peach pome fruit strawberries grapes
Switzerland 1 0.2 3 7 0.2 0.1 3 3 3 3
United Kingdom
(proposed) 1 0.5 10 0.5 10 5 5 10
USA 1 0.2 15 10 2 1 15 7 5 10
USSR 1 0.5 10 10 2 0.5 10 5 5 5
Yugoslavia 0.1 7 0.5 0.1 2 0.5 2
a From: FAO/WHO (1988a)
Table 5. Proposed Maximum Residue Limits for carbendazim from any
sourcea
Commodity MRL (mg/kg) Applicationb
Apricot 10c B,C
Asparagus 0.1d B,T
Avocado 0.5 B
Banana 1c B,C,T
Barley straw and fodder, dry 2 B
Bean fodder 50 C
Beans, dry 2 B
Berries and other small fruit 5 B,C,T
Brussel sprouts 0.5 B
Broad bean 2 T
Carrot 5c C,T
Cattle meat 0.1d B
Celery 2 B,C
Cereal grains 0.5 B,C,T
Cherries 10c B,C,T
Citrus fruits 10c B,C,T
Coffee beans 0.1d C
Common beane 2 C
Cucumber 0.5 B,C,T
Eggs (poultry) 0.1d B,T
Egg plant 0.5 C
Gherkin 2 C,T
Hops, dry 50 C
Lettuce, head 5 B,C,T
Mango 2 B
Melons, except watermelons 2c B,C
Milk 0.1d B
Mushrooms 1 B,C,T
Nectarine 2 B
Onion, bulb 2 C,T
Peach 10c B,C,T
Peanut 0.1d B,C
Peanut fodder 5 B,C
Peppers 5 C
Pineapple 20c B
Plums (including prunes) 2c B,C,T
Pome fruit 5c B,C,T
Potato 3c,f B,C
Poultry meat 0.1d B,T
Rape seed 0.05d C
Rice straw and fodder, dry 15 B,C,T
Sheep meat 0.1d B
Soya bean, dry 0.2 C
Soya bean fodder 0.1d C
Table 5 (contd).
Commodity MRL (mg/kg) Applicationb
Squash, summer 0.5 B
Sugar beet 0.1d B,C,T
Sugar beet leaves on tops 10 B,C,T
Swedeg 0.1d C
Sweet potato 1 B
Taro 0.1d B
Tomato 5 B,C,T
Tree nuts 0.1d B
Wheat straw and fodder, dry 5 B
Winter squash 0.5 B
a From: FAO/WHO (1988b)
b B = benomyl; C = carbendazim; T = thiophanate-methyl
c MRL based on post-harvest use
d At or about the limit of detection
e JMPR recommended 2 mg/kg for dry, dwarf, lima and snap beans.
These are all covered by "VP 0526, Common bean" and "VP 0071,
Beans, dry" in the new classification
f washed before analysis
g Described as rutabagas in 1983 recommendation
5.2.3 National maximum residue limits
National MRLs for certain commodities are listed in Table 4
(FAO/WHO, 1988a).
A complete list of MRLs for carbendazim, including new
proposals and an indication of the source of the data (application
of benomyl, carbendazim, or thiophanate-methyl) on which the MRL is
based, is given in Table 5 (FAO/WHO, 1988b).
5.3 Occupational exposure during manufacture, formulation, or use
5.3.1 Exposure during manufacture
Levels of carbendazim, monitored as worker inhalation exposure,
in a major manufacturing facility (Du Pont) were reviewed from 1986
to 1989. The average level of carbendazim was less than 0.3 mg/m3.
Table 6 lists established inhalation exposure limits for benomyl and
carbendazim.
� 5.3.2 Exposure during use
Although no studies have been performed with carbendazim,
potential dermal and respiratory exposure to benomyl wettable powder
formulation in actual-use situations was determined for tank loading
and mixing for aerial application, re-entry into treated crops, and
home use (garden, ornamental and greenhouse). For crop treatments,
approximately 17 kg benomyl (formulation) was handled per cycle.
Maximum exposure occurred in the loading and mixing operation for
aerial application, where dermal exposure was 26 mg benomyl per
mixing cycle, primarily to hands and forearms (90%), and respiratory
exposure averaged 0.08 mg benomyl. Re-entry data revealed dermal and
respiratory exposures of 5.9 mg/h and < 0.002 mg/h, respectively.
Home-use situations (application of 7 to 8 litres benomyl in
hand-held compressed air sprayers) produced exposures of 1 mg and
0.003 mg per application cycle for the dermal and respiratory
routes, respectively (Everhart & Holt 1982).
Table 6. Established inhalation exposure limitsa
Country and agency Compound TWAb STELc
(mg/m3) (mg/m3)
Australia benomyl 10 -
Belgium benomyl 10 -
Denmark benomyl 5 -
Finland benomyl 10 30
France benomyl 10 -
Switzerland benomyl 10 -
United Kingdom benomyl 10 15
USA: ACGIHd benomyl 10 -
USA: NIOSHe/OSHAf benomyl 10 -
(inhalable dust)
USA: NIOSH/OSHA benomyl 5 -
(respirable dust)
USSR carbendazim - 0.1
a From: ILO (1991)
b Time-weighted average
c Short-term exposure limit
d American Conference of Governmental Industrial Hygienists
e National Institute of Occupational Safety and Health
f Occupational Safety and Health Administration
6. KINETICS AND METABOLISM
Carbendazim is extensively metabolized by animals as described
in detail in section 6.3. Metabolite names and structures are given
in Table 7, Fig. 1 and Fig. 2.
6.1 Absorption
Male albino rats were orally administered a single
14C-carbendazim dose of 12 mg/kg as a solution in diethyl
glycol-ethanol. Based on urinary excretion of 14C-carbendazim and
its metabolites, 5-HBC and 2-AB, the absorption was determined to be
about 85% (Krechniak & Klosowska, 1986).
In a study by Monson (1990), young male and female
Sprague-Dawley rats were administered a single dose via gavage with
14C-radiolabelled [phenyl(U)-14C]-carbendazim (94% pure,
suspended in corn oil) at either a low (50 mg/kg) or a high (1000
mg/kg) dose, and excretion of the radiolabel was monitored every 12
h for 72 h. Most of the radioactivity was excreted by 72 h. The
percentages of originally administered radioactivity (i.e.
14C-carbendazim equivalent) recovered were: a) in urine, 61.7
(male), 53.8 (female) for low dose, 41.4 (male) and 40.7 (female)
for high dose; and b) in faeces, 24.4 (male), 33.2 (female) for low
dose and 61.9 (male) and 69.5 (female) for high dose. The author
concluded that: a) > 98% of the administered dose was recovered in
urine and faeces in all test groups; and b) the absorption
efficiency of carbendazim was approximately 80% of the actual
administered dose for all dose levels, based on the level of
radioactivity appearing in urine and the sum of all metabolites
appearing in faeces as the result of hepatic metabolism (Monson,
1990).
6.2 Distribution and accumulation
In a rat gavage study, [phenyl(U)-14C]-carbendazim was
administered to Sprague-Dawley rats (five rats per sex per group)
using three dosing regimes: a single oral dose of 50 mg/kg; a single
oral dose of 50 mg/kg following pre-conditioning gavage of
non-labelled carbendazim (50 mg/kg) for 14 days; and a single oral
dose of 1000 mg/kg. All rats were sacrificed 72 h after the last
dose. Tissue distribution data showed lack of bioconcentration of
radiolabelled compound. The highest concentrations of radio labelled
tissue residues (less than 1% of the dose) were detected in the
residual carcass and liver (Monson, 1990). This study is discussed
in detail in sections 6.3 and 6.4.
Table 7. Carbendazim and its metabolites in animalsa
Code name Chemical name
Carbendazim (MBC) methyl (1-H-benzimidazol-2-yl)carbamate
5-HBC methyl (5-hydroxy-1H-benzimidazol-2-yl)
carbamate
4-HBC methyl (4-hydroxy-1H-benzimidazol-2-yl)
carbamate
5-HBC-Sb 2-[(methoxycarbonyl)amino]-1H-benzimidazol-5-
yl hydrogen sulfate
5-HBC-Gc 2-[(methoxycarbonyl)amino]-1H-benzimidazol-
5-yl] ß-D-glucopyranosiduronic acid
MBC-4,5-epoxide
MBC-5,6-epoxide
MBC-4,5-dihydrodiol (4,5-dihydro-4,5-dihydroxy-1H-benzimidazol-
2-yl) carbamate
MBC-5,6-dihydrodiol (5,6-dihydro-5,6-dihydroxy-1H-benzimidazol-
2-yl) carbamate
MBC-4,5-diol
MBC-5,6-diol
5-OH-6-GS-MBCd S-[5,6-dihydro-5-hydroxy-2-(methoxycarbonyl
amino)-1H-benzimidazol-4-yl]glutathione
5-OH-4-GS-MBC S-[4,5-dihydro-5-hydroxy-2-(methoxycarbonyl
amino)-1H-benzimidazol-4-yl]glutathione
5,6-HOBC-N-oxide methyl (6-hydroxy-5-oxo-5H-benzimidazol-2-
yl)-carbamate-N-oxide
5,6-HOBC-N-oxide-G 2-[(methoxycarbonyl)amino]-6-oxo-6H-
benzimidazol-5-yl] ß-D-glucopyranosiduronic
acid-N-oxide
5,6-DHBC methyl (5,6-dihydroxy-1H-benzimidazol-2-yl)
carbamate
Table 7 (contd).
Code name Chemical name
5,6-DHBC-G 6-hydroxy-2-[(methoxycarbonyl)amino]-1H-
benzimidazol-5-yl] ß-D-glucopyranosiduronic
acid
5,6-DHBC-S 6-hydroxy-2-[(methoxycarbonyl)amino]-1H-
benzimidazol-5-yl 5-(hydrogen sulfate)
2-AB 2-aminobenzimidazole
2-AB dihydrodiol 2-amino-4,5-dihydro-4,5-dihydroxy-1H-
benzimidazol
5-HAB 5-hydroxy-2-aminobenzimidazole
a From: Krechniak & Klosowska (1986); Monson (1986, 1990)
b S = conjugate with sulfuric acid
c G = conjugate with glucuronic acid
d GS = conjugate with glutathione
In a study by Krechniak & Klosowska (1986), male albino rats
were administered by gavage a single dose of 12 mg carbendazim per
kg and the distribution of carbendazim and its metabolites among
subcellular liver fractions was determined 1.5 h after dosing. The
distribution was not uniform and was not dependent on the lipid
content of the fractions. The highest relative concentration of
unchanged carbendazim was in the mitochondria. The highest relative
concentration of 5-HBC was in the cytosol and that of 2-AB was in
the microsomes.
Ten hens were individually dosed for six consecutive days with
0.625 mg [2-14C]-carbendazim at a rate equivalent to 5 mg/kg in
the average total daily feed. An additional 10 hens were dosed daily
with 12.5 mg at a rate equivalent to 120 mg/kg in the average total
daily feed. The hens were sacrificed 24 h after the sixth dose and
muscle, kidney, liver and fat were sampled. Carbendazim was
extensively metabolized to 5-HBC and methyl (4,5-dihydro-4,5-
dihydroxy-1H-benzimidazol-2-yl) carbamate. The concentration of
radioactivity, calculated as mg carbendazim per kg, in the high-dose
hens was 2.63 (liver), 1.74 (kidney), 0.06 (thigh muscle), 0.05
(breast muscle), 0.03 (fat) and 0.63 (day-6 eggs). Approximately 73%
of the total radioactivity in the day-6 eggs was identified as 5-HBC
(0.26 mg/kg) and unchanged carbendazim (0.15 mg/kg) (Monson, 1986).
In another feeding study, groups of 20 hens were fed
carbendazim daily at levels of 0, 5, 15 and 100 mg/kg diet for 28
days. Eggs were collected daily and faeces once a week. After 28
days, 15 hens in each group were sacrificed and samples taken for
analysis. The remaining hens were fed a carbendazim-free diet for a
further week and then sacrificed. The majority of the material fed
was rapidly eliminated as parent compound or as 5-HBC. The only
residues found in any of the samples of blood, fat, liver, kidney,
tissue or eggs were in the eggs from the 100-mg/kg group. However,
the residues (up to 0.1 mg carbendazim/kg and 0.36 mg 5-HBC/kg each)
decreased to < 0.05 mg/kg within 4 days of withdrawal of treatment
(Eckert et al., 1985).
A lactating Holstein cow was dosed by capsule twice daily (483
mg [2-14C]-carbendazim each dose), equivalent to 50 mg/kg in the
average total daily feed, for five consecutive days. Samples of milk
were collected at each dosing. Approximately 17 h after the tenth
dose, the cow was sacrificed. Small amounts of radioactivity
corresponding to carbendazim or its metabolites were found in the
liver (2.62 mg/kg) and kidney (0.45 mg/kg), but no significant
amounts (< 0.09 mg/kg) were detected in other tissues or fat. Only
2.7% of the liver radioactivity was 5-HBC whereas 41% and 3% of the
kidney radioactivity corresponded to 5-HBC and 4-HBC, respectively.
14C residue levels in the milk averaged 0.25 mg/kg (calculated as
carbendazim) of which 0.11 mg/kg was 5-HBC and 0.05 mg/kg 4-HBC. No
carbendazim (< 0.01 mg/kg) was detected in the milk (Monson, 1985).
Twelve non-lactating female goats were administered a
feed-rate-equivalent [phenyl(U)-14C]-carbendazim dose of at least
50 mg/kg (range: 50-101 mg/kg) once a day for up to 30 days. A
plateau of 14C residues in the liver was achieved within 2 weeks
of dose initiation and was calculated to be 9.48 mg/kg of liver
(group mean of the total radiolabelled liver residues for goats
sacrificed 2, 3 and 4 weeks after initiation of dosing). The total
14C residue levels in the liver decreased to 5.17, 3.55 and 1.67
mg/kg at 1, 2 and 3 weeks, respectively, after discontinuing dosing.
Based on this data, the elimination half-life for the total 14C
residues from the liver was calculated to be approximately 9 days.
The half-life for removal of carbendazim from the general
circulation, based on 14C-carbendazim equivalent whole blood
levels, was approximately 10 h. The results of this study suggest
that levels of carbendazim-derived residues do not continue to
accumulate beyond 2 weeks when goats are exposed to a constant
feed-level of 50 mg carbendazim/kg. Furthermore, discontinuation of
exposure results in a clearing of residues from the liver (Johnson,
1988).
6.3 Metabolic transformation
In a rat gavage study, carbendazim was found to be extensively
metabolized. Three dosing regimes (five rats of each sex per group)
were used: a single oral dose of 50 mg/kg (low dose); a single oral
dose of 50 mg/kg following pre-conditioning gavage with
non-radiolabelled carbendazim of 50 mg/kg for 14 days
(pre-conditioned low dose); and a single oral dose of 1000 mg/kg
(high dose). The 48-h urine from the low-dose and the high-dose
rats, and the 14-day urine from the pre-conditioned low-dose group
were collected. The total recovery from urine was 61.5 and 61.7% of
given doses for the low-dose and pre-conditioned low-dose male
groups, 53.2 and 59.3% for the low-dose and pre-conditioned low-dose
female groups, and 39 and 41% for both male and female high-dose
groups, respectively. 5-HBC-S (21-43% of given dose) was identified
as the main metabolite, except in the case of the pre-conditioned
low-dose and the high-dose female rat groups (5.5-10%), while in all
female rat groups 5,6-HOBC-N-oxide-G (10-19%) was predominant. Both
5,6-DHBC-S and 5,6-DHBC-G were identified as minor metabolites.
In the same study, the faeces were collected at the same
periods as the urine. The total recovery from faeces was about 24%
for the low-dose and pre-conditioned low-dose male groups, 33-38%
for the low-dose and pre-conditioned low-dose female groups, and
higher (> 60%) for both male and female high-dose groups. Unchanged
carbendazim was about 10-15% of the given dose in the faeces of
high-dose rats (Monson, 1990). The proposed metabolic pathway for
carbendazim in rats is given in Fig. 1.
NMRI mice and Wistar rats of both sexes were given carbendazim,
via gavage, as a single dose of 3 and 300 mg/kg, respectively. Urine
was collected during the first 6 h, after which the animals were
killed. Almost all the metabolites in urine were conjugated with
sulfuric acid. Cleavage of these conjugates by ß-glucuronidase/
arylsulfatase released 5-HBC as the only metabolite extractable from
water. Mouse urine contained a greater amount of compounds that
remained polar after enzyme treatment than the corresponding urine
of rats. Analyses revealed no sex differences (Dorn et al., 1983).
In a further study, male albino rats were administered a single
intravenous dose of 12 mg carbendazim/kg as a solution in
diethylene-glycol. The composition of the measured radioactivity in
urine 12 h after dosing was 94% as 5-HBC, 3% as 2-AB, and 3% as
carbendazim (Krechniak & Klosowska, 1986).
In hens dosed with [2-14C]-carbendazim (5 and 120 mg/kg in
the daily feed), carbendazim was metabolized to 5-HBC, 4-HBC,
4,5-dihydrodiol-MBC and its sulfuric acid conjugate, and also to
2-AB (Monson, 1986). The proposed metabolic pathway in laying hens
is given in Fig. 2.
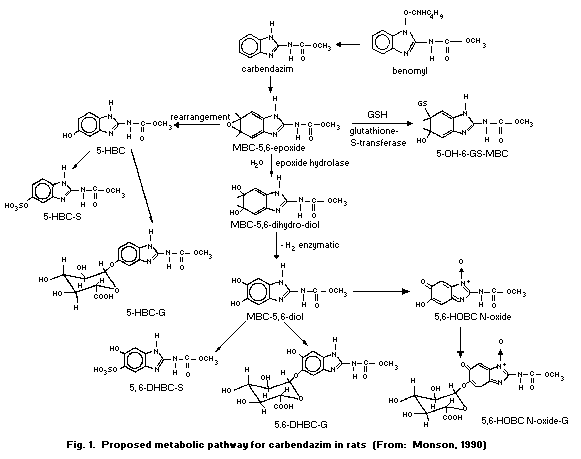
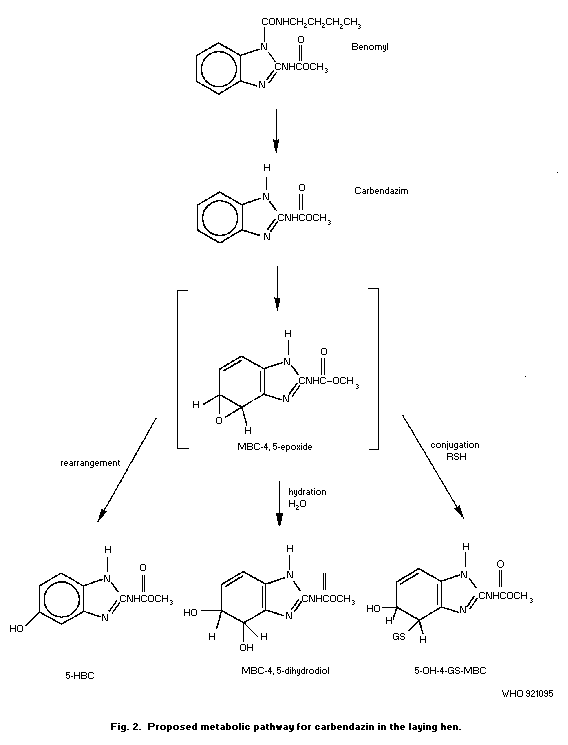 The metabolic fate of carbendazim in the liver was examined in
non-lactating female goats administered a feed-rate-equivalent
[phenyl(U)-14C]-carbendazim dose of 50 mg/kg once a day for 30
days. Extraction of liver homogenate from goats sacrificed 4 weeks
after initiation of dosing, i.e. when the 14C residues in the
liver had reached a plateau, indicated that the major
ethyl-acetate-extractable and identifiable radiolabelled residues in
the liver were 5-HBC (2 to 3 mg/kg) and carbendazim (approximately
0.2 mg/kg). Bound non-extractable 14C residues in the liver
reached a plateau level of approximately 1 mg/kg (Johnson, 1988).
Monson (1991) analysed, via Raney nickel desulfurization and
acid dehydration, the release and characterization of bound
carbendazim metabolites in diary cow, goat, hen and rat liver after
treatment with 14C-carbendazim. Using this technique, he was able
to show that bound 14C residue was released from the liver of cows
(76% bound before desulfurization and 36% bound after
desulfurization) and hens (58% bound before desulfurization and 19%
bound after desulfurization). The major part of the reduced residue
was identified as 5-HBC, 5,6-HOBC or carbendazim, suggesting that
the bound liver residue consisted of conjugates of
benzimidazole-related products and not natural products resulting
from breakdown and incorporation.
Benomyl and carbendazim are metabolized in fish to 5-HBC (Du
Pont, 1972).
6.4 Elimination and excretion
Carbendazim is rapidly excreted in the urine and faeces.
In a rat gavage study (Monson, 1990), [phenyl(U)-14C]-
carbendazim was administered to Sprague-Dawley rats using three
dosing regimes: a single oral dose of 50 mg/kg (low dose); a single
oral dose of 50 mg/kg following pre-conditioning gavage with
unlabelled carbendazim of 50 mg/kg for 14 days (pre-conditioned low
dose); and a single oral dose of 1000 mg/kg (high dose). Each dosing
group comprised five animals of each sex. A preliminary study
conducted with two rats of each sex, each rat having received a
single oral dose of 50 mg/kg, demonstrated that 95% of the
radioactivity excreted in the urine and faeces was recovered within
72 h after dosing and that < 1% of the dose was expired as volatile
metabolites. In the full study, > 98% of the recovered
radioactivity was excreted by the time of sacrifice (i.e. 72 h after
dosing) in the case of each group of rats. Urinary excretion
accounted for 62% to 66% of the dose in males and 54% to 62% of the
dose in low-dose and pre-conditioned low-dose females. In the
high-dose group, this pathway accounted for 41% of the dose in all
animals. Elimination of radiolabel in faeces accounted for virtually
all of the remaining radiolabel.
A lactating Holstein cow was dosed by capsule twice daily (483
mg [2-14C]-carbendazim each dose), equivalent to 50 mg/kg in the
average total daily diet, for five consecutive days, and samples of
urine, faeces and milk were collected at each dosing. Five days
after the initial dose, 65% of the radiolabel had been excreted in
the urine, 21% in the faeces and 0.4% in the milk (a total of
86.4%). Radioactive residues in the urine comprised 48% 5-HBC and 3%
polar water-soluble metabolites (Monson, 1985).
In a further study, lactating Holstein cows were dosed by the
dietary route with 0, 2, 10 or 50 mg carbendazim/kg diet for 28
days. The highest levels of carbendazim metabolites in the urine and
faeces were found in cows fed 50 mg/kg. The highest levels found in
urine were 12.56 mg 5-HBC/litre and 1.29 mg 4-HBC per litre. In the
faeces 3.81 mg 5-HBC/kg and 0.99 mg carbendazim/kg were detected
(but not in the same cow). No carbendazim residues were found in the
urine (Hughes, 1984).
Groups of hens were dosed with [2-14C]-carbendazim at a rate
equivalent to 5 and 120 mg/kg in the average total daily feed for
six consecutive days and were sacrificed 24 h after the sixth dose.
At sacrifice, an average of 95% and 92% of the radioactive doses had
been excreted in the low-dose and high-dose hens, respectively
(Monson, 1986).
6.5 Reaction with body components
In a study by Guengerich (1981), the effects of carbendazim on
hepatic enzyme induction were studied in male and female Crl-CD
rats. The treatment groups included animals fed for 28 days with
diets that contained carbendazim at concentrations of 0, 10, 30,
100, 300, 1000 or 3000 mg/kg. Microsomal epoxide hydrolase and
cytosolic glutathione- S-transferase were monitored in subcellular
fractions isolated from the livers of animals in each treatment
group. Liver weights were also recorded. Elevated liver weights were
observed at 1000 and 3000 mg carbendazim/kg in both male and female
rats. No apparent liver toxicity or effect on body weight was
observed. Carbendazim induced epoxide hydrolase in both sexes of
rats and mice at 1000 and 3000 mg/kg. Induction of
glutathione- S-transferase was observed at 3000 mg/kg. In general,
the level of induction seemed to be slightly greater in females than
males.
In the same study, but in a separate test, CD-1 male mice were
treated via gavage with carbendazim suspended in corn oil (0, 100
and 1000 mg/kg per day) for 5 days. The liver samples were
homogenized and subcellular fractions were prepared as above. Wet
liver weights, microsomal cytochrome P-450, NADPH-cytochrome-c
reductase, styrene-7,8-hydrolase, benzphetamine- N-demethylase,
benzo(a)pyrene hydroxylase and 7-ethoxy coumarin O-deethylase, and
cytosolic glutathione- S-transferase were measured. Those
parameters showing statistically significant increases over the
control values were styrene-7,8-hydrolase and glutathione- S-
transferase; 7-ethoxycoumarin O-deethylase showed a significant
decrease. It is noteworthy that the total microsomal cytochrome
P-450 level did not increase, indicating that a whole scale
microsomal induction phenomena was not induced by carbendazim even
at the higher treatment level. However, as shown by the increase in
microsomal styrene-7,8-hydrolase, some hepatic microsomal enzymes
are induced by in vivo carbendazim treatment (Guengerich, 1981).
There did not appear to be any substantial difference in enzyme
induction between rats and mice.
7. EFFECTS ON LABORATORY MAMMALS AND IN VITRO TEST SYSTEMS
7.1 Single exposure
The acute toxicity of carbendazim in several animal species is
summarized in Table 8. The LD50 values range from > 2000 to > 15
000 mg/kg for a wide variety of test animals and routes of
administration. A description of toxic effects is given in section
7.5.1.
7.2 Short-term exposure
7.2.1 Gavage
Groups of ChR-CD male rats (six per dose level) were gavaged
with 200, 3400 and 5000 mg carbendazim/kg per day, five times/week,
for two weeks. Two out of six rats died at the dose level of 3400
mg/kg per day. At all dose levels, gross and microscopic evidence of
adverse effects on testes and reduction or absence of sperm in the
epididymides was seen. Testes were small and discoloured, with
tubular degeneration and evidence of aspermatogenesis. At the dose
level of 3400 mg/kg per day, there were also morphological changes
in the duodenum (oedema and focal necrosis), bone marrow (reduction
in the blood-forming elements) and liver (decrease in the large
globular-shaped vacuoles) (Sherman, 1965; Sherman & Krauss, 1966).
Subchronic administration of 0, 16, 32 or 64 mg carbendazim per
kg per day by gavage for 90 days to four groups of 10 male and 10
female litter-mate weaning Wistar rats was carried out. The
erythrocyte counts for treated rats were lower than those of the
controls after 15 days of exposure. However, no clear dose-response
relationship was demonstrated after 30, 60 or 90 days of exposure. A
decrease was noted in leucocyte counts at 15 days. After 30 and 60
days, both sexes demonstrated transient decreases in lymphocyte
counts compared to controls. However, no clear dose-response
relationship was observed among the treated groups. No change was
noted in the activity of whole blood cholinesterase. Male rats
showed significantly increased alkaline phosphatase activity at a
dose level of 64 mg/kg per day. Blood urea levels were lower in
males at dose levels of 32 and 64 mg/kg per day after 90 days.
Increased serum bilirubin concentrations were observed in males and
females at 32 and 64 mg/kg per day and were attributable to
parenchymal cell damage, as shown by increased glutamic-pyruvic
transaminase activity. Dose-related changes in the liver ranged from
sparse infiltration by inflammatory cells to inflammatory and
degenerative changes. Tubular dilation and hydropic degeneration
were noted in the kidneys of the low-dose rats, and fibrosis and
congestion in the medium- and high-dose rats. Increased lung weight
was correlated with bronchopneumonic changes. Slight changes in
weights were reported for several organs (Janardhan et al., 1987).
The published information was difficult to evaluate due to the
variability of the results and absence of the raw data.
Table 8. Acute toxicity of carbendazim in animals
Chemical Species Sex Animals Route Vehicle Concentrationa Reference
per group (mg/kg body weight)
Carbendazim rat M/F 10 oral sesame oil LD50 > 15 000 Kramer & Weigand (1971)
(MBC) rat M 10 intraperitoneal 0.9% saline LD50 > 2000 Goodman & Sherman (1974)
and Tween 80
rat M 6 inhalation (1 h) dust ALC > 5.9 mg/litreb Sarver (1975)
rat F 5 dermal sesame oil ALD > 2000 Kramer & Weigand (1971)
mouse M 10 oral propylene glycol LD50 > 15 000 Til & Beems (1981)
mouse M/F 10 intraperitoneal sesame oil LD50 > 15 000 Scholz & Weigand (1972)
dog M/F 2 oral sesame oil ALD50 > 5000 Scholz & Weigand (1972)
guinea-pig M 10 oral corn oil LD50 > 5000 Dashiell (1975)
rabbit M 10 dermal aqueous paste LD50 > 10 000 Edwards (1974a)
75% wettable rat M/F 5 oral corn oil LD50 > 5000 Hinckle (1981)
powder
rat M/F 10 inhalation dust LC50 > 5 mg/litre Nash & Ferenz (1982)
rabbit M/F 5 dermal physiological saline LD50 > 2000 Ford (1982)
Benlate C rat M/F 5 oral water LD50 > 5000 Grandizio & Sarver (1987)
(50% wettable rabbit M/F 5 dermal aqueous paste LD50 > 2000 Vick & Brock (1987b)
powder)
a ALC = approximate lethal concentration; ALD = approximate lethal dose
b Time-weighted concentration
7.2.2 Feeding
7.2.2.1 Rat
Groups of ChR-CD rats (16 males and 16 females per group) were
fed carbendazim (72% a.i.) in the diet for 90 days at levels of 0,
100, 500 and 2500 mg/kg. The animals were observed daily for
behavioural changes and body weight, and food consumption were
recorded at weekly intervals. Haematological examinations were
conducted on 10 male and 10 female rats in each group at 30, 60 and
90 days. Routine urinalyses were performed on the same animals, and
plasma alkaline phosphatase and glutamic-pyruvic transaminase levels
were determined. After 90 to 98 days of continuous feeding, 10 male
and 10 female rats in each group were killed and selected organs
weighed. These and other organs were preserved for microscopic
examination. The six male and six female rats remaining in each
group after terminal sacrifice were used in a reproduction study
(see section 7.5.1). There were no signs of poisoning and no
compound-related effects on weight gain, food consumption or
haematological parameters. There were no control data for
biochemical determinations, urinalysis or differential white blood
counts. The average dose for the high-dose animals was 360 mg/kg per
day initially and 123-152 mg/kg per day at sacrifice. The
liver-to-body-weight ratio in females fed 2500 mg/kg diet was
slightly increased compared with control rats. There were no effects
on testicular weights in any of the treatment groups. Microscopic
examination of selected tissues and organs in the control and
high-dose groups demonstrated no adverse effects attributable to
carbendazim (Sherman, 1968).
7.2.2.2 Dog
Groups of one-year-old beagles (four males and four females per
group) were administered carbendazim (53% a.i.) in the diet for
three months at dietary levels of 0, 100, 500 and 2500 mg/kg. The
highest level was reduced to 1500 mg/kg because of reduced food
intake and decreased body weight. However, compound administration
was interrupted when animals were fed a control diet for a few days
and then fed with 1500 mg/kg again.
Food consumption and body weight data were recorded weekly, and
clinical laboratory examinations (including haematological,
biochemical and urinalysis measurements) were performed pre-test and
after 1, 2 and 3 months of feeding. At the end of the study all
animals were killed, selected organs were weighed, and these and
other organs were subjected to gross and microscopic evaluations.
No mortality or adverse clinical signs were observed over the
course of the study, and growth and food consumption were normal
(except at 1500-2500 mg/kg). Urinalysis measurements were unaffected
by treatment, and there were no dose-related effects on the
haematological values. Females at the mid-dose level showed a trend
toward increased cholesterol levels at 1, 2 and 3 months compared
with the pre-test and control values. High-dose females had
similarly elevated cholesterol levels. Organ-to-body weight changes
were observed in the case of the thymus of low- and mid-dose males
and the prostate of mid-dose males. All the weights for these organs
were increased compared with control values. However, only the
liver, kidney and testes were examined histologically in the low-
and mid-dose groups. Limited histopathological data did not indicate
compound-related effects (Sherman, 1970).
In a study by Til et al. (1972), groups of beagles (four males
and four females per group) were administered carbendazim in the
diet at levels of 0, 100, 300 and 1000 mg/kg for 13 weeks. The
highest level was increased to 2000 mg/kg after six weeks of
treatment. Body weight, haematological and blood chemistry
measurements, urinalyses and liver/kidney function tests were
performed periodically. Gross and microscopic examinations of all
animals were performed at the end of the study. There were no
reported compound-related effects on clinical behaviour, body
weight, food consumption, haematological parameters, kidney function
(phenol red excretion) or liver function (BSP retention)
examinations. Blood chemistry measurements were normal, except for a
slight decrease in albumin in mid- and high-dose males at 12 weeks.
These values differed from week 0 measurements only in high-dose
males. Urinalysis values were normal except for a high bacterial
count in high-dose females at week 13. The blood clotting time was
slightly reduced in high-dose dogs at week 12. There were slight
increases in relative liver and thyroid weights and a decrease in
relative heart weights in the highest-dose group compared with
controls. No microscopic changes that could be associated with
treatment were observed in these or any other organs. Carbendazim
appeared to be without adverse effects on beagles when incorporated
in the diet for 13 weeks at dietary levels of 300 mg/kg or less.
7.2.3 Dermal
New Zealand albino rabbits (six males/group) were treated with
0 or 2000 mg/kg of carbendazim, applied as a 50% aqueous paste to
shaved intact dorsal skin. The material was applied repeatedly, 6
h/day, for ten consecutive days. There were no adverse effects on
body weight, clinical symptoms, organ weights, gross pathology or
histopathology of selected organs. However, there was focal necrosis
of the epidermis and polymorphonuclear cell infiltration of the
dermis in five out of six exposed rabbits. No other effects were
observed (Dashiell, 1975).
7.3 Skin and eye irritation; sensitization
7.3.1 Dermal
The primary dermal irritation potential of Benlate C (50%
wettable powder) was evaluated by applying a 5-g aliquot for 4 h to
the clipped intact skin of six New Zealand White rabbits. Test sites
were evaluated for erythema, oedema and other evidence of dermal
effects, and were scored according to the Draize scale at 4, 24, 48
and 72 h after application. No dermal irritation was seen at any
time during the study (Vick & Brock, 1987a).
7.3.2 Eye
No eye irritation potential for technical carbendazim was seen
in six albino rabbits (Edwards, 1974b). In a study by Vick &
Valentine (1987), a 50% wettable powder (formulation) was evaluated
for acute eye irritation potential in six male New Zealand white
rabbits. The formulation produced slight corneal opacity, mild or
moderate conjunctival redness, and slight or mild conjunctival
oedema in all the rabbits. In addition, there was moderate iritis in
three of the rabbits and minimal blood-tinged discharge in one
rabbit. Microscopic examinations revealed no corneal injury in any
of the treated eyes. The treated eyes of the two other rabbits were
normal by 72 h. It was concluded that this formulation was a
moderate eye irritant.
7.3.3 Sensitization
Albino guinea-pigs (10 males) exposed to carbendazim, either
technical material or a 75% wettable powder formulation, presented
no evidence of dermal sensitization following either intradermal
injections or repeat applications to shaved intact skin (Ford,
1981).
A 50% carbendazim formulation was tested on the shaved intact
skin of 10 male and 10 female Duncan Hartley albino guinea-pigs.
Five male and five female guinea-pigs were treated with 80% ethanol
(in water) and served as vehicle control animals. Two male and two
female guinea-pigs were treated with the test material (as a solid)
at the challenge phase only and served as negative control animals.
1-Chloro-2,4-dinitrobenzene (DNCB) was tested as a 0.3% suspension
in 80% ethanol (in water) on the shaved intact skin of two male and
two female guinea-pigs as a positive control group. No irritation
was observed in the test or vehicle control guinea-pigs or in the
negative controls at the challenge phase. DNCB produced
sensitization in all treated animals (Martin et al., 1987).
7.4 Long-term exposure
7.4.1 Rat
Groups of weanling rats (36 male and 36 female ChR-CD albino
rats/group) were administered carbendazim (50-70% a.i.) in the diet
for 104 weeks at levels of 0, 100, 500, 2500 (increased to 10 000
mg/kg after 20 weeks) and 5000 mg/kg diet. Body weight and food
consumption were recorded weekly for the first year and twice a
month thereafter. Daily observations were made with respect to
behavioural changes and mortality. At periodic intervals throughout
the study, haematological, urinalysis and selected clinical
chemistry examinations were performed. After one year each group was
reduced to 30 male and 30 female rats by interim sacrifice for gross
and microscopic examinations. At the end of the study all surviving
animals were sacrificed and gross examination of tissues and organs
was made. Microscopic examinations were conducted on all tissues and
organs from the control and 2500-mg/kg groups, the livers of the
100- and 500-mg/kg groups, and the livers, kidneys, testes and bone
marrow from the 5000-mg/kg groups. Survival decreased during the
second year to approximately 50% for males and 39% for females in
all groups. Body weight gain was depressed for males and females in
the 2500-mg/kg group and for females in the 5000-mg/kg group,
compared to control groups. Food consumption did not differ among
the various groups. The average daily dose for the 500-mg/kg group
was 65 mg/kg body weight per day initially, 18 mg/kg body weight per
day at one year, and 15 mg/kg body weight per day at two years.
Haematological examinations demonstrated reduced erythrocyte count
and haemoglobin and haematocrit values for females at 9-24 months in
the 2500- and 5000-mg/kg groups, and for males at 24 months in the
2500-mg/kg group. There were no compound-related clinical
manifestations of toxicity and no effects observed in urinalysis
examination. Alkaline phosphatase and glutamic-pyruvic transaminase
activity varied throughout the test at 2500 and 5000 mg/kg but did
not demonstrate a consistent dose-response relationship. There were
no apparent differences in the organ weights or organ-to-body weight
measurements, except in the case of female livers in the 2500- and
5000-mg/kg groups. This increase in the liver-to-body weight ratio
was due to a reduction in body weight. Histopathological examination
of the livers did not demonstrate any compound-related effects.
Males in the 2500-mg/kg group presented a marginal increase in
diffuse testicular atrophy and prostatitis (Sherman, 1972).
In another 2-year rat study, groups of Wistar rats (60 males
and 60 females/group) were administered carbendazim (99% pure) in
the diet at levels of 0, 150, 300 and 2000 mg/kg diet for two years.
The dose of 2000 mg/kg was increased to 5000 mg/kg after one week
and then to 10 000 mg/kg after two weeks for the remainder of the
study. Animals were examined daily for clinical signs of toxicity.
Body weight and food consumption were measured regularly throughout
the study. Haematological (peripheral blood), blood chemistry
(orbital sinus) and urinalysis evaluations were conducted
periodically during the study. All animals were subjected to
complete gross necropsy, and selected organs were weighed. Tissues
were examined microscopically in 20 male and 20 female rats in the
control and high-dose groups. All tumours and gross abnormalities
were also examined histologically. There were no differences between
test groups and control animals concerning clinical signs of
toxicity or food consumption. Body weights were significantly
reduced in low-dose males from week 88 to term and in high-dose
females from week 12 to term. Urinalyses were comparable among all
groups. Of the haematological parameters examined, haemoglobin was
depressed in high-dose females at weeks 26, 52 and 103 and
haematocrit was depressed in high-dose females at week 103. There
were no compound-related effects in males. Serum glutamic
oxaloacetic transaminase (SGOT) activity was decreased in high-dose
males at term, but not in females. High-dose females had increased
serum glutamic pyruvic transaminase (SGPT) activity and decreased
total serum protein at study termination. There were no
compound-related effects on organ weights except for increased
relative liver weights in high-dose females. There were also no
compound-related effects on mortality, this being 50% at week 76 in
control males and at week 92 in treated males. There was 50%
mortality in control and low-dose females at week 88 and in mid- and
high-dose females at 92-96 weeks. Survival at termination of the
study was similar in all groups.
There were no histological differences between control and
treated groups except for an increased incidence of diffuse
proliferation of parafollicular cells of the thyroid in the
high-dose females (Til et al., 1976a).
7.4.2 Dog
In a one-year study, beagle dogs (five of each sex per group)
were fed diets containing 0, 100, 200 or 500 mg carbendazim/kg. The
dogs were weighed at regular intervals, and individual food
consumption was monitored throughout the study. Clinical pathology
evaluations were performed twice prior to the initiation of the
study and five times during the study, at 1, 3, 6, 9 and 12 months.
After one year, all dogs were killed and selected tissues were
examined microscopically. There were no statistical differences in
mean body weight that could be attributed to carbendazim exposure.
The mean daily food consumption in all treated groups of dogs was
similar to controls. None of the clinical observations were
attributable to carbendazim intake. Dogs fed 500 mg/kg had elevated
levels of serum cholesterol. These levels were statistically
significant for the males at 9 months and the females at 1 and 2
months. There were no compound-related microscopic lesions related
to carbendazim intake (Stadler, 1986).
Groups of beagles (four males and four females per group) were
administered carbendazim (53% a.i.) in the diet at dosage levels of
0, 100, 500 and 2500 mg/kg for two years. The dogs were 1-2 years of
age at the start of the test. Food consumption and body weight data
were obtained weekly and animals were examined daily for clinical
signs of toxicity. Haematological, biochemical and urinalysis
examinations were performed periodically throughout the study.
Interim sacrifice after one year was performed on one male and one
female from the control and 500-mg/kg groups. Organ weight, gross
necropsy and histopathological examinations were performed at the
end of the study. Only the livers and testes were examined
histologically in the 100- and 500-mg/kg groups. No mortality was
reported for the control or the 100- and 500-mg/kg dose groups. The
average daily intake for the 500-mg/kg dose group was 15.0-20 mg/kg
body weight per day initially, 14-18 mg/kg body weight per day after
one year and 10-16 mg/kg body weight per day after two years.
Haematological and urinalysis values were unaffected by treatment.
The dogs in the 500-mg/kg groups had increased levels of
cholesterol, blood urea nitrogen (BUN), total protein and SGPT.
Swollen vacuolated hepatic cells and marginal proliferation of the
portal triads with cellular infiltration was observed in one dog
sacrificed after one year, which had been fed 500 mg/kg. No
histopathological liver lesions were observed in animals fed 500
mg/kg diet at the end of the study. Although inflammatory and
fibrotic liver changes were observed in the 2500-mg/kg group, these
changes cannot be evaluated because of the uncertainty of the dosing
regime, of the time of exposure and of the number of dogs. There
were no noticeable effects on organ weights or on organ-to-body
weight ratios. Diffuse testicular atrophy and aspermatogenesis were
observed in males (two out of four) at 100 mg/kg but not at 500
mg/kg (Sherman, 1972).
In another 2-year dog study, groups of beagles (four males and
four females per group) were fed technical carbendazim in the diet
at dosage levels of 0, 150, 300 and 2000 mg/kg for 104 weeks. After
33 weeks the dose 2000 mg/kg was increased to 5000 mg/kg. The dogs
were 22-27 weeks old at the start of the study. Daily examinations
were made for clinical signs of poisoning or adverse behaviour. Body
weight and food consumption data were recorded regularly throughout
the study. At periodic intervals (weeks 13, 26, 52, 78 and 104),
haematological, blood chemistry and urinalysis measurements were
made. Liver function (BSP retention) and kidney function (phenol red
excretion) tests were conducted at weeks 26, 52 and 104. At the
conclusion of the 104 weeks of dietary administration, each dog was
sacrificed and gross and microscopic examination of tissues and
organs was performed. There was no mortality in any group except for
one female in the high-dose group which was killed in a moribund
state after week 36. Body weight was decreased in mid-dose males and
high-dose males and females. Food consumption was comparable among
all groups. Blood clotting times were significantly reduced in
high-dose males from week 13 to term, and slight decreases were
noted in high-dose females. Serum alkaline phosphatase activity was
increased in the high-dose group throughout the study. There were no
compound-related effects on SGPT or SGOT levels. All other
haematological and blood chemistry values were comparable with
control groups. There were no differences for BSP retention, phenol
red excretion or urine analysis values among the various groups.
Absolute liver and thyroid weights were significantly increased in
high-dose dogs. Relative liver, thyroid and pituitary weights were
also significantly increased at the high-dose level. There were no
reported microscopic changes in these organs related to treatment.
There was an increased incidence of prostatitis (3/4 versus 1/4) in
high-dose males compared with controls. Also noted in high-dose
males (1/4) was interstitial mononuclear inflammatory cell
infiltrates and atrophic tubules of the testes. It was concluded
that the feeding of carbendazim in the diet to dogs for two years
was without apparent adverse effects at levels up to and including
300 mg/kg (Reuzel et al., 1976).
7.4.3 Mouse
A description of studies on mice is given in section 7.7.
7.5 Reproduction, embryotoxicity and teratogenicity
7.5.1 Reproduction
Groups of ChR-CD rats (3 male and 16 female rats per group,
except that the high-dose group contained 20 females) were fed
carbendazim in the diet at dose levels of 0, 100, 500, 5000 and 10
000 mg/kg and subjected to a standard two-litter-per-generation,
three-generation reproduction study. The parental animals were fed
the experimental diet at 21 days of age and mated to produce the
F1 litter at 100 days of age. The number of matings, pregnancies
and number of young in each litter at birth was recorded. The
litters were culled to 10 pups/litter on day 4. The number of the
live pups was recorded on days 4, 12 and 21, as was pup weight at
weaning. The parents were mated again to produce the F1B litters.
The F1B litters were maintained on the respective diets for 110
days and then mated to produce the F2A and F2B litters. The
F3A and F3B litters were produced similarly. Gross and
histopathological examinations of selected tissues and organs were
performed on two males and two females in each of five F3litters
from the control, 5000- and 10 000-mg/kg groups. Reproduction
indices, including mating, fecundity, fertility, gestation,
viability and lactation, were calculated and compared with control
values. Carbendazim was without effect on fertility, gestation,
viability and lactation. However, the average litter weights at
weaning were reduced in all generations fed 5000 and 10 000 mg/kg.
Histopathological examination of F3B weanlings did not reveal any
effects that were considered compound-related (Sherman, 1972).
Two additional feeding studies (Sherman, 1968; Koeter et al.,
1976) were reviewed by the 1983 Joint FAO/WHO Meeting on Pesticide
Residues (JMPR), which concluded that both reports had limitations.
For the Sherman (1968) study, it was stated that the "data presented
were extremely limited and submitted as group data only. There were
no pregnancies at 100 mg/kg diet for either F1a or F1b matings.
There were no apparent effects on the reproduction indices on
weanling weights. However, the fertility index for all groups, which
was 33-67% prevents any meaningful interpretation of the data"
(FAO/WHO, 1985b). For the study of Koeter et al. (1976), the 1983
JMPR concluded that "although there were no apparent adverse effects
on reproduction and no teratogenic effects at dietary levels of
carbendazim up to and including 2000 mg/kg, there were no individual
animal data presented. Histopathology of animals in the four-week
study was incomplete and did not include evaluations of spleen or
ovaries. Such additional data are needed to confirm the absence of
adverse effects in this three-generation reproduction study in rats"
(FAO/WHO, 1985b).
A serial breeding technique was used to evaluate the fertility
of male Sprague-Dawley rats after exposure by gavage to 10 daily
doses of carbendazim (400 mg/kg per day). Proven-fertile males (90
days old) were bred with a new female each week. Breeding began on
the third day of treatment and continued for 32 weeks after the last
day of chemical exposure. Twelve days after each breeding period,
the females were killed, their uteri were examined for resorptions,
and the number of dead and viable fetuses was determined. All males
were killed 35 weeks after the exposure, and testicular tissue was
prepared for histopathological examination by vascular perfusion.
The fertility (as indicated by the number of pregnant females) of
males in the carbendazim-treated group was depressed during the
first post-exposure week; 10 of the 24 treated males failed to
induce a pregnancy, as compared with no failure in the control
group. By the fifth post-exposure week, 16 of the 24
carbendazim-treated males were infertile. Of these 16 males, 4
recovered fertility after being infertile for 5-11 consecutive
breeding periods. However, 12 of the males did not recover fertility
during the remainder of the 32 week post-exposure period.
Histological examinations of testicular sections 245 days after the
exposure revealed that the exposure to carbendazim had caused severe
seminiferous tubular atrophy (> 85% of tubules were atrophic) in
those treated males that failed to recover fertility. The
seminiferous tubules of these males often showed "Sertoli cell only"
syndrome epithelium surrounded by a thickened basement membrane.
Less than 2% of the tubules contained spermatozoa in the lumen. The
seminiferous tubules of the carbendazim-treated males that recovered
fertility had various numbers of atrophic tubules (13-85%) 245 days
after the exposure (Carter et al., 1987).
In another study, groups of male and female rats (8-12 of each
sex per group) were administered 0, 50, 100, 200 or 400 mg
carbendazim/kg per day by gavage from weaning through puberty,
gestation and lactation, and they were mated at 84 days of age. The
male rats were killed on day 104-106 while the female rats were
killed on day 27 postpartum. A similar study was conducted with
Syrian hamsters administered 0 or 400 mg/kg per day. In the parental
generation, various landmarks of puberty were measured. In females,
estrous cyclicity, litter size, the number of implants, organ
weights and histological status were assessed. In males, organ
weights, testicular and epididymal sperm counts, sperm motility,
sperm morphology, testicular histological status, and endocrine
parameters were assessed. In addition, the growth, viability and
reproductive function of the offspring (F1) were observed during a
4-month period of continuous breeding. Males were killed at 5 months
for histopathological investigation. In the parental generation of
both species, carbendazim did not alter pubertal development, growth
or viability. The reproductive potential of rats treated with
carbendazim with 200 and 400 mg/kg per day was reduced due to
effects on sperm production and fetal viability. In the male rat,
carbendazim treatment with 200 or 400 mg/kg per day markedly altered
sperm morphology, testicular and epididymal weights, sperm numbers
and testicular histology. Fertility, sperm motility and hormonal
levels were altered primarily in those males fed 200 or 400 mg/kg
per day that exhibited very low sperm counts. A statistically
significant reduction in caudal epididymal sperm count was noted at
dose levels of 50 mg/kg per day or more. Testicular and epididymal
sperm counts in male hamsters were significantly lower (about 21%)
in the carbendazim-treated males compared with the control males. In
the F1 male hamsters, testis and seminal vesicle weights and
epididymal sperm counts were significantly reduced by prenatal
exposure to carbendazim at 400 mg/kg per day. In the parental female
rats, carbendazim administration caused postimplantation losses at
400 mg/kg per day and a few malformed rat pups were found in litters
at 100 and 200 mg/kg per day. Litter size was significantly reduced
at 200 and 400 mg/kg per day. Overall, carbendazim was less toxic to
the hamster than to the rat (Gray et al., 1988, 1990).
Since spermatogenesis is an androgen-dependent process, the
effects of carbendazim (0-400 mg/kg per day by gavage) on the
endocrine function of the rat testes were investigated. Following
subchronic (85 day) exposure, serum hormone levels, including those
of pituitary luteinizing hormone (LH), follicle-stimulating hormone
(FSH), thyroid-stimulating hormone (TSH), prolactin (PRL), and
androgen-binding protein (ABP) and testosterone in testicular fluids
(interstitial fluid and seminiferous tubule fluid) were measured. In
addition, the functional capacity of the Leydig cell to secrete
testosterone was assessed in vitro following human chorionic
gonadotrophin (HCG) challenge. Subchronic treatment with carbendazim
at doses of 50-100 mg/kg per day had no effect on pituitary or
testicular hormone concentrations; dosing with 200 mg/kg per day
elevated the testosterone concentration in the seminiferous tubule
fluid and the ABP tubule fluid without affecting serum testosterone
or ABP concentrations. The dose of 400 mg/kg per day resulted in
increased concentrations of both testosterone and ABP in the
interstitial fluid and seminiferous tubule fluid and of serum ABP,
but no change in serum testosterone level. These hormonal changes
are consistent with carbendazim effects on the gonads and resultant
testicular atrophy. This endocrine profile is similar to the
"Sertoli cell only" syndrome discussed by de Krester (1977). Thus,
the elevated seminiferous tubule fluid testosterone concentrations
may be a result of two factors: (a) greater release of testosterone
by the Leydig cells into the interstitial fluid; and/or (b)
decreased testosterone outflow from the testis into the general
circulation. In addition, increased ABP in the interstitial fluid
may reflect a change in the relative secretion of ABP into the
interstitial fluid and the seminiferous tubules (Rehnberg et al.,
1989).
Since it is possible that extragonadal changes contributed to
the appearance of the altered testicular endocrine profile described
above, a further study focused on the presence of concurrent changes
in the hypothalamus and pituitary control of the testes. In this
study, rats were gavaged with carbendazim (50, 100, 200 or 400 mg/kg
per day) for 85 days (Gray et al., 1988). A dose-related increase in
serum FSH and LH was noted, but values for prolactin and
thyroid-stimulating hormone remained unchanged. No statistical
differences in gonadotropin-releasing hormone (GnRH) concentrations
were present in the mediobasal hypothalamus, although an elevation
in anterior hypothalamic GnRH values was found at the 50-mg/kg dose,
followed by a dose-related decline. These findings would suggest
that carbendazim-induced testicular damage is accompanied by
compensatory changes in the hypothalmic and pituitary regulation of
the testes (Goldman et al., 1989).
The above studies imply that carbendazim acts directly on the
testes to produce a number of hormonal and pathological changes.
Consistent with this hypothesis are the results of the study of
Nakai et al. (1992), in which the effects of carbendazim on the
testes, efferent ductules and spermatozoa were determined after a
single oral dose in male Sprague-Dawley rats (Charles Rivers). In
the first experiment, groups of 86-day-old rats were treated with 0
or 400 mg carbendazim/kg and killed 2, 4 or 8 h later on the same
day or 1, 4, 8, 16 or 32 days after dosing. The effect of
carbendazim was first noted at 8 h as an increase in testicular
weight. Testis weight continued to increase until day 4 and declined
thereafter. On days 16 and 32, testes weights were substantially
lower than those of controls in 5 out of 16 animals, indicating the
variable response of individual animals. A decrease in the
percentage of sonication-resistant sperm heads per testis occurred
at 8 h (4 out of 8 rats affected), but this decrease was not
significant until 24 h, when a mean decrease of 19% was observed.
Maximum decreases in total sperm head counts per testis occurred on
day 8, after which some recovery was apparent. Epididymal weights
were increased on day 4. The percentage of morphologi cally normal
sperm in the cauda epididymus was less on day 4. By day 8, many
spermatozoa heads were separated from their flagella and 10% of the
heads were misshapen. Numerous sloughed round germ cells and
cytoplasmic testicular debris were also evident. No effect on the
percentage of motile sperm was seen at 2, 4 and 8 h or at 1 and 4
days post-treatment. Sperm motility was significantly decreased on
days 8 and 16. Because of the clumping and degeneration of the
spermatozoa, quantitative estimates of the percentage motility could
not be determined on days 8 and 16. Similarly damaged sperm were
seen in 3 out of 8 rats on day 32, although the percentage of motile
sperm that could be measured was similar to that of controls.
In a second experiment by Nakai et al. (1992), groups of rats
(between 97 and 105 days of age) were treated with a single oral
dose of 0, 50, 100, 200, 400 or 800 mg/kg and killed on day 2 or 70
post-treatment. On day 2, a dose-dependent increase in testicular
weight was seen at dose levels 100 mg/kg or more. This was
accompanied by significant increases in mean seminiferous tubular
diameter at 400 and 800 mg/kg. At 50 mg/kg, missing immature germ
cells were noted with round spermatids from stages I and II and
elongated spermatids sloughed from stage VII epithelium. At 100
mg/kg, the disappearance of germ cells was more severe and sloughing
of elongated spermatids extended into stages XII and XIV. A detailed
description of the spermatogenic cycle may be found in Russell et
al. (1990). At doses higher than 100 mg/kg, missing germ cells
extended into all stages except stages IX-XI, while, at doses of
400-800 mg/kg, some seminifer ous epithelia were damaged so severely
that it was difficult to identify the stage. In addition, major
pathological changes were seen in the excurrent ducts of the testis.
The rete testis was swollen with sloughed germ cells indicating that
ductal blockage had occurred further down the tract. Such occlusions
were observed in the efferent ductules of animals treated with 100
mg/kg or more. On day 70, mean testis weight and mean seminiferous
tubule diameter showed a dose-dependent decrease. Histologically,
these decreases were associated with a dose-dependent increase in
seminiferous tubular atrophy. No atrophic tubules were seen in the
control rats. The atrophied tubules contained primarily Sertoli
cells and occasional spermatogonia, and were surrounded by a
thickened basement membrane. Pathological alterations were also seen
in the efferent ductules of the treated animals, 50% or more of the
ducts being occluded in rats dosed with 100 mg or more. The
occlusions were characterized as compacted luminal contents,
spermatic granulomas, mineralizations and obliteration of the
original lumen by fibrotic connective tissue (Nakai et al., 1992).
In a further study, carbendazim was administered to female
Holtzmann rats (8 rats per group) by gavage (0, 25, 50, 100, 200,
400 or 1000 mg/kg per day) during early pregnancy (days 1 to 8). A
range of maternal parameters, including the number of implantation
sites, body weight gain, uterine weight, implantation site size, and
serum ovarian and pituitary hormones, was assessed following
sacrifice at day 9. At dosages up to 400 mg/kg per day, carbendazim
had no significant effect on any of the measured parameters but a
trend towards increased resorptions was evident. The highest dosage
produced reductions in maternal body weight gain, implantation site
weight and serum LH, and an increase in serum estradiol (Cummings et
al., 1990).
Female hamsters were treated with a single gavage dose of
carbendazim to identify effects of this compound during the
perifertilization period. The exposure times were selected to
coincide with either of the two microtubule-dependent events
initiated by the ovulatory surge of LH, i.e. oocyte maturation
(first meiotic division, occurring late on vaginal proestrous) or
fertilization (second meiotic division, occurring early on vaginal
estrous). In the first experiment, 0, 250, 500, 750 or 1000 mg
carbendazim per kg (10 hamsters per group) was administered during
meiosis I. Pregnancy outcome was assessed on day 15. The percentage
of pregnant hamsters was significantly reduced at 750 and 1000
mg/kg. In those animals that became pregnant, the average number of
live pups was reduced at all dose rates. In a second experiment,
female hamsters (10 hamsters per group) were bred overnight and
administered a single dose of 0 or 1000 mg/kg during meiosis II on
the morning following breeding. The percentage of pregnant hamsters
was unaffected, but the average number of live pups (measured at 15
days) was reduced. These results show that carbendazim
administration by gavage at microtubule-dependent meiotic events can
result in early pregnancy loss in hamsters (Perreault et al., 1992).
In a separate study, pseudopregnant rats (induced by
stimulation of the uterine cervix with a small brass rod on
proestrous and estrous) were administered 0 or 400 mg carbendazim/kg
per day during days 1-8. On day 4 of pseudopregnancy, a uterine
decidual cell response was induced and the females were killed on
day 9. The decidual cell response, evaluated as a measure of uterine
competency, was significantly less in the treated rats than in the
controls (Cummings et al., 1990).
Groups of male mice (12 per group) were administered
carbendazim (0, 250, 500 or 1000 mg/kg per day) by gavage for 5
consecutive days. Body weights, testis weights and sperm parameters
were measured at 7, 24 and 39 days post-treatment. Body weights were
unaffected. Testis weights were reduced only in the highest-dose
group at 7 and 24 days but had recovered by 39 days. Flow cytometry
measurements of testicular cells showed that the relative
percentages of certain testicular populations (round, elongating and
elongated spermatids) at the highest-dose group were different from
the control pattern 7 and 24 day after treatment (Evenson et al.,
1987).
7.5.2 Embryotoxicity and teratogenicity
Carbendazim (95% purity) was administered by gavage to female
Holtzman rats at dose levels of 0, 100, 200, 400 or 600 mg/kg body
weight per day during days 1-8 of gestation, and the rats were then
sacrificed on days 11 or 20. Maternal toxicity was not observed at
any dose level. At day 11, the crown-rump length, head length,
number of somites and number of embryos per dam were significantly
reduced in groups receiving dosages of 200 mg/kg body weight per day
or more. Open posterior neuropores and limb anomalies were observed
more frequently at dosages equal to or greater than 200 mg/kg body
weight per day. At day 20, increased resorptions, decreased live
litter size and fetal body weight and delayed ossification was
observed at all dosages. Skeletal malformations at the high dosage
levels were attributed to carbendazim exposure. The authors noted
that developmental alterations occurred at stages after the
termination of dosing, suggesting that either the anomalies
represent delays in development or the embryonic cells are
vulnerable at earlier stages than was previously thought (Cummings
et al., 1992).
Carbendazim, in a 0.5% aqueous suspension of methyl cellulose,
was administered by gavage to Crl:CDBR rats (25/dose group) on days
7-16 of gestation at daily doses of 0, 5, 10, 20 or 90 mg/kg per
day. Maternal toxicity was seen only at the highest dosage in the
form of depressed weight gains during the dosing periods and prior
to sacrifice on day 22. Mean values for liver weight and
liver-to-body weight ratio were increased. Decreased pregnancy rate
was observed at the highest dosage. An increase in the incidence of
early resorptions per dam, decreased litter size, and the total
resorption of three litters occurred at the highest dosage, only the
reduction in females per litter being significant. Significant
reductions in mean fetal weight were observed at both 20 and 90
mg/kg per day. A significant increase in the incidence of fetal
malformations was also seen at 90 mg/kg per day. The malformations
consisted primarily of hydrocephaly, microphthalmia, anophthalmia,
malformed scapulae and axial skeletal malformations (vertebral, rib
and sternebral fusions, exencephaly, hemivertebrae and rib
hyperplasia) (Alverez, 1987). The no-observed-effect levels (NOEL)
for the dam and fetus were 20 and 10 mg/kg per day, respectively.
In a further study, Delatour & Besse (1990) reported
embryotoxic effects in eleven Sprague-Dawley rats administered 19.1
mg carbendazim/kg per day by gavage from days 8 to 15 of gestation
and sacrificed at day 21. The authors reported statistically
significant (P < 0.05) increases in dead fetuses and fetal skeletal
and external malformations, and a statistically significant decrease
in fetal weight in the treated group. Since only one dose was used,
it was not possible to determine a NOEL.
The effects of carbendazim on fetal survival and development
were studied by Janardhan et al. (1984) in rats and rabbits. Female
Wistar-rats (8-10 per group) were given carbendazim (98% purity) by
gavage (0, 20, 40 and 80 mg/kg per day) on days 6 to 15 of
pregnancy. Half of each group of animals was killed on day 21 of
gestation and half was allowed to deliver normally. Female albino
rabbits were given 0, 40, 80 and 160 mg carbendazim/kg per day by
gavage on days 6 to 18 of pregnancy and were sacrificed on day 31.
All sacrificed animals were scored for live and dead fetuses and for
resorptions; live fetuses were killed and examined for
abnormalities. After normal deliveries, neonatal deaths and
survivors were counted; survivors were weighed and examined for
gross abnormalities. In rats sacrificed on day 21, dead and resorbed
fetuses accounted for 29% of conceptuses in controls, 48% at a
dosage of 20 mg/kg per day, and 64 to 73% at dosages of 40 and 80
mg/kg per day. In rabbits, dead and resorbed fetuses formed 0% of
total conceptuses in controls and 15, 21.7 and 33.3% at 40, 80 and
160 mg/kg per day, respectively. There were no differences among the
various groups of rats or rabbits with respect to mean weight of
live fetuses, and there were no malformations. In rats giving birth,
the average number of live pups per litter was close to 8 in
controls, 6 at 20 mg/kg per day, and about 5 at 40 and 80 mg/kg per
day. Mean fetal weight was increased by about 13% over controls at
the two highest dosages. There were no still births, neonatal deaths
or gross abnormalities, but mortality at 21 days postpartum was 3.0
to 3.5 times greater at the two highest-doses compared to control
(Janardhan et al., 1984).
Suspensions of carbendazim (in aqueous 0.5% carboxymethyl-
cellulose) were administered by gavage on days 7 to 19 of presumed
gestation to artificially inseminated New Zealand white rabbits (20
rabbits per group). Dose levels were 0, 10, 20 and 125 mg/kg per day
(based on active ingredient) in a volume of 5 ml/kg. The
administration of 125 mg/kg per day inhibited average maternal
weight until day 16 of gestation. There was a slightly decreased
implantation rate at 20 and 125 mg/kg per day and an increased
incidence of resorption at 125 mg/kg per day. These effects resulted
in a decreased live litter size at these two dosages. At 125 mg/kg
per day, litters showed decreased fetal body weight, but the effect
was not statistically significant. The average percentage of
malformed fetuses per litter was significantly increased at 125
mg/kg per day. Compound-related malformations at the highest dosage
consisted of malformed cervical vertebrae and interrelated
malformation of the ribs and proximate thoracic vertebrae (Christian
et al., 1985). The NOEL for maternal toxicity was 20 mg/kg per day
and the NOEL for developmental toxicity was 10 mg/kg per day.
Groups of ChR-CD rats (27-28 pregnant rats per group) were
administered carbendazim (53% a.i.) in their diet at levels of 0,
100, 500, 2500, 5000, 7500 and 10 000 mg/kg from day 6 to day 15 of
gestation. Average doses were equivalent to 0, 8.9, 45.9, 218.4,
431.6, 625.5 and 746.9 mg/kg body weight per day, respectively. On
day 20 of gestation, all pregnant animals were sacrificed and
fetuses delivered by Caesarean section. There was no mortality, no
adverse effect on body weight or clinical signs of toxicity. Food
intake was reduced at the highest-dose level during the period the
test diet was administered, but returned to control levels from days
16 to 20. The number of implantation sites, resorption sites and
live/dead fetuses were not adversely affected by carbendazim
(Sherman, 1970). After evaluating this study, the 1983 JMPR
concluded that there were no external or internal abnormalities
reported that could be considered compound-related. However, no
individual litter data were presented (FAO/WHO, 1985b).
In a further study (Koeter, 1975a), pregnant Wistar-SPF rats
(18-22 per group) were administered carbendazim in the diet at
dosage levels of 0, 600, 2000 and 6000 mg/kg from days 6 to 15 of
gestation. No individual animal or litter data were reported, and
variations in ossification and other skeletal abnormalities were
presented as percentages. Therefore, the teratogenic or fetotoxic
potential of carbendazim to pregnant Wistar-SPF rats cannot be
determined from the results and data presented (FAO/WHO, 1985b).
Groups of pregnant New Zealand albino rabbits (3-11 per group)
were administered carbendazim in the diet at dosage levels of 0,
600, 2000 and 6000 mg/kg from day 6 to 18 of gestation (Koeter,
1975b). There was a significant increase in the number of
supernumery ribs (bilateral) and skull bones in the highest-dose
group and ossification was significantly delayed or absent in these
fetuses. However, there were no individual animal or litter data,
variations in ossification were presented as percentages, and
visceral anomalies were evaluated in only two of the four groups.
The teratogenic potential of carbendazim to pregnant New Zealand
albino rabbits, therefore, cannot be ascertained from the results
and data presented (FAO/WHO, 1985b).
7.6 Mutagenicity and related end-points
Numerous studies have been conducted to assess the mutagenic
potential of carbendazim. Many of the results are conflicting and
many of the study reports do not provide sufficient detail to
evaluate the reasons for the conflicting data. Prior to the
mid-1980s, industrially produced carbendazim contained phenazine
impurities. Therefore, different degrees of purity of carbendazim
might account for some of the discrepancies. Table 9 presents those
studies that reported sufficient experimental detail and data.
Carbendazim is not a heritable gene mutagen. It does not
interact with cellular DNA, induce point mutations or result in germ
cell mutations. This has been demonstrated in both mammalian and
non-mammalian systems in vitro and in vivo, and in somatic cells
as well as in germ cells. Positive results have occasionally been
obtained in gene mutation studies, but this was due to the presence
of phenazines. These contaminants are mutagenic at very low
concentrations in the Salmonella typhimurium Ames test and also in
the mouse lymphoma Y5178Y TK+/- gene mutation assay.
Concentrations of > 4 mg diaminophenazine/kg and 10 mg
aminohydroxyphenazine/kg resulted in positive Salmonella
typhimurium Ames test results. Process changes by some of the
major carbendazim manufacturers have removed the phenazines. This
contaminant is not present when other benzimidazoles such as benomyl
or thiophanate-methyl are metabolized to carbendazim. Carbendazim
does cause numerical chromosome aberrations (aneuploidy and/or
polyploidy) in experimental systems in vitro and in vivo.
Table 9. Studies on mutagenicity of carbendazim
End-points/Tests Species, strains Concentration Activation Result Reference
1. DNA damage and repair
Bacillus subtilis 20-1000 µg per without negative Shirasu et al.
disk (1977)
Salmonella typhimurium 125-2000 µg/plate NR negative Rashid & Mumma
(TA1535, TA1938) (1986)
Escherichia Coli, K-12 125-2000 µg/plate NR
E. Coli, WP2 125-2000 µg/plate NR negative
Sister chromatid exchanges CHO cells 0.13-40.0 µg/ml without negative Ivett (1984)
1.25-40.0 µg/ml with negative
Sister chromatid exchanges human lymphocytes 0.03-30.0 µg/ml with and negative Banduhn & Obe
without (1985)
Sister chromatid exchanges human lymphocytes 1-60 µg/ml with and marginal positive Pandita (1988)
(technical grade) without at 30 µg/ml and above
Unscheduled DNA synthesis male mouse hepatocytes 0.125-12.5 µg/ml Tong (1981a,b)
male rat hepatocytes 0.0125-12.5 µg/ml
Unscheduled DNA synthesis rat hepatocytes 1.04-104.0 µg/ml negative Litton (1981)
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
2. Gene mutation
(1) Bacterial gene S. typhimurium, TA1535, 4-2500 µg/plate with and negative Gericke (1977)
mutation assays TA1537, TA98, TA100 without
Overlay spot test S. typhimurium, his G46, 50-100 µg/spot weak mutagenic Fiscor et al.
TA1530, TA1950 activity at 100 µg/spot (1978)
Plate incorporation assay TA100 50-200 µg/plate NR negative Fiscor et al. (1978)
Spot, liquid culture & S. typhimurium, his 0.25-10 000 µg/ml negative Fiscor et al.
host mediated assays G46, TA1530, TA1535, (1978)
TA1950
Plate incorporation assay S. typhimurium (TA1535, 5-1000 µg/plate with and negative Shirasu et al.
TA1537, TA1538, TA100) without (1977)
E. coli, WP2 hcr 5-1000 µg/plate with and negative Shirasu et al.
without (1977)
S. typhimurium (TA98, 1-300 µg/plate with and negative Pandita (1988)
TA100) without
S. typhimurium (TA1537, 5000 µg/plate positive in TA1537 Albertini (1989)
TA1538, TA97, TA98) & TA97 (using a
pre-incubation assay),
positive in TA1538 &
TA98 only with
activation
S. typhimurium (TA1535, 100-10 000a µg per with and positve at 5000 µg/ Donovan (1982)
TA1537, TA98, TA100) plate without plate with activation
in TA98, TA1537
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
S. typhimurium (TA1535, up to 10 000b µg with and negative up to Russell (1983)
TA1537, TA98, TA100) per plate without 10 000 µg/plate
S. typhimurium positive if Arce (1984a,b)
carbendazim samples
with 4 ppm DAP or
10 ppm AHP impurities
Host mediated assay male ICR mice Total dose of mice negative Shirasu et al.
S. typhimurium his G46 1000, 4000 mg/kg (1977)
S. typhimurium (TA1535, 2-AB with and negative Donovan (1983);
TA1537, TA98, TA100) without Russell (1977)
100-10 000 µg/plate
5-HBC 200-20000 µg/plate negative
(2) Yeast & fungal A. nidulans 2.77 µM without increase in frequency Speakman & Nirenberg
mutation assays (0.53 µg/ml) of colonies resistant (1981)
to carbendazim
Cladosporium cucumerinum 0.58 µM without no effect on frequency Speakman & Nirenberg
(0.11 µg/ml) of colonies resistant (1981)
to carbendazim
Saccharomyces cerevisiae 10-40 µg/ml negative Albertini (1991)
D7
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
(3) In vitro gene mutation
assays
HGPRT CHO cells 3-628 µM with negative Waterer (1980)
3-654 µM without
forward mutation mouse L5178Y 50-250 µM with and positive with Jotz (1980)
at TK locus lymphoma cells without activation at 100 µM,
negative without
activation
(4) Insect germ cell
gene mutation
Sex-linked recessive Drosophila melanogaster 0.5 mg/ml negative Lamb & Lilly
lethals in DMSO (1980)
Germline aneuploidy D. melanogaster, FIX, 400-50 000 ppm negative Osgood et al.
ZESTE feeding to larvae or (1991)
young adult females
(5) In vivo mammalian mouse embryos treated in 200 mg/kg positive Fahrig & Seiler
gene mutation assays utero by dosing the mother (1979)
100-300 mg/kg orally
3. Chromosomal effects
(1) In vitro chromosomal S. cerevisiae 0.001-0.100 mg/ml maximum responses Whittaker et al.
effect for mitotic & meiotic (1990)
chromosome gain at 0.05
and 0.025 mg/ml
S. cerevisiae 5 µg/ml positive for Albertini (1991)
aneuploidy
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
human lymphocytes 0.1-10.0 µM with and not clastogenic, Banduhn & Obe
without but induce (1985)
micronuclei at > 10 µM
human lymphocytes 0.5 mg/ml without did not increase Lamb & Lilly
chromosome (1980)
aberration frequency
(2) In vivo mammalian
chromosomal effect
male & female Sprague- single oral dose did not increase BASF (1975);
Dawley rat bone marrow 300 mg/kg chromosome FAO/WHO (1985b)
cells aberration frequency
Chinese hamster bone oral, 1000 mg/kg did not produce Seiler (1976)
marrow cells chromosome breakage
ICR mice nucleated 2 oral dose of exhibit lagging Seiler (1976)
anaphase cells 1000 mg/kg chromosome, bridge
formation, tripolar
spindle formation, etc
Mouse bone marrow 500 mg/kg did not detect Seiler (1976)
micronuclei
Swiss albino mice 0, 200, 1000, 2000, dose dependent Pandita (1988)
4000 or 6000 mg/kg i.p. increase in
micronucleus
male Chinese hamsters 0, 100, 500 or no effect on Pandita (1988)
bone marrow 1000 mg/kg i.p. hyperploidy
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
(3) In vivo germ cell
chromosomal mutation
Mouse dominant lethal NMRI mice 500 mg/kg i.p. per did not exhibit a Hoechst (1974)
test day, 5 days dominant lethal effect
male NMRI mice 300 mg/kg gavage did not exhibit a Hoechst (1974)
per day, 5 days dominant lethal effect
a Phenazin concentration in carbendazim: 9 ppm DAP, 5 ppm AHP
b Phenazin concentration in carbendazim: < 0.6 ppm DAP, < 4 ppm AHP
DAP = diaminophenazine; AHP = aminohydroxyphenazine; i.p. = intraperitoneal; NR = not reported
7.7 Carcinogenicity
In a study by Wood (1982), groups of CD-1 mice (80 males and 80
females per group) were administered carbendazim (99% a.i.) in the
diet at dose levels of 0, 500, 1500 and 7500 mg/kg diet for two
years. The highest-dose was reduced to 3750 mg/kg after 66 weeks for
the males because of increased mortality (62 surviving controls, 32
surviving at 7500 mg/kg); females, however, received 7500 mg/kg
throughout the study period. Animals were 6-7 weeks old at the start
of this study. Mortality was compound-related in male mice. The
high-dose males were sacrificed at week 73 because of a significant
increase in mortality (23 surviving). Only nine males in the
1500-mg/kg group survived to week 104, compared with 18 surviving
control males. Females did not show this increase in mortality.
There were no dose-related effects on body weight or food
consumption at any time during the study, although terminal body
weights for the low- and mid-dose males were less than those of the
control and high-dose males. Clinical data were similar for all
treatment and control groups. Haematological measures were
unaffected.
Both absolute and relative thymus weights were significantly
decreased in females at 500 and 1500 mg/kg, but not in the high-dose
group. Absolute liver weight was increased in the high-dose females
and relative liver weight at the two highest-dose levels. The organ
weights of male mice were variable and only the kidney and thymus
weights appeared to be decreased as a result of treatment. Absolute
kidney and thymus weights were depressed in male mice at all
treatment levels, but relative kidney and thymus weights were
significantly decreased only in the high-dose males. Histological
examination revealed dose-related changes in the thymus (lymphoid
depletion) and accumulation of yellow-brown pigment in the renal
tubules for mid- and high-dose male mice.
Examination of the testes demonstrated an increase in the
frequency of sperm stasis in mid- and high-dose males, together with
increased germinal cell atrophy (bilateral only). There was no
trend, however, towards unilateral germinal cell atrophy, the
incidence in controls being greater than or equal to that of treated
males. These effects are, therefore, not considered to be
compound-related.
The examination of male mice livers revealed a significant
hepatotoxic effect at 1500 and 7500 mg/kg, demonstrated by
centrilobular hypertrophy, necrosis and swelling. There was no
increase in the frequency of hepatocellular adenomas; these occurred
with equal frequency in control and treatment groups. There was a
significant increase in hepatocellular carcinomas but only at 1500
mg/kg. However, too few males survived at the high-dose level to 17
months (510 days) to support the conclusion of no oncogenic effect
at that dose level.
The occurrence of total hepatic tumours (hepatocellular
carcinomas, hepatocellular adenomas and hepatoblastomas) is given in
Table 10. This was statistically increased (P < 0.05) for the low-,
mid-, and high-dose females. It was also statistically increased (P
< 0.05) in mid-dose males but was not evaluated in high-dose males
because of the high rate of mortality.
In conclusion, this study on CD-1 mice showed that there were
statistically significant increases (P < 0.05) in the incidence of
hepatocellular carcinoma at 1500 mg/kg for males and at all dose
levels for females, the response being dose-related (P < 0.05).
Because of high mortality in the high-dose males, a dose-response
relationship could not be determined. In addition, the high mor
tality rate in male controls further hampered the interpretation of
results. Histopathological analysis of the hepatocellular tumours in
the test animals showed no difference from controls, and the median
latent period for development of these hepatocellular carcinomas
showed no significant decrease in the test animals. No carcinogenic
effect was observed in tissues other than the liver (Wood, 1982).
In a study on SPF Swiss mice (100 males and 100 females per
group), carbendazim was administered in the diet at dosage levels of
0, 150, 300 and 1000 mg/kg diet for 80 weeks. The highest-dose was
increased to 2000 mg/kg at week 4 and to 5000 mg/kg at week 8 for
the remainder of the study. Animals were examined for behaviour and
clinical signs of toxicity, and body weight measurements were
determined throughout the study. Gross pathology investigations were
performed on all animals, liver and kidney weights were recorded,
and tissues were examined microscopically. There were no
compound-related effects on general condition, mortality or body
weight. Survival at term was 70% for males and 80% for females.
Relative liver weights for high-dose males and females were
significantly different from those of controls, but there were no
changes in kidney weights. The results are shown in Table 11. The
combined incidence of hepatocellular adenomas and hepatocellular
carcinomas increased with increasing dose levels for both males and
females. Males showed a more pronounced induction of liver tumours
and a more frequent occurrence of hepatocellular carcinomas which
were often found simultaneously with hepatocellular adenomas.
Females, on the contrary, showed only hepatocellular adenomas in
most cases (Beems et al., 1976; Mohr, 1977).
Table 10. Incidence of combined primary hepatocellular tumours and mean and median period for development
after treatment with carbendazim in CD-1 micea
Male mice Female mice
Carbendazim concentration (mg/kg diet): 0 500 1500 7500 (3750) 0 500 1500 7500
Combined incidences of hepatic tumours 13 20 23b NAc 1 9b 21b 15b
Median period for development (days) 633 697 651 NAc 732 724 697 733
Mean period for development (days) 628 671 628 NAc 732 706 689 653
Number of mice examinedd 80 80 80 80 79 78 80 78
Median survival time (weeks) 79 72 69 64 91 91 91 91
Number of mice alive at termination 18 14 9 23 22 15 13 20
a From: Wood (1982)
b Significant by Fisher's Exact Test at P < 0.05 level of probability
c NA = not applicable. This group was terminated after 516 days on test (test week 73) due to a high rate of mortality. In
view of the early termination of this test group and low incidence of hepatic tumours observed in this group, the inclusion
of its data in statistical evaluation of hepatic tumour incidence is considered inappropriate.
d Mice found dead or sacrificed in extremis prior to each group's termination were subjected to gross pathological evaluation.
Table 11. Incidence of proliferation lesions of the hepatocytes in Swiss micea
Male mice Female mice
Carbendazim concentration (mg/kg diet): 0 150 300 5000 0 150 300 5000
Number of animals examinedb 100 94 98 100 94 99 98 95
Liver nodular hyperplasiac 0 8 11 25 0 5 3 11
Hepatocellular adenoma 9 5 13 14 1 1 3 8
Hepatocellular carcinoma 1 3 4 9 1 0 0 0
a Adapted from: Mohr (1977)
b A number of animals could not be examined because of well-advanced autolysis. In addition, slides were not available for
some of the animals.
c Values for this non-neoplastic lesion included all types of cellular alteration. A statistical analysis was not conducted.
In a study on HOE NMRKf (SPF 71) mice (100-120 males and
females per group), carbendazim was administered in the diet for 96
weeks at dosage levels of 0, 50, 150, 300 and 1000 mg/kg diet. The
highest-dose was increased to 2000 mg/kg at week 4 and to 5000 mg/kg
at week 8 for the remainder of the study. Animals were examined for
behaviour and general condition, as well as for body weight,
food/water consumption and mortality. Gross necropsies were
performed on all animals, liver and lung weights were recorded, and
all organs and tissues were examined microscopically. An interim
sacrifice was made at 18 months of 20 males and 20 females from the
control group and the highest-dose group.
There were no compound-related effects on behaviour, body
weight gain, food/water consumption or mortality. At 22 months there
was 24-31% mortality in male mice and 37-52% mortality in females in
all groups of mice. As there was no difference between the treated
and control groups, it was concluded mortality was not influenced by
the feeding of carbendazim. The mean daily consumption of
carbendazim in mg/kg body weight for males and females,
respectively, was 5.8 and 7.1 at 50 mg/kg diet, 17.1 and 21.2 at 150
mg/kg diet, 34.4 and 41.9 at 300 mg/kg diet, and 548.4 and 682.3 at
5000 mg/kg diet.
Examination of lung and liver weights at 18 and 22 months
demonstrated an increase in absolute and relative liver weights in
both male and female mice at 5000 mg/kg diet. Macroscopic and
microscopic examination of 20 male and 20 female animals killed
after 18 months of receiving 5000 mg carbendazim/kg revealed
compound-related effects on the liver. All animals demonstrated
centrilobular hypertrophy, single cell necrosis, mitotic cells and
pigmented Kupffer cells. The tissues of the remaining 100 males and
100 females exposed to 5000 mg/kg were evaluated at 22 months. There
was marked liver cell hypertrophy (greater than in animals treated
for 18 months only), clear cell foci, mitosis, inclusion bodies in
enlarged cell nuclei, multiple cell necrosis, and greenish yellow
pigment in Kupffer cells.
Neoplastic nodules (adenomas), carcinomas, fibrosarcomas and
other tumorigenic responses in the liver were equally distributed
among all groups (Tables 12 and 13). Although haemangiomas of the
liver (Table 14) were found in all treated groups (but not in
controls), no dose-related response was evident. Lung adenomatosis
was equally distributed among all groups. There was no effect of
carbendazim on the incidence or time of onset of tumours, and the
total number of benign and malignant tumours was comparable among
the various groups of mice. Thus, there was no evidence of a
carcinogenic effect from carbendazim when administered in the diet
to mice at doses up to and including 5000 mg/kg diet for 22 months
(Donaubauer et al., 1982).
Table 12. Incidence of tumours in HOE NMRKf mice after 18 months of
exposurea
Mice treated with
Control 5000 mg/kg diet
male female male female
Lung
adenoma 3 5 4 -
Uterus
stromal sarcoma 1 -
Ovary
tubular adenoma 1 -
granulosa cell tumour 1 -
luteoma and tubular adenoma 1 -
Skin and subcutaneous tissue
round cell sarcoma - - - 1
adenocarcinoma - - 1 -
Lymph nodes
lymphosarcoma - - - 1
3 9 5 2
a Adapted from Donaubauer et al. (1982)
7.8 Neurotoxicity
Carbendazim (500, 2500 and 5000 mg/kg) has been evaluated for
neurotoxic potential in white Leghorn hens (10 per group). Controls
consisted of the vehicle (corn oil) and the neurotoxin tri- o-tolyl
phosphate (TOTP). The hens were observed daily for mortality and
clinical neurotoxicity for four weeks. Neurotoxic signs, consisting
of leg weakness, ataxia and/or "goose-stepping" gait, were observed
in hens treated with TOTP. Less severe and reversible signs,
consisting of slight leg weakness and ataxia, were observed in hens
treated with 5000 mg carbendazim/kg, but no neurotoxic signs were
observed for those treated with 500 or 2500 mg/kg. Microscopic
examination indicated that there was no axonal degeneration or
demyelination in carbendazim-treated animals (Goldenthal, 1978).
Table 13. Possible preneoplastic changes and primary neoplasms in the livers and lungs of HOE NMRKf mice
after 22 months of exposurea
Carbendazim concentration: 0 50 150 300 5000
(mg/kg diet) male female male female male female male female male female
Liver
Neoplastic nodules
(adenomas) 3 2 1 1
Clear cell foci 3 4
Basophilic foci 1 1
Haemangiomas (liver) 2 3 2 2 1
Lungs
Adenomatosis 29 10 31 8 30 10 25 9 22 12
Adenocarcinomas 1 1
Keratinizing squamous
cell carcinomas 1
Cavernous haemangioma 1
32 10 35 11 34 11 27 12 27 17
a Adapted from Donaubauer et al. (1982)
Table 14. Haemangiomas of the liver in HOE NMRKf mice after
22 months of exposurea
Dose (mg/kg diet) Male Female
0 0 0
50 2 0
150 3 0
300 2 2
5000 0 1
a Adapted from Donaubauer et al. (1982)
7.9 Toxicity of contaminants
When manufactured under certain conditions, carbendazim
contains a class of contaminants called phenazines. These
contaminants are mutagenic at very low concentrations in the
Salmonella typhimurium Ames test and also in the mouse lymphoma
Y5178Y TK+/- gene mutation assay. Concentrations of > 4 ppm
diaminophenazine or 10 ppm aminohydroxyphenazine yielded positive
results. Process changes by some of carbendazim manufacturers have
now removed the phenazines.
7.10 Mechanisms of toxicity - mode of action
Biochemical studies on the mechanism of action of benzimidazole
compounds have shown that their biological effects are caused by
interactions with cell microtubules (Davidse & Flach, 1977). These
cellular structures are present in all eukaryotic cells and are
involved in several vital functions, such as intracellular
transports and cell division. Benzimidazole compounds have been used
as anticancer drugs and as anthelminthic drugs in animals and humans
because they act as spindle poisons by interfering with the
formation and/or functioning of microtubules. However, eukaryotes
are known to be unequally sensitive to each benzimidazol compound,
which explains the use of these compounds in helminthiases.
Selective toxicity of benomyl and carbendazim for fungi has been
explained by comparing their binding to fungal and mammalian
tubulin. The different sensitivity of several fungi has also been
explained by the different affinity of benomyl and carbendazim for
fungal tubulin.
Benomyl has been found to bind to fungal tubulin but not to
porcine brain tubulin, indicating that mammalian tubulin has no, or
at least low, affinity for benomyl (Davidse & Flach, 1977). This is
in agreement with the observation that benomyl at concentrations
that are lethal for sensitive fungi does not interact with in vitro
microtubule assembly in these brain extracts. In vitro ID50
values for several mycelial extracts of various fungal species
sensitive to benomyl were all below 5 µmol/litre (Davidse & Flach,
1977). In vitro rat brain tubulin polymerization was inhibited to
about 20% at benomyl or carbendazim concentrations of 25 µmol/litre
(De Brabander et al., 1976b). For comparison, a standard antitubulin
drug in humans such as vincristine inhibited 50% tubulin assembly at
0.1 µmol/litre in the same experiment. The assembly of sheep and
calf brain microtubule was also found to be unaffected by
carbendazim concentrations higher than 100 µmol/litre (Ireland et
al., 1979).
Mitotic arrest by benzimidazole and six analogues at metaphase
was evaluated in human lymphocyte cultures. Structure-activity
relationships indicate that antimitotic activity is related to C6
substitution of the benzimidazole moiety (Holden et al., 1980). In
this study, however, benomyl and carbendazim were not tested. The
question of whether all C6 unsubstituted benzimidazoles, such as
benomyl and carbendazim, have no effect on mitosis of human
lymphocytes in cell cultures is therefore unresolved.
A link between the effects of benomyl and carbendazim on
tubulin and their teratogenic effects has been postulated (Ellis et
al., 1987, 1988).
8. EFFECTS ON HUMANS
8.1 General population exposure
No references to carbendazim poisoning have been documented in
the literature. Recent data to estimate dietary exposure of the
benzimidazole benomyl, based on food consumption patterns within the
USA and elsewhere, indicate exposure below the NOELs from animal
toxicity tests. Further information is given in section 5.2.1 of
Environmental Health Criteria 148: Benomyl (WHO, 1993).
8.2 Occupational exposure
Selected blood profiles from 50 factory workers involved in the
manufacture of benomyl and carbendazim were compared to those of a
control group of 48 workers who were not exposed to these two
fungicides. White blood cell count, red blood cell count, and
haemoglobin and haematocrit values were comparable among the two
groups. There were no quantitative estimates of exposure given for
the factory workers. No female employees were included in the
control group (Everhart, 1979; FAO/WHO, 1985a).
A study was performed to determine whether exposure to benomyl
and carbendazim had an adverse effect on the fertility of 298 male
manufacturing workers exposed to benomyl between 1970 and 1977. The
workers ranged from 19 to 64 years of age (79% were between 20 and
39, and 78% of the spouses were similarly aged between 20 and 39
years). Exposure duration ranged from less than one month to 95
months, and more than 51% of the workers were potentially exposed
from 1 to 5 months. The birth rates of exposed workers' spouses were
compared with those of four comparison populations from the same
county, state, region and country (USA). There was no reduction in
fertility as shown by the birth rates for the study population,
which were generally higher than the comparison populations.
Spermatogenesis among workers was not examined (Gooch, 1978;
FAO/WHO, 1985a).
9. EFFECTS ON OTHER ORGANISMS IN THE LABORATORY AND FIELD
9.1 Microorganisms
Studies on soil effects have concentrated on benomyl. However,
carbendazim, the breakdown product of benomyl, would be expected to
produce comparable results.
Soil respiration has been found to be little influenced by
benomyl at concentrations below 10 mg/kg, which is the maximum soil
concentration expected after use at recommended application rates
(Hofer et al., 1971; van Fassen, 1974; Peeples, 1974; Weeks &
Hedrick, 1975).
A study on the influence of benomyl on soil nitrogen
mineralization showed that the release of ammonia was not decreased
by benomyl, whereas the influence of the fungicide on nitrification
varied from a stimulation (van Fassen, 1974), through no effect
(Mazur & Hughes, 1975), to a decreased nitrification (Hofer et al.,
1971; Wainwright & Pugh, 1974). The differences may be related to
the soil composition since Hofer et al. (1971) found a greater
effect in sandy than in organic soil.
Benomyl, in combination with eleven other pesticides that were
used in an orchard spray programme, had only a minimal and
short-term effect on respiration, ammonification and nitrification
at concentrations expected after recommended use of benomyl over a
spraying season. Ten times the recommended application rates had a
pronounced effect on both respiration and nitrification, which
lasted for more than 4 weeks (Helweg, 1985).
The influence of benomyl and carbendazim on soil microbial
activity was studied in Sweden following repeated annual
applications during autumn to winter cereals for a period of 3 to 5
years. The effects of the fungicides on straw decomposition, balance
of straw fungal flora and nitrogen mineralization in the soil were
investigated in field and laboratory experiments. The decomposition
of straw in the field was not affected in clay soils by annual
applications of up to 2 kg/ha. In sandy soils, rates of up to 0.5
kg/ha had no effect, but in one case at 2 kg/ha the initial stages
of straw decomposition were slightly inhibited. All doses tested in
both clay and sandy soils caused changes in the composition of the
straw fungal flora (Torstensson & Wessen, 1984).
Tests were conducted to determine the effects of the fungicide
carbendazim (Bavistin), the herbicide fluometuron (Cotoran) and the
insecticide curacron on Egyptian soil fungi when applied at the
recommended field rates and at 4 and 8 times these rates.
Carbendazim produced a significant inhibition of the total count of
fungi at all three rates after 5 and 40 days and at the higher
application rates after 80 days. The response of Aspergillus sp.
alone to this fungicide reflected the effects on the total fungal
count (Abdel-Fattah et al., 1982).
An application of carbendazim (5 mg formulation/kg soil) and
Calixin (1.5 mg formulation/kg soil) significantly enhanced the
microbial activity, as well as the availability of ammonium nitrogen
and phosphate in the soil, while nitrite, nitrate and available
potassium were found to decrease. During incubation, CO2 evolution
decreased during the first 21 days, increased up to 42 days, and,
thereafter, decreased with both treatments (Khan et al., 1987).
9.2 Aquatic organisms
The effect of carbendazim was monitored using the green alga
Selenastrum capricornutum in an OECD guideline test (201). The
EC50 (based on total growth) at 72 h was 1.3 mg/litre and at 120 h
was 1.6 mg/litre. The no-observed-effect concentration (NOEC) was
0.5 mg/litre. To study whether carbendazim was algistatic or
algicidal, organisms were recultured at the end of the initial 120 h
of incubation. Regrowth occurred in the control but not in the test
cultures (8.0 mg/litre) after a period of 9 days. Carbendazim was,
therefore, considered to be algicidal (Douglas & Handley, 1987). In
another study using Chlorella pyrenoidosa, the 48-h EC50 for
growth inhibition was calculated to be 0.54 mg/litre (Canton, 1976).
The acute toxicity of carbendazim to a variety of aquatic
organisms is summarized in Table 15. For 96-h tests, LC50 values
ranged from 0.007 mg/litre for channel catfish ( Ictalurus punctatus
yolk-sac fry) to 5.5 mg/litre for bluegill sunfish (Palawski &
Knowles, 1986).
In a static renewal 21-day test using Daphnia magna, onset of
reproduction was the most sensitive indicator, being significantly
delayed at 0.025 mg carbendazim/litre (measured concentration).
Other parameters (number of days reproducing, total number of young,
and young produced per day) were affected significantly at 0.05
mg/litre. The NOEC was 0.013 mg/litre (measured concentration)
(Hutton et al., 1986).
Gillet & Roubaud (1983) showed that the toxicity of carbendazim
to carp ( Cyprinus carpio) was greater for the fertilization and
early development stages than for the adult stages. They found the
30-min LC100 during the fertilization stage to be < 5 mg/litre at
pH 9 and < 2.5 mg/litre at pH 7. The rainbow trout ( Oncorhynchus
mykiss) was more sensitive, with an LC100 (30 min, pH 9) of 0.5
mg/litre and 71% survival, compared to controls, at 0.05 mg/litre.
Table 15. Toxicity of carbendazim to aquatic organisms
Organism Size/a Stat/ Temperature Hardnessb pH Duration LC50 Reference
age flow (°C) (mg/litre) (h) (mg/litre)
Freshwater
Water flea < 24 h stat 20 179 7.5 48 0.087 Hutton (1988)
(Daphnia magna) < 24 h stat 20 48 0.46 Canton (1976)
Rainbow trout 3 months stat 15 48 1.8 Canton (1976)
(Oncorhynchus mykiss) 0.8 g stat 13 7.5 96 0.36 Heinemann (1971)
yolk-sac fry stat 10 40-48 7.4 96 0.145 Palawski & Knowles (1986)
swimup fry stat 10 40-48 7.4 96 0.32 Palawski & Knowles (1986)
0.2 g stat 10 40-48 7.4 96 0.37 Palawski & Knowles (1986)
1.2 g stat 10 40-48 7.4 96 0.87 Palawski & Knowles (1986)
stat 12 40-48 7.4 96 > 1.8 Palawski & Knowles (1986)
stat 17 40-48 7.4 96 0.87 Palawski & Knowles (1986)
stat 22 40-48 7.4 96 0.1 Palawski & Knowles (1986)
stat 10 40-48 6.5 96 0.64 Palawski & Knowles (1986)
stat 10 40-48 7.5 96 0.41 Palawski & Knowles (1986)
stat 10 40-48 8.5 96 0.34 Palawski & Knowles (1986)
stat 10 40 7.4 96 0.78 Palawski & Knowles (1986)
stat 10 320 7.4 96 0.88 Palawski & Knowles (1986)
Bluegill sunfish 0.95 g stat 22-23 67 7.0 96 5.5 Wetzel & Hutton (1984)
(Lepomis macrochirus) 0.2 g stat 22 40-48 7.4 96 > 3.2 Palawski & Knowles (1986)
Channel catfish yolk-sac fry stat 22 40-48 7.4 96 0.007 Palawski & Knowles (1986)
(Ictalurus punctatus) swimup fry stat 22 40-48 7.4 96 0.012 Palawski & Knowles (1986)
0.2 g stat 22 40-48 7.4 96 0.01 Palawski & Knowles (1986)
1.2 g stat 22 40-48 7.4 96 0.019 Palawski & Knowles (1986)
stat 12 40-48 7.4 96 > 0.56 Palawski & Knowles (1986)
stat 17 40-48 7.4 96 0.14 Palawski & Knowles (1986)
stat 22 40-48 7.4 96 0.032 Palawski & Knowles (1986)
stat 22 40-48 6.5 96 0.023 Palawski & Knowles (1986)
Table 15 (contd).
Organism Size/a Stat/ Temperature Hardnessb pH Duration LC50 Reference
age flow (°C) (mg/litre) (h) (mg/litre)
stat 22 40-48 7.5 96 0.014 Palawski & Knowles (1986)
stat 22 40-48 8.5 96 0.023 Palawski & Knowles (1986)
stat 22 40 7.4 96 0.018 Palawski & Knowles (1986)
stat 22 320 7.4 96 0.024 Palawski & Knowles (1986)
Toad tadpole stat 25 48 3.5 Nishiuchi & Yoshida (1974)
(Bufo bufo japonicus)
Marine and Estuarine
Eastern oyster 25-50 mm flow 18-20 96 > 1.145c Boeri (1988a)
(Crassostrea virginica)
Mysid shrimp stat 23 96 0.098 Boeri (1988c)
(Mysidopsis bahia)
Sheepshead minnow 0.14 g stat 22 96 > 1.158 Boeri (1988b)
(Cyprinodon variegatus)
a stat = static conditons (water unchanged for duration of test); flow = flow-through conditions (carbendazim concentration
in water continuously maintained)
b hardness given as mg CaCO3/litre
c EC50 based on rate of shell deposition
Bluegill sunfish were exposed to benomyl, carbendazim and 2-AB
at concentrations of 0.05 mg/litre (measured concentrations 0.01 to
0.04 mg/litre) and 5.0 mg/litre (measured concentration 2 to 5
mg/litre). No residue was found in the tissues of fish exposed to
low levels of these compounds, but detectable residues were found in
the tissues of fish exposed to the high levels. However, there was
no build-up or bioconcentration with time (DuPont, 1972).
9.3 Terrestrial organisms
Van Gestel et al. (1992) exposed red earthworms ( Eisenia
andrei) to carbendazim, added as an aqueous solution of the
formulation Derosal, to artificial soil. The final concentrations in
soil were 0, 1, 3.2, 10, 32 and 100 mg formulation/kg dry soil,
corresponding to 0, 0.6, 1.92, 6.0, 19.2 and 60 mg carbendazim/kg.
The worms had been acclimatised for 1 week in the artificial soil,
which contained 8 g cow dung per kg soil as food for the worms. The
soil pH (7.3) was higher than optimum for this species and resulted
in a lower cocoon production in this test than expected. At the two
highest concentrations of 19.2 and 60 mg a.i./kg, all worms died
over the 3 week experimental period; an LC50 of 5.7 (4.7-6.9) mg
a.i./kg was calculated. Growth was significantly reduced at 6.0
mg/kg soil, and reproduction (measured as cocoon production, number
of fertile cocoons and numbers of juveniles) was significantly
reduced at > 1.92 mg a.i./kg soil. The EC50 for cocoon
production was calculated to be 2.9 (2.2-3.8) mg a.i./kg dry soil.
The response curve for reproduction and growth effects of
carbendazim was steep. Vonk et al. (1986) had reported comparable
results for a second Eisenia species, E. foetida, in a different
artifical soil; the LC50 was 9.3 mg/kg soil and the NOEC for
cocoon production 2.0 mg/kg soil.
Carbendazim (99.3% purity) was evaluated for acute contact
toxicity after thoracic application in honey-bees ( Apis mellifera).
Each treatment level consisted of four replicates of ten bees each.
Forty bees served as positive control (using carbaryl) and forty as
negative control. No deaths occurred after the application of 50 µg
carbendazim/bee, the highest rate tested. Carbendazim is, therefore,
classified as "relatively non-toxic" to the honey-bee (Meade, 1984).
The LD50 for bobwhite quail, with an observation period of 14
days following the single oral dose, was > 2250 mg/kg body weight
(Beavers, 1985). For mallard ducks the dietary 5-day LC50 was >
10 000 mg carbendazim/kg diet (Fink, 1975).
9.4 Population and ecosystem effects
Van Gestel (1992) has summarized reports of toxicity of
carbendazim on earthworms from field studies based on different soil
types, application rates and crops (Table 16). Estimated soil
concentrations in this table are based on application rates; it has
been assumed that there is no mobility of the compound beyond the
top 2.5 cm of soil and homogeneous distribution of carbendazim in
this layer. For orchard application, it was further assumed that 50%
of the applied active ingredient reached the soil. Reported effects
include reduced numbers, reduced activity (as reduced removal of
leaf litter) and reduced reproduction of worms. Application rates
were within the recommended rates for carbendazim as a fungicide on
these crops.
Table 16. Summary of earthworm toxicity data on carbendazim in field studies
Crop/soil type Dosage Estimated soil Time Effect Reference
(kg/ha) concentration (days)
(mg/kg)
Grass 0.3 0.9 12 11% reduction in number Ammon (1985)
Grass 0.56 1.6 49 72% reduction in cast production Keogh & Whitehead
42% more litter left (1975)
Winter wheat/clay 0.15 0.4 275 14% reduction in juveniles Lofs-Holmin (1981)
50% increase in adult
Allolobophora caliginosa
25% increase in juveniles
25% reduction in adult
A. chlorotica Lofs-Holmin (1981)
Apples/loam 0.375a 1.6 245 reduction in litter removal Cook & Swait (1975)
a Three separate applications of carbendazim
10. EVALUATION OF HUMAN HEALTH RISKS AND EFFECTS ON THE ENVIRONMENT
Benomyl and carbendazim are two different fungicides in their
own right. However, carbendazim is also the main metabolite of
benomyl in mammals and the degradation product of benomyl in the
environment. Butyl isocyanate is the chemical moiety removed from
benomyl when carbendazim is formed. Given the similar toxicities
caused by benomyl and carbendazim and the different toxicological
profile of butyl isocyanate, the two fungicides are evaluated
together in this monograph.
10.1 Evaluation of human health risks
The two primary routes of exposure to humans are through diet
and through manufacturing or use of the product.
There is limited information on actual dietary exposure to
benomyl and carbendazim. Dietary exposure has been estimated in the
Netherlands and the USA. In the USA, exposure based on dietary
habits, measured residue levels, and the percentage of crop treated
has been calculated for various subgroups of population. These
calculations indicate that the estimated benomyl exposure is
0.144-1.479 µg/kg per day (section 5.2.1 of Environmental Health
Criteria 148: Benomyl (WHO, 1993)). In the Netherlands, the mean
dietary intake was estimated to be 0.83 µg/kg per day (0.05 mg/day
per person). These levels of exposure are below the recommended ADI
of 0.01 (carbendazim) and 0.02 (benomyl) mg/kg body weight.
The average air levels of benomyl and carbendazim have been
determined in a manufacturing facility (section 5.3) and found to be
less than 0.2 and 0.3 mg/m3, respectively. Both values are below
the Threshold Limit Value of 5-10 mg benomyl/m3 established by a
number of governmental agencies.
In one study, potential respiratory and dermal exposure to
benomyl wettable powder formulation was determined under several
agricultural use situations (section 5.3). The highest rates of
exposure occurred in situations of mixing and loading in preparation
for aerial application; dermal and respiratory exposures were,
respectively, 26 and 0.08 mg/person per cycle. Home users and
agricultural workers re-entering treated fields were estimated to be
exposed to about 1 mg/person per cycle and 5.9 mg/person per hour,
principally through the dermal route.
Because of the low mammalian toxicity, acute benomyl or
carbendazim poisonings are unlikely to occur under conditions of
normal use.
Several studies of agricultural workers (section 8.2 of
Environmental Health Criteria 148: Benomyl (WHO, 1993)) have shown
some cases of contact dermatitis after exposure to benomyl. These
effects can be significantly reduced or eliminated by wearing
long-sleeved shirts, long trousers and gloves.
There is little information on health effects in humans as a
consequence of exposure to either benomyl or carbendazim. Two
studies have been conducted on factory workers involved in the
manufacture of benomyl (section 8.2). In one study, haematological
profiles from 50 factory workers involved in the manufacture of
benomyl were comparable to those from a control group of 48 workers.
A second study found no decrease in the birth rate of the wives of
298 factory workers exposed to benomyl.
Extensive studies of various species of laboratory animals show
reproductive, developmental, mutagenic and carcinogenic effects
associated with both benomyl and carbendazim. The effects observed
on rat fetuses were microphthalmia, hydrocephaly and encephaloceles.
The no-observed-effect levels (NOEL) of benomyl for developmental
toxicity are equal to or greater than 10 mg/kg body weight per day,
depending upon the species and route of administration. Similarly,
the NOEL of benomyl for reproductive effects in the male rat appears
to be 15 mg/kg body weight per day after gavage dosing. In feeding
studies with both benomyl and carbendazim, the NOEL appears to be
500 mg/kg diet (equivalent to 25 mg/kg body weight per day).
However, one benomyl feeding study reported a NOEL of less than 1
mg/kg diet (0.05 mg/kg body weight per day) for male reproductive
effects. The reason for the discrepancy between the NOEL in this
latter study and other investigations is unknown.
The only consistent genotoxic effect noted in animal studies is
the induction of numerical chromosomal aberrations. These effects
are consistent with the interaction of benomyl and carbendazim with
microtubule formation.
Rat carcinogenicity studies did not show any carcinogenic
effect for either compound. Benomyl and carbendazim induce
hepatocellular tumours in CD-1 and SPF Swiss mice but not in NMRKf
mice. This finding in mice is not considered to be a result of a
direct genotoxic action. Rather, it appears to be associated with
liver toxicity in strains of mice that are highly susceptible to
tumour formation at this site.
Benomyl and carbendazim are spindle poisons. Effects on target
cells are consequences of binding to microtubules, giving toxicities
similar to those of other spindle poisons such as colchicine and
vincristine. Benzimidazole compounds in general and benomyl and
carbendazim in particular have selective effects on the microtubules
of different eukaryotes. Reasons for this selectivity include the
binding capability to different tubulins and pharmacokinetic
differences across species. In vitro concentrations of benomyl
used to kill sensitive fungi were found to be ineffective in
disturbing mammalian microtubular functions. These studies on the
mechanism of action of benomyl and carbendazim indicate a selective
effect of these compounds for target species.
In summary, the LD50, as determined in a number of test
species, for benomyl ranges from > 2000 to > 12 000 mg/kg and for
carbendazim from > 2000 to > 15 000 mg/kg. There are no known
reports of human poisoning for either compound. This, coupled with
the low estimated environmental levels of both compounds, would
suggest that the possibility of acute poisoning by benomyl or
carbendazim is very remote. Similarly, the data available on test
species make it unlikely that either benomyl or carbendazim is
carcinogenic for humans. The NOELs for both reproductive and
teratogenic effects of benomyl and carbendazim (i.e. 10-15 mg/kg) do
raise a possibility that an accidental ingestion of either fungicide
could adversely alter reproductive outcome in humans, but the
likelihood that such poisoning would occur is remote. The
selectivity of these two benzimidazole compounds for the tubulin of
the target species (fungi) and their relative ineffectiveness to
disturb mammalian microtubule function further reduce the
possibility of their having toxic effects in humans.
10.2 Evaluation of effects on the environment
Benomyl is rapidly converted to carbendazim in various
environmental compartments, the half-lives being 2 and 19 h in water
and soil, respectively. Therefore, data from studies on both benomyl
and carbendazim are relevant for the evaluation of environmental
effects.
Carbendazim persists on leaf surfaces and in leaf litter. In
soil the half-life is between 3 and 12 months, and the compound may
be detected for up to 3 years. However, in many cases, major
residues will be lost within a single season. Residues of carben
dazim and its metabolites are strongly bound or incorporated into
soil organic matter. The strong adsorption (Koc = approximately
2000) of carbendazim to soil and sediment particles reduces its
bioavailability to terrestrial and aquatic organisms. Similarly, the
mobility of carbendazim in soil is limited, and it is not expected
to leach to ground water.
Benomyl and carbendazim are highly toxic to some aquatic
organisms in laboratory tests, the most sensitive species being the
channel catfish with a 96-h LC50 for yolk-sac fry of 0.006 mg
benomyl/litre. However, this toxicity is unlikely to be manifest in
the environment for most aquatic organisms because of the low
bioavailability in surface waters. The exposure of sediment-living
organisms could be greater, but no test results are available for
these organisms.
Benomyl and carbendazim affect groups of fungi in soil but do
not seem to modify the overall microbial activity of the soil when
used at normal field rates.
In both the laboratory and field, benomyl and carbendazim
applied at recommended rates cause deaths and sublethal reproductive
effects on earthworms of many different species. Surface-feeding
species eating leaf litter are most at risk. Populations may take
more than 2 years to recover. There are no studies available on
other litter and soil invertebrates.
Benomyl and carbendazim have low toxicity for birds and
carbendazim is classified as "relatively non-toxic" to honey-bees.
10.3 Conclusions
Benomyl causes dermal sensitization in humans. Both benomyl and
carbendazim represent a very low risk for acute poisoning in humans.
Given the current exposures and the low rate of dermal absorption of
benomyl and carbendazim, it is unlikely that they would cause
systemic toxicity effects either in the general population or in
occupationally exposed subjects. These conclusions are drawn from
animal data and from the limited human data available, but these
extrapolations are supported by the understanding of the mode of
action of carbendazim and benomyl in both target and non-target
species.
Further elucidation of the mechanism of toxicity of carbendazim
and benomyl in mammals will perhaps permit a better determination of
no-observed-effect levels. Binding studies on tubulins of target
cells (testis and embryonic tissues) will facilitate comparisons
across species.
Carbendazim is strongly adsorbed to soil organic matter and
remains in the soil for up to 3 years. It persists on leaf surfaces
and, therefore, in leaf litter. Earthworms have been shown to be
adversely affected (population and reproductive effects) at
recommended application rates. There is no information on other soil
or litter arthropods that would be similarly exposed.
The high toxicity to aquatic organisms in laboratory tests is
unlikely to be seen in the field because of the low bioavailability
of sediment-bound residues of carbendazim. However, no information
is available on sediment-living species which would receive the
highest exposure.
11. FURTHER RESEARCH
1. Comparative binding studies of carbendazim to tubulins of
target tissues from various species should be undertaken.
2. Further clarification of the fate of 1,2-diaminobenzene and
bound residues in the environment is needed.
3. The effects of benomyl and carbendazim on sediment-dwelling
organisms needs to be investigated.
12. PREVIOUS EVALUATIONS BY INTERNATIONAL BODIES
Carbendazim was evaluated by the Joint FAO/WHO Meeting on
Pesticide Residues (JMPR) in 1973, 1976, 1977, 1978, 1983 and 1988.
The 1978 meeting agreed that the MRLs for benomyl, carbendazim and
thiophanate-methyl should be combined and expressed as carbendazim.
Carbendazim residues were last evaluated by the 1988 meeting
(FAO/WHO, 1988b) and the MRLs were updated at that time. These MRLs
(expressed as carbendazim) are listed in Table 4. The 1983 meeting
(FAO/WHO, 1985a) evaluated carbendazim toxicology and set the
following NOEL levels and ADI:
Rat: 500 mg/kg diet, equivalent to 25 mg/kg body weight
Dog: 100 mg/kg diet, equivalent to 2.5 mg/kg body weight
Rat: teratology - 30 mg/kg body weight per day (benomyl)
The estimated ADI for carbendazim was established to be 0-0.01
mg/kg body weight.
Carbendazim has not been evaluated by the International Agency
for Research on Cancer (IARC).
REFERENCES
Abdel-Fattah HM, Abdel-Kader M, & Hamida S (1982) Effect of
Bavistin, Cotoran and Curacron on Egyptian soil fungi.
Mycopathologia, 80(2): 101-106.
Albertini S (1989) Influence of different factors on the induction
of chromosome malsegregation in Saccharomyces cerevisiae D61.M by
Bavistan and assessment of its genotoxic property in the Ames test
and in Saccharomyces cerevisiae D7. Mutat Res, 216: 327-340.
Albertini S (1991) Reevaluation of the 9 compounds reported
conclusive positive in yeast Saccharomyces cerevisiae aneuploidy
test systems by the Gene-Tox Program using strain D61.M of
Saccharomyces cerevisiae. Mutat Res, 260: 165-180.
Alverez L (1987) Teratogenicity study of INE-965 (carbendazim) in
rats. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc.,
Haskell Laboratory (Unpublished report No. HLR 281-87).
Amacher DE, Paillet S, & Ray VA (1979) Point mutations at the
thymidine kinase locus in L5178Y mouse lymphoma cells: Application
to genetic toxicological testing. Mutat Res, 64: 391-406.
Ames BN, McCann J, & Yamasaki E (1975) Methods of detecting
carcinogens and mutagens with the Salmonella/mammalian microsome
mutagenicity test. Mutat Res, 31: 347-364.
Ammon HU (1985) Worm toxicity tests using Tubifex tubifex. In:
Comportement et effets secondaires des pesticides dans le sol. Les
Colloques de l'INRA 31, Versailles, 4-8 juin 1984. Paris, Institut
national de la Recherche agronomique, pp 303-317.
Arce GT (1984a) Mutagenicity evaluation in Salmonella typhimurium.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 72-84).
Arce GT (1984b) Mutagenicity evaluation in Salmonella typhimurium.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 71-84).
Arthur MF, Marsh BH, Fadel LC, & Zwick TC (1989a) Anaerobic aquatic
metabolism of [phenyl(U)-14C] benomyl in West Jefferson, Ohio,
pond water and sediment (Battelle Project No. N-0799-8800).
Columbus, Ohio, Battelle, Environmental Sciences Department
(Unpublished report No. AMR 770-87, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Arthur MF, Schweitzer KL, Fadel LC, Marsh BH, & Marsh SS (1989b)
Aerobic aquatic metabolism of [phenyl(U)-14C] benomyl in
Greenville, Mississippi, water and sediment (Battelle Project No.
N-0966-7301). Columbus, Ohio, Battelle, Environmental Sciences
Department (Unpublished report No. AMR 1452-89, prepared for E.I. Du
Pont de Nemours and Co., Inc.).
Bamburg JR, Shooter EM, & Wilson L (1973) Developmental changes in
microtubule protein of the chick brain. Biochemistry, 12: 1476-1482.
Banduhn N & Obe G (1985) Mutagenicity of methyl
2-benzimidazolecarbamate, diethylstilbestrol and estradiol:
Structural chromosomal aberrations, sister-chromatid exchanges,
C-mitoses, polyploidies and micronuclei. Mutat Res, 156: 199-218.
BASF (1975) Dominant lethal study: mice. Ludwigshafen, Germany, BASF
AG.
Baude FJ, Gardiner JA, & Han JC-Y (1973) Characterization of
residues on plants following foliar spray applications of benomyl. J
Agric Food Chem, 21: 1084-1090.
Baude FJ, Pease HL, & Holt RF (1974) Fate of benomyl on field soil
and turf. J Agric Food Chem, 22: 413.
Beavers JB (1985) An acute oral toxicity study in the bobwhite quail
with H-15,607 (MBC). St. Michaels, Maryland, Wildlife International
Ltd (Unpublished report No. HLO 96-85, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Beems RB, Til HP, & van der Heijden CA (1976) Carcinogenicity study
with carbendazim (99% MBC) in mice. The Hague, Central Institute for
Nutrition and Food Research (TNO) (Summary Report No. R-4936,
prepared for Hoechst AG, Frankfurt and BASF AG, Ludwigshafen).
Boeri RL (1988a) Flow through acute toxicity of carbendazim
technical to the eastern oyster, Crassostrea virginica.
Marblehead, Massachusetts, Enseco (Unpublished report No. HLO 15-89,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Boeri RL (1988b) Static acute toxicity of carbendazim technical to
the sheepshead minnow, Cyprinodon variegatus. Marblehead,
Massachusetts, Enseco (Unpublished report No. HLO 12-89, prepared
for E.I. Du Pont de Nemours and Co., Inc.).
Boeri RL (1988c) Static acute toxicity of carbendazim technical to
the mysid, Mysidopsis bahia. Marblehead, Massachusetts, Enseco
(Unpublished report No. HLO 829-88, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Bushway RJ, Hurst HL, Kugabalasooriar J, & Perkins LB (1991)
Determination of carbendazim in blueberries by reversed-phase high
performance liquid chromatography. J Chromatogr, 555: 321-324.
Canton JH (1976) The toxicity of benomyl, thiophanate-methyl, and
BLM to four freshwater organisms. Bull Environ Contam Toxicol, 16:
214-218.
Carter SD, Hess RA, & Laskey JW (1987) The fungicide methyl
2-benzimidazolecarbamate causes infertility in male Sprague-Dawley
rats. Biol Reprod, 37(3): 709-718.
Chiba M & Veres DF (1981) Fate of benomyl and its degradation
compound methyl 2-benzimidazole carbamate on apple foliage. J Agric
Food Chem, 29: 588-590.
Christian MS, Hoberman AM, & Feussner EL (1985) Developmental
toxicity study of carbendazim administered via gavage to New Zealand
white rabbits. Horsham, Pennsylvania, Argus Research Laboratories,
Inc. (Unpublished report No. HLO 515-85, prepared for E.I. Du Pont
de Nemours and Co., Inc.).
Clive D, Johnson KO, Spector JFS, Batson AG, & Moore-Brown MM (1979)
Validation and characterization of the L5178Y/TK +/- mouse
lymphoma mutagen assay system. Mutat Res, 59: 61-108.
Cook ME & Swait AAJ (1975) Effects of some fungicide treatments on
earthworm populations and leaf removal in apple orchards. J Hortic
Sci, 50: 495-499.
County Natwest Woodmac (1992) The fungicide market. London, County
Natwest Woodmac, Agrochemical Service, pp 49-51.
Cox DM & Puck TT (1969) Chromosomal non-disjunction: The action of
colcemid on Chinese hamster ovary cells in vitro. Cytogenetics, 8:
158-169.
Cummings AM, Harris ST, & Rehnberg GL (1990) Effects of methyl
benzimidazole carbamate during early pregnancy in the rat. Fundam
Appl Toxicol, 15: 528-535.
Cummings AM, Ebron-McCoy MT, Rogers JM, Barbee BD, & Harris ST
(1992) Developmental effects of methyl benzimidazole carbamate
following exposure during early pregnancy. Fundam Appl Toxicol, 18:
288-293.
Dashiell OL (1975) Oral LD50 test in guinea pigs (Carbendazim).
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 846-74).
Davidse LC (1973) Antimitotic activity of methyl benzimidazol-2-yl
carbamate (MBC) in Aspergillus nidulans. Pestic Biochem Physiol,
3: 317-325.
Davidse LC (1975) Antimitotic activity of methyl
benzimidazol-2-ylcarbamate in fungi and its binding to cellular
protein. In: Borgers M & de Brabander M ed. Microtubules and
microtubule inhibitors. Amsterdam, Elsevier/North-Holland Publishing
Company, pp 483-495.
Davidse LC & Flach W (1977) Differential binding of methyl
benzimidazol-2-yl carbamate to fungal tubulin as a mechanism of
resistance to this antimitotic agent in mutant strains of
Aspergillus nidulans. J Cell Biol, 72: 174-193.
Davidse LC & Flach W (1978) Interaction of thiabendazole with fungal
tubulin. Biochim Biophys Acta, 543: 82-90.
Delatour P & Besse S (1990) Benzimidazole carbamate d'éthyle: effet
tératogène et présence dans le lait de vache après administration de
thiophanate. Ann Rech Vét, 21: 87-92.
Delp CJ (1980) Coping with resistance to plant disease control
agents. Plant Dis, 64: 652-657.
De Brabander M, van de Veire R, Aerts F, Geuens S, & Hoebeke J
(1976a) A new culture model facilitating rapid quantitative testing
of mitotic spindle inhibition in mammalian cells. J Natl Cancer
Inst, 56: 357-363.
De Brabander M, van de Veire R, Aerts F, Brogers M, & Janssen PAJ
(1976b) The effect of methyl [5(2-thienylcarbonyl)-1H-benzimidazol-
2-yl carbamate (R17934; NSC 238159), a new synthetic antitumoral
drug interfering with microtubules, on mammalian cells cultured in
vitro. Cancer Res, 36: 905-916.
De Vos RH, van Dokkum W, Olthof PDA, Quirijns JK, Muys T, & van der
Poll JM (1984) Pesticides and other chemical residues in Dutch total
diet samples (June 1976 - July 1978). Food Chem Toxicol, 22: 11-21.
Donaubauer HH, Schuetz E, Weigand W, & Kramer M (1982) Repeated dose
(24 month) feeding study for determination of the carcinogenic
effect of HOE 17411 OFAT204 (carbendazim) in mice. Frankfurt,
Germany, Hoechst AG, Pharmaceuticals Research, Toxicology Section
(Unpublished report).
Donovan SD (1982) Mutagenicity evaluation in Salmonella
typhimurium. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 678-82).
Donovan SM (1983) Mutagenicity evaluation in Salmonella
typhimurium. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 797-82).
Dorn E, Schmidt E, Kellner HM, & Leist KH (1983) HOE 017411-14-C
(carbendazim-14-C) metabolic fate in rats and mice, a comparison.
Frankfurt, Germany, Hoechst AG.
Douglas MT & Handley JW (1987) The algistatic activity of
carbendazim technical. Huntingdon, United Kingdom, Huntingdon
Research Centre Ltd (Unpublished report No. DPT 171(g)/871604,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Du Pont (1972) Residue studies - fish: benomyl, MBC, and 2-AB.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report).
Eckert J, Stauffer R, & Hoppman D (1985) Determination of MBC, 4-OH
MBC, and 5-OH MBC in chicken eggs, tissues and feces. Bernville,
Pennsylvania, Enviro-Bio-Tech Ltd (Unpublished report No. AMR
260-84, prepared for E.I. Du Pont de Nemours and Co., Inc.).
Edwards DF (1974a) Skin absorption LD50 (carbendazim). Newark,
Delaware, Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 798-74).
Edwards DF (1974b) Federal Hazardous Substances Act - Eye irritation
test in rabbits (carbendazim). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 799-74).
Ellis WG, Semple JL, Hoogenboom ER, Kavlock RJ, & Zeman FJ (1987)
Benomyl-induced craniocerebral anomalies in fetuses of adequately
nourished and protein-deprived rats. Teratog Carcinog Mutagen, 7:
357-375.
Ellis WG, De Roos F, Kavlock RJ, & Zeman FJ (1988) Relationship of
periventricular overgrowth to hydrocephalus in brains of fetal rats
exposed to benomyl. Teratog Carcinog Mutagen, 8: 377-391.
Evenson DP, Janca FC, & Jost LK (1987) Effects of the fungicide
methyl-benzimidazol-2-yl carbamate (MBC) on mouse germ cells as
determined by flow cytometry. J Toxicol Environ Health, 20: 387-399.
Everhart LP (1979) Benlate dust exposure survey. Wilmington,
Delaware, E.I. Du Pont de Nemours and Co., Inc. (Unpublished report
No. B/TOX 6).
Everhart LP & Holt RF (1982) Potential benlate fungicide exposure
during mixer/loader operations, crop harvest and home use. J Agric
Food Chem, 30: 222-227.
Fahrig R & Seiler JP (1979) Dose and effect of methyl
2-benzimidazolyl-carbamate in the "mammalian spot test", an in vivo
method for the detection of genetic alterations in somatic cells of
mice. Chem-Biol Interact, 26: 115-120.
FAO/WHO (1985a) Benomyl. In: Pesticide residues in food -
Evaluations 1983. Rome, Food and Agriculture Organization of the
United Nations, pp 7-46 (FAO Plant Production and Protection Paper
61).
FAO/WHO (1985b) Carbendazim. In: Pesticide residues in food -
Evaluations 1983. Rome, Food and Agriculture Organization of the
United Nations, pp 89-121 (FAO Plant Production and Protection Paper
61).
FAO/WHO (1988a) Benomyl. In: Pesticide residues in food -
Evaluations 1988. Part I: Residues. Rome, Food and Agriculture
Organization of the United Nations, pp 5-15 (FAO Plant Production
and Protection Paper 93/1).
FAO/WHO (1988b) Carbendazim. In: Pesticide residues in food -
Evaluations 1988. Part I: Residues. Rome, Food and Agriculture
Organization of the United Nations, pp 41-54 (FAO Plant Production
and Protection Paper 93/1).
Fielding M, Barcelo D, Helweg A, Galassi S, Torstensson L, van
Zoonen P, Wolter R, & Angeletti G (1992) Pesticides in ground and
drinking water. Brussels, Commission of the European Communities,
Directorate-General for Science, Research and Development,
Environment and Waste Recycling, 135 pp (Water Pollution Research
Report 27).
Fink R (1975) Eight-day dietary LC50 - Mallard ducks. Sterling,
Virginia, Truslow Farms, Inc. (Unpublished report No. HLO 108-75,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Fiscor G, Bordas S, & Steward SJ (1978) Mutagenicity testing of
benomyl, methyl-2-benzimidazole carbamate, streptozotocin and
N-methyl-N'-nitro-N-nitrosoguanine in Salmonella typhimurium
in vitro and in rodent host-mediated assays. Mutat Res, 51:
151-164.
Fleeker JR & Lacy HM (1977) Photolysis of methyl
2-benzimidazole-carbamate. J Agric Food Chem, 25: 51-55.
Ford LS (1981) Primary skin irritation and sensitization on guinea
pigs. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc.,
Haskell Laboratory (Unpublished report No. HLR 729-81).
Ford LS (1982) Acute skin absorption LD50 test on rabbits
(carbendazim 75% wettable powder). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 822-81).
Freidman PA & Platzer EG (1978) Interaction of anthelmintic
benzimidazoles and benzimidazoles derivatives with bovine brain
tubulin. Biochim Biophys Acta, 544: 605-614.
Galloway SM & Ivett JL (1986) Chemically induced aneuploidy in
mammalian cells in culture. Mutat Res, 167: 89-105.
Gericke D (1977) Test for mutagenicity in bacteria strains in the
absence and presence of a liver preparation. Frankfurt, Germany,
Hoechst AG, Laboratory for Cancer Research (Report No. A/12202).
Gillet C & Roubaud P (1983) Prehatching survival of carp eggs after
treatment during fertilization and early development with the
antimitotic fungicide carbendazim. Water Res, 17: 1343-1348.
Goldenthal EI (1978) Neurotoxicity in hens. Mattawan, Michigan,
International Research and Development Corporation (Unpublished
report No. HLO 27-79, prepared for E.I. Du Pont de Nemours and Co.,
Inc.).
Goldman JM, Rehnberg GL, Cooper RL, Gray LE Jr, Hein JF, & McElroy
WK (1989) Effects of the benomyl metabolite, carbendazim, on the
hypothalamic-pituitary reproductive axis in the male rat.
Toxicology, 57(2): 173-182.
Gooch JJ (1978) Fertility of workers potentially exposed to benomyl.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report No. B/TOX 7).
Goodman NC (1974) Intraperitoneal LD50 test [in rats using
technical carbendazim (less than 98%)]. Newark, Delaware, E.I. Du
Pont de Nemours and Co., Inc., Haskell Laboratory (Unpublished
report No. HLR 845-74).
Goodman NC & Sherman H (1975) Oral LD50 tests (fasted male and
female rats). Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 847-74).
Grandizio AM & Sarver JW (1987) Acute oral toxicity study with
INE-965-261 (Benlate C) in male and female rats. Newark, Delaware,
E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 358-87).
Gray LE, Ostby J, Sigmon R, Ferrell J, Rehnberg G, Linder R, Cooper
R, Goldman J, & Laskey J (1988) The development of a protocol to
assess reproductive effects of toxicants in the rat. Reprod Toxicol,
2: 281-287.
Gray LE, Ostby J, Linder R, Goldman J, Rehnberg G, & Cooper R (1990)
Carbendazim-induced alterations of reproductive development and
function in the rat and hamster. Fundam Appl Toxicol, 15: 281-297.
Guengerich FP (1981) Enzyme induction with Du Pont compounds
H11,201-02 and H10,962-02. Nashville, Tennessee, Vanderbilt
University, School of Medicine (Unpublished report No. HLO-850-81,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Gupta VK & Sharma SD (1989) Degradation of carbendazim in soil by
microorganisms. Indian J Agric Sci, 59: 21-24.
Gustafson DI (1989) Groundwater ubiquity score: A simple method for
assessing pesticide leachability. Environ Toxicol Chem, 8: 339-357.
Hammerschlag RS & Sisler HD (1973) Benomyl and
methyl-2-benzimidazole (MBC): biochemical, cytological, and chemical
aspects of toxicity to Ustilage maydis and Saccharomyces
cerevesiae. Pestic Biochem Physiol, 3: 42-54.
Han JC-Y (1983a) Characterization of residues in bean plants
following foliar spray applications with methyl
2-14C-benzimidazolecarbamate (MBC). Wilmington, Delaware, E.I. Du
Pont de Nemours and Co., Inc. (Unpublished report No. MBC/ME-4).
Han JC-Y (1983b) Anaerobic soil metabolism of 2-14C-benomyl and
methyl 2-14C-benzimidazolecarbamate. Wilmington, Delaware, E.I. Du
Pont de Nemours and Co., Inc. (Unpublished report No. B/ME-44).
Heinemann JM (1971) Acute static fish (rainbow trout) toxicity study
- freshwater (MBC). Vienna, Virginia, Hazelton Laboratories
(Unpublished report No. HLO-150-71, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Helweg A (1977) Degradation and adsorption of carbendazim and
2-aminobenzimidazole in soil. Pestic Sci, 8: 71-78.
Helweg A (1979) Influence of temperature, humidity, and inoculation
on the degradation of 14C-labelled 2-aminobenzimidazole in soil.
Water Air Soil Pollut, 12: 275-281.
Helweg A (1985) Side effects caused by pesticide combinations. In:
Jensen V, Kjoller A, & Sorensen LH ed. Proceedings of the FEMS
Symposium on Microbial Communities in Soil. Amsterdam, Oxford, New
York, Elsevier Science Publishers, pp 385-395.
Hill EF & Camardese MB (1986) Lethal toxicities of environmental
contaminants and pesticides to Coturnix. Washington, DC, US
Department of the Interior, Fish and Wildlife Service (Fish and
Wildlife Technical Report No. 2).
Hinckle L (1981) Oral LD50 test in rats - EPA proposed guidelines.
Newark, Delaware, E.I. du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 769-81).
Hoechst (1974) Mouse dominant lethal study on MBC. Frankfurt,
Germany, Hoechst AG.
Hofer I, Beck T, & Wallnöfer P (1971) [The influence of the
fungicide benomyl on soil microflora.] Z Pflanzenkr Pflanzenschutz,
78: 399-405 (in German).
Holden HE, Crider PA, & Wahrenburg MG (1980) Mitotic arrest by
benzimidazole analogs in human lymphocyte cultures. Environ Mutat,
2: 67-73.
Hughes DL (1984) Residue study of the fungicide MBC in lactating
dairy cattle. Madison, Wisconsin, Hazelton Laboratories America,
Inc. (Unpublished report No. AMR 429-85, prepared for E.I. Du Pont
de Nemours and Co., Inc.).
Hutton DG (1988) Static acute 48-hours EC50 of carbamic acid,
1H-benzimidazol-2-yl-, methyl ester to fed Daphnia magna. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 638-88).
Hutton DG, Kasprzak DJ, & Priester TM (1984) Laboratory studies of
[2-14C] carbendazim bioconcentration in bluegill sunfish. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 428-84).
Hutton DG, Lieder PH, & Grapel GJ (1986) Chronic toxicity of
carbamic acid, 1H-benzimidazol-2-yl-, methyl ester to Daphnia
magna. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc.,
Haskell Laboratory (Unpublished report No. HLR 241-86).
ILO (1991) Occupational exposure limits for airborne toxic
substances, 3rd ed. Geneva, International Labour Office, 455 pp
(Occupational Safety and Health Series No. 37).
Ireland CM, Gull K, Gutteridge WE, & Pogson CI (1979) The
interaction of benzimidazole carbamates with mammalian microtubule
protein. Biochem Pharmacol, 28: 2680-2682.
Ivett JL (1984) Mutagenicity evaluation of H#15,152 in an in vitro
sister chromatid exchange assay in Chinese hamster ovary (CHO)
cells. Kensington, Maryland, Litton Bionetics, Inc. (Unpublished
report No. HLO 254-84, prepared for E.I. Du Pont de Nemours and Co.,
Inc.).
Janardhan A, Sattur PB, & Sisodia P (1984) Teratology of methyl
benzimidazole carbamate in rats and rabbits. Bull Environ Contam
Toxicol, 33(3): 257-263.
Janardhan A, Rao AB, & Sisodia P (1987) Sub-chronic toxicity of
methyl benzimidazole carbamate in rats. Bull Environ Contam Toxicol,
38(5): 890-898.
Jansz GF & Pomerantz DK (1985) The effect of prenatal treatment with
busulfan on in vitro androgen production by testes from rats of
various ages. Can J Physiol Pharmacol, 63: 1155-1158.
Johnson JD (1988) Determination of the plateau level of bound
[phenyl(U)-14C]carbendazim residues in goat liver. Columbus, Ohio,
Battelle Columbus Division (Unpublished report No. AMR-779-87,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Jotz MM (1980) An evaluation of mutagenic potential of benomyl
employing the L5178Y TK+/- mouse lymphoma assay (SRI Project No.
LSU 7558). Menlo Park, California, SRI International (Unpublished
report prepared for the US Environmental Protection Agency).
Keogh RG & Whitehead PH (1975) Observations on some effects of
pasture spraying with benomyl and carbendazim on earthworm activity
and litter removal from pasture. NZ J Exp Agric, 3: 103-104.
Khan S, Khan NN, & Khan FA (1987) The influence of bavistin and
calixin on carbon dioxide-evolution and the major nutrient status
(nitrogen, phosphorus, potassium) of the soil. Environ Pollut,
47(2): 115-122.
Kirkland JJ (1973) Method for high-speed liquid chromatographic
analysis of benomyl and/or metabolite residue in cow milk, urine,
feces and tissues. J Agric Food Chem, 21: 171-177.
Kirkland JJ, Holt RF, & Pease HL (1973) Determination of benomyl
residues in soils and plant tissues by high-speed cation exchange
liquid chromatography. J Agric Food Chem, 21: 368-371.
Koeter HBWM (1975a) Effect of HOE 17411F (=BAS 3460F) on pregnancy
of the rat. The Hague, Central Institute of Nutrition and Food
Research (TNO) (Unpublished report No. R-4726, prepared for Hoechst
AG, Frankfurt and BASF AG, Ludwigshafen).
Koeter HBWM (1975b) Effect of HOE 17411F (=BAS 3460F) on pregnancy
of the New Zealand white rabbit. The Hague, Central Institute for
Nutrition and Food Research (TNO) (Unpublished report No. R-4881,
prepared for Hoechst AG, Frankfurt and BASF AG, Ludwigshafen).
Kramer M & Weigand W (1971) (HOE 17411 O F) Toxicological
examination. Frankfurt, Germany, Hoechst AG, Pharmaceuticals
Research, Toxicology Section (Unpublished report No. A-00936).
Krechniak J & Klosowska B (1986) The fate of 14C-carbendazim in
rat. Xenobiotica, 16(9): 809-815.
Lamb MJ & Lilly LJ (1980) An investigation of some genetic
toxicological effects of the fungicide benomyl. Toxicology, 17:
83-95.
Litton (1981) Evaluation of HOE 17411 O F AT204 in the primary rat
hepatocyte unscheduled DNA synthesis assay (LBI Project No. 21001).
Kensington, Maryland, Litton Bionetics, Inc. (Unpublished report
prepared for Hoechst AG, Frankfurt).
Lofs-Holmin A (1981) Influence in field experiments of benomyl and
carbendazim on earthworms ( Lumbricidae) in relation to soil
texture. Swed J Agric Res, 11: 141-147.
Lofs-Holmin A (1982) Influence of routine pesticide spraying on
earthworms ( Lumbricidea) in field experiments with winter wheat.
Swed J Agric Res, 12: 121-124.
Marsh BH & Arthur MF (1989) Aerobic metabolism of
[phenyl(U)-14C]benomyl in keyport silt loam. Columbus, Ohio,
Battelle Memorial Institute (Unpublished report No. AMR 1112-88,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Martin DA, Henry JE, & Brock WJ (1987) Closed-patch repeated insult
dermal sensitization study (Buehler Method) with Benlate C fungicide
in guinea-pigs. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 510-87).
Marvin Ch, Birndle ID, Hall CD, & Chiba M (1991) Rapid on-line
precolumn high performance liquid chromatographic method for the
determination of benomyl, carbendazim and aldicarb species in
drinking water. J Chromatogr, 555: 147-154.
Mayer FL & Ellersieck MR (1986) Manual of acute toxicity:
Interpretation and data base for 410 chemicals and 66 species of
freshwater animals. Washington, DC, US Department of the Interior,
Fish and Wildlife Service, pp 45-46 (Resource Publication No. 160).
Mazur AR & Hughes TD (1975) Nitrogen transformations in soil as
affected by the fungicides benomyl, dyrene and maneb. Agron J, 67:
755-758.
Meade AB (1984) Acute contact LD50 toxicity study in honey bees
( Apis mellifera L.) with INE965-212 (MBC). Wilmington, Delaware,
E.I. Du Pont de Nemours and Co., Inc. (Unpublished report No.
ABM-84-5).
Mohr U (1977) Review of liver sections from mice and rats fed with
carbendazim. Hannover, Germany, Medical School, Department of
Experimental Pathology (Unpublished report No. MBC/TOX 6, prepared
for Hoechst AG, Frankfurt and BASF AG, Ludwigshafen).
Monson KD (1985) Metabolism of [2-14C]-carbendazim in the
lactating dairy cow. Wilmington, Delaware, E.I. Du Pont de Nemours
and Co., Inc. (Unpublished report No. AMR 248-84).
Monson KD (1986) Metabolism of [2-14C]-carbendazim in laying hens.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report No. AMR 264-84).
Monson KD (1990) Metabolism of [phenyl(U)-14C]carbendazim in rats.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report No. AMR-1141-88).
Monson KD (1991) Release and characterization of bound benomyl and
carbendazim metabolites in animal tissues via Raney nickel
desulfurization and acid dehydration. J Agric Food Chem, 39:
1808-1811.
Nakai M, Hess RA, Moore BJ, Guttroff RF, Strader LF, & Linder RE
(1992) Acute and long-term effects of a single dose of the fungicide
carbendazim (methyl 2-benzimidazole carbamate) on the male
reproductive system in the rat. J Androl, 13: 507-518.
Nash SD & Ferenz R (1982) Inhalation median lethal concentration
(LC50 in rats -EPA protocol (carbendazim 75% wettable powder).
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 365-82).
Newsome WH & Collins PG (1987) Enzyme-linked immunosorbent assay of
benomyl and thiabendazole in some foods. J Assoc Off Anal Chem, 70:
1025-1027.
Newsome WH & Shields JB (1981) A radioimmunoassay for benomyl and
methyl 2-benzi midazolecarbamate on food. J Agric Food Chem, 29:
220-222.
Nishiuchi Y & Yoshida K (1974) Toxicity of new agricultural
chemicals to tadpoles. Bull Agric Chem Insp Stn, 14: 66-68.
Osgood C, Zimmering S, & Mason JM (1991) Aneuploidy in Drosophila.
II. Further validation of the FIX and ZESTE genetic test systems
employing female Drosophila melanogaster. Mutat Res, 259: 147-163.
Palawski DU & Knowles CO (1986) Toxicological studies of benomyl and
carbendazim in rainbow trout, channel catfish and bluegills. Environ
Toxicol Chem, 5: 1039-1046.
Pandita TK (1988) Assessment of the mutagenic potential of a
fungicide Bavistin using multiple assays. Mutat Res, 204: 627-643.
Parsons DW & Witt JM (1988) Pesticides in groundwater in the United
States of America: A report of a 1988 survey of State lead agencies.
Corvallis, Oregon, Oregon State University Extension Service.
Peeples JL (1974) Microbial activity in benomyl-treated soil.
Phytopathology, 64: 857-860.
Perreault SD, Jeffay S, Poss P, & Laskey JW (1992) Use of the
fungicide carbendazim as a model compound to determine the impact of
acute chemical exposure during oocyte maturation and fertilization
on pregnancy outcome in the hamster. Toxicol Appl Pharmacol, 114:
225-231.
Priester TM (1984) Hydrolysis of carbendazim [2-14C]. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. AMR 265-84).
Purser D (1987) Carbendazim: behaviour in water. Harrogate, North
Yorkshire, United Kingdom, Hazelton Laboratories (Unpublished report
No. 5220-269/12, prepared for E.I. Du Pont de Nemours and Co.,
Inc.).
Ramakrishna C, Gowda TKS, & Sethunathan N (1979) Effect of benomyl
and its hydrolysis products, MBC and AB on nitrification in flooded
soil. Bull Environ Contam Toxicol, 21: 328-333.
Rashid MA & Mumma RO (1986) Screening pesticides for their
availability to damage bacterial DNA. J Environ Sci Health, B21:
319-334.
Raw F (1962) Studies of earthworm populations in orchards. Ann Appl
Biol, 50: 389-404.
Rehnberg GL, Cooper RL, Goldman JM, Gray LE, Hein JF, & McElroy WK
(1989) Serum and testicular testosterone and androgen binding
protein profiles following subchronic treatment with carbendazim.
Toxicol Appl Pharmacol, 101: 55-61.
Reuzel PGJ, Hendriksen CFM, & Til HP (1976) Long-term (two-year)
toxicity study with carbendazim in beagle dogs. The Hague, Central
Institute for Nutrition and Food Research (TNO) (Unpublished report
prepared for BASF AG, Ludwigshafen and Hoechst AG, Frankfurt).
Rhodes RC (1987) Greenhouse crop-rotation study with
[2-14C]carbendazim. Wilmington, Delaware, E.I. Du Pont de Nemours
and Co., Inc. (Unpublished report No. AMR-495-86).
Rhodes RC & Long JD (1983) Run-off and leaching studies with methyl
2-14C-benzimidazolecarbamate on soil. Wilmington, Delaware, E.I.
Du Pont de Nemours and Co., Inc. (Unpublished report No. MBC/ME-9).
Rhodes RC, Pease HL, & Holt RF (1983) Greenhouse studies on crop
uptake of MBC and 2-AB from soil. Wilmington, Delaware, E.I. Du Pont
de Nemours and Co., Inc. (Unpublished Report No. MBC/ME-5).
Roberts BL & Dorough HW (1984) Relative toxicities on chemicals to
the earthworm Eisenia foetida. Environ Toxicol Chem, 3: 67-78.
Russel LD, Ettlin RA, Sinha Hikim AP, & Clegg ER (1990) Histological
and histopathological evaluation of the testis. Clearwater, Florida,
Cache River Press.
Russell JF (1977) Mutagenicity evaluation of 2-benzimidazolecarbamic
acid, 5-hydroxy-, methyl in Salmonella typhimurium. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report).
Russell JF (1983) Mutagenicity evaluation in Salmonella
typhimurium. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 179-83).
Sarver JW (1975) Acute inhalation toxicity - one hour head only
(carbendazim). Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 58-75).
Scholz J & Weigand W (1972) Carbendazim (HOE 17411 O F, batch
496/I). Toxicological examination. Frankfurt, Germany, Hoechst AG,
Pharmaceuticals Research, Toxicology Section (Unpublished report No.
A-00935).
Seiler JP (1976) The mutagenicity of benzimidazole and benzimidazole
derivatives. VI. Cytogenetic effects of benzimidazole derivatives in
the bone marrow of the mouse and the Chinese hamster. Mutat Res, 40:
339-348.
Seiler JP (1977) Apparent and real thresholds: A study on two
mutagens. In: Scott D, Bridges BA, & Sobels FH ed. Progress in
genetic toxicology. Amsterdam, Elsevier/North-Holland Biomedical
Press, pp 233-238.
Sheir-Neiss F, Nardi RV, Gealt A, & Morris NR (1976) Tubulin-like
protein from Aspergillus nidulans. Biochem Biophys Res Commun, 69:
285-290.
Sherman H (1965) Acute oral test. Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 125-65).
Sherman H (1968) Ninety-day feeding study in rats using a wettable
powder formulation (70% MBC). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 95-68).
Sherman H (1970) Three-month feeding study in dogs using a wettable
powder formulation (50% MBC). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 283-70).
Sherman H (1972) Long-term feeding studies in rats and dogs with
2-benzimadazole carbamic acid, methyl ester (INE-965) (50% and 70%
MBC wettable powder formulations). Parts I and II. Newark, Delaware,
E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 195-72).
Sherman H & Krauss WC (1966) Acute oral test (carbendazim). Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 99-66).
Sheu CW, Lee JK, Arras CA, Jones RL, & Lavappa KS (1990) The
feasibility of using Chinese hamster as an animal model for
aneuploidy. Environ Mol Mutagen, 16: 320-323.
Shirasu Y, Moriya M, & Kato K (1977) Mutagenicity testing on
fungicide 1991 in microbial systems. Tokyo, Institute of
Environmental Toxicology, Kodaira Laboratories (Unpublished report
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Shukla Y, Antony M, & Mehrotra NK (1989) Studies on g-glutamyl
transpeptidase in rodents exposed to benomyl. Bull Environ Contam
Toxicol, 42: 301-306.
Siebert D, Zimmermann FK, & Lemperle E (1970) Genetic effects of
fungicides. Mutat Res, 10: 533-543.
Speakman JB & Nirenberg HI (1981) Mutagenicity of methyl
benzimidazole-2-yl carbamate (MBC) towards Aspergillus nidulans
(Eidam) Winter and Cladosporium cucumerinum Ellis and Arth. Mutat
Res, 88: 45-51.
Stadler JC (1986) One-year feeding study in dogs with carbendazim.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 291-86).
Staub T & Sozzi D (1984) Fungicide resistance: A continuing
challenge. Plant Dis, 68: 1026-1031.
Stringer A & Lyons C (1974) The effect of benomyl and
thiophanate-methyl on earthworm populations in apple orchards.
Pestic Sci, 5: 189-196.
Stringer A & Wright MA (1973) The effect of benomyl and some related
compounds on Lumbricus terrestris and other earthworms. Pestic
Sci, 4: 165-170.
Styles JA & Garner R (1974) Benzimidazolecarbamate methyl ester -
evaluation of its effects in vivo and in vitro. Mutat Res, 26:
177-187.
Til HP, van den Muelen HC, Feron VJ, Seinen W, & de Groot
AP (1972) Sub-chronic (90-day) toxicity study with W17411 in beagle
dogs. The Hague, Central Institute for Nutrition and Food Research
(TNO) (Unpublished report No. R-3920, prepared for Hoechst AG,
Frankfurt).
Til HP, Koellen C, & van der Heijden CA (1976a) Combined chronic
toxicity and carcinogenicity study with carbendazim in rats. The
Hague, Central Institute for Nutrition and Food Research (TNO)
(Unpublished report prepared for BASF AG, Ludwigshafen and Hoechst
AG, Frankfurt).
Til HP, Koeter HBWM, & van der Heijden CA (1976b) Multigeneration
study with carbendazim in rats. The Hague, Central Institute for
Nutrition and Food Research (TNO) (Unpublished report No. R-5024,
prepared for Hoechst AG, Frankfurt).
Til HP, Beems RB, & de Groot AP (1981) Determination of the acute
oral toxicity of carbendazim in mice. The Hague, Central Institute
for Nutrition and Food Research (TNO) (Unpublished report No.
A-21604, prepared for BASF AG, Ludwigshafen).
Tong C (198la) Hepatocyte primary culture/DNA repair assay on
compound l1, 201-01 (MBC) using mouse hepatocytes in culture.
Valhalla, New York, Naylor Dana Institute (Unpublished report No.
HLO 743-81, prepared for E.I. Du Pont de Nemours and Co., Inc.).
Tong C (198lb) Hepatocyte primary culture/DNA repair assay on
compound 11,201-01 (MBC) using rat hepatocytes in culture. Valhalla,
New York, Naylor Dana Institute (Unpublished report No. HLO 744-81,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Torstensson L & Wessen B (1984) Interactions between the fungicide
benomyl and soil microorganisms. Soil Biol Biochem, 16: 445-452.
Van Faassen HG (1974) Effect of the fungicide benomyl on some
metabolic processes and on numbers of bacteria and actinomycetes in
the soil. Soil Biol Biochem, 6: 131-133.
Van Gestel CAM (1992) Validation of earthworm toxicity tests by
comparison with field studies: A review of benomyl, carbendazim,
carbofuran, and carbaryl. Ecotoxicol Environ Saf, 23: 221-236.
Van Gestel CAM, Dirven-van Breeman EM, Baerselman R, Emans HJB,
Janssen JAM, Postuma R, & van Vliet PJM (1992) Comparison of
sublethal and lethal criteria for nine different chemicals in
standardized toxicity tests using the earthworm Eisenia andrei.
Ecotoxicol Environ Saf, 23: 206-220.
Vick DA & Brock WJ (1987a) Primary dermal irritation study with
Benlate C fungicide in rabbits. Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 299-87).
Vick DA & Brock WJ (1987b) Acute dermal toxicity study of Benlate C
fungicide in rabbits. Newark, Delaware, E.I. Du Pont de Nemours and
Co., Inc., Haskell Laboratory (Unpublished report No. HLR 303-87).
Vick DA & Valentine R (1987) Primary eye irritation study with
Benlate C fungicide in rabbits. Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 287-87).
Vonk JW, Adema OMM, & Barug D (1986) Comparison of the effects of
several chemicals on microorganisms, higher plants and earthworms.
In: Assink JW & van den Brink WG ed. Contaminated soil. Dordrecht,
Boston, Lancaster, Martinus Nijhoff Publishers, p 191.
Wainwright M & Pugh GJF (1974) The effect of fungicides on certain
chemical and microbial properties of soils. Soil Biol Biochem, 6:
263-267.
Wallin M, Friden B, & Billger M (1989) Studies of the interaction of
chemicals with microtubule assembly in vitro can be used as an
assay for detection of cytotoxic chemicals and possible inducers of
aneuploidy. Mutat Res, 201: 303-311.
Waterer JC (1980) Chinese hamster ovary cell assay for mutagenicity.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 660-88).
Weeks RE & Hedrick HG (1975) Influence of a systemic fungicide on
oxygen uptake by soil microorganisms. Soil Sci, 119: 280-284.
Wetzel JW & Hutton DG (1984) 96-Hour LC50 to bluegill sunfish.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 401-84).
Whittaker SG, Moser SF, Maloney DH, Piegorsch WW, Resnick MA, &
Fogel S (1990) The detection of mitotic and meiotic chromosome gain
in the yeast Saccharomyces cerevisiae: Effects of methyl
benzimidazol-2-yl carbamate, methyl methanesulfonate, ethyl
methanesulfonate, dimethyl sulfoxide, propionitrile and
cyclophosphamide monohydrate. Mutat Res, 242: 231-258.
WHO (1993) Environmental Health Criteria 148: Benomyl. Geneva, World
Health Organization.
Wood CK (1982) Long-term feeding study with 2-benzimidazole-
carbamate, methyl ester (< 99% MBC, INE-965) in mice. Parts I and
II. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 70-82).
Yarden O, Aharonson N, & Katan J (1987) Accelerated microbial
degradation of methyl benzimidizol-2-ylcarbamate in soil and its
control. Soil Biol Biochem, 19(6): 735-739.
RESUME ET CONCLUSIONS
1. Résumé
1.1 Identité, propriétés physiques et chimiques et méthodes d'analyse
Le carbendazime, un solide cristallin blanc, est un fongicide
endothérapique qui appartient à la famille du benzimidazole. Son
point de fusion est d'environ 250 °C et sa tension de vapeur est <
1 x 10-7 Pa (< 1 x 10-9 mbar) à 20 °C. Le carbendazime est
pratiquement insoluble dans l'eau, sa solubilité n'étant que de 8
mg/litre à pH 7 et à 20 °C. Il est stable dans les conditions
normales de stockage.
L'analyse des résidus de même que celle des prélèvements
effectués dans l'environnement comportent une extraction au moyen
d'un solvant organique et une purification de l'extrait par partage
liquide-liquide. Le dosage de ces résidus peut s'effectuer par
chromatographie en phase liquide à haute performance ou par titrage
immunologique.
1.2 Sources d'exposition humaine et environnementale
Le carbendazime est, parmi les fongicides de la famille du
benzimidazole, celui qui est le plus largement utilisé. Il est
présenté sous forme de dispersion ou de suspension aqueuse, de
granulés fluants dispersables dans l'eau et de poudre mouillable.
1.3 Transport, distribution et transformation dans l'environnement
Le carbendazime est d'ailleurs le produit de transformation
dans l'environnement du bénomyl, un autre fongicide
benzimidazolique; la réaction est rapide avec une demi-vie
respective de 2 et 19 heures dans l'eau et le sol. On peut donc
utiliser les résultats des études sur le bénomyl ou sur le
carbendazime pour l'évaluation des effets sur l'environnement.
Dans l'environnement, le carbendazime se décompose avec une
demi-vie de 6 à 12 mois sur le sol nu, de 3 à 6 mois sur le gazon et
de 2 à 25 mois dans l'eau en aérobiose et en anaérobiose,
respectivement. Le carbendazime est principalement décomposé par les
microorganismes. Le 2-aminobenzimidazole (2-AB) en est l'un des
principaux produits de dégradation et il est à son tour décomposé
par les microorganismes. Lors de la décomposition du bénomyl marqué
au 14C sur le noyau phényle, on a constaté que 9% seulement du
carbone-14 était éliminé sous forme de CO2 en une année
d'incubation. Le carbone-14 restant était principalement récupéré
sous forme de carbendazime et de résidus liés. L'étude de la
destinée d'un éventuel produit de dégradation (1,2-diaminobenzène)
pourrait peut-être permettre de mieux définir la voie de dégradation
des fongicides benzimidazoliques dans l'environnement.
Des études effectuées sur le terrain ou sur colonne ont montré
que le carbendazime restait dans les couches superficielles du sol.
On n'a pas mesuré l'adsorption du carbendazime dans le sol, mais on
pense qu'elle doit être aussi forte que dans le cas du bénomyl, avec
des valeurs de Koc allant de 1000 à 3600. Les valeurs de log Kow
sont respectivement de 1,36 et de 1,49 pour le bénomyl et le
carbendazime.
Un modèle de criblage basé sur les données d'adsorption et de
persistance n'a pas révélé de risque de lessivage. Cette observation
est corroborée par des analyses d'eau de pluie effectuées aux
Etats-Unis, analyses qui n'ont pas permis de déceler ce composé dans
l'un quelconque des 212 puits étudiés (la limite de détection n'a
pas été précisée). On estime que le bénomyl et le carbendazime
entraînés par ruissellement correspondent uniquement à la fraction
adsorbée aux particules de sol; d'ailleurs ces composés sont sans
doute fortement adsorbés aux sédiments présents dans l'environnement
aquatique.
Le carbendazime est hydrolysé en 2-AB. Ce produit en constitue
également le principal métabolite dans le sol et les végétaux.
Chez l'animal, le carbendazime est métabolisé en
(5-hydroxy-1H-benzimidazole-2-yl)-carbamate (5-HBC) et autres
métabolites polaires, qui sont rapidement excrétés. On n'a pas
observé d'accumulation de carbendazime dans aucun système
biologique.
1.4 Concentrations dans l'environnement et exposition humaine
Il ne semble pas qu'il existe de données résultant d'une
surveillance du carbendazime dans l'environnement. Toutefois on peut
récapituler comme suit les données d'études portant sur la destinée
écologique de ce produit.
Comme le bénomyl et le carbendazime restent stable pendant des
semaines sur les végétaux, ils peuvent être ingérés par des
organismes qui se nourrissent de feuilles mortes. Des résidus de
carbendazime peuvent subsister jusqu'à 3 ans dans le sol et les
sédiments. Toutefois la forte adsorption du carbendazime aux
particules de sol et de sédiments réduit l'exposition des organismes
terrestres et aquatiques.
Ce sont les résidus de bénomyl et de carbendazime présents sur
les cultures vivrières qui constituent la principale source
d'exposition de la population humaine dans son ensemble. L'analyse
de l'exposition par voie alimentaire qui a été effectuée aux
Etats-Unis (bénomyl plus carbendazime) et aux Pays-Bas (carbendazime
seul) a montré que la quantité moyenne ingérée était
vraisemblablement de l'ordre d'une dixième de la dose journalière
acceptable (DJA) qui est, pour le bénomyl, de 0.02 mg/kg de poids
corporel et pour le carbendazime, de 0,01 mg/kg de poids corporel.
L'exposition professionnelle au cours de la production est
inférieure à la valeur-seuil établie pour le bénomyl. Les ouvriers
agricoles qui préparent les mélanges, effectuent le remplissage ou
retournent dans les champs traités par du bénomyl ou du
carbendazime, courent un risque d'exposition cutanée correspondant à
quelques milligrammes de bénomyl par heure. Le port de dispositifs
de protection permettrait de réduire encore cette exposition. En
outre, étant donné que l'absorption percutanée est vraisemblablement
faible, il est très peu probable que le carbendazime puisse avoir
des effets toxiques généraux sur les populations humaines en étant
absorbé par cette voie.
1.5 Cinétique et métabolisme
Le carbendazime est bien absorbé (à hauteur de 80 à 85%) après
ingestion mais il l'est beaucoup moins après exposition par voie
cutanée. Une fois absorbé, le carbendazime donne naissance dans
l'organisme à de nombreux métabolites. Les principaux d'entre eux
sont le 5-HBC et les 5,6-HOBC-N-oxydes. Le 5,6-DHBC-S et le
5,6-DHBC-G constituent des métabolites mineurs.
D'après la distribution tissulaire du carbendazime, il n'y a
pas de bioconcentration. Chez le rat, la concentration la plus
élevée après administration par voie orale (< 1% de la dose), a été
constatée dans le foie. Le produit se retrouvait sous forme de
carbendazime dans les mitochondries, de 5-HBC dans le cytosol et de
2-AB dans les microsomes. On a également retrouvé le carbendazime et
ses métabolites dans les reins de poulets et de vaches après
exposition, mais il ne se trouvait pas dans les autres tissus en
quantités appréciables. Après administration de carbendazime à des
vaches laitières, on a retrouvé de petites quantités de 5-HBC et de
4-HBC dans le lait.
Soixante-douze heures après avoir été administré par voie orale
à des rats, le carbendazime était excrété dans leurs urines.
Chez les rats et les souris, de fortes doses de carbendazime
administrées, soit mêlées à la nourriture, soit par gavage, ont eu
un effet sur certaines enzymes microsomiques du foie. Ainsi il y a
eu induction de la styrène-7,8-hydrolase et de l'époxyde-hydrolase
mais réduction de l'activité de la 7-hydroxycoumarine-O-déséthylase.
On a également constaté une induction de l'activité de la
glutation-S-transférase du cytosol.
1.6 Effets sur les mammifères de laboratoire et sur les systèmes
d'épreuve in vitro
1.6.1 Exposition unique
Le carbendazime présente une faible toxicité aiguë. Les valeurs
de la DL50 vont de > 2000 à 15 000 mg/kg chez toutes sortes
d'animaux d'expérience exposés par diverses voies. Toutefois on n'a
pas noté, après exposition unique, d'effets nocifs sensibles sur la
reproduction (voir Section 1.6.5).
1.6.2 Exposition de brève durée
Administré en mélange à la nourriture pendant des périodes
allant jusqu'à 90 jours, le carbendazime a entraîné une légère
modification du poids du foie chez des rattes qui en avaient reçu
une dose quotidienne de 360 mg/kg de poids corporel. Une étude de 90
jours au cours de laquelle du carbendazime a été administré par
gavage à des rats, a permis de fixer à 16 mg/kg/jour la dose sans
effets observables, basée sur l'hépatotoxicité. Les études
d'alimentation de courte durée effectuées sur des chiens se sont
révélées insuffisantes pour l'établissement d'une dose sans effets
observables. Une étude de 10 jours, au cours de laquelle de lapins
ont été exposés au carbendazime par voir cutanée n'a pas permis de
mettre en évidence une toxicité générale à la seule dose étudiée
(200 mg/kg).
1.6.3 Irritation et sensibilisation au niveau de la peau et des
yeux
L'application du produit sur l'épiderme de lapins ou de cobayes
n'a produit aucune irritation ni sensibilisation de la peau. Chez le
lapin, l'instillation oculaire n'a produit qu'une irritation légère
à modérée de la conjonctive.
1.6.4 Exposition de longue durée
A l'issue d'une étude d'alimentation où des rats mâles et
femelles avaient reçu 2500 mg de carbendazime/kg de nourriture, on a
constaté une réduction du nombre des érythrocytes ainsi que du taux
d'hémoglobine et de l'hématocrite. Aucun effet toxique sur le foie
n'a été constaté. Parmi les rats mâles qui avaient reçu du
carbendazime à raison de 2500 mg ou davantage/kg de nourriture, on
notait une augmentation peu importante du nombre de cas d'atrophie
testiculaire diffuse et de prostatite. Chez le rat, la dose sans
effets observables est de 500 mg/kg de nourriture. Des chiens qui
avaient reçu pendant une année ou davantage une nourriture contenant
500 mg de carbendazime/kg, ont présenté une élévation du taux de
cholestérol sérique et de l'activité de la phosphatase alcaline
accompagnée d'autres indices d'une hépatotoxicité. Chez le chien, la
dose sans effets observables a été fixée à 300 mg/kg de nourriture.
Chez des souris mâles et femelles qui avaient reçu une
nourriture contenant 5000 mg de carbendazime/kg de nourriture on a
observé une augmentation du poids absolu du foie. Chez des souris
mâles ayant reçu 1500 mg de carbendazime/kg de nourriture on a
également observé une importante hypertrophie centrilobulaire avec
nécrose et hypertrophie générale du foie.
1.6.5 Reproduction, embryotoxicité et tératogénicité
Lors d'une étude de reproduction portant sur trois générations,
on a administré à des rats du carbendazime à des doses allant
jusqu'à 500 mg/kg de nourriture inclusivement; aucun effet nocif n'a
été constaté sur la reproduction de ces animaux. En revanche, après
administration du même produit par gavage pendant 85 jour à raison
de 200 mg/kg/jour, on constatait une réduction de la fertilité des
mâles. A la dose de 50 mg/kg de poids corporel, la même étude a
révélé une réduction sensible du nombre de spermatozoïdes
épididymaires.
Après administration par voie orale d'une dose unique de
carbendazime et examen histologique, on a constaté que dès les deux
premiers jours, il y avait une forte perturbation de la
spermatogénèse avec occlusion des canaux efférents et augmentation
du poids des testicules à la dose de 100 mg/kg de poids corporel.
Aucun effet n'a été observé au cours de la même étude à la dose de
50 mg/kg. Ces effets ont persisté jusqu'au 70ème jour chez les rats
ayant reçu 400 mg/kg.
Administré à des rats en doses quotidiennes supérieures à 10
mg/kg du 7ème au 16ème jour de la gestation, le carbendazime a
provoqué une augmentation du nombre de malformations et d'anomalies
chez le rat. On constatait également une légère diminution du taux
de nidation chez des lapines qui en avaient reçu respectivement 20
ou 125 mg/kg en doses quotidiennes du 7ème au 19ème jour de la
gestation ainsi qu'un accroissement de la fréquence des résorptions
à la dose quotidienne de 125 mg/kg. Le composé s'est révélé toxique
pour la mère, chez la ratte, à la dose quotidienne de 20 mg/kg et
chez la lapine à la dose quotidienne de 125 mg/kg.
Outre que le taux de gravidité était en diminution et que la
fréquence des résorptions précoces était en augmentation chez le
rat, il y avait une réduction sensible du poids des foetus aux doses
quotidiennes respectives de 20 et 90 mg/kg, et un accroissement
également notable des malformations à la dose quotidienne de 90
mg/kg. Il s'agissait essentiellement d'hydrocéphalie, de
microphthalmie, d'anophthalmie, de malformations des omoplates et de
la partie axiale du squelette (fusions vertébrales, costales et
sternébrales, exencéphalie, hémivertèbres et hyperplasie costale).
Toutefois on n'a pas constaté de malformations importantes chez le
lapin.
1.6.6 Mutagénicité et autres points d'aboutissement des effets
toxiques
Les épreuves effectuées in vitro et in vivo sur des
systèmes mammaliens et non-mammaliens ainsi que sur des cellules
somatiques ou germinales montrent que le carbendazime n'interagit
pas avec l'ADN, qu'il ne provoque pas de mutations ponctuelles et ne
cause pas de mutation des cellules germinales.
Toutefois le carbendazime provoque des aberrations dans le
nombre des chromosomes (aneuploïdie et/ou polyloïdie) dans les
systèmes d'épreuve (tant in vitro qu' in vivo).
1.6.7 Cancérogénicité
L'administration de bénomyl et de carbendazime mêlés à la
nourriture a provoqué chez des souris CD-1 et Swiss axéniques une
augmentation dans l'incidence des tumeurs hépatocellulaires.
Une étude de cancérogénicité a été effectuée avec le
carbendazime sur des souris CD-1; elle a révélé une augmentation
statistiquement significative et liée à la dose dans l'incidence des
néoplasmes hépatocellulaires chez les femelles. Il y avait également
une augmentation statistiquement significative à la dose moyenne
(1500 mg/kg) de ces anomalies chez les mâles, augmentation qui n'a
pas été constatée aux doses élevées en raison du taux élevé de
mortalité. Une autre étude de cancérogénicité portant sur le
carbendazime a été effectuée chez une souche génétiquement
apparentée de souris Swiss axéniques et exogames à des doses de 0,
150, 300 et 1000 mg/kg (la dernière dose étant portée 5000 mg/kg au
cours de l'étude); elle a révélé un accroissement dans l'incidence
de l'ensemble de adénomes et carcinomes hépatocellulaires. Une
troisième étude, effectuée cette fois sur des souris NMRKf à des
doses respectives de 0, 50, 150, 300 et 1000 mg/kg de nourriture (la
dernière dose étant également portée à 5000 mg/kg au cours de
l'étude) n'a cette fois pas fait ressortir d'effets cancérogènes. Le
bénomyl ou le carbendazime ont provoqué indifféremment des tumeurs
hépatiques chez deux souches de souris (CD-1 et axéniques), souches
qui présentent toutes deux un taux élevé de tumeurs hépatiques
spontanées. En revanche, le carbendazime n'est pas cancérogène chez
les souris NMRKf dont le taux de tumeurs spontanées du foie est
faible.
Les études de cancérogénicité portant sur le bénomyl et le
carbendazime ont donné des résultats négatifs chez le rat.
1.6.8 Mécanisme de la toxicité - mode d'action
On pense que les effets biologiques du bénomyl et du
carbendazime résultent de leur interaction avec les microtubules
cellulaires. Ces structures interviennent dans des fonctions aussi
importantes que la division cellulaire, qui est inhibée par ces deux
substances. La toxicité du bénomyl et du carbendazime pour les
mammifères est donc liée à une perturbation des fonctions du système
microtubulaire.
Comme les autres dérivés du benzimidazole, le bénomyl et le
carbendazime sont plus ou moins toxiques selon les espèces. Cette
sélectivité toxicologique s'explique au moins en partie par le fait
que les deux substances ne se lient pas de la même manière aux
tubulines des espèces visées et des espèces non-visées.
1.7 Effets sur l'homme
Aucun effet nocif pour la santé humaine n'a été signalé.
1.8 Effets sur les autres êtres vivants, en laboratoire et dans leur
milieu naturel
Le carbendazime n'a guère d'effet sur l'activité microbienne du
sol aux doses d'emploi recommandées. On a cependant signalé
l'existence d'effets nocifs vis-à-vis de certains groupes de
champignons.
On a calculé que la CE50 à 72 heures, fondée sur la
croissance totale, pour les algues bleu-vert du genre Selenastrum
capricornutum, était égale à 1,3 mg/litre; la concentration sans
effets observables était de 0,5 mg/litre. La toxicité du
carbendazime pour les invertébrés aquatiques et les poissons varie
largement, les valeurs de la CL50 à 96 heures allant de 0,007
mg/litre pour des poissons chats du genre Ictalurus, à 5,5
mg/litre pour Lepomis machrochirus. Une étude de 21 jours sur la
daphnie Daphnia magna, a révélé que, à la dose de 0,025 mg/litre,
le déclenchement de la reproduction était sensiblement retardé; la
concentration sans effets observables a été fixée à 0,013 mg/litre
dans ce cas.
Le carbendazime se montre toxique pour les lombrics dans des
expériences de laboratoire reproduisant les conditions réelles
d'exposition qui résultent d'une utilisation dans les conditions
recommandées sur le terrain. Il est "relativement non toxique" pour
les abeilles et peu toxique pour les oiseaux.
2. Conclusions
Le bénomyl, un composé voisin du carbendazime, provoque une
sensibilisation cutanée chez l'homme. Ce composé et le carbendazime
lui-même ne présentent guère de risques d'intoxication aiguë pour
l'homme. Etant donné les conditions actuelles d'exposition et le
faible taux d'absorption percutanée de ces deux composés, il est
improbable qu'ils provoquent des effets toxiques généralisés dans la
population ou chez les personnes exposées pour des raisons
professionnelles. Ces conclusions résultent des données relatives à
l'animal et dans une moindre mesure des données relatives à l'homme;
elles reposent sur la connaissance du mode d'action du carbendazime
et du bénomyl, tant chez les espèces visées que chez les autres.
Grâce à une meilleure connaissance du mécanisme de la toxicité
du bénomyl et du carbendazime chez les mammifères, on pourra
peut-être mieux déterminer quelles sont les doses sans effets
observables. Des études portant sur la liaison de ces composés aux
tubulines des cellules cibles (tissus testiculaires et
embryonnaires) faciliteront sans doute les comparaisons
interspécifiques.
Le carbendazime est fortement adsorbé aux matières organiques
du sol et il y persiste pendant des périodes allant jusqu'à trois
ans. Il persiste également sur le feuillage et se retrouve par
conséquent dans les feuilles mortes. On a montré que les lombrics
pouvaient souffrir (dans leur effectif comme dans leur reproduction)
de l'action de ces composés aux doses d'emploi recommandées. On ne
possède aucune données sur les autres arthropodes qui vivent dans le
sol ou sur les débris organiques et qui pourraient être exposés de
la même manière.
Il est improbable que la forte toxicité que, selon les études
de la laboratoire, le carbendazime présente pour les organismes
aquatiques, s'observe également dans le milieu naturel, du fait de
la faible biodisponibilité des résidus de ce composé liés aux
sédiments. Toutefois on ne possède aucune donnée sur les espèce
sédimenticoles, qui seraient les plus exposées.
RESUMEN Y CONCLUSIONES
1. Resumen
1.1 Identidad, propiedades físicas y químicas y métodos analíticos
La carbendazima es una sustancia sólida cristalina de color
blanco y acción fungicida sistémica que pertenece a la familia del
bencimidazol. Su punto de fusión es de unos 250 °C y tiene una
presión de vapor de < 1 x 10-7 Pa (< 1 x 10-9 mbar). Es
prácticamente insoluble en agua (8 mg/litro) a pH 7 y 20 °C. Es un
compuesto estable en condiciones de almacenamiento normales.
Los análisis de las muestras de residuos y del medio ambiente
se realizan mediante extracción con un disolvente orgánico y
purificación del extracto obtenido utilizando un procedimiento de
reparto líquido-líquido. La valoración de los residuos se puede
realizar mediante cromatografía líquida de alto rendimiento o
inmunoensayo.
1.2 Fuentes de exposición humana y ambiental
La carbendazima es la sustancia más ampliamente utilizada de la
familia de los fungicidas bencimidazólicos. Está formulada como
dispersión acuosa, suspensión acuosa, gránulos fluidos dispersables
en agua y polvo humectable.
1.3 Transporte, distribución y transformación en el medio ambiente
En el medio ambiente, el benomilo se transforma rápidamente en
carbendazima, con una semivida de 2 y 19 h en el agua y en el suelo,
respectivamente. Por consiguiente, para la evaluación de los efectos
sobre el medio ambiente son importantes los datos obtenidos de los
estudios realizados con ambos compuestos.
La carbendazima se descompone en el medio ambiente, con una
semivida de 6 a 12 meses en el suelo desnudo, de 3 a 6 meses en el
césped y de 2 y 25 meses en el agua en condiciones aerobias y
anaerobias, respectivamente. La descomposición se debe sobre todo a
la acción de los microorganismos; el principal producto de su
degradación es el 2-aminobencimidazol (2-AB), que luego se
descompone de nuevo, también por la actividad microbiana. En la
descomposición del benomilo marcado con un grupo fenilo con 14C,
sólo el 9% del 14C formó CO2 durante un año de incubación,
mientras que el resto del 14C se recuperó sobre todo como
carbendazima y en productos unidos a residuos. El destino de un
posible producto de degradación (1,2-diaminobenceno) puede aclarar
ulteriormente la vía de degradación de los fungicidas
bencimidazólicos en el medio ambiente.
En estudios de campo y de columna se ha puesto de manifiesto
que la carbendazima queda retenida en la capa superficial del suelo.
Aunque no se dispone de datos sobre su adsorción en el suelo, es
probable que ésta sea tan intensa como la del benomilo (los valores
de Koc oscilan entre 1000 y 3600). Los valores del log Kow para
el benomilo y la carbendazima son respectivamente 1,36 y 1,49.
En la evaluación realizada en un modelo de selección, basado en
datos de adsorción y persistencia, se puso de manifiesto que no
había riesgo de lixiviación. En los Estados Unidos se han efectuado
análisis de agua de pozos que confirman esto, puesto que no se
encontraron trazas de carbendazima en ninguno de los 212 pozos
muestreados (no se dispone del límite de detección). Es de suponer
que la escorrentía superficial del benomilo y la carbendazima se
deba solamente al fungicida adsorbido en las partículas del suelo y
que en el medio acuoso estén fuertemente adsorbidos en los
sedimentos.
La carbendazima se hidroliza a 2-AB. Este es también el
metabolito primario en el suelo y las plantas.
En los sistemas animales se metaboliza a
(5-hidroxi-1H-bencimidazol-2-il)-carbamato (5-HBC) y otros
metabolitos polares, que se excretan rápidamente. No se ha observado
que la carbendazima se acumule en ningún sistema biológico.
1.4 Niveles medioambientales y exposición humana
No parece que se disponga de datos de vigilancia ambiental para
la carbendazima. Sin embargo, los estudios realizados sobre su
destino en el medio ambiente pueden resumirse como sigue.
Puesto que el benomilo y la carbendazima se mantienen estables
en las plantas durante varias semanas, pueden pasar a los organismos
que se alimentan de las hojas caídas. El suelo y los sedimentos
pueden conservar residuos de carbendazima hasta tres años. Sin
embargo, la fuerte adsorción de este compuesto en las partículas del
suelo y en los sedimentos reduce la exposición de los organismos
terrestres y acuáticos.
La principal fuente de exposición para la población humana
general se debe a los residuos de benomilo y carbendazima en los
cultivos alimentarios. En análisis de la exposición a través de los
alimentos realizados en los Estados Unidos (benomilo y carbendazima
combinados) y en los Países Bajos (con carbendazima) se obtuvo una
ingesta media prevista de alrededor del 10 por ciento de la ingesta
diaria admisible (IDA) recomendada, de 0,02 mg/kg de peso corporal
para el benomilo y de 0,01 mg/kg de peso corporal para la
carbendazima.
La exposición profesional durante el proceso de fabricación es
inferior al valor umbral límite. Se considera que los trabajadores
agrícolas que se ocupan de mezclar y cargar los plaguicidas o que
entran en campos tratados con benomilo sufren exposición cutánea a
unos mg de benomilo por hora. Esta forma de exposición se podría
reducir con algún tipo de protección. Por otra parte, puesto que la
absorción cutánea que cabe prever es baja, la probabilidad de que el
benomilo tenga efectos tóxicos sistémicos a través de esta vía es
muy escasa.
1.5 Cinética y metabolismo
La carbendazima se absorbe bien tras la exposición oral
(80-85%), pero mucho menos después de una exposición cutánea. Una
vez absorbida, se metaboliza formando numerosos compuestos en el
organismo. Los principales metabolitos son el 5-HBC y los óxidos de
5,6-HOBC-N; otros menos son importantes el 5,6-DHBC-S y el
5,6-DHBC-G.
La distribución de la carbendazima en los tejidos demuestra la
ausencia de bioconcentración. En la rata, la concentración más
elevada tras su administración oral (< 1% de la dosis) se produjo
en el hígado. Apareció carbendazima en las mitocondrias, el
metabolito 5-HBC en el citosol y el 2-AB en los microsomas. Aunque
se encontraron también carbendazima y sus metabolitos en el riñón de
gallinas y de vacas, no se detectaron niveles significativos en
otros tejidos. Tras suministrar carbendazima con el pienso a vacas
lactantes, se encontraron pequeñas cantidades de 5-HBC y 4-HBC en la
leche.
La carbendazima se excreta en la orina y las heces en un plazo
de 72 h tras la dosificación oral en ratas.
En ratas y ratones, la administración de dosis altas de
carbendazima en la dieta o con sonda afectó a ciertas enzimas de los
microsomas. Se produjo inducción de la estireno-7,8-hidrolasa y de
la epóxido-hidrolasa, mientras que se redujo la actividad de la
hidroxicumarín-O-deetilasa. También se indujo la actividad de la
glutatión-S-transferasa citosólica.
1.6 Efectos en los mamíferos de laboratorio y en sistemas de prueba
in vitro
1.6.1 Exposición única
La carbendazima tiene una toxicidad aguda baja. Los valores de
la DL50 oscilan entre > 2000 y 15 000 mg/kg en una amplia
variedad de animales de experimentación y de vías de administración.
Sin embargo, se han observado efectos reproductivos adversos
significativos en la salud tras una exposición única (véase la
sección 1.6.5).
1.6.2 Exposición de corta duración
La administración de carbendazima en la dieta durante un
período de hasta 90 días produjo efectos leves en el peso del hígado
de las ratas hembras expuestas a 360 mg/kg de peso corporal al día.
En un estudio de alimentación con sonda a ratas durante 90 días, el
NOEL fue de 16 mg/kg por día, basado en la hepatotoxicidad. Los
estudios de alimentación de corta duración en perros no se
consideraron adecuados para establecer un NOEL. En un estudio
cutáneo de 10 días en conejos no se detectó toxicidad sistémica con
la única dosis ensayada (200 mg/kg).
1.6.3 Irritación y sensibilización cutánea y ocular
La aplicación a la piel de conejos y cobayos no produjo
irritación o sensibilización cutánea. Tras la aplicación a los ojos
de conejos se observó una irritación conjuntival moderada o ligera.
1.6.4 Exposición prolongada
En ratas macho y hembra a las que se administraron 2500 mg/kg
de alimentos se registró una disminución del número de eritrocitos y
de los valores de la hemoglobina y el hematocrito. No se observó
ninguna toxicidad de tipo hepático. Las ratas macho a las que se
administraron 2500 mg/kg de alimentos o más presentaron un aumento
marginal de la atrofia testicular y la prostatitis. El NOEL en las
ratas es de 500 mg/kg de alimentos. En perros que recibieron una
dieta con 500 mg de carbendazima por kg de alimentos durante un año
o más se observó aumento del colesterol sérico, elevación de la
actividad de la fosfatasa alcalina y otras indicaciones de
hepatotoxicidad. El NOEL en perros es de 300 mg/kg de alimentos.
1.6.5 Reproducción, embriotoxicidad y teratogenicidad
La carbendazima no tuvo ningún efecto adverso en la
reproducción cuando se administró a ratas en un estudio de
reproducción de tres generaciones a dosis de hasta 500 mg/kg de
alimentos. Se registró una reducción de la fecundidad masculina en
las ratas a las que se administró carbendazima (200 mg/kg por día)
con sonda durante 85 días. Con una dosis de 50 mg/kg de peso
corporal al día en este estudio disminuyó considerablemente el
número de espermatozoides en el epidídimo.
Tras la administración a ratas de una dosis oral única de 100
mg/kg de peso corporal, el examen histológico puso de manifiesto una
interrupción temprana (0-2 días) de la espermatogénesis, con
oclusión de los conductos eferentes y un aumento del peso de los
testículos. En este estudio de dosis única no se observó ningún
efecto con 50 mg/kg. Los efectos se mantuvieron hasta transcurridos
70 días en ratas tratadas con 400 mg/kg.
La carbendazima provocó un aumento de las malformaciones y las
anomalías en las ratas administrada a dosis diarias superiores a 10
mg/kg en los días 7-16 de la gestación. En conejas se produjo un
ligero descenso de la tasa de implantación en dosis de 20 y 125
mg/kg por día administradas en los días 7-19 de la gestación, y un
aumento de la incidencia de la reabsorción a 125 mg/kg por día. Se
observó toxicidad materna con dosis de 20 y de 125 mg/kg por día en
ratas y conejos, respectivamente.
Además del descenso de la tasa de gestaciones y del aumento de
las reabsorciones precoces en la rata, se produjo una reducción
significativa del peso del feto con dosis de 20 y 90 mg/kg por día,
y un incremento significativo de las malformaciones fetales con la
dosis de 90 mg/kg por día. Estas consistieron fundamentalmente en
hidrocefalia, microftalmia, anoftalmia, malformaciones escapulares y
del esqueleto axial (fusiones vertebrales, costales y esternebrales,
exencefalia, hemivértebras e hiperplasia costal). Sin embargo, en
los conejos no aparecieron malformaciones significativas.
1.6.6 Mutagenicidad y otros efectos finales afines
Los ensayos realizados en sistemas de mamíferos y no mamíferos
in vitro e in vivo y en células somáticas, así como en células
germinales, demuestran que la carbendazima no tiene interacción con
el ADN celular, no induce mutaciones puntuales y tampoco produce
mutaciones de las células germinales.
Sin embargo, la carbendazima provoca aberraciones cromosómicas
numéricas (aneuploidía o poliploidía) en sistemas experimentales in
vitro e in vivo.
1.6.7 Carcinogenicidad
La administración de benomilo y carbendazima con los alimentos
a ratones CD-1 y suizos SPF dio lugar a un aumento de la incidencia
de tumores hepatocelulares.
Un estudio de carcinogenicidad de la carbendazima con ratones
CD-1 puso de manifiesto un aumento estadísticamente signifi cativo,
relacionado con la dosis, de la neoplasia hepatocelular en las
hembras. También se observó un aumento estadísticamente
significativo en los machos tratados con una dosis de nivel medio
(1500 mg/kg de alimentos), pero no en los que recibieron dosis
altas, a causa de la elevada mortalidad. Un estudio sobre la
carcinogenicidad de la carbendazima en una raza de ratones
genéticamente relacionada con la anterior, los ratones SPF (raza
aleatoria suiza), con dosis de 0, 150, 300 y 1000 mg/kg de alimentos
(que se aumentó a 5000 mg/kg durante el estudio), mostró un aumento
de la incidencia de adenomas y carcinomas hepatocelulares
combinados. Un tercer estudio realizado en ratones NMRKf con dosis
de 0, 50, 150, 300 y 1000 mg de carbendazima por kg de alimentos
(que se aumentó a 5000 mg/kg durante el estudio) no puso de
manifiesto efectos carcinogénicos. El benomilo y la carbendazima
provocaron tumores hepáticos en dos razas de ratones (CD-1 y SPF),
ambas con un elevado índice de tumores hepáticos espontáneos. En
cambio, la carbendazima no es carcinogénica en ratones NMRKf, que
tienen un índice bajo de este tipo de tumores espontáneos.
Los estudios de carcinogenicidad del benomilo y la carbendazima
en ratas fueron negativos.
1.6.8 Mecanismo de toxicidad, modo de acción
Los efectos biológicos del benomilo y la carbendazima se deben
a su interacción con los microtúbulos celulares. Estas estructuras
intervienen en funciones esenciales, como la división celular, que
inhiben el benomilo y la carbendazima. La toxicidad de estos
productos en los mamíferos está vinculada a una disfunción
microtubular.
El benomilo y la carbendazima, al igual que otros compuestos
del bencimidazol, tienen una toxicidad selectiva para distintas
especies, que se explica, por lo menos en parte, porque el benomilo
y la carbendazima se unen de manera distinta a los microtúbulos de
las especies específicas en las que actúan y en las que no.
1.7 Efectos en el ser humano
No se ha informado de efectos adversos para la salud humana.
1.8 Efectos en otros organismos en el laboratorio y en el medio
ambiente
Con las dosis de aplicación recomendadas, la carbendazima tiene
pocos efectos sobre la actividad microbiana del suelo. Se han
notificado algunos efectos adversos sobre ciertos tipos de hongos.
La CE50 a las 72 horas para el alga verde Selenastrum
capricornutum, basada en el crecimiento total, se calculó en 1,3
mg/litro; la NOEC fue de 0,5 mg/litro. La toxicidad de la
carbendazima para los invertebrados acuáticos y los peces varía
ampliamente, con una CL50 a las 96 h que oscila entre 0,007
mg/litro para Ictalurus punctatus y 5,5 mg/litro para Lepomis
macrochirus. En una prueba de 21 días con Daphnia magna, el
comienzo de la reproducción se retrasó considerablemente con 0,025
mg/litro; la NOEC fue de 0,013 mg/litro.
La carbendazima es tóxica para las lombrices de tierra en
experimentos de laboratorio con una exposición a concentraciones
normales y con las dosis de aplicación recomendadas en el campo. Es
"relativamente no tóxica" para las abejas de miel y su toxicidad es
baja para las aves.
2. Conclusiones
El benomilo causa sensibilización cutánea en el ser humano.
Tanto el benomilo como la carbendazima representan un riesgo muy
pequeño de intoxicación aguda. Dados los niveles de exposición
actuales y el bajo índice de absorción cutánea de estos dos
compuestos, no es probable que puedan tener efectos de toxicidad
sistémica en la población general o en personas expuestas
profesionalmente. Estas son las conclusiones que se pueden sacar de
los datos obtenidos en animales y de los limitados datos sobre el
ser humano de que se dispone, pero estas extrapolaciones están
respaldadas por el conocimiento del modo de acción de la
carbendazima y el benomilo en especies en las que actúan y en las
que no.
Una mayor clarificación del mecanismo de toxicidad de ambos
compuestos en los mamíferos permitirá quizás definir mejor los
niveles sin efectos observados. El estudio de su unión a los
microtúbulos de las células destinatarias (tejidos testicular y
embrionario) facilitará la comparación entre distintas especies.
La carbendazima se adsorbe fuertemente en la materia orgánica
del suelo, que la conserva durante un período de hasta tres años.
Persiste en la superficie de las hojas y, por consiguiente, en las
hojas caídas. Se ha demostrado que las dosis recomendadas de
aplicación afectan negativamente a las lombrices de tierra (con
efectos sobre la población y la reproducción). No se dispone de
datos acerca de sus efectos sobre otros artrópodos del suelo o de la
maleza, que estarían igualmente expuestos.
No es probable que se pueda observar en el medio ambiente la
elevada toxicidad demostrada en las pruebas de laboratorio para los
organismos acuáticos, debido a la baja biodisponibilidad de los
residuos de carbendazima unidos a los sedimentos. Sin embargo, no se
dispone de información acerca de sus efectos en las especies que
viven en los sedimentos, que sufrirían la exposición más intensa.
The metabolic fate of carbendazim in the liver was examined in
non-lactating female goats administered a feed-rate-equivalent
[phenyl(U)-14C]-carbendazim dose of 50 mg/kg once a day for 30
days. Extraction of liver homogenate from goats sacrificed 4 weeks
after initiation of dosing, i.e. when the 14C residues in the
liver had reached a plateau, indicated that the major
ethyl-acetate-extractable and identifiable radiolabelled residues in
the liver were 5-HBC (2 to 3 mg/kg) and carbendazim (approximately
0.2 mg/kg). Bound non-extractable 14C residues in the liver
reached a plateau level of approximately 1 mg/kg (Johnson, 1988).
Monson (1991) analysed, via Raney nickel desulfurization and
acid dehydration, the release and characterization of bound
carbendazim metabolites in diary cow, goat, hen and rat liver after
treatment with 14C-carbendazim. Using this technique, he was able
to show that bound 14C residue was released from the liver of cows
(76% bound before desulfurization and 36% bound after
desulfurization) and hens (58% bound before desulfurization and 19%
bound after desulfurization). The major part of the reduced residue
was identified as 5-HBC, 5,6-HOBC or carbendazim, suggesting that
the bound liver residue consisted of conjugates of
benzimidazole-related products and not natural products resulting
from breakdown and incorporation.
Benomyl and carbendazim are metabolized in fish to 5-HBC (Du
Pont, 1972).
6.4 Elimination and excretion
Carbendazim is rapidly excreted in the urine and faeces.
In a rat gavage study (Monson, 1990), [phenyl(U)-14C]-
carbendazim was administered to Sprague-Dawley rats using three
dosing regimes: a single oral dose of 50 mg/kg (low dose); a single
oral dose of 50 mg/kg following pre-conditioning gavage with
unlabelled carbendazim of 50 mg/kg for 14 days (pre-conditioned low
dose); and a single oral dose of 1000 mg/kg (high dose). Each dosing
group comprised five animals of each sex. A preliminary study
conducted with two rats of each sex, each rat having received a
single oral dose of 50 mg/kg, demonstrated that 95% of the
radioactivity excreted in the urine and faeces was recovered within
72 h after dosing and that < 1% of the dose was expired as volatile
metabolites. In the full study, > 98% of the recovered
radioactivity was excreted by the time of sacrifice (i.e. 72 h after
dosing) in the case of each group of rats. Urinary excretion
accounted for 62% to 66% of the dose in males and 54% to 62% of the
dose in low-dose and pre-conditioned low-dose females. In the
high-dose group, this pathway accounted for 41% of the dose in all
animals. Elimination of radiolabel in faeces accounted for virtually
all of the remaining radiolabel.
A lactating Holstein cow was dosed by capsule twice daily (483
mg [2-14C]-carbendazim each dose), equivalent to 50 mg/kg in the
average total daily diet, for five consecutive days, and samples of
urine, faeces and milk were collected at each dosing. Five days
after the initial dose, 65% of the radiolabel had been excreted in
the urine, 21% in the faeces and 0.4% in the milk (a total of
86.4%). Radioactive residues in the urine comprised 48% 5-HBC and 3%
polar water-soluble metabolites (Monson, 1985).
In a further study, lactating Holstein cows were dosed by the
dietary route with 0, 2, 10 or 50 mg carbendazim/kg diet for 28
days. The highest levels of carbendazim metabolites in the urine and
faeces were found in cows fed 50 mg/kg. The highest levels found in
urine were 12.56 mg 5-HBC/litre and 1.29 mg 4-HBC per litre. In the
faeces 3.81 mg 5-HBC/kg and 0.99 mg carbendazim/kg were detected
(but not in the same cow). No carbendazim residues were found in the
urine (Hughes, 1984).
Groups of hens were dosed with [2-14C]-carbendazim at a rate
equivalent to 5 and 120 mg/kg in the average total daily feed for
six consecutive days and were sacrificed 24 h after the sixth dose.
At sacrifice, an average of 95% and 92% of the radioactive doses had
been excreted in the low-dose and high-dose hens, respectively
(Monson, 1986).
6.5 Reaction with body components
In a study by Guengerich (1981), the effects of carbendazim on
hepatic enzyme induction were studied in male and female Crl-CD
rats. The treatment groups included animals fed for 28 days with
diets that contained carbendazim at concentrations of 0, 10, 30,
100, 300, 1000 or 3000 mg/kg. Microsomal epoxide hydrolase and
cytosolic glutathione- S-transferase were monitored in subcellular
fractions isolated from the livers of animals in each treatment
group. Liver weights were also recorded. Elevated liver weights were
observed at 1000 and 3000 mg carbendazim/kg in both male and female
rats. No apparent liver toxicity or effect on body weight was
observed. Carbendazim induced epoxide hydrolase in both sexes of
rats and mice at 1000 and 3000 mg/kg. Induction of
glutathione- S-transferase was observed at 3000 mg/kg. In general,
the level of induction seemed to be slightly greater in females than
males.
In the same study, but in a separate test, CD-1 male mice were
treated via gavage with carbendazim suspended in corn oil (0, 100
and 1000 mg/kg per day) for 5 days. The liver samples were
homogenized and subcellular fractions were prepared as above. Wet
liver weights, microsomal cytochrome P-450, NADPH-cytochrome-c
reductase, styrene-7,8-hydrolase, benzphetamine- N-demethylase,
benzo(a)pyrene hydroxylase and 7-ethoxy coumarin O-deethylase, and
cytosolic glutathione- S-transferase were measured. Those
parameters showing statistically significant increases over the
control values were styrene-7,8-hydrolase and glutathione- S-
transferase; 7-ethoxycoumarin O-deethylase showed a significant
decrease. It is noteworthy that the total microsomal cytochrome
P-450 level did not increase, indicating that a whole scale
microsomal induction phenomena was not induced by carbendazim even
at the higher treatment level. However, as shown by the increase in
microsomal styrene-7,8-hydrolase, some hepatic microsomal enzymes
are induced by in vivo carbendazim treatment (Guengerich, 1981).
There did not appear to be any substantial difference in enzyme
induction between rats and mice.
7. EFFECTS ON LABORATORY MAMMALS AND IN VITRO TEST SYSTEMS
7.1 Single exposure
The acute toxicity of carbendazim in several animal species is
summarized in Table 8. The LD50 values range from > 2000 to > 15
000 mg/kg for a wide variety of test animals and routes of
administration. A description of toxic effects is given in section
7.5.1.
7.2 Short-term exposure
7.2.1 Gavage
Groups of ChR-CD male rats (six per dose level) were gavaged
with 200, 3400 and 5000 mg carbendazim/kg per day, five times/week,
for two weeks. Two out of six rats died at the dose level of 3400
mg/kg per day. At all dose levels, gross and microscopic evidence of
adverse effects on testes and reduction or absence of sperm in the
epididymides was seen. Testes were small and discoloured, with
tubular degeneration and evidence of aspermatogenesis. At the dose
level of 3400 mg/kg per day, there were also morphological changes
in the duodenum (oedema and focal necrosis), bone marrow (reduction
in the blood-forming elements) and liver (decrease in the large
globular-shaped vacuoles) (Sherman, 1965; Sherman & Krauss, 1966).
Subchronic administration of 0, 16, 32 or 64 mg carbendazim per
kg per day by gavage for 90 days to four groups of 10 male and 10
female litter-mate weaning Wistar rats was carried out. The
erythrocyte counts for treated rats were lower than those of the
controls after 15 days of exposure. However, no clear dose-response
relationship was demonstrated after 30, 60 or 90 days of exposure. A
decrease was noted in leucocyte counts at 15 days. After 30 and 60
days, both sexes demonstrated transient decreases in lymphocyte
counts compared to controls. However, no clear dose-response
relationship was observed among the treated groups. No change was
noted in the activity of whole blood cholinesterase. Male rats
showed significantly increased alkaline phosphatase activity at a
dose level of 64 mg/kg per day. Blood urea levels were lower in
males at dose levels of 32 and 64 mg/kg per day after 90 days.
Increased serum bilirubin concentrations were observed in males and
females at 32 and 64 mg/kg per day and were attributable to
parenchymal cell damage, as shown by increased glutamic-pyruvic
transaminase activity. Dose-related changes in the liver ranged from
sparse infiltration by inflammatory cells to inflammatory and
degenerative changes. Tubular dilation and hydropic degeneration
were noted in the kidneys of the low-dose rats, and fibrosis and
congestion in the medium- and high-dose rats. Increased lung weight
was correlated with bronchopneumonic changes. Slight changes in
weights were reported for several organs (Janardhan et al., 1987).
The published information was difficult to evaluate due to the
variability of the results and absence of the raw data.
Table 8. Acute toxicity of carbendazim in animals
Chemical Species Sex Animals Route Vehicle Concentrationa Reference
per group (mg/kg body weight)
Carbendazim rat M/F 10 oral sesame oil LD50 > 15 000 Kramer & Weigand (1971)
(MBC) rat M 10 intraperitoneal 0.9% saline LD50 > 2000 Goodman & Sherman (1974)
and Tween 80
rat M 6 inhalation (1 h) dust ALC > 5.9 mg/litreb Sarver (1975)
rat F 5 dermal sesame oil ALD > 2000 Kramer & Weigand (1971)
mouse M 10 oral propylene glycol LD50 > 15 000 Til & Beems (1981)
mouse M/F 10 intraperitoneal sesame oil LD50 > 15 000 Scholz & Weigand (1972)
dog M/F 2 oral sesame oil ALD50 > 5000 Scholz & Weigand (1972)
guinea-pig M 10 oral corn oil LD50 > 5000 Dashiell (1975)
rabbit M 10 dermal aqueous paste LD50 > 10 000 Edwards (1974a)
75% wettable rat M/F 5 oral corn oil LD50 > 5000 Hinckle (1981)
powder
rat M/F 10 inhalation dust LC50 > 5 mg/litre Nash & Ferenz (1982)
rabbit M/F 5 dermal physiological saline LD50 > 2000 Ford (1982)
Benlate C rat M/F 5 oral water LD50 > 5000 Grandizio & Sarver (1987)
(50% wettable rabbit M/F 5 dermal aqueous paste LD50 > 2000 Vick & Brock (1987b)
powder)
a ALC = approximate lethal concentration; ALD = approximate lethal dose
b Time-weighted concentration
7.2.2 Feeding
7.2.2.1 Rat
Groups of ChR-CD rats (16 males and 16 females per group) were
fed carbendazim (72% a.i.) in the diet for 90 days at levels of 0,
100, 500 and 2500 mg/kg. The animals were observed daily for
behavioural changes and body weight, and food consumption were
recorded at weekly intervals. Haematological examinations were
conducted on 10 male and 10 female rats in each group at 30, 60 and
90 days. Routine urinalyses were performed on the same animals, and
plasma alkaline phosphatase and glutamic-pyruvic transaminase levels
were determined. After 90 to 98 days of continuous feeding, 10 male
and 10 female rats in each group were killed and selected organs
weighed. These and other organs were preserved for microscopic
examination. The six male and six female rats remaining in each
group after terminal sacrifice were used in a reproduction study
(see section 7.5.1). There were no signs of poisoning and no
compound-related effects on weight gain, food consumption or
haematological parameters. There were no control data for
biochemical determinations, urinalysis or differential white blood
counts. The average dose for the high-dose animals was 360 mg/kg per
day initially and 123-152 mg/kg per day at sacrifice. The
liver-to-body-weight ratio in females fed 2500 mg/kg diet was
slightly increased compared with control rats. There were no effects
on testicular weights in any of the treatment groups. Microscopic
examination of selected tissues and organs in the control and
high-dose groups demonstrated no adverse effects attributable to
carbendazim (Sherman, 1968).
7.2.2.2 Dog
Groups of one-year-old beagles (four males and four females per
group) were administered carbendazim (53% a.i.) in the diet for
three months at dietary levels of 0, 100, 500 and 2500 mg/kg. The
highest level was reduced to 1500 mg/kg because of reduced food
intake and decreased body weight. However, compound administration
was interrupted when animals were fed a control diet for a few days
and then fed with 1500 mg/kg again.
Food consumption and body weight data were recorded weekly, and
clinical laboratory examinations (including haematological,
biochemical and urinalysis measurements) were performed pre-test and
after 1, 2 and 3 months of feeding. At the end of the study all
animals were killed, selected organs were weighed, and these and
other organs were subjected to gross and microscopic evaluations.
No mortality or adverse clinical signs were observed over the
course of the study, and growth and food consumption were normal
(except at 1500-2500 mg/kg). Urinalysis measurements were unaffected
by treatment, and there were no dose-related effects on the
haematological values. Females at the mid-dose level showed a trend
toward increased cholesterol levels at 1, 2 and 3 months compared
with the pre-test and control values. High-dose females had
similarly elevated cholesterol levels. Organ-to-body weight changes
were observed in the case of the thymus of low- and mid-dose males
and the prostate of mid-dose males. All the weights for these organs
were increased compared with control values. However, only the
liver, kidney and testes were examined histologically in the low-
and mid-dose groups. Limited histopathological data did not indicate
compound-related effects (Sherman, 1970).
In a study by Til et al. (1972), groups of beagles (four males
and four females per group) were administered carbendazim in the
diet at levels of 0, 100, 300 and 1000 mg/kg for 13 weeks. The
highest level was increased to 2000 mg/kg after six weeks of
treatment. Body weight, haematological and blood chemistry
measurements, urinalyses and liver/kidney function tests were
performed periodically. Gross and microscopic examinations of all
animals were performed at the end of the study. There were no
reported compound-related effects on clinical behaviour, body
weight, food consumption, haematological parameters, kidney function
(phenol red excretion) or liver function (BSP retention)
examinations. Blood chemistry measurements were normal, except for a
slight decrease in albumin in mid- and high-dose males at 12 weeks.
These values differed from week 0 measurements only in high-dose
males. Urinalysis values were normal except for a high bacterial
count in high-dose females at week 13. The blood clotting time was
slightly reduced in high-dose dogs at week 12. There were slight
increases in relative liver and thyroid weights and a decrease in
relative heart weights in the highest-dose group compared with
controls. No microscopic changes that could be associated with
treatment were observed in these or any other organs. Carbendazim
appeared to be without adverse effects on beagles when incorporated
in the diet for 13 weeks at dietary levels of 300 mg/kg or less.
7.2.3 Dermal
New Zealand albino rabbits (six males/group) were treated with
0 or 2000 mg/kg of carbendazim, applied as a 50% aqueous paste to
shaved intact dorsal skin. The material was applied repeatedly, 6
h/day, for ten consecutive days. There were no adverse effects on
body weight, clinical symptoms, organ weights, gross pathology or
histopathology of selected organs. However, there was focal necrosis
of the epidermis and polymorphonuclear cell infiltration of the
dermis in five out of six exposed rabbits. No other effects were
observed (Dashiell, 1975).
7.3 Skin and eye irritation; sensitization
7.3.1 Dermal
The primary dermal irritation potential of Benlate C (50%
wettable powder) was evaluated by applying a 5-g aliquot for 4 h to
the clipped intact skin of six New Zealand White rabbits. Test sites
were evaluated for erythema, oedema and other evidence of dermal
effects, and were scored according to the Draize scale at 4, 24, 48
and 72 h after application. No dermal irritation was seen at any
time during the study (Vick & Brock, 1987a).
7.3.2 Eye
No eye irritation potential for technical carbendazim was seen
in six albino rabbits (Edwards, 1974b). In a study by Vick &
Valentine (1987), a 50% wettable powder (formulation) was evaluated
for acute eye irritation potential in six male New Zealand white
rabbits. The formulation produced slight corneal opacity, mild or
moderate conjunctival redness, and slight or mild conjunctival
oedema in all the rabbits. In addition, there was moderate iritis in
three of the rabbits and minimal blood-tinged discharge in one
rabbit. Microscopic examinations revealed no corneal injury in any
of the treated eyes. The treated eyes of the two other rabbits were
normal by 72 h. It was concluded that this formulation was a
moderate eye irritant.
7.3.3 Sensitization
Albino guinea-pigs (10 males) exposed to carbendazim, either
technical material or a 75% wettable powder formulation, presented
no evidence of dermal sensitization following either intradermal
injections or repeat applications to shaved intact skin (Ford,
1981).
A 50% carbendazim formulation was tested on the shaved intact
skin of 10 male and 10 female Duncan Hartley albino guinea-pigs.
Five male and five female guinea-pigs were treated with 80% ethanol
(in water) and served as vehicle control animals. Two male and two
female guinea-pigs were treated with the test material (as a solid)
at the challenge phase only and served as negative control animals.
1-Chloro-2,4-dinitrobenzene (DNCB) was tested as a 0.3% suspension
in 80% ethanol (in water) on the shaved intact skin of two male and
two female guinea-pigs as a positive control group. No irritation
was observed in the test or vehicle control guinea-pigs or in the
negative controls at the challenge phase. DNCB produced
sensitization in all treated animals (Martin et al., 1987).
7.4 Long-term exposure
7.4.1 Rat
Groups of weanling rats (36 male and 36 female ChR-CD albino
rats/group) were administered carbendazim (50-70% a.i.) in the diet
for 104 weeks at levels of 0, 100, 500, 2500 (increased to 10 000
mg/kg after 20 weeks) and 5000 mg/kg diet. Body weight and food
consumption were recorded weekly for the first year and twice a
month thereafter. Daily observations were made with respect to
behavioural changes and mortality. At periodic intervals throughout
the study, haematological, urinalysis and selected clinical
chemistry examinations were performed. After one year each group was
reduced to 30 male and 30 female rats by interim sacrifice for gross
and microscopic examinations. At the end of the study all surviving
animals were sacrificed and gross examination of tissues and organs
was made. Microscopic examinations were conducted on all tissues and
organs from the control and 2500-mg/kg groups, the livers of the
100- and 500-mg/kg groups, and the livers, kidneys, testes and bone
marrow from the 5000-mg/kg groups. Survival decreased during the
second year to approximately 50% for males and 39% for females in
all groups. Body weight gain was depressed for males and females in
the 2500-mg/kg group and for females in the 5000-mg/kg group,
compared to control groups. Food consumption did not differ among
the various groups. The average daily dose for the 500-mg/kg group
was 65 mg/kg body weight per day initially, 18 mg/kg body weight per
day at one year, and 15 mg/kg body weight per day at two years.
Haematological examinations demonstrated reduced erythrocyte count
and haemoglobin and haematocrit values for females at 9-24 months in
the 2500- and 5000-mg/kg groups, and for males at 24 months in the
2500-mg/kg group. There were no compound-related clinical
manifestations of toxicity and no effects observed in urinalysis
examination. Alkaline phosphatase and glutamic-pyruvic transaminase
activity varied throughout the test at 2500 and 5000 mg/kg but did
not demonstrate a consistent dose-response relationship. There were
no apparent differences in the organ weights or organ-to-body weight
measurements, except in the case of female livers in the 2500- and
5000-mg/kg groups. This increase in the liver-to-body weight ratio
was due to a reduction in body weight. Histopathological examination
of the livers did not demonstrate any compound-related effects.
Males in the 2500-mg/kg group presented a marginal increase in
diffuse testicular atrophy and prostatitis (Sherman, 1972).
In another 2-year rat study, groups of Wistar rats (60 males
and 60 females/group) were administered carbendazim (99% pure) in
the diet at levels of 0, 150, 300 and 2000 mg/kg diet for two years.
The dose of 2000 mg/kg was increased to 5000 mg/kg after one week
and then to 10 000 mg/kg after two weeks for the remainder of the
study. Animals were examined daily for clinical signs of toxicity.
Body weight and food consumption were measured regularly throughout
the study. Haematological (peripheral blood), blood chemistry
(orbital sinus) and urinalysis evaluations were conducted
periodically during the study. All animals were subjected to
complete gross necropsy, and selected organs were weighed. Tissues
were examined microscopically in 20 male and 20 female rats in the
control and high-dose groups. All tumours and gross abnormalities
were also examined histologically. There were no differences between
test groups and control animals concerning clinical signs of
toxicity or food consumption. Body weights were significantly
reduced in low-dose males from week 88 to term and in high-dose
females from week 12 to term. Urinalyses were comparable among all
groups. Of the haematological parameters examined, haemoglobin was
depressed in high-dose females at weeks 26, 52 and 103 and
haematocrit was depressed in high-dose females at week 103. There
were no compound-related effects in males. Serum glutamic
oxaloacetic transaminase (SGOT) activity was decreased in high-dose
males at term, but not in females. High-dose females had increased
serum glutamic pyruvic transaminase (SGPT) activity and decreased
total serum protein at study termination. There were no
compound-related effects on organ weights except for increased
relative liver weights in high-dose females. There were also no
compound-related effects on mortality, this being 50% at week 76 in
control males and at week 92 in treated males. There was 50%
mortality in control and low-dose females at week 88 and in mid- and
high-dose females at 92-96 weeks. Survival at termination of the
study was similar in all groups.
There were no histological differences between control and
treated groups except for an increased incidence of diffuse
proliferation of parafollicular cells of the thyroid in the
high-dose females (Til et al., 1976a).
7.4.2 Dog
In a one-year study, beagle dogs (five of each sex per group)
were fed diets containing 0, 100, 200 or 500 mg carbendazim/kg. The
dogs were weighed at regular intervals, and individual food
consumption was monitored throughout the study. Clinical pathology
evaluations were performed twice prior to the initiation of the
study and five times during the study, at 1, 3, 6, 9 and 12 months.
After one year, all dogs were killed and selected tissues were
examined microscopically. There were no statistical differences in
mean body weight that could be attributed to carbendazim exposure.
The mean daily food consumption in all treated groups of dogs was
similar to controls. None of the clinical observations were
attributable to carbendazim intake. Dogs fed 500 mg/kg had elevated
levels of serum cholesterol. These levels were statistically
significant for the males at 9 months and the females at 1 and 2
months. There were no compound-related microscopic lesions related
to carbendazim intake (Stadler, 1986).
Groups of beagles (four males and four females per group) were
administered carbendazim (53% a.i.) in the diet at dosage levels of
0, 100, 500 and 2500 mg/kg for two years. The dogs were 1-2 years of
age at the start of the test. Food consumption and body weight data
were obtained weekly and animals were examined daily for clinical
signs of toxicity. Haematological, biochemical and urinalysis
examinations were performed periodically throughout the study.
Interim sacrifice after one year was performed on one male and one
female from the control and 500-mg/kg groups. Organ weight, gross
necropsy and histopathological examinations were performed at the
end of the study. Only the livers and testes were examined
histologically in the 100- and 500-mg/kg groups. No mortality was
reported for the control or the 100- and 500-mg/kg dose groups. The
average daily intake for the 500-mg/kg dose group was 15.0-20 mg/kg
body weight per day initially, 14-18 mg/kg body weight per day after
one year and 10-16 mg/kg body weight per day after two years.
Haematological and urinalysis values were unaffected by treatment.
The dogs in the 500-mg/kg groups had increased levels of
cholesterol, blood urea nitrogen (BUN), total protein and SGPT.
Swollen vacuolated hepatic cells and marginal proliferation of the
portal triads with cellular infiltration was observed in one dog
sacrificed after one year, which had been fed 500 mg/kg. No
histopathological liver lesions were observed in animals fed 500
mg/kg diet at the end of the study. Although inflammatory and
fibrotic liver changes were observed in the 2500-mg/kg group, these
changes cannot be evaluated because of the uncertainty of the dosing
regime, of the time of exposure and of the number of dogs. There
were no noticeable effects on organ weights or on organ-to-body
weight ratios. Diffuse testicular atrophy and aspermatogenesis were
observed in males (two out of four) at 100 mg/kg but not at 500
mg/kg (Sherman, 1972).
In another 2-year dog study, groups of beagles (four males and
four females per group) were fed technical carbendazim in the diet
at dosage levels of 0, 150, 300 and 2000 mg/kg for 104 weeks. After
33 weeks the dose 2000 mg/kg was increased to 5000 mg/kg. The dogs
were 22-27 weeks old at the start of the study. Daily examinations
were made for clinical signs of poisoning or adverse behaviour. Body
weight and food consumption data were recorded regularly throughout
the study. At periodic intervals (weeks 13, 26, 52, 78 and 104),
haematological, blood chemistry and urinalysis measurements were
made. Liver function (BSP retention) and kidney function (phenol red
excretion) tests were conducted at weeks 26, 52 and 104. At the
conclusion of the 104 weeks of dietary administration, each dog was
sacrificed and gross and microscopic examination of tissues and
organs was performed. There was no mortality in any group except for
one female in the high-dose group which was killed in a moribund
state after week 36. Body weight was decreased in mid-dose males and
high-dose males and females. Food consumption was comparable among
all groups. Blood clotting times were significantly reduced in
high-dose males from week 13 to term, and slight decreases were
noted in high-dose females. Serum alkaline phosphatase activity was
increased in the high-dose group throughout the study. There were no
compound-related effects on SGPT or SGOT levels. All other
haematological and blood chemistry values were comparable with
control groups. There were no differences for BSP retention, phenol
red excretion or urine analysis values among the various groups.
Absolute liver and thyroid weights were significantly increased in
high-dose dogs. Relative liver, thyroid and pituitary weights were
also significantly increased at the high-dose level. There were no
reported microscopic changes in these organs related to treatment.
There was an increased incidence of prostatitis (3/4 versus 1/4) in
high-dose males compared with controls. Also noted in high-dose
males (1/4) was interstitial mononuclear inflammatory cell
infiltrates and atrophic tubules of the testes. It was concluded
that the feeding of carbendazim in the diet to dogs for two years
was without apparent adverse effects at levels up to and including
300 mg/kg (Reuzel et al., 1976).
7.4.3 Mouse
A description of studies on mice is given in section 7.7.
7.5 Reproduction, embryotoxicity and teratogenicity
7.5.1 Reproduction
Groups of ChR-CD rats (3 male and 16 female rats per group,
except that the high-dose group contained 20 females) were fed
carbendazim in the diet at dose levels of 0, 100, 500, 5000 and 10
000 mg/kg and subjected to a standard two-litter-per-generation,
three-generation reproduction study. The parental animals were fed
the experimental diet at 21 days of age and mated to produce the
F1 litter at 100 days of age. The number of matings, pregnancies
and number of young in each litter at birth was recorded. The
litters were culled to 10 pups/litter on day 4. The number of the
live pups was recorded on days 4, 12 and 21, as was pup weight at
weaning. The parents were mated again to produce the F1B litters.
The F1B litters were maintained on the respective diets for 110
days and then mated to produce the F2A and F2B litters. The
F3A and F3B litters were produced similarly. Gross and
histopathological examinations of selected tissues and organs were
performed on two males and two females in each of five F3litters
from the control, 5000- and 10 000-mg/kg groups. Reproduction
indices, including mating, fecundity, fertility, gestation,
viability and lactation, were calculated and compared with control
values. Carbendazim was without effect on fertility, gestation,
viability and lactation. However, the average litter weights at
weaning were reduced in all generations fed 5000 and 10 000 mg/kg.
Histopathological examination of F3B weanlings did not reveal any
effects that were considered compound-related (Sherman, 1972).
Two additional feeding studies (Sherman, 1968; Koeter et al.,
1976) were reviewed by the 1983 Joint FAO/WHO Meeting on Pesticide
Residues (JMPR), which concluded that both reports had limitations.
For the Sherman (1968) study, it was stated that the "data presented
were extremely limited and submitted as group data only. There were
no pregnancies at 100 mg/kg diet for either F1a or F1b matings.
There were no apparent effects on the reproduction indices on
weanling weights. However, the fertility index for all groups, which
was 33-67% prevents any meaningful interpretation of the data"
(FAO/WHO, 1985b). For the study of Koeter et al. (1976), the 1983
JMPR concluded that "although there were no apparent adverse effects
on reproduction and no teratogenic effects at dietary levels of
carbendazim up to and including 2000 mg/kg, there were no individual
animal data presented. Histopathology of animals in the four-week
study was incomplete and did not include evaluations of spleen or
ovaries. Such additional data are needed to confirm the absence of
adverse effects in this three-generation reproduction study in rats"
(FAO/WHO, 1985b).
A serial breeding technique was used to evaluate the fertility
of male Sprague-Dawley rats after exposure by gavage to 10 daily
doses of carbendazim (400 mg/kg per day). Proven-fertile males (90
days old) were bred with a new female each week. Breeding began on
the third day of treatment and continued for 32 weeks after the last
day of chemical exposure. Twelve days after each breeding period,
the females were killed, their uteri were examined for resorptions,
and the number of dead and viable fetuses was determined. All males
were killed 35 weeks after the exposure, and testicular tissue was
prepared for histopathological examination by vascular perfusion.
The fertility (as indicated by the number of pregnant females) of
males in the carbendazim-treated group was depressed during the
first post-exposure week; 10 of the 24 treated males failed to
induce a pregnancy, as compared with no failure in the control
group. By the fifth post-exposure week, 16 of the 24
carbendazim-treated males were infertile. Of these 16 males, 4
recovered fertility after being infertile for 5-11 consecutive
breeding periods. However, 12 of the males did not recover fertility
during the remainder of the 32 week post-exposure period.
Histological examinations of testicular sections 245 days after the
exposure revealed that the exposure to carbendazim had caused severe
seminiferous tubular atrophy (> 85% of tubules were atrophic) in
those treated males that failed to recover fertility. The
seminiferous tubules of these males often showed "Sertoli cell only"
syndrome epithelium surrounded by a thickened basement membrane.
Less than 2% of the tubules contained spermatozoa in the lumen. The
seminiferous tubules of the carbendazim-treated males that recovered
fertility had various numbers of atrophic tubules (13-85%) 245 days
after the exposure (Carter et al., 1987).
In another study, groups of male and female rats (8-12 of each
sex per group) were administered 0, 50, 100, 200 or 400 mg
carbendazim/kg per day by gavage from weaning through puberty,
gestation and lactation, and they were mated at 84 days of age. The
male rats were killed on day 104-106 while the female rats were
killed on day 27 postpartum. A similar study was conducted with
Syrian hamsters administered 0 or 400 mg/kg per day. In the parental
generation, various landmarks of puberty were measured. In females,
estrous cyclicity, litter size, the number of implants, organ
weights and histological status were assessed. In males, organ
weights, testicular and epididymal sperm counts, sperm motility,
sperm morphology, testicular histological status, and endocrine
parameters were assessed. In addition, the growth, viability and
reproductive function of the offspring (F1) were observed during a
4-month period of continuous breeding. Males were killed at 5 months
for histopathological investigation. In the parental generation of
both species, carbendazim did not alter pubertal development, growth
or viability. The reproductive potential of rats treated with
carbendazim with 200 and 400 mg/kg per day was reduced due to
effects on sperm production and fetal viability. In the male rat,
carbendazim treatment with 200 or 400 mg/kg per day markedly altered
sperm morphology, testicular and epididymal weights, sperm numbers
and testicular histology. Fertility, sperm motility and hormonal
levels were altered primarily in those males fed 200 or 400 mg/kg
per day that exhibited very low sperm counts. A statistically
significant reduction in caudal epididymal sperm count was noted at
dose levels of 50 mg/kg per day or more. Testicular and epididymal
sperm counts in male hamsters were significantly lower (about 21%)
in the carbendazim-treated males compared with the control males. In
the F1 male hamsters, testis and seminal vesicle weights and
epididymal sperm counts were significantly reduced by prenatal
exposure to carbendazim at 400 mg/kg per day. In the parental female
rats, carbendazim administration caused postimplantation losses at
400 mg/kg per day and a few malformed rat pups were found in litters
at 100 and 200 mg/kg per day. Litter size was significantly reduced
at 200 and 400 mg/kg per day. Overall, carbendazim was less toxic to
the hamster than to the rat (Gray et al., 1988, 1990).
Since spermatogenesis is an androgen-dependent process, the
effects of carbendazim (0-400 mg/kg per day by gavage) on the
endocrine function of the rat testes were investigated. Following
subchronic (85 day) exposure, serum hormone levels, including those
of pituitary luteinizing hormone (LH), follicle-stimulating hormone
(FSH), thyroid-stimulating hormone (TSH), prolactin (PRL), and
androgen-binding protein (ABP) and testosterone in testicular fluids
(interstitial fluid and seminiferous tubule fluid) were measured. In
addition, the functional capacity of the Leydig cell to secrete
testosterone was assessed in vitro following human chorionic
gonadotrophin (HCG) challenge. Subchronic treatment with carbendazim
at doses of 50-100 mg/kg per day had no effect on pituitary or
testicular hormone concentrations; dosing with 200 mg/kg per day
elevated the testosterone concentration in the seminiferous tubule
fluid and the ABP tubule fluid without affecting serum testosterone
or ABP concentrations. The dose of 400 mg/kg per day resulted in
increased concentrations of both testosterone and ABP in the
interstitial fluid and seminiferous tubule fluid and of serum ABP,
but no change in serum testosterone level. These hormonal changes
are consistent with carbendazim effects on the gonads and resultant
testicular atrophy. This endocrine profile is similar to the
"Sertoli cell only" syndrome discussed by de Krester (1977). Thus,
the elevated seminiferous tubule fluid testosterone concentrations
may be a result of two factors: (a) greater release of testosterone
by the Leydig cells into the interstitial fluid; and/or (b)
decreased testosterone outflow from the testis into the general
circulation. In addition, increased ABP in the interstitial fluid
may reflect a change in the relative secretion of ABP into the
interstitial fluid and the seminiferous tubules (Rehnberg et al.,
1989).
Since it is possible that extragonadal changes contributed to
the appearance of the altered testicular endocrine profile described
above, a further study focused on the presence of concurrent changes
in the hypothalamus and pituitary control of the testes. In this
study, rats were gavaged with carbendazim (50, 100, 200 or 400 mg/kg
per day) for 85 days (Gray et al., 1988). A dose-related increase in
serum FSH and LH was noted, but values for prolactin and
thyroid-stimulating hormone remained unchanged. No statistical
differences in gonadotropin-releasing hormone (GnRH) concentrations
were present in the mediobasal hypothalamus, although an elevation
in anterior hypothalamic GnRH values was found at the 50-mg/kg dose,
followed by a dose-related decline. These findings would suggest
that carbendazim-induced testicular damage is accompanied by
compensatory changes in the hypothalmic and pituitary regulation of
the testes (Goldman et al., 1989).
The above studies imply that carbendazim acts directly on the
testes to produce a number of hormonal and pathological changes.
Consistent with this hypothesis are the results of the study of
Nakai et al. (1992), in which the effects of carbendazim on the
testes, efferent ductules and spermatozoa were determined after a
single oral dose in male Sprague-Dawley rats (Charles Rivers). In
the first experiment, groups of 86-day-old rats were treated with 0
or 400 mg carbendazim/kg and killed 2, 4 or 8 h later on the same
day or 1, 4, 8, 16 or 32 days after dosing. The effect of
carbendazim was first noted at 8 h as an increase in testicular
weight. Testis weight continued to increase until day 4 and declined
thereafter. On days 16 and 32, testes weights were substantially
lower than those of controls in 5 out of 16 animals, indicating the
variable response of individual animals. A decrease in the
percentage of sonication-resistant sperm heads per testis occurred
at 8 h (4 out of 8 rats affected), but this decrease was not
significant until 24 h, when a mean decrease of 19% was observed.
Maximum decreases in total sperm head counts per testis occurred on
day 8, after which some recovery was apparent. Epididymal weights
were increased on day 4. The percentage of morphologi cally normal
sperm in the cauda epididymus was less on day 4. By day 8, many
spermatozoa heads were separated from their flagella and 10% of the
heads were misshapen. Numerous sloughed round germ cells and
cytoplasmic testicular debris were also evident. No effect on the
percentage of motile sperm was seen at 2, 4 and 8 h or at 1 and 4
days post-treatment. Sperm motility was significantly decreased on
days 8 and 16. Because of the clumping and degeneration of the
spermatozoa, quantitative estimates of the percentage motility could
not be determined on days 8 and 16. Similarly damaged sperm were
seen in 3 out of 8 rats on day 32, although the percentage of motile
sperm that could be measured was similar to that of controls.
In a second experiment by Nakai et al. (1992), groups of rats
(between 97 and 105 days of age) were treated with a single oral
dose of 0, 50, 100, 200, 400 or 800 mg/kg and killed on day 2 or 70
post-treatment. On day 2, a dose-dependent increase in testicular
weight was seen at dose levels 100 mg/kg or more. This was
accompanied by significant increases in mean seminiferous tubular
diameter at 400 and 800 mg/kg. At 50 mg/kg, missing immature germ
cells were noted with round spermatids from stages I and II and
elongated spermatids sloughed from stage VII epithelium. At 100
mg/kg, the disappearance of germ cells was more severe and sloughing
of elongated spermatids extended into stages XII and XIV. A detailed
description of the spermatogenic cycle may be found in Russell et
al. (1990). At doses higher than 100 mg/kg, missing germ cells
extended into all stages except stages IX-XI, while, at doses of
400-800 mg/kg, some seminifer ous epithelia were damaged so severely
that it was difficult to identify the stage. In addition, major
pathological changes were seen in the excurrent ducts of the testis.
The rete testis was swollen with sloughed germ cells indicating that
ductal blockage had occurred further down the tract. Such occlusions
were observed in the efferent ductules of animals treated with 100
mg/kg or more. On day 70, mean testis weight and mean seminiferous
tubule diameter showed a dose-dependent decrease. Histologically,
these decreases were associated with a dose-dependent increase in
seminiferous tubular atrophy. No atrophic tubules were seen in the
control rats. The atrophied tubules contained primarily Sertoli
cells and occasional spermatogonia, and were surrounded by a
thickened basement membrane. Pathological alterations were also seen
in the efferent ductules of the treated animals, 50% or more of the
ducts being occluded in rats dosed with 100 mg or more. The
occlusions were characterized as compacted luminal contents,
spermatic granulomas, mineralizations and obliteration of the
original lumen by fibrotic connective tissue (Nakai et al., 1992).
In a further study, carbendazim was administered to female
Holtzmann rats (8 rats per group) by gavage (0, 25, 50, 100, 200,
400 or 1000 mg/kg per day) during early pregnancy (days 1 to 8). A
range of maternal parameters, including the number of implantation
sites, body weight gain, uterine weight, implantation site size, and
serum ovarian and pituitary hormones, was assessed following
sacrifice at day 9. At dosages up to 400 mg/kg per day, carbendazim
had no significant effect on any of the measured parameters but a
trend towards increased resorptions was evident. The highest dosage
produced reductions in maternal body weight gain, implantation site
weight and serum LH, and an increase in serum estradiol (Cummings et
al., 1990).
Female hamsters were treated with a single gavage dose of
carbendazim to identify effects of this compound during the
perifertilization period. The exposure times were selected to
coincide with either of the two microtubule-dependent events
initiated by the ovulatory surge of LH, i.e. oocyte maturation
(first meiotic division, occurring late on vaginal proestrous) or
fertilization (second meiotic division, occurring early on vaginal
estrous). In the first experiment, 0, 250, 500, 750 or 1000 mg
carbendazim per kg (10 hamsters per group) was administered during
meiosis I. Pregnancy outcome was assessed on day 15. The percentage
of pregnant hamsters was significantly reduced at 750 and 1000
mg/kg. In those animals that became pregnant, the average number of
live pups was reduced at all dose rates. In a second experiment,
female hamsters (10 hamsters per group) were bred overnight and
administered a single dose of 0 or 1000 mg/kg during meiosis II on
the morning following breeding. The percentage of pregnant hamsters
was unaffected, but the average number of live pups (measured at 15
days) was reduced. These results show that carbendazim
administration by gavage at microtubule-dependent meiotic events can
result in early pregnancy loss in hamsters (Perreault et al., 1992).
In a separate study, pseudopregnant rats (induced by
stimulation of the uterine cervix with a small brass rod on
proestrous and estrous) were administered 0 or 400 mg carbendazim/kg
per day during days 1-8. On day 4 of pseudopregnancy, a uterine
decidual cell response was induced and the females were killed on
day 9. The decidual cell response, evaluated as a measure of uterine
competency, was significantly less in the treated rats than in the
controls (Cummings et al., 1990).
Groups of male mice (12 per group) were administered
carbendazim (0, 250, 500 or 1000 mg/kg per day) by gavage for 5
consecutive days. Body weights, testis weights and sperm parameters
were measured at 7, 24 and 39 days post-treatment. Body weights were
unaffected. Testis weights were reduced only in the highest-dose
group at 7 and 24 days but had recovered by 39 days. Flow cytometry
measurements of testicular cells showed that the relative
percentages of certain testicular populations (round, elongating and
elongated spermatids) at the highest-dose group were different from
the control pattern 7 and 24 day after treatment (Evenson et al.,
1987).
7.5.2 Embryotoxicity and teratogenicity
Carbendazim (95% purity) was administered by gavage to female
Holtzman rats at dose levels of 0, 100, 200, 400 or 600 mg/kg body
weight per day during days 1-8 of gestation, and the rats were then
sacrificed on days 11 or 20. Maternal toxicity was not observed at
any dose level. At day 11, the crown-rump length, head length,
number of somites and number of embryos per dam were significantly
reduced in groups receiving dosages of 200 mg/kg body weight per day
or more. Open posterior neuropores and limb anomalies were observed
more frequently at dosages equal to or greater than 200 mg/kg body
weight per day. At day 20, increased resorptions, decreased live
litter size and fetal body weight and delayed ossification was
observed at all dosages. Skeletal malformations at the high dosage
levels were attributed to carbendazim exposure. The authors noted
that developmental alterations occurred at stages after the
termination of dosing, suggesting that either the anomalies
represent delays in development or the embryonic cells are
vulnerable at earlier stages than was previously thought (Cummings
et al., 1992).
Carbendazim, in a 0.5% aqueous suspension of methyl cellulose,
was administered by gavage to Crl:CDBR rats (25/dose group) on days
7-16 of gestation at daily doses of 0, 5, 10, 20 or 90 mg/kg per
day. Maternal toxicity was seen only at the highest dosage in the
form of depressed weight gains during the dosing periods and prior
to sacrifice on day 22. Mean values for liver weight and
liver-to-body weight ratio were increased. Decreased pregnancy rate
was observed at the highest dosage. An increase in the incidence of
early resorptions per dam, decreased litter size, and the total
resorption of three litters occurred at the highest dosage, only the
reduction in females per litter being significant. Significant
reductions in mean fetal weight were observed at both 20 and 90
mg/kg per day. A significant increase in the incidence of fetal
malformations was also seen at 90 mg/kg per day. The malformations
consisted primarily of hydrocephaly, microphthalmia, anophthalmia,
malformed scapulae and axial skeletal malformations (vertebral, rib
and sternebral fusions, exencephaly, hemivertebrae and rib
hyperplasia) (Alverez, 1987). The no-observed-effect levels (NOEL)
for the dam and fetus were 20 and 10 mg/kg per day, respectively.
In a further study, Delatour & Besse (1990) reported
embryotoxic effects in eleven Sprague-Dawley rats administered 19.1
mg carbendazim/kg per day by gavage from days 8 to 15 of gestation
and sacrificed at day 21. The authors reported statistically
significant (P < 0.05) increases in dead fetuses and fetal skeletal
and external malformations, and a statistically significant decrease
in fetal weight in the treated group. Since only one dose was used,
it was not possible to determine a NOEL.
The effects of carbendazim on fetal survival and development
were studied by Janardhan et al. (1984) in rats and rabbits. Female
Wistar-rats (8-10 per group) were given carbendazim (98% purity) by
gavage (0, 20, 40 and 80 mg/kg per day) on days 6 to 15 of
pregnancy. Half of each group of animals was killed on day 21 of
gestation and half was allowed to deliver normally. Female albino
rabbits were given 0, 40, 80 and 160 mg carbendazim/kg per day by
gavage on days 6 to 18 of pregnancy and were sacrificed on day 31.
All sacrificed animals were scored for live and dead fetuses and for
resorptions; live fetuses were killed and examined for
abnormalities. After normal deliveries, neonatal deaths and
survivors were counted; survivors were weighed and examined for
gross abnormalities. In rats sacrificed on day 21, dead and resorbed
fetuses accounted for 29% of conceptuses in controls, 48% at a
dosage of 20 mg/kg per day, and 64 to 73% at dosages of 40 and 80
mg/kg per day. In rabbits, dead and resorbed fetuses formed 0% of
total conceptuses in controls and 15, 21.7 and 33.3% at 40, 80 and
160 mg/kg per day, respectively. There were no differences among the
various groups of rats or rabbits with respect to mean weight of
live fetuses, and there were no malformations. In rats giving birth,
the average number of live pups per litter was close to 8 in
controls, 6 at 20 mg/kg per day, and about 5 at 40 and 80 mg/kg per
day. Mean fetal weight was increased by about 13% over controls at
the two highest dosages. There were no still births, neonatal deaths
or gross abnormalities, but mortality at 21 days postpartum was 3.0
to 3.5 times greater at the two highest-doses compared to control
(Janardhan et al., 1984).
Suspensions of carbendazim (in aqueous 0.5% carboxymethyl-
cellulose) were administered by gavage on days 7 to 19 of presumed
gestation to artificially inseminated New Zealand white rabbits (20
rabbits per group). Dose levels were 0, 10, 20 and 125 mg/kg per day
(based on active ingredient) in a volume of 5 ml/kg. The
administration of 125 mg/kg per day inhibited average maternal
weight until day 16 of gestation. There was a slightly decreased
implantation rate at 20 and 125 mg/kg per day and an increased
incidence of resorption at 125 mg/kg per day. These effects resulted
in a decreased live litter size at these two dosages. At 125 mg/kg
per day, litters showed decreased fetal body weight, but the effect
was not statistically significant. The average percentage of
malformed fetuses per litter was significantly increased at 125
mg/kg per day. Compound-related malformations at the highest dosage
consisted of malformed cervical vertebrae and interrelated
malformation of the ribs and proximate thoracic vertebrae (Christian
et al., 1985). The NOEL for maternal toxicity was 20 mg/kg per day
and the NOEL for developmental toxicity was 10 mg/kg per day.
Groups of ChR-CD rats (27-28 pregnant rats per group) were
administered carbendazim (53% a.i.) in their diet at levels of 0,
100, 500, 2500, 5000, 7500 and 10 000 mg/kg from day 6 to day 15 of
gestation. Average doses were equivalent to 0, 8.9, 45.9, 218.4,
431.6, 625.5 and 746.9 mg/kg body weight per day, respectively. On
day 20 of gestation, all pregnant animals were sacrificed and
fetuses delivered by Caesarean section. There was no mortality, no
adverse effect on body weight or clinical signs of toxicity. Food
intake was reduced at the highest-dose level during the period the
test diet was administered, but returned to control levels from days
16 to 20. The number of implantation sites, resorption sites and
live/dead fetuses were not adversely affected by carbendazim
(Sherman, 1970). After evaluating this study, the 1983 JMPR
concluded that there were no external or internal abnormalities
reported that could be considered compound-related. However, no
individual litter data were presented (FAO/WHO, 1985b).
In a further study (Koeter, 1975a), pregnant Wistar-SPF rats
(18-22 per group) were administered carbendazim in the diet at
dosage levels of 0, 600, 2000 and 6000 mg/kg from days 6 to 15 of
gestation. No individual animal or litter data were reported, and
variations in ossification and other skeletal abnormalities were
presented as percentages. Therefore, the teratogenic or fetotoxic
potential of carbendazim to pregnant Wistar-SPF rats cannot be
determined from the results and data presented (FAO/WHO, 1985b).
Groups of pregnant New Zealand albino rabbits (3-11 per group)
were administered carbendazim in the diet at dosage levels of 0,
600, 2000 and 6000 mg/kg from day 6 to 18 of gestation (Koeter,
1975b). There was a significant increase in the number of
supernumery ribs (bilateral) and skull bones in the highest-dose
group and ossification was significantly delayed or absent in these
fetuses. However, there were no individual animal or litter data,
variations in ossification were presented as percentages, and
visceral anomalies were evaluated in only two of the four groups.
The teratogenic potential of carbendazim to pregnant New Zealand
albino rabbits, therefore, cannot be ascertained from the results
and data presented (FAO/WHO, 1985b).
7.6 Mutagenicity and related end-points
Numerous studies have been conducted to assess the mutagenic
potential of carbendazim. Many of the results are conflicting and
many of the study reports do not provide sufficient detail to
evaluate the reasons for the conflicting data. Prior to the
mid-1980s, industrially produced carbendazim contained phenazine
impurities. Therefore, different degrees of purity of carbendazim
might account for some of the discrepancies. Table 9 presents those
studies that reported sufficient experimental detail and data.
Carbendazim is not a heritable gene mutagen. It does not
interact with cellular DNA, induce point mutations or result in germ
cell mutations. This has been demonstrated in both mammalian and
non-mammalian systems in vitro and in vivo, and in somatic cells
as well as in germ cells. Positive results have occasionally been
obtained in gene mutation studies, but this was due to the presence
of phenazines. These contaminants are mutagenic at very low
concentrations in the Salmonella typhimurium Ames test and also in
the mouse lymphoma Y5178Y TK+/- gene mutation assay.
Concentrations of > 4 mg diaminophenazine/kg and 10 mg
aminohydroxyphenazine/kg resulted in positive Salmonella
typhimurium Ames test results. Process changes by some of the
major carbendazim manufacturers have removed the phenazines. This
contaminant is not present when other benzimidazoles such as benomyl
or thiophanate-methyl are metabolized to carbendazim. Carbendazim
does cause numerical chromosome aberrations (aneuploidy and/or
polyploidy) in experimental systems in vitro and in vivo.
Table 9. Studies on mutagenicity of carbendazim
End-points/Tests Species, strains Concentration Activation Result Reference
1. DNA damage and repair
Bacillus subtilis 20-1000 µg per without negative Shirasu et al.
disk (1977)
Salmonella typhimurium 125-2000 µg/plate NR negative Rashid & Mumma
(TA1535, TA1938) (1986)
Escherichia Coli, K-12 125-2000 µg/plate NR
E. Coli, WP2 125-2000 µg/plate NR negative
Sister chromatid exchanges CHO cells 0.13-40.0 µg/ml without negative Ivett (1984)
1.25-40.0 µg/ml with negative
Sister chromatid exchanges human lymphocytes 0.03-30.0 µg/ml with and negative Banduhn & Obe
without (1985)
Sister chromatid exchanges human lymphocytes 1-60 µg/ml with and marginal positive Pandita (1988)
(technical grade) without at 30 µg/ml and above
Unscheduled DNA synthesis male mouse hepatocytes 0.125-12.5 µg/ml Tong (1981a,b)
male rat hepatocytes 0.0125-12.5 µg/ml
Unscheduled DNA synthesis rat hepatocytes 1.04-104.0 µg/ml negative Litton (1981)
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
2. Gene mutation
(1) Bacterial gene S. typhimurium, TA1535, 4-2500 µg/plate with and negative Gericke (1977)
mutation assays TA1537, TA98, TA100 without
Overlay spot test S. typhimurium, his G46, 50-100 µg/spot weak mutagenic Fiscor et al.
TA1530, TA1950 activity at 100 µg/spot (1978)
Plate incorporation assay TA100 50-200 µg/plate NR negative Fiscor et al. (1978)
Spot, liquid culture & S. typhimurium, his 0.25-10 000 µg/ml negative Fiscor et al.
host mediated assays G46, TA1530, TA1535, (1978)
TA1950
Plate incorporation assay S. typhimurium (TA1535, 5-1000 µg/plate with and negative Shirasu et al.
TA1537, TA1538, TA100) without (1977)
E. coli, WP2 hcr 5-1000 µg/plate with and negative Shirasu et al.
without (1977)
S. typhimurium (TA98, 1-300 µg/plate with and negative Pandita (1988)
TA100) without
S. typhimurium (TA1537, 5000 µg/plate positive in TA1537 Albertini (1989)
TA1538, TA97, TA98) & TA97 (using a
pre-incubation assay),
positive in TA1538 &
TA98 only with
activation
S. typhimurium (TA1535, 100-10 000a µg per with and positve at 5000 µg/ Donovan (1982)
TA1537, TA98, TA100) plate without plate with activation
in TA98, TA1537
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
S. typhimurium (TA1535, up to 10 000b µg with and negative up to Russell (1983)
TA1537, TA98, TA100) per plate without 10 000 µg/plate
S. typhimurium positive if Arce (1984a,b)
carbendazim samples
with 4 ppm DAP or
10 ppm AHP impurities
Host mediated assay male ICR mice Total dose of mice negative Shirasu et al.
S. typhimurium his G46 1000, 4000 mg/kg (1977)
S. typhimurium (TA1535, 2-AB with and negative Donovan (1983);
TA1537, TA98, TA100) without Russell (1977)
100-10 000 µg/plate
5-HBC 200-20000 µg/plate negative
(2) Yeast & fungal A. nidulans 2.77 µM without increase in frequency Speakman & Nirenberg
mutation assays (0.53 µg/ml) of colonies resistant (1981)
to carbendazim
Cladosporium cucumerinum 0.58 µM without no effect on frequency Speakman & Nirenberg
(0.11 µg/ml) of colonies resistant (1981)
to carbendazim
Saccharomyces cerevisiae 10-40 µg/ml negative Albertini (1991)
D7
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
(3) In vitro gene mutation
assays
HGPRT CHO cells 3-628 µM with negative Waterer (1980)
3-654 µM without
forward mutation mouse L5178Y 50-250 µM with and positive with Jotz (1980)
at TK locus lymphoma cells without activation at 100 µM,
negative without
activation
(4) Insect germ cell
gene mutation
Sex-linked recessive Drosophila melanogaster 0.5 mg/ml negative Lamb & Lilly
lethals in DMSO (1980)
Germline aneuploidy D. melanogaster, FIX, 400-50 000 ppm negative Osgood et al.
ZESTE feeding to larvae or (1991)
young adult females
(5) In vivo mammalian mouse embryos treated in 200 mg/kg positive Fahrig & Seiler
gene mutation assays utero by dosing the mother (1979)
100-300 mg/kg orally
3. Chromosomal effects
(1) In vitro chromosomal S. cerevisiae 0.001-0.100 mg/ml maximum responses Whittaker et al.
effect for mitotic & meiotic (1990)
chromosome gain at 0.05
and 0.025 mg/ml
S. cerevisiae 5 µg/ml positive for Albertini (1991)
aneuploidy
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
human lymphocytes 0.1-10.0 µM with and not clastogenic, Banduhn & Obe
without but induce (1985)
micronuclei at > 10 µM
human lymphocytes 0.5 mg/ml without did not increase Lamb & Lilly
chromosome (1980)
aberration frequency
(2) In vivo mammalian
chromosomal effect
male & female Sprague- single oral dose did not increase BASF (1975);
Dawley rat bone marrow 300 mg/kg chromosome FAO/WHO (1985b)
cells aberration frequency
Chinese hamster bone oral, 1000 mg/kg did not produce Seiler (1976)
marrow cells chromosome breakage
ICR mice nucleated 2 oral dose of exhibit lagging Seiler (1976)
anaphase cells 1000 mg/kg chromosome, bridge
formation, tripolar
spindle formation, etc
Mouse bone marrow 500 mg/kg did not detect Seiler (1976)
micronuclei
Swiss albino mice 0, 200, 1000, 2000, dose dependent Pandita (1988)
4000 or 6000 mg/kg i.p. increase in
micronucleus
male Chinese hamsters 0, 100, 500 or no effect on Pandita (1988)
bone marrow 1000 mg/kg i.p. hyperploidy
Table 9 (contd).
End-points/Tests Species, strains Concentration Activation Result Reference
(3) In vivo germ cell
chromosomal mutation
Mouse dominant lethal NMRI mice 500 mg/kg i.p. per did not exhibit a Hoechst (1974)
test day, 5 days dominant lethal effect
male NMRI mice 300 mg/kg gavage did not exhibit a Hoechst (1974)
per day, 5 days dominant lethal effect
a Phenazin concentration in carbendazim: 9 ppm DAP, 5 ppm AHP
b Phenazin concentration in carbendazim: < 0.6 ppm DAP, < 4 ppm AHP
DAP = diaminophenazine; AHP = aminohydroxyphenazine; i.p. = intraperitoneal; NR = not reported
7.7 Carcinogenicity
In a study by Wood (1982), groups of CD-1 mice (80 males and 80
females per group) were administered carbendazim (99% a.i.) in the
diet at dose levels of 0, 500, 1500 and 7500 mg/kg diet for two
years. The highest-dose was reduced to 3750 mg/kg after 66 weeks for
the males because of increased mortality (62 surviving controls, 32
surviving at 7500 mg/kg); females, however, received 7500 mg/kg
throughout the study period. Animals were 6-7 weeks old at the start
of this study. Mortality was compound-related in male mice. The
high-dose males were sacrificed at week 73 because of a significant
increase in mortality (23 surviving). Only nine males in the
1500-mg/kg group survived to week 104, compared with 18 surviving
control males. Females did not show this increase in mortality.
There were no dose-related effects on body weight or food
consumption at any time during the study, although terminal body
weights for the low- and mid-dose males were less than those of the
control and high-dose males. Clinical data were similar for all
treatment and control groups. Haematological measures were
unaffected.
Both absolute and relative thymus weights were significantly
decreased in females at 500 and 1500 mg/kg, but not in the high-dose
group. Absolute liver weight was increased in the high-dose females
and relative liver weight at the two highest-dose levels. The organ
weights of male mice were variable and only the kidney and thymus
weights appeared to be decreased as a result of treatment. Absolute
kidney and thymus weights were depressed in male mice at all
treatment levels, but relative kidney and thymus weights were
significantly decreased only in the high-dose males. Histological
examination revealed dose-related changes in the thymus (lymphoid
depletion) and accumulation of yellow-brown pigment in the renal
tubules for mid- and high-dose male mice.
Examination of the testes demonstrated an increase in the
frequency of sperm stasis in mid- and high-dose males, together with
increased germinal cell atrophy (bilateral only). There was no
trend, however, towards unilateral germinal cell atrophy, the
incidence in controls being greater than or equal to that of treated
males. These effects are, therefore, not considered to be
compound-related.
The examination of male mice livers revealed a significant
hepatotoxic effect at 1500 and 7500 mg/kg, demonstrated by
centrilobular hypertrophy, necrosis and swelling. There was no
increase in the frequency of hepatocellular adenomas; these occurred
with equal frequency in control and treatment groups. There was a
significant increase in hepatocellular carcinomas but only at 1500
mg/kg. However, too few males survived at the high-dose level to 17
months (510 days) to support the conclusion of no oncogenic effect
at that dose level.
The occurrence of total hepatic tumours (hepatocellular
carcinomas, hepatocellular adenomas and hepatoblastomas) is given in
Table 10. This was statistically increased (P < 0.05) for the low-,
mid-, and high-dose females. It was also statistically increased (P
< 0.05) in mid-dose males but was not evaluated in high-dose males
because of the high rate of mortality.
In conclusion, this study on CD-1 mice showed that there were
statistically significant increases (P < 0.05) in the incidence of
hepatocellular carcinoma at 1500 mg/kg for males and at all dose
levels for females, the response being dose-related (P < 0.05).
Because of high mortality in the high-dose males, a dose-response
relationship could not be determined. In addition, the high mor
tality rate in male controls further hampered the interpretation of
results. Histopathological analysis of the hepatocellular tumours in
the test animals showed no difference from controls, and the median
latent period for development of these hepatocellular carcinomas
showed no significant decrease in the test animals. No carcinogenic
effect was observed in tissues other than the liver (Wood, 1982).
In a study on SPF Swiss mice (100 males and 100 females per
group), carbendazim was administered in the diet at dosage levels of
0, 150, 300 and 1000 mg/kg diet for 80 weeks. The highest-dose was
increased to 2000 mg/kg at week 4 and to 5000 mg/kg at week 8 for
the remainder of the study. Animals were examined for behaviour and
clinical signs of toxicity, and body weight measurements were
determined throughout the study. Gross pathology investigations were
performed on all animals, liver and kidney weights were recorded,
and tissues were examined microscopically. There were no
compound-related effects on general condition, mortality or body
weight. Survival at term was 70% for males and 80% for females.
Relative liver weights for high-dose males and females were
significantly different from those of controls, but there were no
changes in kidney weights. The results are shown in Table 11. The
combined incidence of hepatocellular adenomas and hepatocellular
carcinomas increased with increasing dose levels for both males and
females. Males showed a more pronounced induction of liver tumours
and a more frequent occurrence of hepatocellular carcinomas which
were often found simultaneously with hepatocellular adenomas.
Females, on the contrary, showed only hepatocellular adenomas in
most cases (Beems et al., 1976; Mohr, 1977).
Table 10. Incidence of combined primary hepatocellular tumours and mean and median period for development
after treatment with carbendazim in CD-1 micea
Male mice Female mice
Carbendazim concentration (mg/kg diet): 0 500 1500 7500 (3750) 0 500 1500 7500
Combined incidences of hepatic tumours 13 20 23b NAc 1 9b 21b 15b
Median period for development (days) 633 697 651 NAc 732 724 697 733
Mean period for development (days) 628 671 628 NAc 732 706 689 653
Number of mice examinedd 80 80 80 80 79 78 80 78
Median survival time (weeks) 79 72 69 64 91 91 91 91
Number of mice alive at termination 18 14 9 23 22 15 13 20
a From: Wood (1982)
b Significant by Fisher's Exact Test at P < 0.05 level of probability
c NA = not applicable. This group was terminated after 516 days on test (test week 73) due to a high rate of mortality. In
view of the early termination of this test group and low incidence of hepatic tumours observed in this group, the inclusion
of its data in statistical evaluation of hepatic tumour incidence is considered inappropriate.
d Mice found dead or sacrificed in extremis prior to each group's termination were subjected to gross pathological evaluation.
Table 11. Incidence of proliferation lesions of the hepatocytes in Swiss micea
Male mice Female mice
Carbendazim concentration (mg/kg diet): 0 150 300 5000 0 150 300 5000
Number of animals examinedb 100 94 98 100 94 99 98 95
Liver nodular hyperplasiac 0 8 11 25 0 5 3 11
Hepatocellular adenoma 9 5 13 14 1 1 3 8
Hepatocellular carcinoma 1 3 4 9 1 0 0 0
a Adapted from: Mohr (1977)
b A number of animals could not be examined because of well-advanced autolysis. In addition, slides were not available for
some of the animals.
c Values for this non-neoplastic lesion included all types of cellular alteration. A statistical analysis was not conducted.
In a study on HOE NMRKf (SPF 71) mice (100-120 males and
females per group), carbendazim was administered in the diet for 96
weeks at dosage levels of 0, 50, 150, 300 and 1000 mg/kg diet. The
highest-dose was increased to 2000 mg/kg at week 4 and to 5000 mg/kg
at week 8 for the remainder of the study. Animals were examined for
behaviour and general condition, as well as for body weight,
food/water consumption and mortality. Gross necropsies were
performed on all animals, liver and lung weights were recorded, and
all organs and tissues were examined microscopically. An interim
sacrifice was made at 18 months of 20 males and 20 females from the
control group and the highest-dose group.
There were no compound-related effects on behaviour, body
weight gain, food/water consumption or mortality. At 22 months there
was 24-31% mortality in male mice and 37-52% mortality in females in
all groups of mice. As there was no difference between the treated
and control groups, it was concluded mortality was not influenced by
the feeding of carbendazim. The mean daily consumption of
carbendazim in mg/kg body weight for males and females,
respectively, was 5.8 and 7.1 at 50 mg/kg diet, 17.1 and 21.2 at 150
mg/kg diet, 34.4 and 41.9 at 300 mg/kg diet, and 548.4 and 682.3 at
5000 mg/kg diet.
Examination of lung and liver weights at 18 and 22 months
demonstrated an increase in absolute and relative liver weights in
both male and female mice at 5000 mg/kg diet. Macroscopic and
microscopic examination of 20 male and 20 female animals killed
after 18 months of receiving 5000 mg carbendazim/kg revealed
compound-related effects on the liver. All animals demonstrated
centrilobular hypertrophy, single cell necrosis, mitotic cells and
pigmented Kupffer cells. The tissues of the remaining 100 males and
100 females exposed to 5000 mg/kg were evaluated at 22 months. There
was marked liver cell hypertrophy (greater than in animals treated
for 18 months only), clear cell foci, mitosis, inclusion bodies in
enlarged cell nuclei, multiple cell necrosis, and greenish yellow
pigment in Kupffer cells.
Neoplastic nodules (adenomas), carcinomas, fibrosarcomas and
other tumorigenic responses in the liver were equally distributed
among all groups (Tables 12 and 13). Although haemangiomas of the
liver (Table 14) were found in all treated groups (but not in
controls), no dose-related response was evident. Lung adenomatosis
was equally distributed among all groups. There was no effect of
carbendazim on the incidence or time of onset of tumours, and the
total number of benign and malignant tumours was comparable among
the various groups of mice. Thus, there was no evidence of a
carcinogenic effect from carbendazim when administered in the diet
to mice at doses up to and including 5000 mg/kg diet for 22 months
(Donaubauer et al., 1982).
Table 12. Incidence of tumours in HOE NMRKf mice after 18 months of
exposurea
Mice treated with
Control 5000 mg/kg diet
male female male female
Lung
adenoma 3 5 4 -
Uterus
stromal sarcoma 1 -
Ovary
tubular adenoma 1 -
granulosa cell tumour 1 -
luteoma and tubular adenoma 1 -
Skin and subcutaneous tissue
round cell sarcoma - - - 1
adenocarcinoma - - 1 -
Lymph nodes
lymphosarcoma - - - 1
3 9 5 2
a Adapted from Donaubauer et al. (1982)
7.8 Neurotoxicity
Carbendazim (500, 2500 and 5000 mg/kg) has been evaluated for
neurotoxic potential in white Leghorn hens (10 per group). Controls
consisted of the vehicle (corn oil) and the neurotoxin tri- o-tolyl
phosphate (TOTP). The hens were observed daily for mortality and
clinical neurotoxicity for four weeks. Neurotoxic signs, consisting
of leg weakness, ataxia and/or "goose-stepping" gait, were observed
in hens treated with TOTP. Less severe and reversible signs,
consisting of slight leg weakness and ataxia, were observed in hens
treated with 5000 mg carbendazim/kg, but no neurotoxic signs were
observed for those treated with 500 or 2500 mg/kg. Microscopic
examination indicated that there was no axonal degeneration or
demyelination in carbendazim-treated animals (Goldenthal, 1978).
Table 13. Possible preneoplastic changes and primary neoplasms in the livers and lungs of HOE NMRKf mice
after 22 months of exposurea
Carbendazim concentration: 0 50 150 300 5000
(mg/kg diet) male female male female male female male female male female
Liver
Neoplastic nodules
(adenomas) 3 2 1 1
Clear cell foci 3 4
Basophilic foci 1 1
Haemangiomas (liver) 2 3 2 2 1
Lungs
Adenomatosis 29 10 31 8 30 10 25 9 22 12
Adenocarcinomas 1 1
Keratinizing squamous
cell carcinomas 1
Cavernous haemangioma 1
32 10 35 11 34 11 27 12 27 17
a Adapted from Donaubauer et al. (1982)
Table 14. Haemangiomas of the liver in HOE NMRKf mice after
22 months of exposurea
Dose (mg/kg diet) Male Female
0 0 0
50 2 0
150 3 0
300 2 2
5000 0 1
a Adapted from Donaubauer et al. (1982)
7.9 Toxicity of contaminants
When manufactured under certain conditions, carbendazim
contains a class of contaminants called phenazines. These
contaminants are mutagenic at very low concentrations in the
Salmonella typhimurium Ames test and also in the mouse lymphoma
Y5178Y TK+/- gene mutation assay. Concentrations of > 4 ppm
diaminophenazine or 10 ppm aminohydroxyphenazine yielded positive
results. Process changes by some of carbendazim manufacturers have
now removed the phenazines.
7.10 Mechanisms of toxicity - mode of action
Biochemical studies on the mechanism of action of benzimidazole
compounds have shown that their biological effects are caused by
interactions with cell microtubules (Davidse & Flach, 1977). These
cellular structures are present in all eukaryotic cells and are
involved in several vital functions, such as intracellular
transports and cell division. Benzimidazole compounds have been used
as anticancer drugs and as anthelminthic drugs in animals and humans
because they act as spindle poisons by interfering with the
formation and/or functioning of microtubules. However, eukaryotes
are known to be unequally sensitive to each benzimidazol compound,
which explains the use of these compounds in helminthiases.
Selective toxicity of benomyl and carbendazim for fungi has been
explained by comparing their binding to fungal and mammalian
tubulin. The different sensitivity of several fungi has also been
explained by the different affinity of benomyl and carbendazim for
fungal tubulin.
Benomyl has been found to bind to fungal tubulin but not to
porcine brain tubulin, indicating that mammalian tubulin has no, or
at least low, affinity for benomyl (Davidse & Flach, 1977). This is
in agreement with the observation that benomyl at concentrations
that are lethal for sensitive fungi does not interact with in vitro
microtubule assembly in these brain extracts. In vitro ID50
values for several mycelial extracts of various fungal species
sensitive to benomyl were all below 5 µmol/litre (Davidse & Flach,
1977). In vitro rat brain tubulin polymerization was inhibited to
about 20% at benomyl or carbendazim concentrations of 25 µmol/litre
(De Brabander et al., 1976b). For comparison, a standard antitubulin
drug in humans such as vincristine inhibited 50% tubulin assembly at
0.1 µmol/litre in the same experiment. The assembly of sheep and
calf brain microtubule was also found to be unaffected by
carbendazim concentrations higher than 100 µmol/litre (Ireland et
al., 1979).
Mitotic arrest by benzimidazole and six analogues at metaphase
was evaluated in human lymphocyte cultures. Structure-activity
relationships indicate that antimitotic activity is related to C6
substitution of the benzimidazole moiety (Holden et al., 1980). In
this study, however, benomyl and carbendazim were not tested. The
question of whether all C6 unsubstituted benzimidazoles, such as
benomyl and carbendazim, have no effect on mitosis of human
lymphocytes in cell cultures is therefore unresolved.
A link between the effects of benomyl and carbendazim on
tubulin and their teratogenic effects has been postulated (Ellis et
al., 1987, 1988).
8. EFFECTS ON HUMANS
8.1 General population exposure
No references to carbendazim poisoning have been documented in
the literature. Recent data to estimate dietary exposure of the
benzimidazole benomyl, based on food consumption patterns within the
USA and elsewhere, indicate exposure below the NOELs from animal
toxicity tests. Further information is given in section 5.2.1 of
Environmental Health Criteria 148: Benomyl (WHO, 1993).
8.2 Occupational exposure
Selected blood profiles from 50 factory workers involved in the
manufacture of benomyl and carbendazim were compared to those of a
control group of 48 workers who were not exposed to these two
fungicides. White blood cell count, red blood cell count, and
haemoglobin and haematocrit values were comparable among the two
groups. There were no quantitative estimates of exposure given for
the factory workers. No female employees were included in the
control group (Everhart, 1979; FAO/WHO, 1985a).
A study was performed to determine whether exposure to benomyl
and carbendazim had an adverse effect on the fertility of 298 male
manufacturing workers exposed to benomyl between 1970 and 1977. The
workers ranged from 19 to 64 years of age (79% were between 20 and
39, and 78% of the spouses were similarly aged between 20 and 39
years). Exposure duration ranged from less than one month to 95
months, and more than 51% of the workers were potentially exposed
from 1 to 5 months. The birth rates of exposed workers' spouses were
compared with those of four comparison populations from the same
county, state, region and country (USA). There was no reduction in
fertility as shown by the birth rates for the study population,
which were generally higher than the comparison populations.
Spermatogenesis among workers was not examined (Gooch, 1978;
FAO/WHO, 1985a).
9. EFFECTS ON OTHER ORGANISMS IN THE LABORATORY AND FIELD
9.1 Microorganisms
Studies on soil effects have concentrated on benomyl. However,
carbendazim, the breakdown product of benomyl, would be expected to
produce comparable results.
Soil respiration has been found to be little influenced by
benomyl at concentrations below 10 mg/kg, which is the maximum soil
concentration expected after use at recommended application rates
(Hofer et al., 1971; van Fassen, 1974; Peeples, 1974; Weeks &
Hedrick, 1975).
A study on the influence of benomyl on soil nitrogen
mineralization showed that the release of ammonia was not decreased
by benomyl, whereas the influence of the fungicide on nitrification
varied from a stimulation (van Fassen, 1974), through no effect
(Mazur & Hughes, 1975), to a decreased nitrification (Hofer et al.,
1971; Wainwright & Pugh, 1974). The differences may be related to
the soil composition since Hofer et al. (1971) found a greater
effect in sandy than in organic soil.
Benomyl, in combination with eleven other pesticides that were
used in an orchard spray programme, had only a minimal and
short-term effect on respiration, ammonification and nitrification
at concentrations expected after recommended use of benomyl over a
spraying season. Ten times the recommended application rates had a
pronounced effect on both respiration and nitrification, which
lasted for more than 4 weeks (Helweg, 1985).
The influence of benomyl and carbendazim on soil microbial
activity was studied in Sweden following repeated annual
applications during autumn to winter cereals for a period of 3 to 5
years. The effects of the fungicides on straw decomposition, balance
of straw fungal flora and nitrogen mineralization in the soil were
investigated in field and laboratory experiments. The decomposition
of straw in the field was not affected in clay soils by annual
applications of up to 2 kg/ha. In sandy soils, rates of up to 0.5
kg/ha had no effect, but in one case at 2 kg/ha the initial stages
of straw decomposition were slightly inhibited. All doses tested in
both clay and sandy soils caused changes in the composition of the
straw fungal flora (Torstensson & Wessen, 1984).
Tests were conducted to determine the effects of the fungicide
carbendazim (Bavistin), the herbicide fluometuron (Cotoran) and the
insecticide curacron on Egyptian soil fungi when applied at the
recommended field rates and at 4 and 8 times these rates.
Carbendazim produced a significant inhibition of the total count of
fungi at all three rates after 5 and 40 days and at the higher
application rates after 80 days. The response of Aspergillus sp.
alone to this fungicide reflected the effects on the total fungal
count (Abdel-Fattah et al., 1982).
An application of carbendazim (5 mg formulation/kg soil) and
Calixin (1.5 mg formulation/kg soil) significantly enhanced the
microbial activity, as well as the availability of ammonium nitrogen
and phosphate in the soil, while nitrite, nitrate and available
potassium were found to decrease. During incubation, CO2 evolution
decreased during the first 21 days, increased up to 42 days, and,
thereafter, decreased with both treatments (Khan et al., 1987).
9.2 Aquatic organisms
The effect of carbendazim was monitored using the green alga
Selenastrum capricornutum in an OECD guideline test (201). The
EC50 (based on total growth) at 72 h was 1.3 mg/litre and at 120 h
was 1.6 mg/litre. The no-observed-effect concentration (NOEC) was
0.5 mg/litre. To study whether carbendazim was algistatic or
algicidal, organisms were recultured at the end of the initial 120 h
of incubation. Regrowth occurred in the control but not in the test
cultures (8.0 mg/litre) after a period of 9 days. Carbendazim was,
therefore, considered to be algicidal (Douglas & Handley, 1987). In
another study using Chlorella pyrenoidosa, the 48-h EC50 for
growth inhibition was calculated to be 0.54 mg/litre (Canton, 1976).
The acute toxicity of carbendazim to a variety of aquatic
organisms is summarized in Table 15. For 96-h tests, LC50 values
ranged from 0.007 mg/litre for channel catfish ( Ictalurus punctatus
yolk-sac fry) to 5.5 mg/litre for bluegill sunfish (Palawski &
Knowles, 1986).
In a static renewal 21-day test using Daphnia magna, onset of
reproduction was the most sensitive indicator, being significantly
delayed at 0.025 mg carbendazim/litre (measured concentration).
Other parameters (number of days reproducing, total number of young,
and young produced per day) were affected significantly at 0.05
mg/litre. The NOEC was 0.013 mg/litre (measured concentration)
(Hutton et al., 1986).
Gillet & Roubaud (1983) showed that the toxicity of carbendazim
to carp ( Cyprinus carpio) was greater for the fertilization and
early development stages than for the adult stages. They found the
30-min LC100 during the fertilization stage to be < 5 mg/litre at
pH 9 and < 2.5 mg/litre at pH 7. The rainbow trout ( Oncorhynchus
mykiss) was more sensitive, with an LC100 (30 min, pH 9) of 0.5
mg/litre and 71% survival, compared to controls, at 0.05 mg/litre.
Table 15. Toxicity of carbendazim to aquatic organisms
Organism Size/a Stat/ Temperature Hardnessb pH Duration LC50 Reference
age flow (°C) (mg/litre) (h) (mg/litre)
Freshwater
Water flea < 24 h stat 20 179 7.5 48 0.087 Hutton (1988)
(Daphnia magna) < 24 h stat 20 48 0.46 Canton (1976)
Rainbow trout 3 months stat 15 48 1.8 Canton (1976)
(Oncorhynchus mykiss) 0.8 g stat 13 7.5 96 0.36 Heinemann (1971)
yolk-sac fry stat 10 40-48 7.4 96 0.145 Palawski & Knowles (1986)
swimup fry stat 10 40-48 7.4 96 0.32 Palawski & Knowles (1986)
0.2 g stat 10 40-48 7.4 96 0.37 Palawski & Knowles (1986)
1.2 g stat 10 40-48 7.4 96 0.87 Palawski & Knowles (1986)
stat 12 40-48 7.4 96 > 1.8 Palawski & Knowles (1986)
stat 17 40-48 7.4 96 0.87 Palawski & Knowles (1986)
stat 22 40-48 7.4 96 0.1 Palawski & Knowles (1986)
stat 10 40-48 6.5 96 0.64 Palawski & Knowles (1986)
stat 10 40-48 7.5 96 0.41 Palawski & Knowles (1986)
stat 10 40-48 8.5 96 0.34 Palawski & Knowles (1986)
stat 10 40 7.4 96 0.78 Palawski & Knowles (1986)
stat 10 320 7.4 96 0.88 Palawski & Knowles (1986)
Bluegill sunfish 0.95 g stat 22-23 67 7.0 96 5.5 Wetzel & Hutton (1984)
(Lepomis macrochirus) 0.2 g stat 22 40-48 7.4 96 > 3.2 Palawski & Knowles (1986)
Channel catfish yolk-sac fry stat 22 40-48 7.4 96 0.007 Palawski & Knowles (1986)
(Ictalurus punctatus) swimup fry stat 22 40-48 7.4 96 0.012 Palawski & Knowles (1986)
0.2 g stat 22 40-48 7.4 96 0.01 Palawski & Knowles (1986)
1.2 g stat 22 40-48 7.4 96 0.019 Palawski & Knowles (1986)
stat 12 40-48 7.4 96 > 0.56 Palawski & Knowles (1986)
stat 17 40-48 7.4 96 0.14 Palawski & Knowles (1986)
stat 22 40-48 7.4 96 0.032 Palawski & Knowles (1986)
stat 22 40-48 6.5 96 0.023 Palawski & Knowles (1986)
Table 15 (contd).
Organism Size/a Stat/ Temperature Hardnessb pH Duration LC50 Reference
age flow (°C) (mg/litre) (h) (mg/litre)
stat 22 40-48 7.5 96 0.014 Palawski & Knowles (1986)
stat 22 40-48 8.5 96 0.023 Palawski & Knowles (1986)
stat 22 40 7.4 96 0.018 Palawski & Knowles (1986)
stat 22 320 7.4 96 0.024 Palawski & Knowles (1986)
Toad tadpole stat 25 48 3.5 Nishiuchi & Yoshida (1974)
(Bufo bufo japonicus)
Marine and Estuarine
Eastern oyster 25-50 mm flow 18-20 96 > 1.145c Boeri (1988a)
(Crassostrea virginica)
Mysid shrimp stat 23 96 0.098 Boeri (1988c)
(Mysidopsis bahia)
Sheepshead minnow 0.14 g stat 22 96 > 1.158 Boeri (1988b)
(Cyprinodon variegatus)
a stat = static conditons (water unchanged for duration of test); flow = flow-through conditions (carbendazim concentration
in water continuously maintained)
b hardness given as mg CaCO3/litre
c EC50 based on rate of shell deposition
Bluegill sunfish were exposed to benomyl, carbendazim and 2-AB
at concentrations of 0.05 mg/litre (measured concentrations 0.01 to
0.04 mg/litre) and 5.0 mg/litre (measured concentration 2 to 5
mg/litre). No residue was found in the tissues of fish exposed to
low levels of these compounds, but detectable residues were found in
the tissues of fish exposed to the high levels. However, there was
no build-up or bioconcentration with time (DuPont, 1972).
9.3 Terrestrial organisms
Van Gestel et al. (1992) exposed red earthworms ( Eisenia
andrei) to carbendazim, added as an aqueous solution of the
formulation Derosal, to artificial soil. The final concentrations in
soil were 0, 1, 3.2, 10, 32 and 100 mg formulation/kg dry soil,
corresponding to 0, 0.6, 1.92, 6.0, 19.2 and 60 mg carbendazim/kg.
The worms had been acclimatised for 1 week in the artificial soil,
which contained 8 g cow dung per kg soil as food for the worms. The
soil pH (7.3) was higher than optimum for this species and resulted
in a lower cocoon production in this test than expected. At the two
highest concentrations of 19.2 and 60 mg a.i./kg, all worms died
over the 3 week experimental period; an LC50 of 5.7 (4.7-6.9) mg
a.i./kg was calculated. Growth was significantly reduced at 6.0
mg/kg soil, and reproduction (measured as cocoon production, number
of fertile cocoons and numbers of juveniles) was significantly
reduced at > 1.92 mg a.i./kg soil. The EC50 for cocoon
production was calculated to be 2.9 (2.2-3.8) mg a.i./kg dry soil.
The response curve for reproduction and growth effects of
carbendazim was steep. Vonk et al. (1986) had reported comparable
results for a second Eisenia species, E. foetida, in a different
artifical soil; the LC50 was 9.3 mg/kg soil and the NOEC for
cocoon production 2.0 mg/kg soil.
Carbendazim (99.3% purity) was evaluated for acute contact
toxicity after thoracic application in honey-bees ( Apis mellifera).
Each treatment level consisted of four replicates of ten bees each.
Forty bees served as positive control (using carbaryl) and forty as
negative control. No deaths occurred after the application of 50 µg
carbendazim/bee, the highest rate tested. Carbendazim is, therefore,
classified as "relatively non-toxic" to the honey-bee (Meade, 1984).
The LD50 for bobwhite quail, with an observation period of 14
days following the single oral dose, was > 2250 mg/kg body weight
(Beavers, 1985). For mallard ducks the dietary 5-day LC50 was >
10 000 mg carbendazim/kg diet (Fink, 1975).
9.4 Population and ecosystem effects
Van Gestel (1992) has summarized reports of toxicity of
carbendazim on earthworms from field studies based on different soil
types, application rates and crops (Table 16). Estimated soil
concentrations in this table are based on application rates; it has
been assumed that there is no mobility of the compound beyond the
top 2.5 cm of soil and homogeneous distribution of carbendazim in
this layer. For orchard application, it was further assumed that 50%
of the applied active ingredient reached the soil. Reported effects
include reduced numbers, reduced activity (as reduced removal of
leaf litter) and reduced reproduction of worms. Application rates
were within the recommended rates for carbendazim as a fungicide on
these crops.
Table 16. Summary of earthworm toxicity data on carbendazim in field studies
Crop/soil type Dosage Estimated soil Time Effect Reference
(kg/ha) concentration (days)
(mg/kg)
Grass 0.3 0.9 12 11% reduction in number Ammon (1985)
Grass 0.56 1.6 49 72% reduction in cast production Keogh & Whitehead
42% more litter left (1975)
Winter wheat/clay 0.15 0.4 275 14% reduction in juveniles Lofs-Holmin (1981)
50% increase in adult
Allolobophora caliginosa
25% increase in juveniles
25% reduction in adult
A. chlorotica Lofs-Holmin (1981)
Apples/loam 0.375a 1.6 245 reduction in litter removal Cook & Swait (1975)
a Three separate applications of carbendazim
10. EVALUATION OF HUMAN HEALTH RISKS AND EFFECTS ON THE ENVIRONMENT
Benomyl and carbendazim are two different fungicides in their
own right. However, carbendazim is also the main metabolite of
benomyl in mammals and the degradation product of benomyl in the
environment. Butyl isocyanate is the chemical moiety removed from
benomyl when carbendazim is formed. Given the similar toxicities
caused by benomyl and carbendazim and the different toxicological
profile of butyl isocyanate, the two fungicides are evaluated
together in this monograph.
10.1 Evaluation of human health risks
The two primary routes of exposure to humans are through diet
and through manufacturing or use of the product.
There is limited information on actual dietary exposure to
benomyl and carbendazim. Dietary exposure has been estimated in the
Netherlands and the USA. In the USA, exposure based on dietary
habits, measured residue levels, and the percentage of crop treated
has been calculated for various subgroups of population. These
calculations indicate that the estimated benomyl exposure is
0.144-1.479 µg/kg per day (section 5.2.1 of Environmental Health
Criteria 148: Benomyl (WHO, 1993)). In the Netherlands, the mean
dietary intake was estimated to be 0.83 µg/kg per day (0.05 mg/day
per person). These levels of exposure are below the recommended ADI
of 0.01 (carbendazim) and 0.02 (benomyl) mg/kg body weight.
The average air levels of benomyl and carbendazim have been
determined in a manufacturing facility (section 5.3) and found to be
less than 0.2 and 0.3 mg/m3, respectively. Both values are below
the Threshold Limit Value of 5-10 mg benomyl/m3 established by a
number of governmental agencies.
In one study, potential respiratory and dermal exposure to
benomyl wettable powder formulation was determined under several
agricultural use situations (section 5.3). The highest rates of
exposure occurred in situations of mixing and loading in preparation
for aerial application; dermal and respiratory exposures were,
respectively, 26 and 0.08 mg/person per cycle. Home users and
agricultural workers re-entering treated fields were estimated to be
exposed to about 1 mg/person per cycle and 5.9 mg/person per hour,
principally through the dermal route.
Because of the low mammalian toxicity, acute benomyl or
carbendazim poisonings are unlikely to occur under conditions of
normal use.
Several studies of agricultural workers (section 8.2 of
Environmental Health Criteria 148: Benomyl (WHO, 1993)) have shown
some cases of contact dermatitis after exposure to benomyl. These
effects can be significantly reduced or eliminated by wearing
long-sleeved shirts, long trousers and gloves.
There is little information on health effects in humans as a
consequence of exposure to either benomyl or carbendazim. Two
studies have been conducted on factory workers involved in the
manufacture of benomyl (section 8.2). In one study, haematological
profiles from 50 factory workers involved in the manufacture of
benomyl were comparable to those from a control group of 48 workers.
A second study found no decrease in the birth rate of the wives of
298 factory workers exposed to benomyl.
Extensive studies of various species of laboratory animals show
reproductive, developmental, mutagenic and carcinogenic effects
associated with both benomyl and carbendazim. The effects observed
on rat fetuses were microphthalmia, hydrocephaly and encephaloceles.
The no-observed-effect levels (NOEL) of benomyl for developmental
toxicity are equal to or greater than 10 mg/kg body weight per day,
depending upon the species and route of administration. Similarly,
the NOEL of benomyl for reproductive effects in the male rat appears
to be 15 mg/kg body weight per day after gavage dosing. In feeding
studies with both benomyl and carbendazim, the NOEL appears to be
500 mg/kg diet (equivalent to 25 mg/kg body weight per day).
However, one benomyl feeding study reported a NOEL of less than 1
mg/kg diet (0.05 mg/kg body weight per day) for male reproductive
effects. The reason for the discrepancy between the NOEL in this
latter study and other investigations is unknown.
The only consistent genotoxic effect noted in animal studies is
the induction of numerical chromosomal aberrations. These effects
are consistent with the interaction of benomyl and carbendazim with
microtubule formation.
Rat carcinogenicity studies did not show any carcinogenic
effect for either compound. Benomyl and carbendazim induce
hepatocellular tumours in CD-1 and SPF Swiss mice but not in NMRKf
mice. This finding in mice is not considered to be a result of a
direct genotoxic action. Rather, it appears to be associated with
liver toxicity in strains of mice that are highly susceptible to
tumour formation at this site.
Benomyl and carbendazim are spindle poisons. Effects on target
cells are consequences of binding to microtubules, giving toxicities
similar to those of other spindle poisons such as colchicine and
vincristine. Benzimidazole compounds in general and benomyl and
carbendazim in particular have selective effects on the microtubules
of different eukaryotes. Reasons for this selectivity include the
binding capability to different tubulins and pharmacokinetic
differences across species. In vitro concentrations of benomyl
used to kill sensitive fungi were found to be ineffective in
disturbing mammalian microtubular functions. These studies on the
mechanism of action of benomyl and carbendazim indicate a selective
effect of these compounds for target species.
In summary, the LD50, as determined in a number of test
species, for benomyl ranges from > 2000 to > 12 000 mg/kg and for
carbendazim from > 2000 to > 15 000 mg/kg. There are no known
reports of human poisoning for either compound. This, coupled with
the low estimated environmental levels of both compounds, would
suggest that the possibility of acute poisoning by benomyl or
carbendazim is very remote. Similarly, the data available on test
species make it unlikely that either benomyl or carbendazim is
carcinogenic for humans. The NOELs for both reproductive and
teratogenic effects of benomyl and carbendazim (i.e. 10-15 mg/kg) do
raise a possibility that an accidental ingestion of either fungicide
could adversely alter reproductive outcome in humans, but the
likelihood that such poisoning would occur is remote. The
selectivity of these two benzimidazole compounds for the tubulin of
the target species (fungi) and their relative ineffectiveness to
disturb mammalian microtubule function further reduce the
possibility of their having toxic effects in humans.
10.2 Evaluation of effects on the environment
Benomyl is rapidly converted to carbendazim in various
environmental compartments, the half-lives being 2 and 19 h in water
and soil, respectively. Therefore, data from studies on both benomyl
and carbendazim are relevant for the evaluation of environmental
effects.
Carbendazim persists on leaf surfaces and in leaf litter. In
soil the half-life is between 3 and 12 months, and the compound may
be detected for up to 3 years. However, in many cases, major
residues will be lost within a single season. Residues of carben
dazim and its metabolites are strongly bound or incorporated into
soil organic matter. The strong adsorption (Koc = approximately
2000) of carbendazim to soil and sediment particles reduces its
bioavailability to terrestrial and aquatic organisms. Similarly, the
mobility of carbendazim in soil is limited, and it is not expected
to leach to ground water.
Benomyl and carbendazim are highly toxic to some aquatic
organisms in laboratory tests, the most sensitive species being the
channel catfish with a 96-h LC50 for yolk-sac fry of 0.006 mg
benomyl/litre. However, this toxicity is unlikely to be manifest in
the environment for most aquatic organisms because of the low
bioavailability in surface waters. The exposure of sediment-living
organisms could be greater, but no test results are available for
these organisms.
Benomyl and carbendazim affect groups of fungi in soil but do
not seem to modify the overall microbial activity of the soil when
used at normal field rates.
In both the laboratory and field, benomyl and carbendazim
applied at recommended rates cause deaths and sublethal reproductive
effects on earthworms of many different species. Surface-feeding
species eating leaf litter are most at risk. Populations may take
more than 2 years to recover. There are no studies available on
other litter and soil invertebrates.
Benomyl and carbendazim have low toxicity for birds and
carbendazim is classified as "relatively non-toxic" to honey-bees.
10.3 Conclusions
Benomyl causes dermal sensitization in humans. Both benomyl and
carbendazim represent a very low risk for acute poisoning in humans.
Given the current exposures and the low rate of dermal absorption of
benomyl and carbendazim, it is unlikely that they would cause
systemic toxicity effects either in the general population or in
occupationally exposed subjects. These conclusions are drawn from
animal data and from the limited human data available, but these
extrapolations are supported by the understanding of the mode of
action of carbendazim and benomyl in both target and non-target
species.
Further elucidation of the mechanism of toxicity of carbendazim
and benomyl in mammals will perhaps permit a better determination of
no-observed-effect levels. Binding studies on tubulins of target
cells (testis and embryonic tissues) will facilitate comparisons
across species.
Carbendazim is strongly adsorbed to soil organic matter and
remains in the soil for up to 3 years. It persists on leaf surfaces
and, therefore, in leaf litter. Earthworms have been shown to be
adversely affected (population and reproductive effects) at
recommended application rates. There is no information on other soil
or litter arthropods that would be similarly exposed.
The high toxicity to aquatic organisms in laboratory tests is
unlikely to be seen in the field because of the low bioavailability
of sediment-bound residues of carbendazim. However, no information
is available on sediment-living species which would receive the
highest exposure.
11. FURTHER RESEARCH
1. Comparative binding studies of carbendazim to tubulins of
target tissues from various species should be undertaken.
2. Further clarification of the fate of 1,2-diaminobenzene and
bound residues in the environment is needed.
3. The effects of benomyl and carbendazim on sediment-dwelling
organisms needs to be investigated.
12. PREVIOUS EVALUATIONS BY INTERNATIONAL BODIES
Carbendazim was evaluated by the Joint FAO/WHO Meeting on
Pesticide Residues (JMPR) in 1973, 1976, 1977, 1978, 1983 and 1988.
The 1978 meeting agreed that the MRLs for benomyl, carbendazim and
thiophanate-methyl should be combined and expressed as carbendazim.
Carbendazim residues were last evaluated by the 1988 meeting
(FAO/WHO, 1988b) and the MRLs were updated at that time. These MRLs
(expressed as carbendazim) are listed in Table 4. The 1983 meeting
(FAO/WHO, 1985a) evaluated carbendazim toxicology and set the
following NOEL levels and ADI:
Rat: 500 mg/kg diet, equivalent to 25 mg/kg body weight
Dog: 100 mg/kg diet, equivalent to 2.5 mg/kg body weight
Rat: teratology - 30 mg/kg body weight per day (benomyl)
The estimated ADI for carbendazim was established to be 0-0.01
mg/kg body weight.
Carbendazim has not been evaluated by the International Agency
for Research on Cancer (IARC).
REFERENCES
Abdel-Fattah HM, Abdel-Kader M, & Hamida S (1982) Effect of
Bavistin, Cotoran and Curacron on Egyptian soil fungi.
Mycopathologia, 80(2): 101-106.
Albertini S (1989) Influence of different factors on the induction
of chromosome malsegregation in Saccharomyces cerevisiae D61.M by
Bavistan and assessment of its genotoxic property in the Ames test
and in Saccharomyces cerevisiae D7. Mutat Res, 216: 327-340.
Albertini S (1991) Reevaluation of the 9 compounds reported
conclusive positive in yeast Saccharomyces cerevisiae aneuploidy
test systems by the Gene-Tox Program using strain D61.M of
Saccharomyces cerevisiae. Mutat Res, 260: 165-180.
Alverez L (1987) Teratogenicity study of INE-965 (carbendazim) in
rats. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc.,
Haskell Laboratory (Unpublished report No. HLR 281-87).
Amacher DE, Paillet S, & Ray VA (1979) Point mutations at the
thymidine kinase locus in L5178Y mouse lymphoma cells: Application
to genetic toxicological testing. Mutat Res, 64: 391-406.
Ames BN, McCann J, & Yamasaki E (1975) Methods of detecting
carcinogens and mutagens with the Salmonella/mammalian microsome
mutagenicity test. Mutat Res, 31: 347-364.
Ammon HU (1985) Worm toxicity tests using Tubifex tubifex. In:
Comportement et effets secondaires des pesticides dans le sol. Les
Colloques de l'INRA 31, Versailles, 4-8 juin 1984. Paris, Institut
national de la Recherche agronomique, pp 303-317.
Arce GT (1984a) Mutagenicity evaluation in Salmonella typhimurium.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 72-84).
Arce GT (1984b) Mutagenicity evaluation in Salmonella typhimurium.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 71-84).
Arthur MF, Marsh BH, Fadel LC, & Zwick TC (1989a) Anaerobic aquatic
metabolism of [phenyl(U)-14C] benomyl in West Jefferson, Ohio,
pond water and sediment (Battelle Project No. N-0799-8800).
Columbus, Ohio, Battelle, Environmental Sciences Department
(Unpublished report No. AMR 770-87, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Arthur MF, Schweitzer KL, Fadel LC, Marsh BH, & Marsh SS (1989b)
Aerobic aquatic metabolism of [phenyl(U)-14C] benomyl in
Greenville, Mississippi, water and sediment (Battelle Project No.
N-0966-7301). Columbus, Ohio, Battelle, Environmental Sciences
Department (Unpublished report No. AMR 1452-89, prepared for E.I. Du
Pont de Nemours and Co., Inc.).
Bamburg JR, Shooter EM, & Wilson L (1973) Developmental changes in
microtubule protein of the chick brain. Biochemistry, 12: 1476-1482.
Banduhn N & Obe G (1985) Mutagenicity of methyl
2-benzimidazolecarbamate, diethylstilbestrol and estradiol:
Structural chromosomal aberrations, sister-chromatid exchanges,
C-mitoses, polyploidies and micronuclei. Mutat Res, 156: 199-218.
BASF (1975) Dominant lethal study: mice. Ludwigshafen, Germany, BASF
AG.
Baude FJ, Gardiner JA, & Han JC-Y (1973) Characterization of
residues on plants following foliar spray applications of benomyl. J
Agric Food Chem, 21: 1084-1090.
Baude FJ, Pease HL, & Holt RF (1974) Fate of benomyl on field soil
and turf. J Agric Food Chem, 22: 413.
Beavers JB (1985) An acute oral toxicity study in the bobwhite quail
with H-15,607 (MBC). St. Michaels, Maryland, Wildlife International
Ltd (Unpublished report No. HLO 96-85, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Beems RB, Til HP, & van der Heijden CA (1976) Carcinogenicity study
with carbendazim (99% MBC) in mice. The Hague, Central Institute for
Nutrition and Food Research (TNO) (Summary Report No. R-4936,
prepared for Hoechst AG, Frankfurt and BASF AG, Ludwigshafen).
Boeri RL (1988a) Flow through acute toxicity of carbendazim
technical to the eastern oyster, Crassostrea virginica.
Marblehead, Massachusetts, Enseco (Unpublished report No. HLO 15-89,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Boeri RL (1988b) Static acute toxicity of carbendazim technical to
the sheepshead minnow, Cyprinodon variegatus. Marblehead,
Massachusetts, Enseco (Unpublished report No. HLO 12-89, prepared
for E.I. Du Pont de Nemours and Co., Inc.).
Boeri RL (1988c) Static acute toxicity of carbendazim technical to
the mysid, Mysidopsis bahia. Marblehead, Massachusetts, Enseco
(Unpublished report No. HLO 829-88, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Bushway RJ, Hurst HL, Kugabalasooriar J, & Perkins LB (1991)
Determination of carbendazim in blueberries by reversed-phase high
performance liquid chromatography. J Chromatogr, 555: 321-324.
Canton JH (1976) The toxicity of benomyl, thiophanate-methyl, and
BLM to four freshwater organisms. Bull Environ Contam Toxicol, 16:
214-218.
Carter SD, Hess RA, & Laskey JW (1987) The fungicide methyl
2-benzimidazolecarbamate causes infertility in male Sprague-Dawley
rats. Biol Reprod, 37(3): 709-718.
Chiba M & Veres DF (1981) Fate of benomyl and its degradation
compound methyl 2-benzimidazole carbamate on apple foliage. J Agric
Food Chem, 29: 588-590.
Christian MS, Hoberman AM, & Feussner EL (1985) Developmental
toxicity study of carbendazim administered via gavage to New Zealand
white rabbits. Horsham, Pennsylvania, Argus Research Laboratories,
Inc. (Unpublished report No. HLO 515-85, prepared for E.I. Du Pont
de Nemours and Co., Inc.).
Clive D, Johnson KO, Spector JFS, Batson AG, & Moore-Brown MM (1979)
Validation and characterization of the L5178Y/TK +/- mouse
lymphoma mutagen assay system. Mutat Res, 59: 61-108.
Cook ME & Swait AAJ (1975) Effects of some fungicide treatments on
earthworm populations and leaf removal in apple orchards. J Hortic
Sci, 50: 495-499.
County Natwest Woodmac (1992) The fungicide market. London, County
Natwest Woodmac, Agrochemical Service, pp 49-51.
Cox DM & Puck TT (1969) Chromosomal non-disjunction: The action of
colcemid on Chinese hamster ovary cells in vitro. Cytogenetics, 8:
158-169.
Cummings AM, Harris ST, & Rehnberg GL (1990) Effects of methyl
benzimidazole carbamate during early pregnancy in the rat. Fundam
Appl Toxicol, 15: 528-535.
Cummings AM, Ebron-McCoy MT, Rogers JM, Barbee BD, & Harris ST
(1992) Developmental effects of methyl benzimidazole carbamate
following exposure during early pregnancy. Fundam Appl Toxicol, 18:
288-293.
Dashiell OL (1975) Oral LD50 test in guinea pigs (Carbendazim).
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 846-74).
Davidse LC (1973) Antimitotic activity of methyl benzimidazol-2-yl
carbamate (MBC) in Aspergillus nidulans. Pestic Biochem Physiol,
3: 317-325.
Davidse LC (1975) Antimitotic activity of methyl
benzimidazol-2-ylcarbamate in fungi and its binding to cellular
protein. In: Borgers M & de Brabander M ed. Microtubules and
microtubule inhibitors. Amsterdam, Elsevier/North-Holland Publishing
Company, pp 483-495.
Davidse LC & Flach W (1977) Differential binding of methyl
benzimidazol-2-yl carbamate to fungal tubulin as a mechanism of
resistance to this antimitotic agent in mutant strains of
Aspergillus nidulans. J Cell Biol, 72: 174-193.
Davidse LC & Flach W (1978) Interaction of thiabendazole with fungal
tubulin. Biochim Biophys Acta, 543: 82-90.
Delatour P & Besse S (1990) Benzimidazole carbamate d'éthyle: effet
tératogène et présence dans le lait de vache après administration de
thiophanate. Ann Rech Vét, 21: 87-92.
Delp CJ (1980) Coping with resistance to plant disease control
agents. Plant Dis, 64: 652-657.
De Brabander M, van de Veire R, Aerts F, Geuens S, & Hoebeke J
(1976a) A new culture model facilitating rapid quantitative testing
of mitotic spindle inhibition in mammalian cells. J Natl Cancer
Inst, 56: 357-363.
De Brabander M, van de Veire R, Aerts F, Brogers M, & Janssen PAJ
(1976b) The effect of methyl [5(2-thienylcarbonyl)-1H-benzimidazol-
2-yl carbamate (R17934; NSC 238159), a new synthetic antitumoral
drug interfering with microtubules, on mammalian cells cultured in
vitro. Cancer Res, 36: 905-916.
De Vos RH, van Dokkum W, Olthof PDA, Quirijns JK, Muys T, & van der
Poll JM (1984) Pesticides and other chemical residues in Dutch total
diet samples (June 1976 - July 1978). Food Chem Toxicol, 22: 11-21.
Donaubauer HH, Schuetz E, Weigand W, & Kramer M (1982) Repeated dose
(24 month) feeding study for determination of the carcinogenic
effect of HOE 17411 OFAT204 (carbendazim) in mice. Frankfurt,
Germany, Hoechst AG, Pharmaceuticals Research, Toxicology Section
(Unpublished report).
Donovan SD (1982) Mutagenicity evaluation in Salmonella
typhimurium. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 678-82).
Donovan SM (1983) Mutagenicity evaluation in Salmonella
typhimurium. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 797-82).
Dorn E, Schmidt E, Kellner HM, & Leist KH (1983) HOE 017411-14-C
(carbendazim-14-C) metabolic fate in rats and mice, a comparison.
Frankfurt, Germany, Hoechst AG.
Douglas MT & Handley JW (1987) The algistatic activity of
carbendazim technical. Huntingdon, United Kingdom, Huntingdon
Research Centre Ltd (Unpublished report No. DPT 171(g)/871604,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Du Pont (1972) Residue studies - fish: benomyl, MBC, and 2-AB.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report).
Eckert J, Stauffer R, & Hoppman D (1985) Determination of MBC, 4-OH
MBC, and 5-OH MBC in chicken eggs, tissues and feces. Bernville,
Pennsylvania, Enviro-Bio-Tech Ltd (Unpublished report No. AMR
260-84, prepared for E.I. Du Pont de Nemours and Co., Inc.).
Edwards DF (1974a) Skin absorption LD50 (carbendazim). Newark,
Delaware, Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 798-74).
Edwards DF (1974b) Federal Hazardous Substances Act - Eye irritation
test in rabbits (carbendazim). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 799-74).
Ellis WG, Semple JL, Hoogenboom ER, Kavlock RJ, & Zeman FJ (1987)
Benomyl-induced craniocerebral anomalies in fetuses of adequately
nourished and protein-deprived rats. Teratog Carcinog Mutagen, 7:
357-375.
Ellis WG, De Roos F, Kavlock RJ, & Zeman FJ (1988) Relationship of
periventricular overgrowth to hydrocephalus in brains of fetal rats
exposed to benomyl. Teratog Carcinog Mutagen, 8: 377-391.
Evenson DP, Janca FC, & Jost LK (1987) Effects of the fungicide
methyl-benzimidazol-2-yl carbamate (MBC) on mouse germ cells as
determined by flow cytometry. J Toxicol Environ Health, 20: 387-399.
Everhart LP (1979) Benlate dust exposure survey. Wilmington,
Delaware, E.I. Du Pont de Nemours and Co., Inc. (Unpublished report
No. B/TOX 6).
Everhart LP & Holt RF (1982) Potential benlate fungicide exposure
during mixer/loader operations, crop harvest and home use. J Agric
Food Chem, 30: 222-227.
Fahrig R & Seiler JP (1979) Dose and effect of methyl
2-benzimidazolyl-carbamate in the "mammalian spot test", an in vivo
method for the detection of genetic alterations in somatic cells of
mice. Chem-Biol Interact, 26: 115-120.
FAO/WHO (1985a) Benomyl. In: Pesticide residues in food -
Evaluations 1983. Rome, Food and Agriculture Organization of the
United Nations, pp 7-46 (FAO Plant Production and Protection Paper
61).
FAO/WHO (1985b) Carbendazim. In: Pesticide residues in food -
Evaluations 1983. Rome, Food and Agriculture Organization of the
United Nations, pp 89-121 (FAO Plant Production and Protection Paper
61).
FAO/WHO (1988a) Benomyl. In: Pesticide residues in food -
Evaluations 1988. Part I: Residues. Rome, Food and Agriculture
Organization of the United Nations, pp 5-15 (FAO Plant Production
and Protection Paper 93/1).
FAO/WHO (1988b) Carbendazim. In: Pesticide residues in food -
Evaluations 1988. Part I: Residues. Rome, Food and Agriculture
Organization of the United Nations, pp 41-54 (FAO Plant Production
and Protection Paper 93/1).
Fielding M, Barcelo D, Helweg A, Galassi S, Torstensson L, van
Zoonen P, Wolter R, & Angeletti G (1992) Pesticides in ground and
drinking water. Brussels, Commission of the European Communities,
Directorate-General for Science, Research and Development,
Environment and Waste Recycling, 135 pp (Water Pollution Research
Report 27).
Fink R (1975) Eight-day dietary LC50 - Mallard ducks. Sterling,
Virginia, Truslow Farms, Inc. (Unpublished report No. HLO 108-75,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Fiscor G, Bordas S, & Steward SJ (1978) Mutagenicity testing of
benomyl, methyl-2-benzimidazole carbamate, streptozotocin and
N-methyl-N'-nitro-N-nitrosoguanine in Salmonella typhimurium
in vitro and in rodent host-mediated assays. Mutat Res, 51:
151-164.
Fleeker JR & Lacy HM (1977) Photolysis of methyl
2-benzimidazole-carbamate. J Agric Food Chem, 25: 51-55.
Ford LS (1981) Primary skin irritation and sensitization on guinea
pigs. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc.,
Haskell Laboratory (Unpublished report No. HLR 729-81).
Ford LS (1982) Acute skin absorption LD50 test on rabbits
(carbendazim 75% wettable powder). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 822-81).
Freidman PA & Platzer EG (1978) Interaction of anthelmintic
benzimidazoles and benzimidazoles derivatives with bovine brain
tubulin. Biochim Biophys Acta, 544: 605-614.
Galloway SM & Ivett JL (1986) Chemically induced aneuploidy in
mammalian cells in culture. Mutat Res, 167: 89-105.
Gericke D (1977) Test for mutagenicity in bacteria strains in the
absence and presence of a liver preparation. Frankfurt, Germany,
Hoechst AG, Laboratory for Cancer Research (Report No. A/12202).
Gillet C & Roubaud P (1983) Prehatching survival of carp eggs after
treatment during fertilization and early development with the
antimitotic fungicide carbendazim. Water Res, 17: 1343-1348.
Goldenthal EI (1978) Neurotoxicity in hens. Mattawan, Michigan,
International Research and Development Corporation (Unpublished
report No. HLO 27-79, prepared for E.I. Du Pont de Nemours and Co.,
Inc.).
Goldman JM, Rehnberg GL, Cooper RL, Gray LE Jr, Hein JF, & McElroy
WK (1989) Effects of the benomyl metabolite, carbendazim, on the
hypothalamic-pituitary reproductive axis in the male rat.
Toxicology, 57(2): 173-182.
Gooch JJ (1978) Fertility of workers potentially exposed to benomyl.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report No. B/TOX 7).
Goodman NC (1974) Intraperitoneal LD50 test [in rats using
technical carbendazim (less than 98%)]. Newark, Delaware, E.I. Du
Pont de Nemours and Co., Inc., Haskell Laboratory (Unpublished
report No. HLR 845-74).
Goodman NC & Sherman H (1975) Oral LD50 tests (fasted male and
female rats). Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 847-74).
Grandizio AM & Sarver JW (1987) Acute oral toxicity study with
INE-965-261 (Benlate C) in male and female rats. Newark, Delaware,
E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 358-87).
Gray LE, Ostby J, Sigmon R, Ferrell J, Rehnberg G, Linder R, Cooper
R, Goldman J, & Laskey J (1988) The development of a protocol to
assess reproductive effects of toxicants in the rat. Reprod Toxicol,
2: 281-287.
Gray LE, Ostby J, Linder R, Goldman J, Rehnberg G, & Cooper R (1990)
Carbendazim-induced alterations of reproductive development and
function in the rat and hamster. Fundam Appl Toxicol, 15: 281-297.
Guengerich FP (1981) Enzyme induction with Du Pont compounds
H11,201-02 and H10,962-02. Nashville, Tennessee, Vanderbilt
University, School of Medicine (Unpublished report No. HLO-850-81,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Gupta VK & Sharma SD (1989) Degradation of carbendazim in soil by
microorganisms. Indian J Agric Sci, 59: 21-24.
Gustafson DI (1989) Groundwater ubiquity score: A simple method for
assessing pesticide leachability. Environ Toxicol Chem, 8: 339-357.
Hammerschlag RS & Sisler HD (1973) Benomyl and
methyl-2-benzimidazole (MBC): biochemical, cytological, and chemical
aspects of toxicity to Ustilage maydis and Saccharomyces
cerevesiae. Pestic Biochem Physiol, 3: 42-54.
Han JC-Y (1983a) Characterization of residues in bean plants
following foliar spray applications with methyl
2-14C-benzimidazolecarbamate (MBC). Wilmington, Delaware, E.I. Du
Pont de Nemours and Co., Inc. (Unpublished report No. MBC/ME-4).
Han JC-Y (1983b) Anaerobic soil metabolism of 2-14C-benomyl and
methyl 2-14C-benzimidazolecarbamate. Wilmington, Delaware, E.I. Du
Pont de Nemours and Co., Inc. (Unpublished report No. B/ME-44).
Heinemann JM (1971) Acute static fish (rainbow trout) toxicity study
- freshwater (MBC). Vienna, Virginia, Hazelton Laboratories
(Unpublished report No. HLO-150-71, prepared for E.I. Du Pont de
Nemours and Co., Inc.).
Helweg A (1977) Degradation and adsorption of carbendazim and
2-aminobenzimidazole in soil. Pestic Sci, 8: 71-78.
Helweg A (1979) Influence of temperature, humidity, and inoculation
on the degradation of 14C-labelled 2-aminobenzimidazole in soil.
Water Air Soil Pollut, 12: 275-281.
Helweg A (1985) Side effects caused by pesticide combinations. In:
Jensen V, Kjoller A, & Sorensen LH ed. Proceedings of the FEMS
Symposium on Microbial Communities in Soil. Amsterdam, Oxford, New
York, Elsevier Science Publishers, pp 385-395.
Hill EF & Camardese MB (1986) Lethal toxicities of environmental
contaminants and pesticides to Coturnix. Washington, DC, US
Department of the Interior, Fish and Wildlife Service (Fish and
Wildlife Technical Report No. 2).
Hinckle L (1981) Oral LD50 test in rats - EPA proposed guidelines.
Newark, Delaware, E.I. du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 769-81).
Hoechst (1974) Mouse dominant lethal study on MBC. Frankfurt,
Germany, Hoechst AG.
Hofer I, Beck T, & Wallnöfer P (1971) [The influence of the
fungicide benomyl on soil microflora.] Z Pflanzenkr Pflanzenschutz,
78: 399-405 (in German).
Holden HE, Crider PA, & Wahrenburg MG (1980) Mitotic arrest by
benzimidazole analogs in human lymphocyte cultures. Environ Mutat,
2: 67-73.
Hughes DL (1984) Residue study of the fungicide MBC in lactating
dairy cattle. Madison, Wisconsin, Hazelton Laboratories America,
Inc. (Unpublished report No. AMR 429-85, prepared for E.I. Du Pont
de Nemours and Co., Inc.).
Hutton DG (1988) Static acute 48-hours EC50 of carbamic acid,
1H-benzimidazol-2-yl-, methyl ester to fed Daphnia magna. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 638-88).
Hutton DG, Kasprzak DJ, & Priester TM (1984) Laboratory studies of
[2-14C] carbendazim bioconcentration in bluegill sunfish. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 428-84).
Hutton DG, Lieder PH, & Grapel GJ (1986) Chronic toxicity of
carbamic acid, 1H-benzimidazol-2-yl-, methyl ester to Daphnia
magna. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc.,
Haskell Laboratory (Unpublished report No. HLR 241-86).
ILO (1991) Occupational exposure limits for airborne toxic
substances, 3rd ed. Geneva, International Labour Office, 455 pp
(Occupational Safety and Health Series No. 37).
Ireland CM, Gull K, Gutteridge WE, & Pogson CI (1979) The
interaction of benzimidazole carbamates with mammalian microtubule
protein. Biochem Pharmacol, 28: 2680-2682.
Ivett JL (1984) Mutagenicity evaluation of H#15,152 in an in vitro
sister chromatid exchange assay in Chinese hamster ovary (CHO)
cells. Kensington, Maryland, Litton Bionetics, Inc. (Unpublished
report No. HLO 254-84, prepared for E.I. Du Pont de Nemours and Co.,
Inc.).
Janardhan A, Sattur PB, & Sisodia P (1984) Teratology of methyl
benzimidazole carbamate in rats and rabbits. Bull Environ Contam
Toxicol, 33(3): 257-263.
Janardhan A, Rao AB, & Sisodia P (1987) Sub-chronic toxicity of
methyl benzimidazole carbamate in rats. Bull Environ Contam Toxicol,
38(5): 890-898.
Jansz GF & Pomerantz DK (1985) The effect of prenatal treatment with
busulfan on in vitro androgen production by testes from rats of
various ages. Can J Physiol Pharmacol, 63: 1155-1158.
Johnson JD (1988) Determination of the plateau level of bound
[phenyl(U)-14C]carbendazim residues in goat liver. Columbus, Ohio,
Battelle Columbus Division (Unpublished report No. AMR-779-87,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Jotz MM (1980) An evaluation of mutagenic potential of benomyl
employing the L5178Y TK+/- mouse lymphoma assay (SRI Project No.
LSU 7558). Menlo Park, California, SRI International (Unpublished
report prepared for the US Environmental Protection Agency).
Keogh RG & Whitehead PH (1975) Observations on some effects of
pasture spraying with benomyl and carbendazim on earthworm activity
and litter removal from pasture. NZ J Exp Agric, 3: 103-104.
Khan S, Khan NN, & Khan FA (1987) The influence of bavistin and
calixin on carbon dioxide-evolution and the major nutrient status
(nitrogen, phosphorus, potassium) of the soil. Environ Pollut,
47(2): 115-122.
Kirkland JJ (1973) Method for high-speed liquid chromatographic
analysis of benomyl and/or metabolite residue in cow milk, urine,
feces and tissues. J Agric Food Chem, 21: 171-177.
Kirkland JJ, Holt RF, & Pease HL (1973) Determination of benomyl
residues in soils and plant tissues by high-speed cation exchange
liquid chromatography. J Agric Food Chem, 21: 368-371.
Koeter HBWM (1975a) Effect of HOE 17411F (=BAS 3460F) on pregnancy
of the rat. The Hague, Central Institute of Nutrition and Food
Research (TNO) (Unpublished report No. R-4726, prepared for Hoechst
AG, Frankfurt and BASF AG, Ludwigshafen).
Koeter HBWM (1975b) Effect of HOE 17411F (=BAS 3460F) on pregnancy
of the New Zealand white rabbit. The Hague, Central Institute for
Nutrition and Food Research (TNO) (Unpublished report No. R-4881,
prepared for Hoechst AG, Frankfurt and BASF AG, Ludwigshafen).
Kramer M & Weigand W (1971) (HOE 17411 O F) Toxicological
examination. Frankfurt, Germany, Hoechst AG, Pharmaceuticals
Research, Toxicology Section (Unpublished report No. A-00936).
Krechniak J & Klosowska B (1986) The fate of 14C-carbendazim in
rat. Xenobiotica, 16(9): 809-815.
Lamb MJ & Lilly LJ (1980) An investigation of some genetic
toxicological effects of the fungicide benomyl. Toxicology, 17:
83-95.
Litton (1981) Evaluation of HOE 17411 O F AT204 in the primary rat
hepatocyte unscheduled DNA synthesis assay (LBI Project No. 21001).
Kensington, Maryland, Litton Bionetics, Inc. (Unpublished report
prepared for Hoechst AG, Frankfurt).
Lofs-Holmin A (1981) Influence in field experiments of benomyl and
carbendazim on earthworms ( Lumbricidae) in relation to soil
texture. Swed J Agric Res, 11: 141-147.
Lofs-Holmin A (1982) Influence of routine pesticide spraying on
earthworms ( Lumbricidea) in field experiments with winter wheat.
Swed J Agric Res, 12: 121-124.
Marsh BH & Arthur MF (1989) Aerobic metabolism of
[phenyl(U)-14C]benomyl in keyport silt loam. Columbus, Ohio,
Battelle Memorial Institute (Unpublished report No. AMR 1112-88,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Martin DA, Henry JE, & Brock WJ (1987) Closed-patch repeated insult
dermal sensitization study (Buehler Method) with Benlate C fungicide
in guinea-pigs. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 510-87).
Marvin Ch, Birndle ID, Hall CD, & Chiba M (1991) Rapid on-line
precolumn high performance liquid chromatographic method for the
determination of benomyl, carbendazim and aldicarb species in
drinking water. J Chromatogr, 555: 147-154.
Mayer FL & Ellersieck MR (1986) Manual of acute toxicity:
Interpretation and data base for 410 chemicals and 66 species of
freshwater animals. Washington, DC, US Department of the Interior,
Fish and Wildlife Service, pp 45-46 (Resource Publication No. 160).
Mazur AR & Hughes TD (1975) Nitrogen transformations in soil as
affected by the fungicides benomyl, dyrene and maneb. Agron J, 67:
755-758.
Meade AB (1984) Acute contact LD50 toxicity study in honey bees
( Apis mellifera L.) with INE965-212 (MBC). Wilmington, Delaware,
E.I. Du Pont de Nemours and Co., Inc. (Unpublished report No.
ABM-84-5).
Mohr U (1977) Review of liver sections from mice and rats fed with
carbendazim. Hannover, Germany, Medical School, Department of
Experimental Pathology (Unpublished report No. MBC/TOX 6, prepared
for Hoechst AG, Frankfurt and BASF AG, Ludwigshafen).
Monson KD (1985) Metabolism of [2-14C]-carbendazim in the
lactating dairy cow. Wilmington, Delaware, E.I. Du Pont de Nemours
and Co., Inc. (Unpublished report No. AMR 248-84).
Monson KD (1986) Metabolism of [2-14C]-carbendazim in laying hens.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report No. AMR 264-84).
Monson KD (1990) Metabolism of [phenyl(U)-14C]carbendazim in rats.
Wilmington, Delaware, E.I. Du Pont de Nemours and Co., Inc.
(Unpublished report No. AMR-1141-88).
Monson KD (1991) Release and characterization of bound benomyl and
carbendazim metabolites in animal tissues via Raney nickel
desulfurization and acid dehydration. J Agric Food Chem, 39:
1808-1811.
Nakai M, Hess RA, Moore BJ, Guttroff RF, Strader LF, & Linder RE
(1992) Acute and long-term effects of a single dose of the fungicide
carbendazim (methyl 2-benzimidazole carbamate) on the male
reproductive system in the rat. J Androl, 13: 507-518.
Nash SD & Ferenz R (1982) Inhalation median lethal concentration
(LC50 in rats -EPA protocol (carbendazim 75% wettable powder).
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 365-82).
Newsome WH & Collins PG (1987) Enzyme-linked immunosorbent assay of
benomyl and thiabendazole in some foods. J Assoc Off Anal Chem, 70:
1025-1027.
Newsome WH & Shields JB (1981) A radioimmunoassay for benomyl and
methyl 2-benzi midazolecarbamate on food. J Agric Food Chem, 29:
220-222.
Nishiuchi Y & Yoshida K (1974) Toxicity of new agricultural
chemicals to tadpoles. Bull Agric Chem Insp Stn, 14: 66-68.
Osgood C, Zimmering S, & Mason JM (1991) Aneuploidy in Drosophila.
II. Further validation of the FIX and ZESTE genetic test systems
employing female Drosophila melanogaster. Mutat Res, 259: 147-163.
Palawski DU & Knowles CO (1986) Toxicological studies of benomyl and
carbendazim in rainbow trout, channel catfish and bluegills. Environ
Toxicol Chem, 5: 1039-1046.
Pandita TK (1988) Assessment of the mutagenic potential of a
fungicide Bavistin using multiple assays. Mutat Res, 204: 627-643.
Parsons DW & Witt JM (1988) Pesticides in groundwater in the United
States of America: A report of a 1988 survey of State lead agencies.
Corvallis, Oregon, Oregon State University Extension Service.
Peeples JL (1974) Microbial activity in benomyl-treated soil.
Phytopathology, 64: 857-860.
Perreault SD, Jeffay S, Poss P, & Laskey JW (1992) Use of the
fungicide carbendazim as a model compound to determine the impact of
acute chemical exposure during oocyte maturation and fertilization
on pregnancy outcome in the hamster. Toxicol Appl Pharmacol, 114:
225-231.
Priester TM (1984) Hydrolysis of carbendazim [2-14C]. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. AMR 265-84).
Purser D (1987) Carbendazim: behaviour in water. Harrogate, North
Yorkshire, United Kingdom, Hazelton Laboratories (Unpublished report
No. 5220-269/12, prepared for E.I. Du Pont de Nemours and Co.,
Inc.).
Ramakrishna C, Gowda TKS, & Sethunathan N (1979) Effect of benomyl
and its hydrolysis products, MBC and AB on nitrification in flooded
soil. Bull Environ Contam Toxicol, 21: 328-333.
Rashid MA & Mumma RO (1986) Screening pesticides for their
availability to damage bacterial DNA. J Environ Sci Health, B21:
319-334.
Raw F (1962) Studies of earthworm populations in orchards. Ann Appl
Biol, 50: 389-404.
Rehnberg GL, Cooper RL, Goldman JM, Gray LE, Hein JF, & McElroy WK
(1989) Serum and testicular testosterone and androgen binding
protein profiles following subchronic treatment with carbendazim.
Toxicol Appl Pharmacol, 101: 55-61.
Reuzel PGJ, Hendriksen CFM, & Til HP (1976) Long-term (two-year)
toxicity study with carbendazim in beagle dogs. The Hague, Central
Institute for Nutrition and Food Research (TNO) (Unpublished report
prepared for BASF AG, Ludwigshafen and Hoechst AG, Frankfurt).
Rhodes RC (1987) Greenhouse crop-rotation study with
[2-14C]carbendazim. Wilmington, Delaware, E.I. Du Pont de Nemours
and Co., Inc. (Unpublished report No. AMR-495-86).
Rhodes RC & Long JD (1983) Run-off and leaching studies with methyl
2-14C-benzimidazolecarbamate on soil. Wilmington, Delaware, E.I.
Du Pont de Nemours and Co., Inc. (Unpublished report No. MBC/ME-9).
Rhodes RC, Pease HL, & Holt RF (1983) Greenhouse studies on crop
uptake of MBC and 2-AB from soil. Wilmington, Delaware, E.I. Du Pont
de Nemours and Co., Inc. (Unpublished Report No. MBC/ME-5).
Roberts BL & Dorough HW (1984) Relative toxicities on chemicals to
the earthworm Eisenia foetida. Environ Toxicol Chem, 3: 67-78.
Russel LD, Ettlin RA, Sinha Hikim AP, & Clegg ER (1990) Histological
and histopathological evaluation of the testis. Clearwater, Florida,
Cache River Press.
Russell JF (1977) Mutagenicity evaluation of 2-benzimidazolecarbamic
acid, 5-hydroxy-, methyl in Salmonella typhimurium. Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report).
Russell JF (1983) Mutagenicity evaluation in Salmonella
typhimurium. Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 179-83).
Sarver JW (1975) Acute inhalation toxicity - one hour head only
(carbendazim). Newark, Delaware, E.I. Du Pont de Nemours and Co.,
Inc., Haskell Laboratory (Unpublished report No. HLR 58-75).
Scholz J & Weigand W (1972) Carbendazim (HOE 17411 O F, batch
496/I). Toxicological examination. Frankfurt, Germany, Hoechst AG,
Pharmaceuticals Research, Toxicology Section (Unpublished report No.
A-00935).
Seiler JP (1976) The mutagenicity of benzimidazole and benzimidazole
derivatives. VI. Cytogenetic effects of benzimidazole derivatives in
the bone marrow of the mouse and the Chinese hamster. Mutat Res, 40:
339-348.
Seiler JP (1977) Apparent and real thresholds: A study on two
mutagens. In: Scott D, Bridges BA, & Sobels FH ed. Progress in
genetic toxicology. Amsterdam, Elsevier/North-Holland Biomedical
Press, pp 233-238.
Sheir-Neiss F, Nardi RV, Gealt A, & Morris NR (1976) Tubulin-like
protein from Aspergillus nidulans. Biochem Biophys Res Commun, 69:
285-290.
Sherman H (1965) Acute oral test. Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 125-65).
Sherman H (1968) Ninety-day feeding study in rats using a wettable
powder formulation (70% MBC). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 95-68).
Sherman H (1970) Three-month feeding study in dogs using a wettable
powder formulation (50% MBC). Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 283-70).
Sherman H (1972) Long-term feeding studies in rats and dogs with
2-benzimadazole carbamic acid, methyl ester (INE-965) (50% and 70%
MBC wettable powder formulations). Parts I and II. Newark, Delaware,
E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 195-72).
Sherman H & Krauss WC (1966) Acute oral test (carbendazim). Newark,
Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell Laboratory
(Unpublished report No. HLR 99-66).
Sheu CW, Lee JK, Arras CA, Jones RL, & Lavappa KS (1990) The
feasibility of using Chinese hamster as an animal model for
aneuploidy. Environ Mol Mutagen, 16: 320-323.
Shirasu Y, Moriya M, & Kato K (1977) Mutagenicity testing on
fungicide 1991 in microbial systems. Tokyo, Institute of
Environmental Toxicology, Kodaira Laboratories (Unpublished report
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Shukla Y, Antony M, & Mehrotra NK (1989) Studies on g-glutamyl
transpeptidase in rodents exposed to benomyl. Bull Environ Contam
Toxicol, 42: 301-306.
Siebert D, Zimmermann FK, & Lemperle E (1970) Genetic effects of
fungicides. Mutat Res, 10: 533-543.
Speakman JB & Nirenberg HI (1981) Mutagenicity of methyl
benzimidazole-2-yl carbamate (MBC) towards Aspergillus nidulans
(Eidam) Winter and Cladosporium cucumerinum Ellis and Arth. Mutat
Res, 88: 45-51.
Stadler JC (1986) One-year feeding study in dogs with carbendazim.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 291-86).
Staub T & Sozzi D (1984) Fungicide resistance: A continuing
challenge. Plant Dis, 68: 1026-1031.
Stringer A & Lyons C (1974) The effect of benomyl and
thiophanate-methyl on earthworm populations in apple orchards.
Pestic Sci, 5: 189-196.
Stringer A & Wright MA (1973) The effect of benomyl and some related
compounds on Lumbricus terrestris and other earthworms. Pestic
Sci, 4: 165-170.
Styles JA & Garner R (1974) Benzimidazolecarbamate methyl ester -
evaluation of its effects in vivo and in vitro. Mutat Res, 26:
177-187.
Til HP, van den Muelen HC, Feron VJ, Seinen W, & de Groot
AP (1972) Sub-chronic (90-day) toxicity study with W17411 in beagle
dogs. The Hague, Central Institute for Nutrition and Food Research
(TNO) (Unpublished report No. R-3920, prepared for Hoechst AG,
Frankfurt).
Til HP, Koellen C, & van der Heijden CA (1976a) Combined chronic
toxicity and carcinogenicity study with carbendazim in rats. The
Hague, Central Institute for Nutrition and Food Research (TNO)
(Unpublished report prepared for BASF AG, Ludwigshafen and Hoechst
AG, Frankfurt).
Til HP, Koeter HBWM, & van der Heijden CA (1976b) Multigeneration
study with carbendazim in rats. The Hague, Central Institute for
Nutrition and Food Research (TNO) (Unpublished report No. R-5024,
prepared for Hoechst AG, Frankfurt).
Til HP, Beems RB, & de Groot AP (1981) Determination of the acute
oral toxicity of carbendazim in mice. The Hague, Central Institute
for Nutrition and Food Research (TNO) (Unpublished report No.
A-21604, prepared for BASF AG, Ludwigshafen).
Tong C (198la) Hepatocyte primary culture/DNA repair assay on
compound l1, 201-01 (MBC) using mouse hepatocytes in culture.
Valhalla, New York, Naylor Dana Institute (Unpublished report No.
HLO 743-81, prepared for E.I. Du Pont de Nemours and Co., Inc.).
Tong C (198lb) Hepatocyte primary culture/DNA repair assay on
compound 11,201-01 (MBC) using rat hepatocytes in culture. Valhalla,
New York, Naylor Dana Institute (Unpublished report No. HLO 744-81,
prepared for E.I. Du Pont de Nemours and Co., Inc.).
Torstensson L & Wessen B (1984) Interactions between the fungicide
benomyl and soil microorganisms. Soil Biol Biochem, 16: 445-452.
Van Faassen HG (1974) Effect of the fungicide benomyl on some
metabolic processes and on numbers of bacteria and actinomycetes in
the soil. Soil Biol Biochem, 6: 131-133.
Van Gestel CAM (1992) Validation of earthworm toxicity tests by
comparison with field studies: A review of benomyl, carbendazim,
carbofuran, and carbaryl. Ecotoxicol Environ Saf, 23: 221-236.
Van Gestel CAM, Dirven-van Breeman EM, Baerselman R, Emans HJB,
Janssen JAM, Postuma R, & van Vliet PJM (1992) Comparison of
sublethal and lethal criteria for nine different chemicals in
standardized toxicity tests using the earthworm Eisenia andrei.
Ecotoxicol Environ Saf, 23: 206-220.
Vick DA & Brock WJ (1987a) Primary dermal irritation study with
Benlate C fungicide in rabbits. Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 299-87).
Vick DA & Brock WJ (1987b) Acute dermal toxicity study of Benlate C
fungicide in rabbits. Newark, Delaware, E.I. Du Pont de Nemours and
Co., Inc., Haskell Laboratory (Unpublished report No. HLR 303-87).
Vick DA & Valentine R (1987) Primary eye irritation study with
Benlate C fungicide in rabbits. Newark, Delaware, E.I. Du Pont de
Nemours and Co., Inc., Haskell Laboratory (Unpublished report No.
HLR 287-87).
Vonk JW, Adema OMM, & Barug D (1986) Comparison of the effects of
several chemicals on microorganisms, higher plants and earthworms.
In: Assink JW & van den Brink WG ed. Contaminated soil. Dordrecht,
Boston, Lancaster, Martinus Nijhoff Publishers, p 191.
Wainwright M & Pugh GJF (1974) The effect of fungicides on certain
chemical and microbial properties of soils. Soil Biol Biochem, 6:
263-267.
Wallin M, Friden B, & Billger M (1989) Studies of the interaction of
chemicals with microtubule assembly in vitro can be used as an
assay for detection of cytotoxic chemicals and possible inducers of
aneuploidy. Mutat Res, 201: 303-311.
Waterer JC (1980) Chinese hamster ovary cell assay for mutagenicity.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 660-88).
Weeks RE & Hedrick HG (1975) Influence of a systemic fungicide on
oxygen uptake by soil microorganisms. Soil Sci, 119: 280-284.
Wetzel JW & Hutton DG (1984) 96-Hour LC50 to bluegill sunfish.
Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 401-84).
Whittaker SG, Moser SF, Maloney DH, Piegorsch WW, Resnick MA, &
Fogel S (1990) The detection of mitotic and meiotic chromosome gain
in the yeast Saccharomyces cerevisiae: Effects of methyl
benzimidazol-2-yl carbamate, methyl methanesulfonate, ethyl
methanesulfonate, dimethyl sulfoxide, propionitrile and
cyclophosphamide monohydrate. Mutat Res, 242: 231-258.
WHO (1993) Environmental Health Criteria 148: Benomyl. Geneva, World
Health Organization.
Wood CK (1982) Long-term feeding study with 2-benzimidazole-
carbamate, methyl ester (< 99% MBC, INE-965) in mice. Parts I and
II. Newark, Delaware, E.I. Du Pont de Nemours and Co., Inc., Haskell
Laboratory (Unpublished report No. HLR 70-82).
Yarden O, Aharonson N, & Katan J (1987) Accelerated microbial
degradation of methyl benzimidizol-2-ylcarbamate in soil and its
control. Soil Biol Biochem, 19(6): 735-739.
RESUME ET CONCLUSIONS
1. Résumé
1.1 Identité, propriétés physiques et chimiques et méthodes d'analyse
Le carbendazime, un solide cristallin blanc, est un fongicide
endothérapique qui appartient à la famille du benzimidazole. Son
point de fusion est d'environ 250 °C et sa tension de vapeur est <
1 x 10-7 Pa (< 1 x 10-9 mbar) à 20 °C. Le carbendazime est
pratiquement insoluble dans l'eau, sa solubilité n'étant que de 8
mg/litre à pH 7 et à 20 °C. Il est stable dans les conditions
normales de stockage.
L'analyse des résidus de même que celle des prélèvements
effectués dans l'environnement comportent une extraction au moyen
d'un solvant organique et une purification de l'extrait par partage
liquide-liquide. Le dosage de ces résidus peut s'effectuer par
chromatographie en phase liquide à haute performance ou par titrage
immunologique.
1.2 Sources d'exposition humaine et environnementale
Le carbendazime est, parmi les fongicides de la famille du
benzimidazole, celui qui est le plus largement utilisé. Il est
présenté sous forme de dispersion ou de suspension aqueuse, de
granulés fluants dispersables dans l'eau et de poudre mouillable.
1.3 Transport, distribution et transformation dans l'environnement
Le carbendazime est d'ailleurs le produit de transformation
dans l'environnement du bénomyl, un autre fongicide
benzimidazolique; la réaction est rapide avec une demi-vie
respective de 2 et 19 heures dans l'eau et le sol. On peut donc
utiliser les résultats des études sur le bénomyl ou sur le
carbendazime pour l'évaluation des effets sur l'environnement.
Dans l'environnement, le carbendazime se décompose avec une
demi-vie de 6 à 12 mois sur le sol nu, de 3 à 6 mois sur le gazon et
de 2 à 25 mois dans l'eau en aérobiose et en anaérobiose,
respectivement. Le carbendazime est principalement décomposé par les
microorganismes. Le 2-aminobenzimidazole (2-AB) en est l'un des
principaux produits de dégradation et il est à son tour décomposé
par les microorganismes. Lors de la décomposition du bénomyl marqué
au 14C sur le noyau phényle, on a constaté que 9% seulement du
carbone-14 était éliminé sous forme de CO2 en une année
d'incubation. Le carbone-14 restant était principalement récupéré
sous forme de carbendazime et de résidus liés. L'étude de la
destinée d'un éventuel produit de dégradation (1,2-diaminobenzène)
pourrait peut-être permettre de mieux définir la voie de dégradation
des fongicides benzimidazoliques dans l'environnement.
Des études effectuées sur le terrain ou sur colonne ont montré
que le carbendazime restait dans les couches superficielles du sol.
On n'a pas mesuré l'adsorption du carbendazime dans le sol, mais on
pense qu'elle doit être aussi forte que dans le cas du bénomyl, avec
des valeurs de Koc allant de 1000 à 3600. Les valeurs de log Kow
sont respectivement de 1,36 et de 1,49 pour le bénomyl et le
carbendazime.
Un modèle de criblage basé sur les données d'adsorption et de
persistance n'a pas révélé de risque de lessivage. Cette observation
est corroborée par des analyses d'eau de pluie effectuées aux
Etats-Unis, analyses qui n'ont pas permis de déceler ce composé dans
l'un quelconque des 212 puits étudiés (la limite de détection n'a
pas été précisée). On estime que le bénomyl et le carbendazime
entraînés par ruissellement correspondent uniquement à la fraction
adsorbée aux particules de sol; d'ailleurs ces composés sont sans
doute fortement adsorbés aux sédiments présents dans l'environnement
aquatique.
Le carbendazime est hydrolysé en 2-AB. Ce produit en constitue
également le principal métabolite dans le sol et les végétaux.
Chez l'animal, le carbendazime est métabolisé en
(5-hydroxy-1H-benzimidazole-2-yl)-carbamate (5-HBC) et autres
métabolites polaires, qui sont rapidement excrétés. On n'a pas
observé d'accumulation de carbendazime dans aucun système
biologique.
1.4 Concentrations dans l'environnement et exposition humaine
Il ne semble pas qu'il existe de données résultant d'une
surveillance du carbendazime dans l'environnement. Toutefois on peut
récapituler comme suit les données d'études portant sur la destinée
écologique de ce produit.
Comme le bénomyl et le carbendazime restent stable pendant des
semaines sur les végétaux, ils peuvent être ingérés par des
organismes qui se nourrissent de feuilles mortes. Des résidus de
carbendazime peuvent subsister jusqu'à 3 ans dans le sol et les
sédiments. Toutefois la forte adsorption du carbendazime aux
particules de sol et de sédiments réduit l'exposition des organismes
terrestres et aquatiques.
Ce sont les résidus de bénomyl et de carbendazime présents sur
les cultures vivrières qui constituent la principale source
d'exposition de la population humaine dans son ensemble. L'analyse
de l'exposition par voie alimentaire qui a été effectuée aux
Etats-Unis (bénomyl plus carbendazime) et aux Pays-Bas (carbendazime
seul) a montré que la quantité moyenne ingérée était
vraisemblablement de l'ordre d'une dixième de la dose journalière
acceptable (DJA) qui est, pour le bénomyl, de 0.02 mg/kg de poids
corporel et pour le carbendazime, de 0,01 mg/kg de poids corporel.
L'exposition professionnelle au cours de la production est
inférieure à la valeur-seuil établie pour le bénomyl. Les ouvriers
agricoles qui préparent les mélanges, effectuent le remplissage ou
retournent dans les champs traités par du bénomyl ou du
carbendazime, courent un risque d'exposition cutanée correspondant à
quelques milligrammes de bénomyl par heure. Le port de dispositifs
de protection permettrait de réduire encore cette exposition. En
outre, étant donné que l'absorption percutanée est vraisemblablement
faible, il est très peu probable que le carbendazime puisse avoir
des effets toxiques généraux sur les populations humaines en étant
absorbé par cette voie.
1.5 Cinétique et métabolisme
Le carbendazime est bien absorbé (à hauteur de 80 à 85%) après
ingestion mais il l'est beaucoup moins après exposition par voie
cutanée. Une fois absorbé, le carbendazime donne naissance dans
l'organisme à de nombreux métabolites. Les principaux d'entre eux
sont le 5-HBC et les 5,6-HOBC-N-oxydes. Le 5,6-DHBC-S et le
5,6-DHBC-G constituent des métabolites mineurs.
D'après la distribution tissulaire du carbendazime, il n'y a
pas de bioconcentration. Chez le rat, la concentration la plus
élevée après administration par voie orale (< 1% de la dose), a été
constatée dans le foie. Le produit se retrouvait sous forme de
carbendazime dans les mitochondries, de 5-HBC dans le cytosol et de
2-AB dans les microsomes. On a également retrouvé le carbendazime et
ses métabolites dans les reins de poulets et de vaches après
exposition, mais il ne se trouvait pas dans les autres tissus en
quantités appréciables. Après administration de carbendazime à des
vaches laitières, on a retrouvé de petites quantités de 5-HBC et de
4-HBC dans le lait.
Soixante-douze heures après avoir été administré par voie orale
à des rats, le carbendazime était excrété dans leurs urines.
Chez les rats et les souris, de fortes doses de carbendazime
administrées, soit mêlées à la nourriture, soit par gavage, ont eu
un effet sur certaines enzymes microsomiques du foie. Ainsi il y a
eu induction de la styrène-7,8-hydrolase et de l'époxyde-hydrolase
mais réduction de l'activité de la 7-hydroxycoumarine-O-déséthylase.
On a également constaté une induction de l'activité de la
glutation-S-transférase du cytosol.
1.6 Effets sur les mammifères de laboratoire et sur les systèmes
d'épreuve in vitro
1.6.1 Exposition unique
Le carbendazime présente une faible toxicité aiguë. Les valeurs
de la DL50 vont de > 2000 à 15 000 mg/kg chez toutes sortes
d'animaux d'expérience exposés par diverses voies. Toutefois on n'a
pas noté, après exposition unique, d'effets nocifs sensibles sur la
reproduction (voir Section 1.6.5).
1.6.2 Exposition de brève durée
Administré en mélange à la nourriture pendant des périodes
allant jusqu'à 90 jours, le carbendazime a entraîné une légère
modification du poids du foie chez des rattes qui en avaient reçu
une dose quotidienne de 360 mg/kg de poids corporel. Une étude de 90
jours au cours de laquelle du carbendazime a été administré par
gavage à des rats, a permis de fixer à 16 mg/kg/jour la dose sans
effets observables, basée sur l'hépatotoxicité. Les études
d'alimentation de courte durée effectuées sur des chiens se sont
révélées insuffisantes pour l'établissement d'une dose sans effets
observables. Une étude de 10 jours, au cours de laquelle de lapins
ont été exposés au carbendazime par voir cutanée n'a pas permis de
mettre en évidence une toxicité générale à la seule dose étudiée
(200 mg/kg).
1.6.3 Irritation et sensibilisation au niveau de la peau et des
yeux
L'application du produit sur l'épiderme de lapins ou de cobayes
n'a produit aucune irritation ni sensibilisation de la peau. Chez le
lapin, l'instillation oculaire n'a produit qu'une irritation légère
à modérée de la conjonctive.
1.6.4 Exposition de longue durée
A l'issue d'une étude d'alimentation où des rats mâles et
femelles avaient reçu 2500 mg de carbendazime/kg de nourriture, on a
constaté une réduction du nombre des érythrocytes ainsi que du taux
d'hémoglobine et de l'hématocrite. Aucun effet toxique sur le foie
n'a été constaté. Parmi les rats mâles qui avaient reçu du
carbendazime à raison de 2500 mg ou davantage/kg de nourriture, on
notait une augmentation peu importante du nombre de cas d'atrophie
testiculaire diffuse et de prostatite. Chez le rat, la dose sans
effets observables est de 500 mg/kg de nourriture. Des chiens qui
avaient reçu pendant une année ou davantage une nourriture contenant
500 mg de carbendazime/kg, ont présenté une élévation du taux de
cholestérol sérique et de l'activité de la phosphatase alcaline
accompagnée d'autres indices d'une hépatotoxicité. Chez le chien, la
dose sans effets observables a été fixée à 300 mg/kg de nourriture.
Chez des souris mâles et femelles qui avaient reçu une
nourriture contenant 5000 mg de carbendazime/kg de nourriture on a
observé une augmentation du poids absolu du foie. Chez des souris
mâles ayant reçu 1500 mg de carbendazime/kg de nourriture on a
également observé une importante hypertrophie centrilobulaire avec
nécrose et hypertrophie générale du foie.
1.6.5 Reproduction, embryotoxicité et tératogénicité
Lors d'une étude de reproduction portant sur trois générations,
on a administré à des rats du carbendazime à des doses allant
jusqu'à 500 mg/kg de nourriture inclusivement; aucun effet nocif n'a
été constaté sur la reproduction de ces animaux. En revanche, après
administration du même produit par gavage pendant 85 jour à raison
de 200 mg/kg/jour, on constatait une réduction de la fertilité des
mâles. A la dose de 50 mg/kg de poids corporel, la même étude a
révélé une réduction sensible du nombre de spermatozoïdes
épididymaires.
Après administration par voie orale d'une dose unique de
carbendazime et examen histologique, on a constaté que dès les deux
premiers jours, il y avait une forte perturbation de la
spermatogénèse avec occlusion des canaux efférents et augmentation
du poids des testicules à la dose de 100 mg/kg de poids corporel.
Aucun effet n'a été observé au cours de la même étude à la dose de
50 mg/kg. Ces effets ont persisté jusqu'au 70ème jour chez les rats
ayant reçu 400 mg/kg.
Administré à des rats en doses quotidiennes supérieures à 10
mg/kg du 7ème au 16ème jour de la gestation, le carbendazime a
provoqué une augmentation du nombre de malformations et d'anomalies
chez le rat. On constatait également une légère diminution du taux
de nidation chez des lapines qui en avaient reçu respectivement 20
ou 125 mg/kg en doses quotidiennes du 7ème au 19ème jour de la
gestation ainsi qu'un accroissement de la fréquence des résorptions
à la dose quotidienne de 125 mg/kg. Le composé s'est révélé toxique
pour la mère, chez la ratte, à la dose quotidienne de 20 mg/kg et
chez la lapine à la dose quotidienne de 125 mg/kg.
Outre que le taux de gravidité était en diminution et que la
fréquence des résorptions précoces était en augmentation chez le
rat, il y avait une réduction sensible du poids des foetus aux doses
quotidiennes respectives de 20 et 90 mg/kg, et un accroissement
également notable des malformations à la dose quotidienne de 90
mg/kg. Il s'agissait essentiellement d'hydrocéphalie, de
microphthalmie, d'anophthalmie, de malformations des omoplates et de
la partie axiale du squelette (fusions vertébrales, costales et
sternébrales, exencéphalie, hémivertèbres et hyperplasie costale).
Toutefois on n'a pas constaté de malformations importantes chez le
lapin.
1.6.6 Mutagénicité et autres points d'aboutissement des effets
toxiques
Les épreuves effectuées in vitro et in vivo sur des
systèmes mammaliens et non-mammaliens ainsi que sur des cellules
somatiques ou germinales montrent que le carbendazime n'interagit
pas avec l'ADN, qu'il ne provoque pas de mutations ponctuelles et ne
cause pas de mutation des cellules germinales.
Toutefois le carbendazime provoque des aberrations dans le
nombre des chromosomes (aneuploïdie et/ou polyloïdie) dans les
systèmes d'épreuve (tant in vitro qu' in vivo).
1.6.7 Cancérogénicité
L'administration de bénomyl et de carbendazime mêlés à la
nourriture a provoqué chez des souris CD-1 et Swiss axéniques une
augmentation dans l'incidence des tumeurs hépatocellulaires.
Une étude de cancérogénicité a été effectuée avec le
carbendazime sur des souris CD-1; elle a révélé une augmentation
statistiquement significative et liée à la dose dans l'incidence des
néoplasmes hépatocellulaires chez les femelles. Il y avait également
une augmentation statistiquement significative à la dose moyenne
(1500 mg/kg) de ces anomalies chez les mâles, augmentation qui n'a
pas été constatée aux doses élevées en raison du taux élevé de
mortalité. Une autre étude de cancérogénicité portant sur le
carbendazime a été effectuée chez une souche génétiquement
apparentée de souris Swiss axéniques et exogames à des doses de 0,
150, 300 et 1000 mg/kg (la dernière dose étant portée 5000 mg/kg au
cours de l'étude); elle a révélé un accroissement dans l'incidence
de l'ensemble de adénomes et carcinomes hépatocellulaires. Une
troisième étude, effectuée cette fois sur des souris NMRKf à des
doses respectives de 0, 50, 150, 300 et 1000 mg/kg de nourriture (la
dernière dose étant également portée à 5000 mg/kg au cours de
l'étude) n'a cette fois pas fait ressortir d'effets cancérogènes. Le
bénomyl ou le carbendazime ont provoqué indifféremment des tumeurs
hépatiques chez deux souches de souris (CD-1 et axéniques), souches
qui présentent toutes deux un taux élevé de tumeurs hépatiques
spontanées. En revanche, le carbendazime n'est pas cancérogène chez
les souris NMRKf dont le taux de tumeurs spontanées du foie est
faible.
Les études de cancérogénicité portant sur le bénomyl et le
carbendazime ont donné des résultats négatifs chez le rat.
1.6.8 Mécanisme de la toxicité - mode d'action
On pense que les effets biologiques du bénomyl et du
carbendazime résultent de leur interaction avec les microtubules
cellulaires. Ces structures interviennent dans des fonctions aussi
importantes que la division cellulaire, qui est inhibée par ces deux
substances. La toxicité du bénomyl et du carbendazime pour les
mammifères est donc liée à une perturbation des fonctions du système
microtubulaire.
Comme les autres dérivés du benzimidazole, le bénomyl et le
carbendazime sont plus ou moins toxiques selon les espèces. Cette
sélectivité toxicologique s'explique au moins en partie par le fait
que les deux substances ne se lient pas de la même manière aux
tubulines des espèces visées et des espèces non-visées.
1.7 Effets sur l'homme
Aucun effet nocif pour la santé humaine n'a été signalé.
1.8 Effets sur les autres êtres vivants, en laboratoire et dans leur
milieu naturel
Le carbendazime n'a guère d'effet sur l'activité microbienne du
sol aux doses d'emploi recommandées. On a cependant signalé
l'existence d'effets nocifs vis-à-vis de certains groupes de
champignons.
On a calculé que la CE50 à 72 heures, fondée sur la
croissance totale, pour les algues bleu-vert du genre Selenastrum
capricornutum, était égale à 1,3 mg/litre; la concentration sans
effets observables était de 0,5 mg/litre. La toxicité du
carbendazime pour les invertébrés aquatiques et les poissons varie
largement, les valeurs de la CL50 à 96 heures allant de 0,007
mg/litre pour des poissons chats du genre Ictalurus, à 5,5
mg/litre pour Lepomis machrochirus. Une étude de 21 jours sur la
daphnie Daphnia magna, a révélé que, à la dose de 0,025 mg/litre,
le déclenchement de la reproduction était sensiblement retardé; la
concentration sans effets observables a été fixée à 0,013 mg/litre
dans ce cas.
Le carbendazime se montre toxique pour les lombrics dans des
expériences de laboratoire reproduisant les conditions réelles
d'exposition qui résultent d'une utilisation dans les conditions
recommandées sur le terrain. Il est "relativement non toxique" pour
les abeilles et peu toxique pour les oiseaux.
2. Conclusions
Le bénomyl, un composé voisin du carbendazime, provoque une
sensibilisation cutanée chez l'homme. Ce composé et le carbendazime
lui-même ne présentent guère de risques d'intoxication aiguë pour
l'homme. Etant donné les conditions actuelles d'exposition et le
faible taux d'absorption percutanée de ces deux composés, il est
improbable qu'ils provoquent des effets toxiques généralisés dans la
population ou chez les personnes exposées pour des raisons
professionnelles. Ces conclusions résultent des données relatives à
l'animal et dans une moindre mesure des données relatives à l'homme;
elles reposent sur la connaissance du mode d'action du carbendazime
et du bénomyl, tant chez les espèces visées que chez les autres.
Grâce à une meilleure connaissance du mécanisme de la toxicité
du bénomyl et du carbendazime chez les mammifères, on pourra
peut-être mieux déterminer quelles sont les doses sans effets
observables. Des études portant sur la liaison de ces composés aux
tubulines des cellules cibles (tissus testiculaires et
embryonnaires) faciliteront sans doute les comparaisons
interspécifiques.
Le carbendazime est fortement adsorbé aux matières organiques
du sol et il y persiste pendant des périodes allant jusqu'à trois
ans. Il persiste également sur le feuillage et se retrouve par
conséquent dans les feuilles mortes. On a montré que les lombrics
pouvaient souffrir (dans leur effectif comme dans leur reproduction)
de l'action de ces composés aux doses d'emploi recommandées. On ne
possède aucune données sur les autres arthropodes qui vivent dans le
sol ou sur les débris organiques et qui pourraient être exposés de
la même manière.
Il est improbable que la forte toxicité que, selon les études
de la laboratoire, le carbendazime présente pour les organismes
aquatiques, s'observe également dans le milieu naturel, du fait de
la faible biodisponibilité des résidus de ce composé liés aux
sédiments. Toutefois on ne possède aucune donnée sur les espèce
sédimenticoles, qui seraient les plus exposées.
RESUMEN Y CONCLUSIONES
1. Resumen
1.1 Identidad, propiedades físicas y químicas y métodos analíticos
La carbendazima es una sustancia sólida cristalina de color
blanco y acción fungicida sistémica que pertenece a la familia del
bencimidazol. Su punto de fusión es de unos 250 °C y tiene una
presión de vapor de < 1 x 10-7 Pa (< 1 x 10-9 mbar). Es
prácticamente insoluble en agua (8 mg/litro) a pH 7 y 20 °C. Es un
compuesto estable en condiciones de almacenamiento normales.
Los análisis de las muestras de residuos y del medio ambiente
se realizan mediante extracción con un disolvente orgánico y
purificación del extracto obtenido utilizando un procedimiento de
reparto líquido-líquido. La valoración de los residuos se puede
realizar mediante cromatografía líquida de alto rendimiento o
inmunoensayo.
1.2 Fuentes de exposición humana y ambiental
La carbendazima es la sustancia más ampliamente utilizada de la
familia de los fungicidas bencimidazólicos. Está formulada como
dispersión acuosa, suspensión acuosa, gránulos fluidos dispersables
en agua y polvo humectable.
1.3 Transporte, distribución y transformación en el medio ambiente
En el medio ambiente, el benomilo se transforma rápidamente en
carbendazima, con una semivida de 2 y 19 h en el agua y en el suelo,
respectivamente. Por consiguiente, para la evaluación de los efectos
sobre el medio ambiente son importantes los datos obtenidos de los
estudios realizados con ambos compuestos.
La carbendazima se descompone en el medio ambiente, con una
semivida de 6 a 12 meses en el suelo desnudo, de 3 a 6 meses en el
césped y de 2 y 25 meses en el agua en condiciones aerobias y
anaerobias, respectivamente. La descomposición se debe sobre todo a
la acción de los microorganismos; el principal producto de su
degradación es el 2-aminobencimidazol (2-AB), que luego se
descompone de nuevo, también por la actividad microbiana. En la
descomposición del benomilo marcado con un grupo fenilo con 14C,
sólo el 9% del 14C formó CO2 durante un año de incubación,
mientras que el resto del 14C se recuperó sobre todo como
carbendazima y en productos unidos a residuos. El destino de un
posible producto de degradación (1,2-diaminobenceno) puede aclarar
ulteriormente la vía de degradación de los fungicidas
bencimidazólicos en el medio ambiente.
En estudios de campo y de columna se ha puesto de manifiesto
que la carbendazima queda retenida en la capa superficial del suelo.
Aunque no se dispone de datos sobre su adsorción en el suelo, es
probable que ésta sea tan intensa como la del benomilo (los valores
de Koc oscilan entre 1000 y 3600). Los valores del log Kow para
el benomilo y la carbendazima son respectivamente 1,36 y 1,49.
En la evaluación realizada en un modelo de selección, basado en
datos de adsorción y persistencia, se puso de manifiesto que no
había riesgo de lixiviación. En los Estados Unidos se han efectuado
análisis de agua de pozos que confirman esto, puesto que no se
encontraron trazas de carbendazima en ninguno de los 212 pozos
muestreados (no se dispone del límite de detección). Es de suponer
que la escorrentía superficial del benomilo y la carbendazima se
deba solamente al fungicida adsorbido en las partículas del suelo y
que en el medio acuoso estén fuertemente adsorbidos en los
sedimentos.
La carbendazima se hidroliza a 2-AB. Este es también el
metabolito primario en el suelo y las plantas.
En los sistemas animales se metaboliza a
(5-hidroxi-1H-bencimidazol-2-il)-carbamato (5-HBC) y otros
metabolitos polares, que se excretan rápidamente. No se ha observado
que la carbendazima se acumule en ningún sistema biológico.
1.4 Niveles medioambientales y exposición humana
No parece que se disponga de datos de vigilancia ambiental para
la carbendazima. Sin embargo, los estudios realizados sobre su
destino en el medio ambiente pueden resumirse como sigue.
Puesto que el benomilo y la carbendazima se mantienen estables
en las plantas durante varias semanas, pueden pasar a los organismos
que se alimentan de las hojas caídas. El suelo y los sedimentos
pueden conservar residuos de carbendazima hasta tres años. Sin
embargo, la fuerte adsorción de este compuesto en las partículas del
suelo y en los sedimentos reduce la exposición de los organismos
terrestres y acuáticos.
La principal fuente de exposición para la población humana
general se debe a los residuos de benomilo y carbendazima en los
cultivos alimentarios. En análisis de la exposición a través de los
alimentos realizados en los Estados Unidos (benomilo y carbendazima
combinados) y en los Países Bajos (con carbendazima) se obtuvo una
ingesta media prevista de alrededor del 10 por ciento de la ingesta
diaria admisible (IDA) recomendada, de 0,02 mg/kg de peso corporal
para el benomilo y de 0,01 mg/kg de peso corporal para la
carbendazima.
La exposición profesional durante el proceso de fabricación es
inferior al valor umbral límite. Se considera que los trabajadores
agrícolas que se ocupan de mezclar y cargar los plaguicidas o que
entran en campos tratados con benomilo sufren exposición cutánea a
unos mg de benomilo por hora. Esta forma de exposición se podría
reducir con algún tipo de protección. Por otra parte, puesto que la
absorción cutánea que cabe prever es baja, la probabilidad de que el
benomilo tenga efectos tóxicos sistémicos a través de esta vía es
muy escasa.
1.5 Cinética y metabolismo
La carbendazima se absorbe bien tras la exposición oral
(80-85%), pero mucho menos después de una exposición cutánea. Una
vez absorbida, se metaboliza formando numerosos compuestos en el
organismo. Los principales metabolitos son el 5-HBC y los óxidos de
5,6-HOBC-N; otros menos son importantes el 5,6-DHBC-S y el
5,6-DHBC-G.
La distribución de la carbendazima en los tejidos demuestra la
ausencia de bioconcentración. En la rata, la concentración más
elevada tras su administración oral (< 1% de la dosis) se produjo
en el hígado. Apareció carbendazima en las mitocondrias, el
metabolito 5-HBC en el citosol y el 2-AB en los microsomas. Aunque
se encontraron también carbendazima y sus metabolitos en el riñón de
gallinas y de vacas, no se detectaron niveles significativos en
otros tejidos. Tras suministrar carbendazima con el pienso a vacas
lactantes, se encontraron pequeñas cantidades de 5-HBC y 4-HBC en la
leche.
La carbendazima se excreta en la orina y las heces en un plazo
de 72 h tras la dosificación oral en ratas.
En ratas y ratones, la administración de dosis altas de
carbendazima en la dieta o con sonda afectó a ciertas enzimas de los
microsomas. Se produjo inducción de la estireno-7,8-hidrolasa y de
la epóxido-hidrolasa, mientras que se redujo la actividad de la
hidroxicumarín-O-deetilasa. También se indujo la actividad de la
glutatión-S-transferasa citosólica.
1.6 Efectos en los mamíferos de laboratorio y en sistemas de prueba
in vitro
1.6.1 Exposición única
La carbendazima tiene una toxicidad aguda baja. Los valores de
la DL50 oscilan entre > 2000 y 15 000 mg/kg en una amplia
variedad de animales de experimentación y de vías de administración.
Sin embargo, se han observado efectos reproductivos adversos
significativos en la salud tras una exposición única (véase la
sección 1.6.5).
1.6.2 Exposición de corta duración
La administración de carbendazima en la dieta durante un
período de hasta 90 días produjo efectos leves en el peso del hígado
de las ratas hembras expuestas a 360 mg/kg de peso corporal al día.
En un estudio de alimentación con sonda a ratas durante 90 días, el
NOEL fue de 16 mg/kg por día, basado en la hepatotoxicidad. Los
estudios de alimentación de corta duración en perros no se
consideraron adecuados para establecer un NOEL. En un estudio
cutáneo de 10 días en conejos no se detectó toxicidad sistémica con
la única dosis ensayada (200 mg/kg).
1.6.3 Irritación y sensibilización cutánea y ocular
La aplicación a la piel de conejos y cobayos no produjo
irritación o sensibilización cutánea. Tras la aplicación a los ojos
de conejos se observó una irritación conjuntival moderada o ligera.
1.6.4 Exposición prolongada
En ratas macho y hembra a las que se administraron 2500 mg/kg
de alimentos se registró una disminución del número de eritrocitos y
de los valores de la hemoglobina y el hematocrito. No se observó
ninguna toxicidad de tipo hepático. Las ratas macho a las que se
administraron 2500 mg/kg de alimentos o más presentaron un aumento
marginal de la atrofia testicular y la prostatitis. El NOEL en las
ratas es de 500 mg/kg de alimentos. En perros que recibieron una
dieta con 500 mg de carbendazima por kg de alimentos durante un año
o más se observó aumento del colesterol sérico, elevación de la
actividad de la fosfatasa alcalina y otras indicaciones de
hepatotoxicidad. El NOEL en perros es de 300 mg/kg de alimentos.
1.6.5 Reproducción, embriotoxicidad y teratogenicidad
La carbendazima no tuvo ningún efecto adverso en la
reproducción cuando se administró a ratas en un estudio de
reproducción de tres generaciones a dosis de hasta 500 mg/kg de
alimentos. Se registró una reducción de la fecundidad masculina en
las ratas a las que se administró carbendazima (200 mg/kg por día)
con sonda durante 85 días. Con una dosis de 50 mg/kg de peso
corporal al día en este estudio disminuyó considerablemente el
número de espermatozoides en el epidídimo.
Tras la administración a ratas de una dosis oral única de 100
mg/kg de peso corporal, el examen histológico puso de manifiesto una
interrupción temprana (0-2 días) de la espermatogénesis, con
oclusión de los conductos eferentes y un aumento del peso de los
testículos. En este estudio de dosis única no se observó ningún
efecto con 50 mg/kg. Los efectos se mantuvieron hasta transcurridos
70 días en ratas tratadas con 400 mg/kg.
La carbendazima provocó un aumento de las malformaciones y las
anomalías en las ratas administrada a dosis diarias superiores a 10
mg/kg en los días 7-16 de la gestación. En conejas se produjo un
ligero descenso de la tasa de implantación en dosis de 20 y 125
mg/kg por día administradas en los días 7-19 de la gestación, y un
aumento de la incidencia de la reabsorción a 125 mg/kg por día. Se
observó toxicidad materna con dosis de 20 y de 125 mg/kg por día en
ratas y conejos, respectivamente.
Además del descenso de la tasa de gestaciones y del aumento de
las reabsorciones precoces en la rata, se produjo una reducción
significativa del peso del feto con dosis de 20 y 90 mg/kg por día,
y un incremento significativo de las malformaciones fetales con la
dosis de 90 mg/kg por día. Estas consistieron fundamentalmente en
hidrocefalia, microftalmia, anoftalmia, malformaciones escapulares y
del esqueleto axial (fusiones vertebrales, costales y esternebrales,
exencefalia, hemivértebras e hiperplasia costal). Sin embargo, en
los conejos no aparecieron malformaciones significativas.
1.6.6 Mutagenicidad y otros efectos finales afines
Los ensayos realizados en sistemas de mamíferos y no mamíferos
in vitro e in vivo y en células somáticas, así como en células
germinales, demuestran que la carbendazima no tiene interacción con
el ADN celular, no induce mutaciones puntuales y tampoco produce
mutaciones de las células germinales.
Sin embargo, la carbendazima provoca aberraciones cromosómicas
numéricas (aneuploidía o poliploidía) en sistemas experimentales in
vitro e in vivo.
1.6.7 Carcinogenicidad
La administración de benomilo y carbendazima con los alimentos
a ratones CD-1 y suizos SPF dio lugar a un aumento de la incidencia
de tumores hepatocelulares.
Un estudio de carcinogenicidad de la carbendazima con ratones
CD-1 puso de manifiesto un aumento estadísticamente signifi cativo,
relacionado con la dosis, de la neoplasia hepatocelular en las
hembras. También se observó un aumento estadísticamente
significativo en los machos tratados con una dosis de nivel medio
(1500 mg/kg de alimentos), pero no en los que recibieron dosis
altas, a causa de la elevada mortalidad. Un estudio sobre la
carcinogenicidad de la carbendazima en una raza de ratones
genéticamente relacionada con la anterior, los ratones SPF (raza
aleatoria suiza), con dosis de 0, 150, 300 y 1000 mg/kg de alimentos
(que se aumentó a 5000 mg/kg durante el estudio), mostró un aumento
de la incidencia de adenomas y carcinomas hepatocelulares
combinados. Un tercer estudio realizado en ratones NMRKf con dosis
de 0, 50, 150, 300 y 1000 mg de carbendazima por kg de alimentos
(que se aumentó a 5000 mg/kg durante el estudio) no puso de
manifiesto efectos carcinogénicos. El benomilo y la carbendazima
provocaron tumores hepáticos en dos razas de ratones (CD-1 y SPF),
ambas con un elevado índice de tumores hepáticos espontáneos. En
cambio, la carbendazima no es carcinogénica en ratones NMRKf, que
tienen un índice bajo de este tipo de tumores espontáneos.
Los estudios de carcinogenicidad del benomilo y la carbendazima
en ratas fueron negativos.
1.6.8 Mecanismo de toxicidad, modo de acción
Los efectos biológicos del benomilo y la carbendazima se deben
a su interacción con los microtúbulos celulares. Estas estructuras
intervienen en funciones esenciales, como la división celular, que
inhiben el benomilo y la carbendazima. La toxicidad de estos
productos en los mamíferos está vinculada a una disfunción
microtubular.
El benomilo y la carbendazima, al igual que otros compuestos
del bencimidazol, tienen una toxicidad selectiva para distintas
especies, que se explica, por lo menos en parte, porque el benomilo
y la carbendazima se unen de manera distinta a los microtúbulos de
las especies específicas en las que actúan y en las que no.
1.7 Efectos en el ser humano
No se ha informado de efectos adversos para la salud humana.
1.8 Efectos en otros organismos en el laboratorio y en el medio
ambiente
Con las dosis de aplicación recomendadas, la carbendazima tiene
pocos efectos sobre la actividad microbiana del suelo. Se han
notificado algunos efectos adversos sobre ciertos tipos de hongos.
La CE50 a las 72 horas para el alga verde Selenastrum
capricornutum, basada en el crecimiento total, se calculó en 1,3
mg/litro; la NOEC fue de 0,5 mg/litro. La toxicidad de la
carbendazima para los invertebrados acuáticos y los peces varía
ampliamente, con una CL50 a las 96 h que oscila entre 0,007
mg/litro para Ictalurus punctatus y 5,5 mg/litro para Lepomis
macrochirus. En una prueba de 21 días con Daphnia magna, el
comienzo de la reproducción se retrasó considerablemente con 0,025
mg/litro; la NOEC fue de 0,013 mg/litro.
La carbendazima es tóxica para las lombrices de tierra en
experimentos de laboratorio con una exposición a concentraciones
normales y con las dosis de aplicación recomendadas en el campo. Es
"relativamente no tóxica" para las abejas de miel y su toxicidad es
baja para las aves.
2. Conclusiones
El benomilo causa sensibilización cutánea en el ser humano.
Tanto el benomilo como la carbendazima representan un riesgo muy
pequeño de intoxicación aguda. Dados los niveles de exposición
actuales y el bajo índice de absorción cutánea de estos dos
compuestos, no es probable que puedan tener efectos de toxicidad
sistémica en la población general o en personas expuestas
profesionalmente. Estas son las conclusiones que se pueden sacar de
los datos obtenidos en animales y de los limitados datos sobre el
ser humano de que se dispone, pero estas extrapolaciones están
respaldadas por el conocimiento del modo de acción de la
carbendazima y el benomilo en especies en las que actúan y en las
que no.
Una mayor clarificación del mecanismo de toxicidad de ambos
compuestos en los mamíferos permitirá quizás definir mejor los
niveles sin efectos observados. El estudio de su unión a los
microtúbulos de las células destinatarias (tejidos testicular y
embrionario) facilitará la comparación entre distintas especies.
La carbendazima se adsorbe fuertemente en la materia orgánica
del suelo, que la conserva durante un período de hasta tres años.
Persiste en la superficie de las hojas y, por consiguiente, en las
hojas caídas. Se ha demostrado que las dosis recomendadas de
aplicación afectan negativamente a las lombrices de tierra (con
efectos sobre la población y la reproducción). No se dispone de
datos acerca de sus efectos sobre otros artrópodos del suelo o de la
maleza, que estarían igualmente expuestos.
No es probable que se pueda observar en el medio ambiente la
elevada toxicidad demostrada en las pruebas de laboratorio para los
organismos acuáticos, debido a la baja biodisponibilidad de los
residuos de carbendazima unidos a los sedimentos. Sin embargo, no se
dispone de información acerca de sus efectos en las especies que
viven en los sedimentos, que sufrirían la exposición más intensa.