
INTERNATIONAL PROGRAMME ON CHEMICAL SAFETY
ENVIRONMENTAL HEALTH CRITERIA 160
ULTRAVIOLET RADIATION
This report contains the collective views of an international group of
experts and does not necessarily represent the decisions or the stated
policy of the United Nations Environment Programme, the International
Labour Organisation, or the World Health Organization.
An Authoritative Scientific Review of Environmental and Health
Effects of UV, with Reference to Global Ozone Layer Depletion
Published under the joint sponsorship of
the United Nations Environment Programme,
the International Labour Organisation,
and the World Health Organization
World Health Orgnization
Geneva, 1994
The International Programme on Chemical Safety (IPCS) is a joint
venture of the United Nations Environment Programme, the International
Labour Organisation, and the World Health Organization. The main
objective of the IPCS is to carry out and disseminate evaluations of
the effects of chemicals on human health and the quality of the
environment. Supporting activities include the development of
epidemiological, experimental laboratory, and risk-assessment methods
that could produce internationally comparable results, and the
development of manpower in the field of toxicology. Other activities
carried out by the IPCS include the development of know-how for coping
with chemical accidents, coordination of laboratory testing and
epidemiological studies, and promotion of research on the mechanisms
of the biological action of chemicals.
WHO Library Cataloguing in Publication Data
Ultraviolet radiation
(Environmental health criteria: 160)
1. Ultraviolet rays 2. Radiation effects
3. Environmental exposure I.Series
ISBN 92 4 157160 8 (NLM Classification: WD 605)
ISSN 0250-863X
The World Health Organization welcomes requests for permission
to reproduce or translate its publications, in part or in full.
Applications and enquiries should be addressed to the Office of
Publications, World Health Organization, Geneva, Switzerland,
Which will be glad to provide the latest information on any changes
made to the text, plans for new editions, and reprints and
translations already available.
(c) World Health Organization 1994
Publications of the World Health Organization enjoy copyright
protection in accordance with the provisions of Protocol 2 of the
Universal Copyright Convention. All rights reserved.
The designations employed and the presentation of the material in
this publication do not imply the impression of any opinion whatsoever
on the part of the Secretariat of the World Health Organization
concerning the legal status of every country, territory, city, or area
or of its authorities, or concerning the delimitation of its frontiers
or boundaries.
The mention of specific companies or of certain manufacturers'
products does not imply that they are endorsed or recommended by the
World Health Organization in preference to others of a similar nature
that are not mentioned. Errors and omissions excepted, the names of
proprietary products are distinguished by initial capital letters.
CONTENTS
PREFACE
1. SUMMARY AND CONCLUSIONS
1.1. Physical characteristics
1.2. Action spectrum and minimum erythemal dose
1.3. Cellular and molecular studies
1.4. Animal studies
1.5. Health effects on humans
1.5.1. Skin
1.5.2. Immune system
1.5.3. Eye
1.6. Environment
1.7. Guidelines on exposure limits and protective measures
2. PHYSICAL CHARACTERISTICS
2.1. Electromagnetic spectrum
2.2. Radiometric quantities and units
2.3. UV production
2.3.1. Thermal emitters
2.3.2. Electrical gaseous discharges
2.3.3. Stimulated emission
2.4. Summary
3. UV SOURCES
3.1. The sun
3.1.1. Factors affecting solar UV levels
3.1.2. Ozone depletion effects
3.1.3. Trends in UV 19
3.1.4. Theoretical models
3.1.5. UV monitoring
3.1.6. Analysis of UV data
3.1.7. Conclusions
3.2. Artificial sources
3.2.1. Incandescent sources
3.2.2. Gaseous discharge sources
3.2.3. Gas welding
3.2.4. Arc welding
3.2.5. Lasers
3.2.6. Sunbeds
4. HUMAN EXPOSURE
4.1. Sunlight
4.2. Skin exposure geometry
4.3. Ocular exposure geometry
4.4. Workplace
4.4.1. Outdoor work
4.4.2. Indoor work
4.4.3. Research
4.4.4. Commerce
4.4.5. Medicine and dentistry
4.5. Elective exposure
5. DOSIMETRIC CONCEPTS
5.1. Interaction of UV with matter
5.2. Biological weighting factors and spectrally
weighted quantities
5.3. Measurement techniques
5.3.1. Detectors
5.3.2. Radiometers
5.3.3. Spectroradiometers
5.3.4. Personal dosimetry
5.4. Calibration
6. CELLULAR AND MOLECULAR STUDIES
6.1. Introduction
6.2. Interactions with biomolecules
6.2.1. Cellular chromophores
6.2.2. Cellular targets
6.3. Action spectra
6.4. Biomolecular damage
6.4.1. Nucleic acids
6.4.2. Membranes
6.4.3. Proteins
6.5. Cellular defenses
6.5.1. DNA
6.5.2. Human excision repair disorders
6.5.3. Antioxidant pathways
6.5.4. Summary
6.6. Cellular consequences of damage
6.6.1. Membrane disruption
6.6.2. Activation of genes
6.6.3. Cell death
6.6.4. Mutation, chromosomal damage and transformation
6.7. Conclusions
7. ANIMAL STUDIES
7.1. Skin carcinogenesis
7.1.1. Domestic animals
7.1.2. Experimental animals
7.1.3. Interactions between radiations
of different wavelengths
7.1.4. Dose response
7.1.5. Effect of pattern of exposure
7.1.6. Action spectrum
7.1.7. Interaction between UV and chemicals
7.1.8. Mechanisms of UV carcinogenesis
7.1.9. Conclusions
7.2. Immune responses
7.2.1. Immune function assays
7.2.2. Susceptibility to rumours
7.2.3. Susceptibility to infectious disease
7.2.4. Susceptibility to immunologically-mediated
disease
7.2.5. Conclusions
7.3. Ocular studies
7.3.1. Introduction
7.3.2. General effects
7.3.3. Caractogenesis
7.3.4. Retinal effects
8. HUMAN STUDIES: THE SKIN
8.1. Characteristics
8.1.1. Structure and optical properties
8.1.2. Skin types
8.2. Beneficial effects
8.2.1. Vitamin D3
8.2.2. Skin adaptation
8.2.3. Other benefits
8.3. Acute effects
8.3.1. Erythema and sunburn
8.3.2. Skin pigmentation and tanning
8.3.3. Photosensitization
8.4. Chronic effects on the skin other than cancer
8.5. Cancer
8.5.1. Nonmelanocytic skin cancer
8.5.2. Cutaneous melanoma
8.5.3. Cancer of the lip
8.5.4. Ocular cancers
8.5.5. Other cancers
8.5.6. Action spectrum
8.5.7. Dose response
8.5.8. Effects of pattern of exposure
8.5.9. Interactions between UV and other agents
8.5.10. Mechanisms of UV carcinogenesis
8.6. Conclusions
9. HUMAN STUDIES: IMMUNE FUNCTION
9.1. Immune function assays
9.2. Susceptibility to tumours, infectious and autoimmune
diseases
9.3. Conclusions
10. HUMAN STUDIES: THE EYE
10.1. Introduction
10.2. The eye
10.3. Study design
10.4. Diseases of the external eye
10.4.1. Photokeratitis and photoconjunctivitis
10.4.2. Climatic droplet keratopathy
10.4.3. Pinguecula
10.4.4. Pterygium
10.4.5. Hyperkeratosis, carcinoma-in-situ, and squamous
cell carcinoma of the conjunctiva
10.5. Diseases of the lens
10.5.1. Cataract
10.5.2. Exfoliation syndrome
10.5.3. Anterior lens capsule
10.6. Diseases of the choroid and retina
10.6.1. Uveal melanoma
10.6.2. Age-related macular degeneration
10.7. Conclusions
11. EFFECTS ON PLANT AND AQUATIC ECOSYSTEMS
11.1. Introduction
11.2. Effects on terrestrial plants
11.2.1. UV penetration into the leaf
11.2.2. Changes in growth
11.2.3. Effects on plant function
11.2.4. Species competition
11.2.5. Plant diseases
11.2.6. UV-protection systems
11.3. Effects on aquatic ecosystems
11.3.1. Effects on phytoplankton
11.3.2. UV increase and primary biomass production
11.4. Conclusions
12. HEALTH HAZARD ASSESSMENT
12.1. Introduction
12.2. Elective exposures
12.2.1. Medical exposures
12.2.2. Phototherapy of seasonal effective disorder
(SED)
12.2.3. Sunbeds
12.2.4. Sunbathing
12.3. Adventitious exposures
12.3.1. Outdoor exposures
12.3.2. Artificial sources
13. INTERNATIONAL GUIDELINES ON EXPOSURE STANDARDS
14. PROTECTIVE MEASURES
14.1. Introduction
14.2. Education
14.3. Protection factors
14.4. Clothing
14.5. Sunscreens
14.6. Tanning devices
14.7. Occupational protection
14.8. Protection in medicine and dentistry
14.9. Nutrition
14.10. Additional protective agents
14.11. Eye protection
15. FUTURE RESEARCH
15.1. Introduction
15.2. INTERSUN
15.3. Solar and personal UV monitoring
15.3.1. Solar monitoring
15.3.2. Personal monitoring
15.4. Terrestrial plants
15.5. Aquatic ecosystems
15.6. Human health
15.6.1. Skin
15.6.2. Immune system
15.6.3. Eye
15.7. Laboratory studies
15.8. Education
15.9. Administration
16. PREVIOUS EVALUATIONS BY INTERNATIONAL BODIES
16.1. United Nations Environment Programme
16.1.1. Ozone
16.1.2. Human health
16.2. International Agency for Research on Cancer
16.3. World Health Organization
16.4. International Commission on Non-Ionising
Radiation Protection
REFERENCES
PREFACE
The World Health Organization (WHO), in collaboration with the
United Nations Environment Programme (UNEP) and the International
Non-Ionizing Radiation Committee (INIRC) of the International
Radiation Protection Association (IRPA), published the first
Environmental Health Criteria (EHC) monograph on Ultraviolet
Radiation in 1979. At the United Nations Conference on the
Environment and Development (UNCED) in 1992 it was declared under
Agenda 21 that there should be activities on the effects of
ultraviolet radiation. Specifically:
"(i) Undertake, as a matter of urgency, research on the effects on
human health of the increasing ultraviolet radiation reaching the
earth's surface as a consequence of depletion of the stratospheric
ozone layer;
(ii) On the basis of the outcome of this research, consider taking
appropriate remedial measures to mitigate the above-mentioned
effects on human beings".
Within the United Nations mandate, and that of the 1993 WHO
Global Strategy for Health and Environment, this monograph has been
drafted to provide the essential authoritative review on which
future research programmes in UV can progress.
IRPA initiated activities in NIR by forming a Working Group on
Non-Ionizing Radiation in 1974. This Working Group later became the
INIRC at the IRPA meeting held in Paris in 1977. In May 1992 the
INIRC was chartered as an independent scientific commission called
the International Commission on Non-Ionizing Radiation Protection
(ICNIRP). ICNIRP continues the work of the IRPA/INIRC by reviewing
the scientific literature on NIR and making assessments of health
risks of human exposure to such radiation. Using the Environmental
Health Criteria monographs, developed in conjunction with WHO,
ICNIRP recommends guidelines on exposure limits, drafts codes of
practice, and works in conjunction with other international
organizations to promote safety and standardization in the
non-ionizing radiation fields.
A UNEP/WHO/ICNIRP Task Group to review the final draft of the
updated Environmental Health Criteria on Ultraviolet (UV) Radiation
met in Geneva from 8-11 December, 1993. Dr W. Kreisel, Executive
Director, World Health Organization opened the meeting on behalf of
WHO. Dr H. Gopalan and Mr R. Matthes welcomed the participants on
behalf of UNEP and ICNIRP respectively.
The first revision of this publication was compiled by members
of the IRPA/INIRC and more recently by members of ICNIRP. Chapters
were prepared by Drs B. Armstrong, J-P. Cesarini, L. Court, P.
Dolan, G. Johnson, A. Kricker, A. McKinlay, M. Repacholi, M.
Selgrade, D. Sliney and R. Tyrrell. These chapters together with the
report of an informal consultation held in Geneva, in August 1993 on
"The Effects of Solar UV Radiation on the Eye", the International
Agency for Research on Cancer (IARC) report of the meeting on
"Health, Solar UV Radiation and Environmental Change " held in Lyon
in October 1992, and the meeting on Immunotoxicology held in
Bilthoven in July 1993, were used by Drs M. Repacholi and D. Sliney
to compile the review draft in September 1993. Comments were
received from a broad cross-section of specialists in the UV fields
and their reviews were gratefully received. These comments were then
incorporated for Task Group review. Final scientific editing of the
text was completed by Drs J-P. Cesarini, A. McKinlay, M. Repacholi
and D. Sliney. Sincere thanks to Christine Cornish and Nancy Smith
for their assistance in the preparation of this text. An editorial
group consisting of Drs M. Repacholi, H. Gopalan and T. Kjellström
coordinated the preparation of this monograph.
This monograph comprises a review of the data on the effects of
exposure on biological systems pertinent to the evaluation of human
health risks. Its purpose is to give an overview of the known
biological effects of UV, identify gaps in knowledge and provide
direction for further research. This monograph will assist health
authorities, regulatory and similar agencies to provide guidance on
health risks from exposure to UV and limits for occupational and
general public exposure.
Earlier reports were not necessarily included, as they were
reviewed in the 1979 monograph. Every effort has been made to
distinguish clearly between established biological effects and those
that have been reported as preliminary or isolated results, or as
hypotheses proposed to explain observed results. The conclusions are
based on established knowledge of interactions of UV with biological
systems.
Subjects reviewed include: the physical characteristics of UV;
measurement techniques; applications and sources of exposure;
mechanisms of interaction; biological effects; guidance on
protective measures; and recommendations on exposure limits.
This monograph will also serve as a scientific database for the
planned WHO/UNEP/IARC/ICNIRP International Research Programme on
Health, Solar UV Radiation and Environmental Change (INTERSUN). The
general objectives of INTERSUN are to:
(i) evaluate the quantitative relationship between solar UV at the
surface of the earth and human health effects, develop reliable
predictions of the health consequences of changes in UV, provide
baseline estimates of the incidence of health effects of UV in
representative populations around the world, and develop practical
ways of monitoring change in these effects over time in relation to
environmental and behavioural change;
(ii) provide essential input into the development of environmental
and public health policies and actions in relation to depletion of
stratospheric ozone; and
(iii) provide a framework for monitoring the effects of solar UV and
the impact of prevention programmes.
Health agencies and regulatory authorities are encouraged to
set up and develop programmes that ensure effective protection
against the health effects of UV. It is hoped that this criteria
monograph will provide useful information for such endeavours.
TASK GROUP MEETING ON THE REVISION OF ENVIRONMENTAL HEALTH CRITERIA
ON UV RADIATION
Members
Dr J.P. Cesarini LRTPH, Fondation Rothschild, Paris, France
Dr L. Court Centre de Recherches du Service de Santé
des Armées, La Tronche Cédex, France
Dr A. McKinlay National Radiological Protection Board,
Didcot, United Kingdom
Dr D. Sliney US Army Environmental Hygiene Agency,
Aberdeen Proving Ground, MD, USA
Dr M.J. Selgrade Immunotoxicology Branch, USEPA, Research
Triangle Park, NC, USA
Dr G. Johnson Institute of Ophthalmology, University of
London, London, United Kingdom
Dr B. Armstrong International Agency for Research on
Cancer, Lyon, France
Dr H. Van Loveren National Institute of Public Health &
Environmental Protection, Bilthoven,
Holland
Dr A. Koppikar Office of Health & Environmental
Assessment, USEPA, Washington DC, USA
Dr R.M. Tyrrell Swiss Institute for Cancer Research,
Epalinges/Lausanne, Switzerland
Dr J.C. van der Leun Academisch Ziekenhuis Utrecht, CX Utrecht,
Holland
Dr I.A. Badr King Khaled Eye Specialist Hospital, King
Saud University, Riyadh, Saudi Arabia
Dr G. Kulandaivelu School of Biological Sciences, Madurai
Kamaraj University, Madurai, India
Dr E. Turowski Institute for Water, Soil and Air Hygiene,
Berlin, Germany
Observers
Dr B. Weatherhead Surface Radiation Research Branch, Boulder,
Colorado, USA
Dr H. Gopalan United Nations Environment Programme
(UNEP), Nairobi, Kenya
Dr A. Kulmula World Meteorological Organization (WMO),
Geneva, Switzerland
Dr A. Kricker International Agency for Research on
Cancer, Lyon, France
Dr U. Feister Deutscher Wetterdienst, Potsdam, Germany
Dr W.H. Weihe President, International Society of
Biometeorology (ISB), Brannenburg, Germany
Dr R. Matthes Bundesamt fur Strahlenschutz, Ingolsädter,
Oberschleissheim, Germany
Dr C. Remé Professor of Ophthalmology,
Universitätsspit, Zurich, Switzerland
WHO Secretariat
Dr W. Kreisel Executive Director, Health and Environment
Mr G. Ozolins Manager, Prevention of
Environmental Pollution
Dr T. Kjellström Prevention of Environmental Pollution
Dr M. Repacholi Prevention of Environmental Pollution
(Secretary)
Dr B. Thylefors Manager, Programme for the Prevention of
Blindness
Dr P.-H. Lambert Chief, Microbiology and Immunology Support
Services
Dr A.-D. Négrel Programme for the Prevention of Blindness
Dr V. Koroltchouk Cancer and Palliative
Care
NOTE TO READERS OF THE CRITERIA MONOGRAPH
Every effort has been made to present information in the
criteria monographs as accurately as possible without unduly
delaying their publication. In the interest of all users of the
Environmental Health Criteria monographs, readers are kindly
requested to communicate any errors that may have occurred to the
Director of the International Programme on Chemical Safety, World
Health Organization, Geneva, Switzerland, in order that they may be
included in corrigenda, which will appear in subsequent volumes.
1. SUMMARY AND CONCLUSIONS
This monograph is concerned with the effects of ultraviolet
(UV) radiation exposure on human health and the environment. Such a
review of the scientific literature is considered timely in view of
the consequences of increased levels of UV at the surface of the
earth resulting from depletion of stratospheric ozone.
1.1 Physical Characteristics
Exposure to UV occurs from both natural and artificial sources.
The sun is the principal source of exposure for most people. Solar
UV undergoes significant absorption by the atmosphere. With
depletion of the stratospheric ozone people and the environment will
be exposed to higher intensities of UV. The consequences of this
added UV exposure are considered so serious that it was a major
topic for discussion at the World Environment Conference, held in
Rio de Janeiro in 1992. In Agenda 21, adopted by the Conference, it
was specifically recommended to "undertake, as a matter of urgency,
research on the effects on human health of the increasing
ultraviolet radiation reaching the earth's surface as the
consequence of depletion of the stratospheric ozone layer." It is
this issue that underscores the current need to better understand
the potential health and environmental risks of UV exposure.
UV is one of the non-ionizing radiations in the electromagnetic
spectrum and lies within the range of wavelengths 100 nm (which
corresponds to a photon energy of approximately 12 eV) to 400 nm.
The short wavelength limit of the UV region is often taken as the
boundary between the ionizing radiation spectrum (wavelengths < 100
nm) and the non-ionizing radiation spectrum. UV can be classified
into UVA (315 - 400 nm), UVB (280 - 315 nm) and UVC (100 - 280 nm)
regions, although other conventions for UVA, UVB and UVC wavelengths
bands are in use.
Most artificial sources of UV, except for lasers, emit a
spectral continuum of UV containing characteristic peaks, troughs
and lines. These sources include various lamps used in medicine,
industry, commerce, research and the home.
UV-induced biological effects depend on the wavelengths of the
radiation. It is necessary for a proper determination of hazard to
have spectral emission data. These consist of spectral irradiance (W
m-2 nm-1) measurements or calculations of emissions from the
source. The total irradiance (W m-2) is obtained by summing over
all wavelengths emitted. The biological or hazard weighted
irradiance (W m-2 effective) is determined by multiplying the
spectral irradiance at each wavelength by the biological or hazard
weighting factor (which quantifies the relative efficacy at each
wavelength for causing the effect) and summing over all wavelengths.
Such factors are obtained from action spectra.
1.2 Action Spectrum and Minimum Erythemal Dose
An action spectrum is a graph of the reciprocal of the radiant
exposure required to produce the given effect at each wavelength.
All the data in such curves are normalized to the datum at the most
efficacious wavelength(s). By summing the biologically effective
irradiance over the exposure period, the biologically effective
radiant exposure (J m-2 effective) can be calculated. For UV
induced erythema, the action spectrum adopted by the International
Commission on Non-Ionizing Radiation Protection (ICNIRP),
International Commission on Illumination (CIE), the International
Electrotechnical Commission (IEC) and various national bodies, is a
composite curve obtained by statistical analysis of many research
results on the minimum radiant exposure of UV at different
wavelengths necessary to just cause erythema.
The most commonly used quantity for describing the erythemal
potential of an exposure to UV is the number of minimum erythemal
doses (MEDs) represented by the exposure. An MED is the radiant
exposure of UV that produces a just noticeable erythema on a
previously unexposed skin. It corresponds to a radiant exposure of
monochromatic radiation at the maximum spectral efficacy for
erythema (around 300 nm) of approximately 150 to 2000 J m-2
effective, depending on skin type. In this text 200 - 300 J m-2
effective is used as the value of 1 MED for comparative safety
purposes for white skin.
1.3 Cellular and Molecular Studies
To produce any change, UV must be absorbed by the biomolecule.
This involves absorption of a single photon by the molecule and the
production of an excited state in which one electron of the
absorbing molecule is raised to a higher energy level. The primary
products caused by UV exposure are generally reactive species or
free radicals which form extremely quickly but which can produce
effects that can last for hours, days or even years. DNA is the most
critical target for damage by UVB and UVC radiations. While a
considerable amount of knowledge is available concerning the
interaction of UV with nucleic acids, controversy exists as to which
lesion constitutes the most important type of pre-mutagenic damage.
Cell death, chromosome changes, mutation and morphological
transformations are observed after UV exposure of procaryotic and
eucaryotic cells. Many different genes and several viruses
(including HIV) are activated by UV exposure. The genes activated by
UVB and UVC are different from those activated by UVA. Studies of
DNA repair defective disorders have clearly established a link
between UV induced DNA damage in skin and various types of cancer.
1.4 Animal Studies
Solar UV has been shown to produce cancers in domestic and food
animals. In experimental animals UV causes predominantly squamous
cell carcinomas. UVB is most effective at producing SCCs, although
they are produced by UVA but at much higher intensities, similar to
the levels needed for erythema and tanning. The effectiveness of UVC
is unknown except at one wavelength (254 nm). At this wavelength the
effectiveness is less than UVB.
Melanomas are much less common and only two animal models have
been found for induction of melanoma by UV alone. An initial action
spectrum determined for a type of hybrid fish indicates a peak in
the UVB range but also shows a high level of effectiveness in the
UVA. Basal cell carcinomas are rare in animals.
Exposure to suberythemal doses of UV have been shown to
exacerbate a variety of infections in rodent models. UV affects
infections both at the site of exposure and at distant sites. Recent
work indicates that systemic infections without skin involvement may
be affected. Enhanced susceptibility appears to result from T-helper
cell activity. The mechanisms associated with this suppression
appear to be the same as those identified with suppression to
contact and delayed type hypersensitivity responses. Suppression of
these immune responses appears to be mediated by release of soluble
mediators from UVB exposed skin which alters the antigen
presentation by Langerhans and other cells so that they fail to
activate TH 1 cells. The resulting immune suppression is antigen
specific, can occur regardless of whether or not antigen is applied
at the site of exposure, and is relatively long lasting. UV exposure
also prevents the development of protection immunity to a variety of
infections in mice and rats.
Many studies in experimental animals have demonstrated that UV
exposure can cause both acute and delayed effects such as cataract,
photokeratitis, damage to the corneal epithelium and various retinal
effects. Studies of photochemical retinal injury in aphakic monkeys
have shown that the retina is six times more vulnerable to
photochemical damage from UV than the visible wavelengths.
1.5 Health Effects on Humans
1.5.1 Skin
Acute effects on the skin consist of solar erythema, "sunburn",
which, if severe enough, may result in blistering and destruction of
the surface of the skin with secondary infection and systemic
effects, similar to those resulting from a first or second degree
heat burn. Although UVC is very efficiently absorbed by nucleic
acids, the overlying dead layers of skin absorb the radiation to
such a degree that there is only mild erythema and, usually, no late
sequelae, even after repeated exposures. Much less is known about
the biological effects of UVA. However, doses of UVA, which alone
may not show any biological effect, can, in the presence of certain
environmental, consumer and medicinal chemical agents, result in
injury to tissues (phototoxicity, photoallergy, enhancement of
photocarcinogenesis).
Chronic skin changes due to UV consist of skin cancer (both
melanoma and non-melanocytic), benign abnormalities of melanocytes
(freckles, melanocytic naevi and solar or senile lentigines), and a
range of other chronic injuries resulting from UV exposure to
keratinocytes, blood vessels and fibrous tissue, often described as
"photoaging" (solar elastosis). The much increased rates of skin
cancer in patients with xeroderma pigmentosum, who have a deficiency
in the capacity to repair UV-induced DNA damage, suggest that direct
UV damage of the DNA may be a step in the cause of these cancers.
This suggestion has also been supported by the observation of UV
specific mutations of the p53 tumour suppressor gene in a proportion
of patients with non-melanocytic skin cancer. Oxidative and immune
suppressant effects may also contribute to the capacity of UV to
cause skin cancers.
Cancer of the lip is much more common in fair than dark skin
populations and is associated with outdoor work. However possible
confounding with tobacco and alcohol use has not been adequately
controlled in any study.
The worldwide incidence of malignant melanoma has continued to
increase. Strong epidemiological evidence exists that sun exposure
causes cutaneous melanoma and non-melanocytic skin cancer. Their
incidence is less in darker than light skin groups living in the
same geographical area. Risk of skin cancer decreases with
increasing pigmentation. The anatomical site most seen for squamous
cell carcinoma (SCC) is the head and neck, areas most exposed to the
sun. Incidence of both melanoma and non-melanocytic skin cancer are
increased in areas of high ambient solar UV radiation. Melanoma is
strongly related to frequency of recreational exposure to the sun
and to history of sunburns.
There is suggestive evidence that exposure to sunlamps may
increase the risk of melanoma, but the studies conducted so far have
not consistently controlled confounding factors.
1.5.2 Immune system
A number of studies suggest that UV exposures at environmental
levels suppress immune responses in both rodents and man. In rodents
this immune suppression results in enhanced susceptibility to
certain infectious diseases with skin involvement and some systemic
infections. Mechanisms associated with UV-induced immunosuppression
and host defence mechanisms which provide for protection against
infectious agents, are similar in rodents and man. It is therefore
reasonable to assume that exposure to UV may enhance the risk of
infection and decrease the effectiveness of vaccines in humans.
However additional research is necessary to substantiate this.
1.5.3 Eye
The acute effects of UV on the eyes consist of the development
of photokeratitis and photoconjunctivitis, which are unpleasant but
usually reversible and easily prevented by appropriate eyewear.
Chronic effects on the eye consist of the development of pterygium
and squamous cell cancer of the conjunctiva and cataracts. A review
of the studies suggests that there is sufficient evidence to link
acute ocular exposure to photokeratitis but our knowledge of the
effects of chronic exposure is less certain. While there is
sufficient evidence that cortical and posterior subcapsular
cataracts (PSC) can be caused by UVB in laboratory animals, there is
limited evidence to link cortical and PSC cataracts in humans to
chronic ocular exposure to UVB. Insufficient information is
available to separate out the other factors contributing to cataract
formation, or to state the proportion of cataracts which can be
attributed to UVB exposure. There is also limited evidence to link
the development of climatic droplet keratopathy and pterygium, but
insufficient evidence to link uveal melanoma with UV exposure.
1.6 Environment
Increased levels of UV due to ozone layer depletion may have
serious consequences for living organisms. A 10% reduction in ozone
could lead to as much as a 15-20% increase in UV exposure depending
on the biological process being considered. While the impact on
human health, crop production, fisheries etc. is largely unknown,
adverse effects of increased exposure to UVB have been reported on
plant growth, photosynthesis and disease resistance. Further, the
impact of increased UV levels on aquatic ecosystems (the major
contributor to the earth's biomass) may be substantial.
Phytoplankton, at the base of the aquatic food chain, serves as food
for larvae of fish and shrimp. These in turn are consumed by fish,
which subsequently provide an essential food source for many human
beings and other animals. A significant reduction in phytoplankton
from increased UVB exposure will directly affect the human and
animal marine food source.
1.7 Guidelines on Exposure Limits and Protective Measures
Guidance on exposure limits for UV are described in chapter 13.
International guidelines define exposure limits (ELs) below which it
is expected that nearly all people may be repeatedly exposed without
adverse effects. The ELs are intended to be used to evaluate
potentially hazardous exposures from, for example, solar radiation,
arcs, gas and vapour discharges, fluorescent lamps and incandescent
sources. The ELs are generally below levels which are often used for
the UV exposure of patients required as part of medical treatment
and below levels associated with sunbed exposure. ELs are not
intended to apply to exposure of pathologically photosensitive
individuals, to people concomitantly exposed to photosensitising
agents or to neonates.
Finally this monograph describes protection and control
measures such as the containment of UV sources, and methods for
personal protection including the use of sunscreen preparations,
clothing, eye and skin protection, and behavioural modifications.
While topical application of sunscreen is a preferred method of
absorbing UVB, some preparations do not absorb the longer wavelength
UVA effectively. Moreover, some have been found to contain
ingredients that are mutagenic in sunlight. There is still much
research necessary before the impact on health of increased levels
of UVA will be known. In the meantime people using sunscreens should
use only those with the highest sun protection factor (SPF) and be
aware that they are for their protection from the sun and not for
tanning purposes. Use of wide brimmed hats, protective clothing and
UV absorbing eye glasses is still the best personal protection
against the adverse effects of UV exposure.
With increasing levels of solar UV resulting from depletion of
the ozone layer, and the continuing rise in the level of melanoma
worldwide, people should become more aware of their UV exposure and
take appropriate precautions. These precautions include staying out
of the sun during the period around noon (the period when the UV
levels are highest), or wearing UV protective clothing, hats and sun
glasses. Broad spectrum (UVB and UVA protective) sunscreens should
be used when other means of protection are not feasible. These
sunscreens should be used to reduce exposure rather than lengthen
the period of exposure to the sun. Protection of young children is
particularly important for the prevention of long-term consequences
of UV exposure. In general behavioural patterns must change to
protect against increasing solar UV levels.
2. PHYSICAL CHARACTERISTICS
2.1 Electromagnetic Spectrum
Oscillations of electromagnetic fields can cause energy to be
transported in the form of electromagnetic radiation. Examples of
this type of radiative transport of energy include radiofrequency
waves, light and x rays. Ultraviolet (UV) radiation is one form of
electromagnetic energy in the optical region of the electromagnetic
spectrum. All electromagnetic radiation is characterized by
frequency f and wavelength lambda. These two quantities are linked
through the relationship:
f = c/lamda
where c is the speed of light (3 108 m s-1). The energy of a
single photon is determined by the wavelength of the photon as
described by the relationship
(photon energy) = hf = hc/lamda where h = 1.24 eV nm
Non-ionizing radiation (NIR) is the term generally applied to
all forms of electromagnetic radiation whose primary mode of
interaction with matter is other than by producing ionization. NIR
refers to electromagnetic fields and radiation with wavelengths
exceeding 100 nm, which is equivalent to quantum (photon) energies
below 12.4 eV, the minimum energy needed to break the weakest
macromolecular bonds. The non-ionizing spectrum encompasses all
fields of radiation from UV to DC fields.
For purposes of health protection, the optical portion of NIR
can be subdivided into several wavelength ranges, as shown in table
2.1. The nomenclature was standardized by the International
Commission on Illumination (CIE) however, some scientists use a
modification of this system by shifting the 315 nm break point
between UVA and UVB to 320 nm; the reader should always check the
definition given in each publication. For the purposes of this
document, the CIE convention is followed.
Table 2.1 Optical Radiation Spectral Bands
Ultraviolet radiation 100 - 400 nm
UVA 315 - 400 nm
UVB 280 - 315 nm
UVC 100 - 280 nm
Visible radiation (light) 400 - 760 nm
Infrared radiation (IR) 760 - 106 nm = 1 mm
UV of wavelengths less than 180 nm has no direct biological
effect on humans since it is effectively absorbed in a few
centimetres of air. For this reason, the spectral region below 180
nm is frequently referred to as the vacuum ultraviolet region.
2.2 Radiometric Quantities and Units
Radiometric quantities are absolute physical quantities used to
describe the characteristics of a source or radiation field. For UV,
ten generic radiometric terms are summarized in table 2.2. Each of
these quantities can be defined for a certain wavelength or
frequency range, or can be integrated over the whole spectrum of a
given source. Since UV is normally absorbed over a surface, with
very limited penetration depth, the most common quantities used to
describe exposure dose and dose rate to UV are: radiant exposure
(incident energy divided by the receptor surface area) and
irradiance (incident power divided by the receptor surface area)
respectively. Radiant exitance is the power per area of the emitted
radiation at the emitting surface. The receptor surface is most
often considered as a flat plane. However, for some biological and
chemical purposes, the radiation incident on a cylindrical or
spherical surface may also be considered.
For the purpose of radiation protection, physical quantities
are needed to describe sources and fields of radiation as well as
the interaction of this optical radiation with matter. These are
described in IRPA/INIRC(1985).
2.3 UV Production
Sources of electromagnetic radiation can be categorized in
several different ways; for example they can be grouped according to
the type of material or the type of equipment that produces the
radiation. At the submicroscopic level, the manner in which the
radiation originates can be described in terms of nuclear,
electronic or molecular transitions between energy states or by the
acceleration of charged particles. Common sources of UV emission
involve energy transitions between electronic states of molecules in
materials.
2.3.1 Thermal emitters
When the temperature of a material increases, electrons in the
molecules are raised to higher energy states, a variety of energy
transitions take place and photons are emitted. The higher the
temperature the greater is the fraction of these photons at higher
energies. Matter at temperatures above 2500 K emits a significant
number of photons in the UV spectral range. The emission spectra of
such incandescent sources are characteristically smooth - so-called
"continuum", possibly with superimposed spectral emission lines.
Table 2.2 Radiometric terminology for UV:
Useful radiometric quantities and units
Quantity Symbol Defining equation Unit and abbreviation
Radiant energy Qe Qe = Integral Operator Phie dt joule (J) = 1 watt second
dQe
Radiant energy We We = joule per cubic metre
density dV (J m-3)
dQe
Radiant flux Phie , P Phie = watt (W)
(radiant power) dt
dPhie
Radiant exitance Me Me = watt per square metre
dA (W m-2)
= Integral Operator Le cosTheta dOmega
Irradiance or dPhie
Radiant Flux Ee Ee = watt per square metre
Density dA (W m-2)
dPhie
Radiant intensity Ie Ie = watt per steradian (W sr-1)
dOmega
d 2Phie
Radiance1 Le Le = watt per steradian per
dOmega dA cosTheta square metre (W sr-1 m-2)
Table 2.2 (contd).
Quantity Symbol Defining equation Unit and abbreviation
Radiant exposure He = dQe joule per square metre
(dose in He (J m-2)
photobiology) dA
P
Radiant efficiency2 ne ne = unitless
(of a source) Pi
Optical density3 De De = - log10(gammae) unitless
NOTE: All terms in this table are radiometric terms and should not be confused with photometric terms.
The symbol A represents surface area, Omega is solid angle, Theta is the incident (zenith) angle,
V represents volume, t is time. The units may also refer to narrow spectral bands in which the term is
preceded by the word spectral and the unit is then per wavelength interval and the symbol has a
subscript lamda. For example, spectral irradiance Egamma has units of W m-2 nm-1 or W cm-2 nm-1. While
the metre is the preferred unit of length, the centimetre is still commonly used for many of the above
terms and the nm or µm are most commonly used to express wavelength.
1. At the source L = dI and at a receptor L = dE
dA cosTheta dOmega cosTheta
2. Pi is electrical input power in watts
3. This formula applies only to situations where the radiation is not scattered, but only absorbed.
In this case gamma represents the fraction of transmitted energy.
The spectral emission of a heated material is governed by basic
physical laws. The Stefan-Boltzmann Law expresses the total
radiation emitted by a heated material as a function of its
temperature. The total radiant power emitted by a theoretically
perfect radiator, a so-called black body, is proportional to the
fourth power of its temperature (in kelvin, K). Any practical
thermal emitter in equilibrium emits less radiant power than its
equivalent black body at the same temperature and unlike a black
body the amount depends on the nature of its surface. The spectral
distribution of black body radiation is described by the Planck
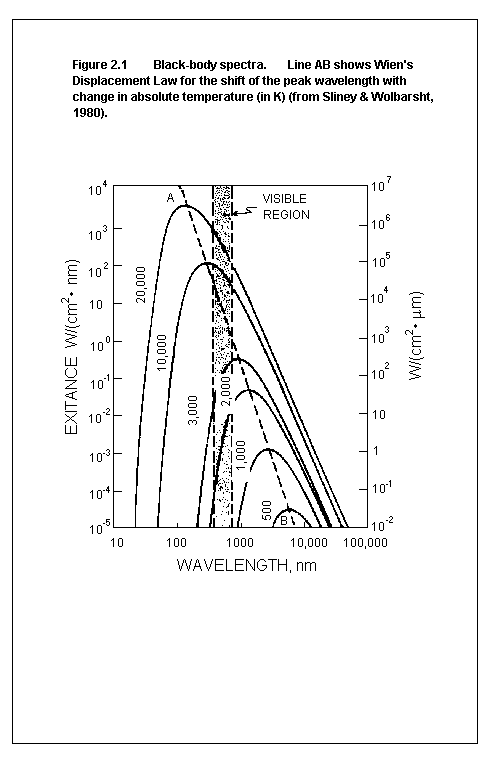
Radiation Law. Wien's Displacement Law is illustrated in figure 2.1,
and describes mathematically the spectral distribution of emission
from a black body and the wavelength of the maximum emission shift
to shorter wavelengths as temperature increases. When a material is
heated so that incandescence occurs, it first appears red and, with
increasing temperature, progresses to white or blue.
2.3.2 Electrical gaseous discharges
Radiation may be produced when an electric current is passed
through a gas or vapour. Atoms may be ionized if sufficient energy
is transferred from a moving electron. Alternatively the moving
electron may not impart sufficient energy for ionization, but
instead may impart energy to raise the electrons of the gas to an
excited (higher) energy level. When they return to a lower energy
level, or their ground state, radiation of one or more
characteristic wavelengths is emitted. The wavelengths of emission
are determined by the type of gas or vapour present in the discharge
and appear as spectral emission lines. The width of the lines and
the amount of radiation in the interval between them depends on the
pressure of the discharge. At low pressure, fine lines with little
or no continuum are produced. As the pressure of the discharge is
raised the lines broaden and their relative magnitudes alter. The
magnitude of the continuum increases. The electrical gaseous
discharge is the basis of operation of many UV emitting lamps.
2.3.3 Stimulated emission
Radiation can be produced by the specific electronic transition
process of stimulated emission. This depends on the ability of the
radiating medium to undergo "population inversion", i.e., achieving
a condition where there are more atoms or molecules in a higher
energy excited state than in a lower one. Once population inversion
occurs an avalanche of photons can be generated by stimulated
emission. Initially, spontaneously emitted photons stimulate other
excited atoms to emit photons of the same energy in phase with one
another. This is the basis of operation of the laser.
 2.4 Summary
UV is a part of the non-ionizing region of the electromagnetic
spectrum. Its position in the spectrum can be characterised by
wavelength and it is quantified using radiometric quantities and
units. Production of UV is possible by different mechanisms that are
all based on atomic or molecular excitation. Excitation in a medium
can be achieved by thermal, electrical or optical energy absorption.
Emission of radiation is caused either by de-excitation or by
stimulated synchronised processes.
3. UV SOURCES
3.1 The Sun
The sun is the main source of UV. The broad spectrum and
intensity of UV from the sun are due to the high temperature at its
surface and its size. The intensity of solar UV reaching the earth's
atmosphere would probably be lethal to most living organisms on the
earth's surface without the shielding afforded by the atmosphere.
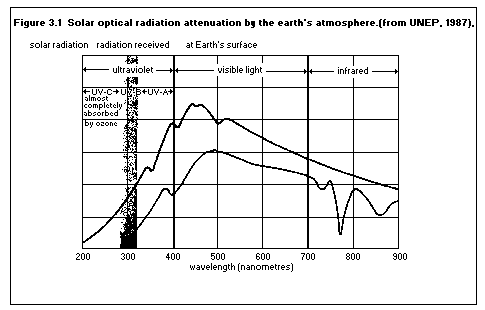
Solar UV undergoes absorption and scattering as it passes through
the earth's atmosphere with absorption by molecular oxygen and ozone
being the most important processes. The ozone layer prevents almost
all UV of wavelengths lamda < 290 nm and a substantial fraction (in
excess of 90% of the total energy) from 290 - 315 nm from reaching
the earth's surface. Thus the terrestrial environment is exposed to
UV between 290 nm and 400 nm. The spectrum, both before passage
through the atmosphere and at sea level, is shown in figure 3.1
The solar radiation reaching the top of the earth's atmosphere
is affected by the solar output and the earth-sun distance.
Variations in solar output are much smaller than the variations
caused by atmospheric attenuation factors. The irradiance incident
on the top of the earth's atmosphere is summarised in table 3.1, but
will vary with the exact distance of the earth from the sun at a
particular time. The extreme values associated with this variation
are approximately ± 3.3% above and below the annual mean and occur
in January and July respectively.
Table 3.1 Spectral distribution of solar radiation prior to
attenuation by the earth's atmosphere (Frederick et al., 1989).
Wavelength band Irradiance (W m-2) Percent of total
UVC 6.4 0.5
UVB 21.1 1.5
UVA 85.7 6.3
Total UV 113.2 8.3
Visible and IR 1254 91.7
The atmosphere has a profound effect on the irradiance which
reaches the surface of the earth. In January (in the northern
hemisphere) or July (in the southern hemisphere) when the solar
elevation is low, direct UV travels a longer path through the
atmosphere and a large amount of scattering occurs. In addition,
much of the resultant scattered UV propagates downwards to the
earth's surface at angles to the horizontal that are larger than the
solar elevation, hence travelling a shorter and less absorptive
path. This results in large ratios of scattered to direct UV. During
the summer the ratio of diffuse to direct UV is smaller.
2.4 Summary
UV is a part of the non-ionizing region of the electromagnetic
spectrum. Its position in the spectrum can be characterised by
wavelength and it is quantified using radiometric quantities and
units. Production of UV is possible by different mechanisms that are
all based on atomic or molecular excitation. Excitation in a medium
can be achieved by thermal, electrical or optical energy absorption.
Emission of radiation is caused either by de-excitation or by
stimulated synchronised processes.
3. UV SOURCES
3.1 The Sun
The sun is the main source of UV. The broad spectrum and
intensity of UV from the sun are due to the high temperature at its
surface and its size. The intensity of solar UV reaching the earth's
atmosphere would probably be lethal to most living organisms on the
earth's surface without the shielding afforded by the atmosphere.
Solar UV undergoes absorption and scattering as it passes through
the earth's atmosphere with absorption by molecular oxygen and ozone
being the most important processes. The ozone layer prevents almost
all UV of wavelengths lamda < 290 nm and a substantial fraction (in
excess of 90% of the total energy) from 290 - 315 nm from reaching
the earth's surface. Thus the terrestrial environment is exposed to
UV between 290 nm and 400 nm. The spectrum, both before passage
through the atmosphere and at sea level, is shown in figure 3.1
The solar radiation reaching the top of the earth's atmosphere
is affected by the solar output and the earth-sun distance.
Variations in solar output are much smaller than the variations
caused by atmospheric attenuation factors. The irradiance incident
on the top of the earth's atmosphere is summarised in table 3.1, but
will vary with the exact distance of the earth from the sun at a
particular time. The extreme values associated with this variation
are approximately ± 3.3% above and below the annual mean and occur
in January and July respectively.
Table 3.1 Spectral distribution of solar radiation prior to
attenuation by the earth's atmosphere (Frederick et al., 1989).
Wavelength band Irradiance (W m-2) Percent of total
UVC 6.4 0.5
UVB 21.1 1.5
UVA 85.7 6.3
Total UV 113.2 8.3
Visible and IR 1254 91.7
The atmosphere has a profound effect on the irradiance which
reaches the surface of the earth. In January (in the northern
hemisphere) or July (in the southern hemisphere) when the solar
elevation is low, direct UV travels a longer path through the
atmosphere and a large amount of scattering occurs. In addition,
much of the resultant scattered UV propagates downwards to the
earth's surface at angles to the horizontal that are larger than the
solar elevation, hence travelling a shorter and less absorptive
path. This results in large ratios of scattered to direct UV. During
the summer the ratio of diffuse to direct UV is smaller.
 3.1.1 Factors affecting solar UV levels
The total solar UV reaching the earth's surface, termed global
UV can be divided into two components: direct and diffuse. Global UV
reaching a horizontal surface is the quantity most often measured.
For biological entities such as people and trees, UV hitting
cylindrical or spherical surfaces may be more important.
The amount and spectral distribution of solar UV irradiance
reaching the earth's surface depends on a number of factors,
including
(a) wavelength of the UV
(b) solar zenith angle, which depends on latitude, date of the year
and time of day
(c) solar source spectrum incident at the top of the atmosphere (d)
ozone column thickness and vertical distribution
(e) molecular absorption and scattering (including localized
gaseous pollutants)
(f) aerosol absorption and scattering (including anthropogenic
(g) absorption, scattering and reflection by clouds
(h) reflectance characteristics (albedo) of the ground
(i) shadowing by surrounding objects
(j) altitude above sea level
The presence of cloud cover, air pollution, haze, or even
scattered clouds, plays a significant role in attenuating UV. UVB
and UVA irradiances are reduced due to scattering by water droplets
and/or ice crystals in the clouds. Clouds can block a significant
portion of the UV which would have otherwise reached the surface.
Cloud cover and type are highly variable. The transmission of
UV radiation through clouds depends on cloud height, type and
optical density. The resultant effect on UV transmission is
difficult to assess particularly in the case of partial cloudiness.
The effect of cloudiness on the solar irradiation of a horizontal
plane can be approximated by
F = 1 - 0.056 C
where C is the total cloud index in tenths of sky covered from 0 to
10, 10 being complete sky cover. Thus for complete cloud cover, the
transmitted UV irradiance would decrease by 72% and for half cloud
cover by 44%. In extreme cases cloud cover can decrease UV
irradiance by over 90%. Estimates of the average reduction of UVB
due to clouds (relative to cloudless skies) based on satellite
measurements of backscattered solar UV are 30% at 60 degrees
latitude, 10% at 20 degrees latitude and 20% at the equator.
3.1.2 Ozone depletion effects
Over 90% of the total atmospheric ozone resides in the
stratosphere (the upper atmosphere). The total ozone column is
important for filtering solar UV. Only UVB is affected by changes in
the ozone column. UVC is almost completely absorbed by ozone and
oxygen in the atmosphere; even with severe ozone reduction UVC would
still be effectively absorbed by the remaining oxygen.
As stratospheric ozone levels decrease, the resulting higher
levels of solar UVB could increase the production of reactive OH
radical molecules, potentially increasing the chemical reactivity of
the troposphere. In polluted areas with sufficient concentrations of
oxides of nitrogen (NOx) (above 0.5 ppb by volume) and hydrocarbon
compounds this enhanced reactivity is calculated to result in
greater levels of tropospheric ozone and other potentially harmful
oxidized products, such as hydrogen peroxide and acids.
The amount of ozone in the free troposphere is increasing for
example over central europe and other parts of the globe. The effect
of these trends on UV radiation needs to be studied further (WMO,
1993). For high sun angles, tropospheric ozone is a more effective
absorber of UV radiation than stratospheric ozone because of the
increased path length of scattered radiation in the lower atmosphere
(Brühl & Crutzen, 1989).
The total ozone column is not uniform but varies with latitude
and time of year. At the same latitude, away from the equator and
tropics, the total ozone column tends to be greater in spring than
in autumn. Thus, even though the sun angles are the same on March 21
and September 21, the differences in total ozone column result in
more UVB in early autumn than in early spring. If cloud cover and
atmospheric pollution are not taken into account, in the tropics,
the relatively constant ozone column and the similar solar angles
throughout the year, result in little variation in solar UVB with
season.
Changes in total column ozone were observed by both ground
based and satellite based instruments from the WMO Global Ozone
Observing System. The measurements show pronounced ozone depletion
over Antarctica in months 9-12 (local early spring) (UNEP-WMO,
1989). Increases in biologically effective UVB were observed during
this same time period (NSF, 1993). Over representation of UVA in the
common broad band meter action spectrum, relative to most biological
action spectra, produces less pronounced relative and absolute
changes in assessed UVB. Therefore under cloud free conditions
common broad band meters are less sensitive in assessing the ground
level UVB effects of ozone depletion than detectors with a steeper
action spectrum response such as for DNA damage or for erythema.
3.1.3 Trends in UV
There have been no significant changes in ozone at the equator.
Total column ozone over the Northern mid-latitudes has decreased by
several percent over the past two decades. Efforts to detect changes
in UV over long time periods have failed due to changes in column
ozone.
Besides the problems indicated in the interpretation and
comparison of measurement data from different sites and sources,
there are also insufficient direct solar terrestrial UVB
measurements for constructing a global climatology or trend
assessment due to some of the following (Driscoll 1992):
(a) problems in establishing appropriate instrumentation: either
highly sophisticated and reliable spectral instruments or broad
band instruments which fairly represent the sensitivities of
different biological and chemical targets (with different
wavelength dependencies and sensitivity to different
orientations),
(b) difficulty of maintaining accurate field instrument
calibrations over many years, and
(c) practical limitations in establishing a global monitoring
network especially with the potential for disturbance from
locally polluted areas.
With insufficient spectral measurement data collected over long
periods, data from the more extensively used broad-band measurement
systems have been scrutinised. UV data from eight stations in the US
showed decreases in UVB between 0.5% and 1.1% per year during the
time period of 1974-1985 (Scotto et al., 1988). However, this
result does not agree with theoretical predictions and may be due to
problems in the long-term calibration of meters or local pollution
in mainly urban or semi-urban sites (Munkata, 1993; Smith & Ryan
1993). In Russia, a 12% decrease of UVB was observed in Moscow
between 1968 and 1983 with a concurrent 15% increase in turbidity
and a 13% increase in cloudiness. At the Jungfraujoch observing
station in the Swiss alps (3.6 km above sea level), increases of
0.7±0.2% per year in UVB were observed under clear sky conditions
between 1981 and 1989. A comparison of summer spectral data weighted
with the CIE erythemal action spectrum using the same
instrumentation at Lauder, New Zealand (45°S) and Neuherberg,
Germany (48°N) showed weighted UV irradiances were 1.6 times larger
in New Zealand due to decreased ozone column thickness (24% lower
than in Germany). Spectral filter measurements at 39o N between 1976
though 1990 showed large increases in monthly maximum values which
were not statistically significant (Correll et al., 1992).
3.1.4 Theoretical models
To overcome some of the deficiencies in interpreting the
results from solar UV measurements, theoretical models have been
used to predict UV levels. However, the theoretical determination of
the spectral distribution of global UV including the effects of
cloud cover and ground reflection is extremely complex. There are
three categories of models to predict UV transmission. Empirical and
semi-empirical models are useful for assessing daily or annual
erythemal doses as a function of latitude, solar zenith angle, cloud
cover and ground albedo. These models have been used for a long time
at a variety of locations. Two of the most common examples are the
models of Green et al. (1974) and Diffey (1977)
Two stream models simulate the radiative transfer through the
atmosphere. They have been well verified with field data from
Antarctica as well as mid-latitudes (e.g. Frederick et al., 1989).
Multistream models allow for the angular distribution of UV
transmission. This is useful for more detailed dosimetry (Stamnes
et al., 1988).
The maximum erythemally effective UVB irradiances, calculated
by the Diffey model, are shown in table 3.2 for the Northern
Hemisphere, as a function of latitude and time of year at sea level
(Driscoll 1992). There are strong seasonal and latitudinal
variations in UVB. Under cloudless skies, the UVB is more intense in
summer and at all times of year is greater at lower latitudes. The
solar elevation angle determines the length of the path of the sun's
rays as they penetrate the atmosphere. When the sun is low in the
sky, the path through the atmosphere is longer and the filtering
action of the air is therefore increased. When the sun is directly
overhead, the sun's rays have the shortest path through the
atmosphere. Approximately 50% of the daily UV is received during the
middle four hours around noon when the sun is high in the sky
(Sliney, 1987).
The number of minimum erythemal doses (MEDs) in a 3 hour
exposure period around noon for fair skin is shown as a function of
latitude and time of year in table 3.3 for the Northern Hemisphere.
For example, at 55°N (Newcastle), a calculated value of 6 MED would
be received for a sensitive skin type in a 3 hour exposure around
noon on a clear day in July (1 MED per half hour exposure).
Comparing the values in table 3.2 with the results of
calculations using the model developed by Frederick et al. (1989)
for clear sky conditions at noon, the latter are typically 50%
higher. This may be due to differences in the biological action
spectra used or in the spectral irradiance data calculated. The R-B
meter response characteristic used by Frederick et al. (1989) does
not follow the CIE reference erythema action spectrum used in the
model developed by Diffey (1977) at wavelengths greater than 300 nm.
3.1.5 UV monitoring
Recent public and scientific concern about ozone depletion and
increased UV have lead to the establishment of many UV monitoring
centres in the last few years. Five years ago less than fifty UV
monitoring stations were operating around the world. Today more than
250 monitoring centres are underway for a variety of reasons.
Governmental agencies, scientific institutions, universities and
private groups have begun to monitor UV. The World Meteorological
Organization (WMO) has established a global network called Global
Atmosphere Watch (GAB). It presently has eight observatory stations
that make continuous spectral and broad band UV measurements. The
Global Environment Facility is supporting the creation of 10-15
additional stations in developing countries. Various national and
multi-national agencies are also operating and establishing UV
monitoring networks.
Table 3.2 Calculated clear sky noontime erythemally effective UVB
irradiances (mW m-2 CIE erythemally weighted) on a horizontal
surface as a function of latitude (°) and time of year for the sea
level in the Northern Hemisphere using typical ozone values
(Driscoll, 1992).
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
0 229 251 260 229 210 207 214 233 278 248 226 221
5 212 232 242 235 214 213 220 241 258 227 207 206
10 203 220 229 248 226 225 232 255 244 215 198 175
15 155 199 201 227 221 226 234 240 223 198 155 135
20 127 183 191 215 234 239 247 227 212 184 127 111
25 95 134 165 185 203 227 219 200 194 143 101 84
30 79 110 157 176 192 215 208 190 186 117 85 56
35 42 80 114 146 160 188 182 166 148 93 50 31
40 28 61 93 139 152 178 172 158 121 73 33 21
45 16 33 66 105 126 158 153 128 93 42 21 13
50 12 22 54 85 119 150 145 104 77 28 15 8
55 6 14 33 65 92 130 114 81 48 18 8 4
60 4 10 21 53 75 105 93 67 31 12 5 2
65 0 6 14 35 59 85 76 42 21 7 2 0
70 0 3 10 23 48 70 62 29 15 4 0 0
75 0 0 6 15 30 53 39 20 9 0 0 0
80 0 0 3 11 20 33 26 14 5 0 0 0
85 0 0 2 7 14 23 18 9 2 0 0 0
90 0 0 0 4 10 16 13 5 0 0 0 0
Table 3.3 Number of MEDs in a 3 h exposure period for a sensitive
skin type (1 MED = 200 J m-2 effective) for the erythemally
effective UVB irradiances given in table 3.2 as a function of
latitude (°) and time of year for the Northern Hemisphere (Driscoll
1992).
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
0 12 14 14 12 11 11 12 13 15 13 12 12
5 11 13 13 13 12 12 12 13 14 12 11 11
10 11 12 12 13 12 12 13 14 13 12 11 9
15 8 11 11 12 12 12 13 13 12 11 8 7
20 7 10 10 12 13 13 13 12 11 10 7 6
25 5 7 9 10 11 12 12 11 10 8 5 5
30 4 6 8 10 10 12 11 10 10 6 5 3
35 2 4 6 8 9 10 10 9 8 5 3 2
40 2 3 5 8 8 10 9 9 7 4 2 1
45 1 2 4 6 7 9 8 7 5 2 1 1
50 1 1 3 5 6 8 8 6 4 2 1 0
55 0 1 2 4 5 7 6 4 3 1 0 0
60 0 1 1 3 4 6 5 4 2 1 0 0
65 0 0 1 2 3 5 4 2 1 0 0 0
70 0 0 1 1 3 4 3 2 1 0 0 0
75 0 0 0 1 2 3 2 1 0 0 0 0
80 0 0 0 1 1 2 1 1 0 0 0 0
85 0 0 0 0 1 1 1 0 0 0 0 0
90 0 0 0 0 1 1 1 0 0 0 0 0
The reasons for monitoring UV are generally divided into the
following four areas:
(i) to provide information to the public on UV levels and
variations,
(ii) establishing a basic UV climatology,
(iii) studying cause and effects of UV transmission, and
(iv) detecting long term variability.
While these reasons are not mutually exclusive they can often
dictate the type of care taken with instrument maintenance. The
quality and care needed for trend detection requires a great deal
more effort than for UV measurements to provide public information.
There are many difficulties in making accurate field
measurements of ambient UV. Care must be taken that no part of the
instrument is shaded from either direct or indirect sun; nearby
surfaces must not change during the time period of monitoring; and
routine maintenance such as snow and debris removal must be
conducted systematically to avoid trend bias. The value of the
measurements for trend detection rely on diligent maintenance of the
site including co-located measurements. The data must have spectral,
temporal and angular resolution appropriate for validation of model
calculations and for evaluation of biological effects (spectral
resolution better than <1 nm). Broad band instruments are useful
for establishing climatology and operations where spectral
instruments are not practical.
Frequent and reliable calibration of the instrumentation is
necessary. UV monitoring for the detection of trends is difficult
and failure to detect a trend may be due to lack of awareness of
changes in instrument behaviour and correction of it.
Intercomparison of different types of UV monitoring instruments have
shown that individual instrument characteristics can cause
substantial differences in measurements. Further intercomparisons
are necessary to understand and reduce the uncertainties of
measurements.
3.1.6 Analysis of UV data
Atmospheric UV data are difficult to analyze. To date all UV
data have been analyzed separately by each investigator and in a
different manner. This variation in analysis makes the
interpretation and comparison of results difficult. Unfortunately,
the data are normally not easily obtained and re-analyzed for
comparative studies.
UV data pose difficult problems because they are not generally
independent and normally distributed, therefore robust time series
techniques should be used to analyze the data correctly. Standard
techniques, depending on how they are applied, can greatly over or
under estimate the confidence in the observed trends, as well as
supply spurious trends when used on UV data. Bishop (1992) has
suggested that using proper statistical techniques, at least ten
years of data will be necessary to correctly detect a trend of 5%
per decade. This estimate was made based on analysis of a single
station's data independently. However, analysing groups of data may
allow trend detection in less time.
3.1.7 Conclusions
The earth's atmosphere has a profound effect on the UV
irradiance reaching the earth's surface. It absorbs a large fraction
of the incident UV and changes part to diffuse radiation. Variations
in UV intensity depend on solar zenith angle, atmospheric ozone,
cloudiness, aerosol load and other factors.
One of the most important tools to determine climatological
values and long-term changes in solar UV radiation is the monitoring
of solar UV. Atmospheric parameters which modulate UV need to be
observed in parallel to help explain the changes observed in the
measurements. The existing data base is much too scarce to derive
climatological values and trends on global or regional scales.
Efforts are being made to establish national and international
networks for measurements of solar UV radiation.
Models that simulate typical values and predictions of changes
in UV radiation for different atmospheric conditions are also used.
The results from these models need to be compared with measurements.
3.2 Artificial Sources
Artificial sources of UV are commonplace. There are few
artificial sources that result in human exposure to UV greater than
that from the sun. However, exceptions are those used for medical
therapy and diagnosis, cosmetic tanning. Industrial sources are
generally effectively enclosed, but accidental exposure may occur.
There are very few non-laser sources of optical radiation that emit
UV solely.
Any unfiltered optical source, whose emissions are due to the
heating of a material e.g. a filament lamp, that emits significant
quantities of UV will also emit visible and infrared radiations. In
the case of high temperature tungsten halogen lamps biologically
significant amounts of shorter wavelength UVB are also emitted.
Essentially the same holds true for high intensity (gaseous)
discharge (HID) lamps. Some incandescent and HID lamps have
sufficient intrinsic filtration in the glass envelope of the lamp.
However, additional filtration, afforded by incorporation of the
lamp in a suitably filtered luminaire, may be necessary.
Most man-made sources of UV can be grouped together in the
categories shown below. The spectrum of the UV emitted varies from
one source to another
Incandescent sources
tungsten lamps
Gas discharges
mercury lamps (low-, medium- and high-pressure)
mercury lamps with metal halides
xenon lamps
hydrogen and deuterium lamps
flash tubes
Electric discharges
welding arcs
carbon arcs
Fluorescent lamps
fluorescent lighting tubes
fluorescent sunlamps (UVB emitters)
fluorescent UVA tubes
Lasers
excimer laser
dye laser
gas laser
3.2.1 Incandescent sources
When a material is heated a large number of energy transitions
occur within its molecules and optical photons are emitted. An
ideally efficient emitter (radiator) is termed a black body
radiator. The total radiant power and its spectral distribution
depend only on the temperature of the black body. The spectral
emissions in terms of spectral radiant exitance of a black body
radiator for different temperatures are illustrated in figure 3.2
and tabulated in table 3.4 (McKinlay et al., 1988). The wavelength
corresponding to the peak of the spectral emission of a black body
radiator varies inversely as its temperature and as the temperature
increases an increasing amount of UV is emitted. Incandescent
sources whose temperatures are greater than about 2900 K emit
significant amounts of UV with respect to possible effects on human
health. The optical emission of the sun corresponds approximately to
that of a black body radiator at a temperature of around 6000 K.
3.1.1 Factors affecting solar UV levels
The total solar UV reaching the earth's surface, termed global
UV can be divided into two components: direct and diffuse. Global UV
reaching a horizontal surface is the quantity most often measured.
For biological entities such as people and trees, UV hitting
cylindrical or spherical surfaces may be more important.
The amount and spectral distribution of solar UV irradiance
reaching the earth's surface depends on a number of factors,
including
(a) wavelength of the UV
(b) solar zenith angle, which depends on latitude, date of the year
and time of day
(c) solar source spectrum incident at the top of the atmosphere (d)
ozone column thickness and vertical distribution
(e) molecular absorption and scattering (including localized
gaseous pollutants)
(f) aerosol absorption and scattering (including anthropogenic
(g) absorption, scattering and reflection by clouds
(h) reflectance characteristics (albedo) of the ground
(i) shadowing by surrounding objects
(j) altitude above sea level
The presence of cloud cover, air pollution, haze, or even
scattered clouds, plays a significant role in attenuating UV. UVB
and UVA irradiances are reduced due to scattering by water droplets
and/or ice crystals in the clouds. Clouds can block a significant
portion of the UV which would have otherwise reached the surface.
Cloud cover and type are highly variable. The transmission of
UV radiation through clouds depends on cloud height, type and
optical density. The resultant effect on UV transmission is
difficult to assess particularly in the case of partial cloudiness.
The effect of cloudiness on the solar irradiation of a horizontal
plane can be approximated by
F = 1 - 0.056 C
where C is the total cloud index in tenths of sky covered from 0 to
10, 10 being complete sky cover. Thus for complete cloud cover, the
transmitted UV irradiance would decrease by 72% and for half cloud
cover by 44%. In extreme cases cloud cover can decrease UV
irradiance by over 90%. Estimates of the average reduction of UVB
due to clouds (relative to cloudless skies) based on satellite
measurements of backscattered solar UV are 30% at 60 degrees
latitude, 10% at 20 degrees latitude and 20% at the equator.
3.1.2 Ozone depletion effects
Over 90% of the total atmospheric ozone resides in the
stratosphere (the upper atmosphere). The total ozone column is
important for filtering solar UV. Only UVB is affected by changes in
the ozone column. UVC is almost completely absorbed by ozone and
oxygen in the atmosphere; even with severe ozone reduction UVC would
still be effectively absorbed by the remaining oxygen.
As stratospheric ozone levels decrease, the resulting higher
levels of solar UVB could increase the production of reactive OH
radical molecules, potentially increasing the chemical reactivity of
the troposphere. In polluted areas with sufficient concentrations of
oxides of nitrogen (NOx) (above 0.5 ppb by volume) and hydrocarbon
compounds this enhanced reactivity is calculated to result in
greater levels of tropospheric ozone and other potentially harmful
oxidized products, such as hydrogen peroxide and acids.
The amount of ozone in the free troposphere is increasing for
example over central europe and other parts of the globe. The effect
of these trends on UV radiation needs to be studied further (WMO,
1993). For high sun angles, tropospheric ozone is a more effective
absorber of UV radiation than stratospheric ozone because of the
increased path length of scattered radiation in the lower atmosphere
(Brühl & Crutzen, 1989).
The total ozone column is not uniform but varies with latitude
and time of year. At the same latitude, away from the equator and
tropics, the total ozone column tends to be greater in spring than
in autumn. Thus, even though the sun angles are the same on March 21
and September 21, the differences in total ozone column result in
more UVB in early autumn than in early spring. If cloud cover and
atmospheric pollution are not taken into account, in the tropics,
the relatively constant ozone column and the similar solar angles
throughout the year, result in little variation in solar UVB with
season.
Changes in total column ozone were observed by both ground
based and satellite based instruments from the WMO Global Ozone
Observing System. The measurements show pronounced ozone depletion
over Antarctica in months 9-12 (local early spring) (UNEP-WMO,
1989). Increases in biologically effective UVB were observed during
this same time period (NSF, 1993). Over representation of UVA in the
common broad band meter action spectrum, relative to most biological
action spectra, produces less pronounced relative and absolute
changes in assessed UVB. Therefore under cloud free conditions
common broad band meters are less sensitive in assessing the ground
level UVB effects of ozone depletion than detectors with a steeper
action spectrum response such as for DNA damage or for erythema.
3.1.3 Trends in UV
There have been no significant changes in ozone at the equator.
Total column ozone over the Northern mid-latitudes has decreased by
several percent over the past two decades. Efforts to detect changes
in UV over long time periods have failed due to changes in column
ozone.
Besides the problems indicated in the interpretation and
comparison of measurement data from different sites and sources,
there are also insufficient direct solar terrestrial UVB
measurements for constructing a global climatology or trend
assessment due to some of the following (Driscoll 1992):
(a) problems in establishing appropriate instrumentation: either
highly sophisticated and reliable spectral instruments or broad
band instruments which fairly represent the sensitivities of
different biological and chemical targets (with different
wavelength dependencies and sensitivity to different
orientations),
(b) difficulty of maintaining accurate field instrument
calibrations over many years, and
(c) practical limitations in establishing a global monitoring
network especially with the potential for disturbance from
locally polluted areas.
With insufficient spectral measurement data collected over long
periods, data from the more extensively used broad-band measurement
systems have been scrutinised. UV data from eight stations in the US
showed decreases in UVB between 0.5% and 1.1% per year during the
time period of 1974-1985 (Scotto et al., 1988). However, this
result does not agree with theoretical predictions and may be due to
problems in the long-term calibration of meters or local pollution
in mainly urban or semi-urban sites (Munkata, 1993; Smith & Ryan
1993). In Russia, a 12% decrease of UVB was observed in Moscow
between 1968 and 1983 with a concurrent 15% increase in turbidity
and a 13% increase in cloudiness. At the Jungfraujoch observing
station in the Swiss alps (3.6 km above sea level), increases of
0.7±0.2% per year in UVB were observed under clear sky conditions
between 1981 and 1989. A comparison of summer spectral data weighted
with the CIE erythemal action spectrum using the same
instrumentation at Lauder, New Zealand (45°S) and Neuherberg,
Germany (48°N) showed weighted UV irradiances were 1.6 times larger
in New Zealand due to decreased ozone column thickness (24% lower
than in Germany). Spectral filter measurements at 39o N between 1976
though 1990 showed large increases in monthly maximum values which
were not statistically significant (Correll et al., 1992).
3.1.4 Theoretical models
To overcome some of the deficiencies in interpreting the
results from solar UV measurements, theoretical models have been
used to predict UV levels. However, the theoretical determination of
the spectral distribution of global UV including the effects of
cloud cover and ground reflection is extremely complex. There are
three categories of models to predict UV transmission. Empirical and
semi-empirical models are useful for assessing daily or annual
erythemal doses as a function of latitude, solar zenith angle, cloud
cover and ground albedo. These models have been used for a long time
at a variety of locations. Two of the most common examples are the
models of Green et al. (1974) and Diffey (1977)
Two stream models simulate the radiative transfer through the
atmosphere. They have been well verified with field data from
Antarctica as well as mid-latitudes (e.g. Frederick et al., 1989).
Multistream models allow for the angular distribution of UV
transmission. This is useful for more detailed dosimetry (Stamnes
et al., 1988).
The maximum erythemally effective UVB irradiances, calculated
by the Diffey model, are shown in table 3.2 for the Northern
Hemisphere, as a function of latitude and time of year at sea level
(Driscoll 1992). There are strong seasonal and latitudinal
variations in UVB. Under cloudless skies, the UVB is more intense in
summer and at all times of year is greater at lower latitudes. The
solar elevation angle determines the length of the path of the sun's
rays as they penetrate the atmosphere. When the sun is low in the
sky, the path through the atmosphere is longer and the filtering
action of the air is therefore increased. When the sun is directly
overhead, the sun's rays have the shortest path through the
atmosphere. Approximately 50% of the daily UV is received during the
middle four hours around noon when the sun is high in the sky
(Sliney, 1987).
The number of minimum erythemal doses (MEDs) in a 3 hour
exposure period around noon for fair skin is shown as a function of
latitude and time of year in table 3.3 for the Northern Hemisphere.
For example, at 55°N (Newcastle), a calculated value of 6 MED would
be received for a sensitive skin type in a 3 hour exposure around
noon on a clear day in July (1 MED per half hour exposure).
Comparing the values in table 3.2 with the results of
calculations using the model developed by Frederick et al. (1989)
for clear sky conditions at noon, the latter are typically 50%
higher. This may be due to differences in the biological action
spectra used or in the spectral irradiance data calculated. The R-B
meter response characteristic used by Frederick et al. (1989) does
not follow the CIE reference erythema action spectrum used in the
model developed by Diffey (1977) at wavelengths greater than 300 nm.
3.1.5 UV monitoring
Recent public and scientific concern about ozone depletion and
increased UV have lead to the establishment of many UV monitoring
centres in the last few years. Five years ago less than fifty UV
monitoring stations were operating around the world. Today more than
250 monitoring centres are underway for a variety of reasons.
Governmental agencies, scientific institutions, universities and
private groups have begun to monitor UV. The World Meteorological
Organization (WMO) has established a global network called Global
Atmosphere Watch (GAB). It presently has eight observatory stations
that make continuous spectral and broad band UV measurements. The
Global Environment Facility is supporting the creation of 10-15
additional stations in developing countries. Various national and
multi-national agencies are also operating and establishing UV
monitoring networks.
Table 3.2 Calculated clear sky noontime erythemally effective UVB
irradiances (mW m-2 CIE erythemally weighted) on a horizontal
surface as a function of latitude (°) and time of year for the sea
level in the Northern Hemisphere using typical ozone values
(Driscoll, 1992).
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
0 229 251 260 229 210 207 214 233 278 248 226 221
5 212 232 242 235 214 213 220 241 258 227 207 206
10 203 220 229 248 226 225 232 255 244 215 198 175
15 155 199 201 227 221 226 234 240 223 198 155 135
20 127 183 191 215 234 239 247 227 212 184 127 111
25 95 134 165 185 203 227 219 200 194 143 101 84
30 79 110 157 176 192 215 208 190 186 117 85 56
35 42 80 114 146 160 188 182 166 148 93 50 31
40 28 61 93 139 152 178 172 158 121 73 33 21
45 16 33 66 105 126 158 153 128 93 42 21 13
50 12 22 54 85 119 150 145 104 77 28 15 8
55 6 14 33 65 92 130 114 81 48 18 8 4
60 4 10 21 53 75 105 93 67 31 12 5 2
65 0 6 14 35 59 85 76 42 21 7 2 0
70 0 3 10 23 48 70 62 29 15 4 0 0
75 0 0 6 15 30 53 39 20 9 0 0 0
80 0 0 3 11 20 33 26 14 5 0 0 0
85 0 0 2 7 14 23 18 9 2 0 0 0
90 0 0 0 4 10 16 13 5 0 0 0 0
Table 3.3 Number of MEDs in a 3 h exposure period for a sensitive
skin type (1 MED = 200 J m-2 effective) for the erythemally
effective UVB irradiances given in table 3.2 as a function of
latitude (°) and time of year for the Northern Hemisphere (Driscoll
1992).
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
0 12 14 14 12 11 11 12 13 15 13 12 12
5 11 13 13 13 12 12 12 13 14 12 11 11
10 11 12 12 13 12 12 13 14 13 12 11 9
15 8 11 11 12 12 12 13 13 12 11 8 7
20 7 10 10 12 13 13 13 12 11 10 7 6
25 5 7 9 10 11 12 12 11 10 8 5 5
30 4 6 8 10 10 12 11 10 10 6 5 3
35 2 4 6 8 9 10 10 9 8 5 3 2
40 2 3 5 8 8 10 9 9 7 4 2 1
45 1 2 4 6 7 9 8 7 5 2 1 1
50 1 1 3 5 6 8 8 6 4 2 1 0
55 0 1 2 4 5 7 6 4 3 1 0 0
60 0 1 1 3 4 6 5 4 2 1 0 0
65 0 0 1 2 3 5 4 2 1 0 0 0
70 0 0 1 1 3 4 3 2 1 0 0 0
75 0 0 0 1 2 3 2 1 0 0 0 0
80 0 0 0 1 1 2 1 1 0 0 0 0
85 0 0 0 0 1 1 1 0 0 0 0 0
90 0 0 0 0 1 1 1 0 0 0 0 0
The reasons for monitoring UV are generally divided into the
following four areas:
(i) to provide information to the public on UV levels and
variations,
(ii) establishing a basic UV climatology,
(iii) studying cause and effects of UV transmission, and
(iv) detecting long term variability.
While these reasons are not mutually exclusive they can often
dictate the type of care taken with instrument maintenance. The
quality and care needed for trend detection requires a great deal
more effort than for UV measurements to provide public information.
There are many difficulties in making accurate field
measurements of ambient UV. Care must be taken that no part of the
instrument is shaded from either direct or indirect sun; nearby
surfaces must not change during the time period of monitoring; and
routine maintenance such as snow and debris removal must be
conducted systematically to avoid trend bias. The value of the
measurements for trend detection rely on diligent maintenance of the
site including co-located measurements. The data must have spectral,
temporal and angular resolution appropriate for validation of model
calculations and for evaluation of biological effects (spectral
resolution better than <1 nm). Broad band instruments are useful
for establishing climatology and operations where spectral
instruments are not practical.
Frequent and reliable calibration of the instrumentation is
necessary. UV monitoring for the detection of trends is difficult
and failure to detect a trend may be due to lack of awareness of
changes in instrument behaviour and correction of it.
Intercomparison of different types of UV monitoring instruments have
shown that individual instrument characteristics can cause
substantial differences in measurements. Further intercomparisons
are necessary to understand and reduce the uncertainties of
measurements.
3.1.6 Analysis of UV data
Atmospheric UV data are difficult to analyze. To date all UV
data have been analyzed separately by each investigator and in a
different manner. This variation in analysis makes the
interpretation and comparison of results difficult. Unfortunately,
the data are normally not easily obtained and re-analyzed for
comparative studies.
UV data pose difficult problems because they are not generally
independent and normally distributed, therefore robust time series
techniques should be used to analyze the data correctly. Standard
techniques, depending on how they are applied, can greatly over or
under estimate the confidence in the observed trends, as well as
supply spurious trends when used on UV data. Bishop (1992) has
suggested that using proper statistical techniques, at least ten
years of data will be necessary to correctly detect a trend of 5%
per decade. This estimate was made based on analysis of a single
station's data independently. However, analysing groups of data may
allow trend detection in less time.
3.1.7 Conclusions
The earth's atmosphere has a profound effect on the UV
irradiance reaching the earth's surface. It absorbs a large fraction
of the incident UV and changes part to diffuse radiation. Variations
in UV intensity depend on solar zenith angle, atmospheric ozone,
cloudiness, aerosol load and other factors.
One of the most important tools to determine climatological
values and long-term changes in solar UV radiation is the monitoring
of solar UV. Atmospheric parameters which modulate UV need to be
observed in parallel to help explain the changes observed in the
measurements. The existing data base is much too scarce to derive
climatological values and trends on global or regional scales.
Efforts are being made to establish national and international
networks for measurements of solar UV radiation.
Models that simulate typical values and predictions of changes
in UV radiation for different atmospheric conditions are also used.
The results from these models need to be compared with measurements.
3.2 Artificial Sources
Artificial sources of UV are commonplace. There are few
artificial sources that result in human exposure to UV greater than
that from the sun. However, exceptions are those used for medical
therapy and diagnosis, cosmetic tanning. Industrial sources are
generally effectively enclosed, but accidental exposure may occur.
There are very few non-laser sources of optical radiation that emit
UV solely.
Any unfiltered optical source, whose emissions are due to the
heating of a material e.g. a filament lamp, that emits significant
quantities of UV will also emit visible and infrared radiations. In
the case of high temperature tungsten halogen lamps biologically
significant amounts of shorter wavelength UVB are also emitted.
Essentially the same holds true for high intensity (gaseous)
discharge (HID) lamps. Some incandescent and HID lamps have
sufficient intrinsic filtration in the glass envelope of the lamp.
However, additional filtration, afforded by incorporation of the
lamp in a suitably filtered luminaire, may be necessary.
Most man-made sources of UV can be grouped together in the
categories shown below. The spectrum of the UV emitted varies from
one source to another
Incandescent sources
tungsten lamps
Gas discharges
mercury lamps (low-, medium- and high-pressure)
mercury lamps with metal halides
xenon lamps
hydrogen and deuterium lamps
flash tubes
Electric discharges
welding arcs
carbon arcs
Fluorescent lamps
fluorescent lighting tubes
fluorescent sunlamps (UVB emitters)
fluorescent UVA tubes
Lasers
excimer laser
dye laser
gas laser
3.2.1 Incandescent sources
When a material is heated a large number of energy transitions
occur within its molecules and optical photons are emitted. An
ideally efficient emitter (radiator) is termed a black body
radiator. The total radiant power and its spectral distribution
depend only on the temperature of the black body. The spectral
emissions in terms of spectral radiant exitance of a black body
radiator for different temperatures are illustrated in figure 3.2
and tabulated in table 3.4 (McKinlay et al., 1988). The wavelength
corresponding to the peak of the spectral emission of a black body
radiator varies inversely as its temperature and as the temperature
increases an increasing amount of UV is emitted. Incandescent
sources whose temperatures are greater than about 2900 K emit
significant amounts of UV with respect to possible effects on human
health. The optical emission of the sun corresponds approximately to
that of a black body radiator at a temperature of around 6000 K.
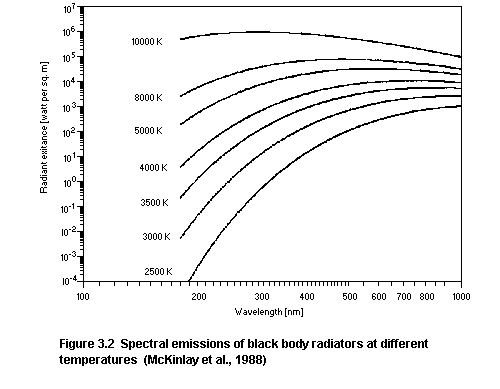 By integrating over all wavelengths the area under an absolute
plot of spectral radiant exitance against wavelength, the total
radiant exitance M can be determined by the formula:
M = sigmaT4
where sigma = Stefan-Boltzmann constant (5.67 x 10-3 W m-2 K-4)
and T = absolute temperature (K).
Table 3.4 Approximate radiant exitances of black body sources at
different temperatures
Temperature UVB/C UVA UVA
K 200-315 nm 315-340 nm 340-400 nm
[W m-2] [W m-2] [W m-2]
1500 8.6 x 10-5 5.6 x 10-4 2.7 x 10-2
2000 2.4 x 10-1 7.9 x 10-1 14.2
2500 29 62 634
3000 760 1147 8095
3500 8050 9220 5 x 104
4000 4.8 x 104 4.4 x 104 2 x 105
5000 6.3 x 105 4 x 105 1.4 x 106
6000 3.6 x 106 1.7 x 106 5 x 106
10000 1.4 x 108 3.2 x 107 6.9 x 107
In practice no material emits radiation with a black body
spectrum. However, tungsten at high temperatures (such as used for
the filaments of ordinary and tungsten halogen incandescent lamps)
and molten metals approximate to theoretical black bodies. Such
sources are termed grey bodies. The ratio of the actual spectral
radiant exitance to the theoretical black body spectral radiant
exitance is a measure of how closely the grey body approximates in
its spectral emission at a particular wavelength to a black body and
is termed the spectral emissivity (epsilonlamda). Similarly, by
summation over all wavelengths, the total radiant exitance M of a
material is given by
M = epsilon sigma T4
where epsilon is the emissivity.
The emissivity depends on the material and on its surface
structure, and is a function of wavelength. For example, for
tungsten at 3000 K (typical of a tungsten halogen lamp filament),
the emissivity varies from an average of about 0.45 in the UVA
region to about 0.2 in the IRB ( 1.4 - 3.0 µm) region.
The incandescent lamp is the oldest type of electric lamp still
in common use. The optical radiation from incandescent lamps results
from the heating of tungsten filaments which, with the exception of
lamps for some photographic and graphic arts applications, are not
heated to temperatures in excess of about 3000 K. Typical gas-filled
incandescent filament lamps operate between 2700 and 3000 K with
electrical input powers up to 500 W and the peaks of their spectral
emissions are in the infra-red (IRA) region. The emissions of UV by
tungsten filament lamps are generally negligible with respect to
human health.
In applications where greater power (up to 5 kW) is required,
tungsten-halogen (often called quartz-halogen) lamps are often used.
These are quartz envelope tungsten filament lamps to which a halogen
vapour (usually iodine) is added. The presence of the vapour enables
operation at an increased gas pressure compared with conventional
incandescent lamps, and evaporation from the filament is minimized.
These features result in improved luminous efficacy and longer life.
In order to operate efficiently, the temperature of the wall of the
bulb must be maintained at not less than 260 °C. The majority of the
bulbs of tungsten-halogen lamps are made from silica (quartz) whose
thermal properties are most suitable. The combination of filament
temperatures which are likely to be in the range 2900 to 3450 K and
quartz bulbs results in a significantly higher level of emission of
potentially harmful UV compared with ordinary tungsten filament
lamps. The incorporation of suitable secondary filtration to reduce
UV emissions to an acceptable level is an important feature of the
design of any illumination system using tungsten halogen lamps
(McKinlay et al., 1989).
3.2.2 Gaseous discharge sources
The electrical excitation of a gas or vapour is a typical
mechanism for generating optical radiation. An electric current is
passed through a gas or a mixture of gases ionized to produce
electrons and positive ions. Emissions are the result of electronic
transitions in the atoms of a material from low to high energy
states (absorption and excitation) followed by transitions from the
high to low energy states (de-excitation and emission). This process
is often combined with the process of luminescence, whereby the
characteristic (line) photon emissions of the gas are absorbed by a
luminescent material (luminophor or phosphor) which in turn emits
optical radiation, typically as a continuum over a range of longer
wavelengths. The 253.7 nm UV emission from a low pressure mercury
vapour discharge is used as a source of excitation in low pressure
fluorescent lamps. By raising the pressure of the discharge to a few
atmospheres the emission lines increasingly broaden effectively
forming a continuum. In some cases the 253.7 nm line emission will
be self-absorbed by the vapour of the discharge.
Low pressure discharge lamps
Germicidal lamps
The low pressure mercury-discharge lamp is often used for the
purpose of germicide and disinfection. Such lamps are very efficient
emitters of UV. Approximately 50% of the electrical power is
converted to UV of which up to 95% is emitted at a wavelength of
253.7 nm (see figure 3.3). Germicidal type lamps are available in a
range of sizes, shapes and powers. Small, low-wattage (5-10 W)
germicidal lamps are often used as fluorescence-inducing lamps for
the purposes of, for example, chromatographic analysis and the
fluorescence identification and authentication of documents. The
quartz envelopes of some lamps in this category transmit 185 nm
wavelength radiation characteristic of mercury vapour.
Fluorescent lamps
The most common application of the low-pressure discharge is
fluorescent lamps. These operate by means of a discharge between two
electrodes through a mixture of mercury vapour and a rare gas,
usually argon. Light is produced by conversion of 253.7 nm mercury
emission to longer wavelength radiations by means of a phosphor
coating on the inside of the glass envelope of the lamp. Lamps are
available with many different phosphors and envelopes to produce a
wide range of spectral emissions covering the visible (light), UVA
and UVB regions. While the continuum emissions of fluorescent lamps
are characteristic of the phosphors the narrow peak, spectral
emissions are dominated by the characteristic line emission spectrum
of the low-pressure mercury vapour discharge.
General lighting fluorescent lamps
These lamps are available in a range of physical sizes, powers
and phosphors. The range of phosphors includes a large selection of
"near white" and "special colour" lamps. In relation to other light
sources the fluorescent lamp is particularly efficient, with about
20% of the input energy resulting in useful light. Detailed spectral
analysis of the UV emissions of different general lighting
fluorescent lamps have shown that in general UVB and UVC emissions
are extremely low due to the marked attenuation of wavelengths <
320 nm afforded by the glass envelope.
By integrating over all wavelengths the area under an absolute
plot of spectral radiant exitance against wavelength, the total
radiant exitance M can be determined by the formula:
M = sigmaT4
where sigma = Stefan-Boltzmann constant (5.67 x 10-3 W m-2 K-4)
and T = absolute temperature (K).
Table 3.4 Approximate radiant exitances of black body sources at
different temperatures
Temperature UVB/C UVA UVA
K 200-315 nm 315-340 nm 340-400 nm
[W m-2] [W m-2] [W m-2]
1500 8.6 x 10-5 5.6 x 10-4 2.7 x 10-2
2000 2.4 x 10-1 7.9 x 10-1 14.2
2500 29 62 634
3000 760 1147 8095
3500 8050 9220 5 x 104
4000 4.8 x 104 4.4 x 104 2 x 105
5000 6.3 x 105 4 x 105 1.4 x 106
6000 3.6 x 106 1.7 x 106 5 x 106
10000 1.4 x 108 3.2 x 107 6.9 x 107
In practice no material emits radiation with a black body
spectrum. However, tungsten at high temperatures (such as used for
the filaments of ordinary and tungsten halogen incandescent lamps)
and molten metals approximate to theoretical black bodies. Such
sources are termed grey bodies. The ratio of the actual spectral
radiant exitance to the theoretical black body spectral radiant
exitance is a measure of how closely the grey body approximates in
its spectral emission at a particular wavelength to a black body and
is termed the spectral emissivity (epsilonlamda). Similarly, by
summation over all wavelengths, the total radiant exitance M of a
material is given by
M = epsilon sigma T4
where epsilon is the emissivity.
The emissivity depends on the material and on its surface
structure, and is a function of wavelength. For example, for
tungsten at 3000 K (typical of a tungsten halogen lamp filament),
the emissivity varies from an average of about 0.45 in the UVA
region to about 0.2 in the IRB ( 1.4 - 3.0 µm) region.
The incandescent lamp is the oldest type of electric lamp still
in common use. The optical radiation from incandescent lamps results
from the heating of tungsten filaments which, with the exception of
lamps for some photographic and graphic arts applications, are not
heated to temperatures in excess of about 3000 K. Typical gas-filled
incandescent filament lamps operate between 2700 and 3000 K with
electrical input powers up to 500 W and the peaks of their spectral
emissions are in the infra-red (IRA) region. The emissions of UV by
tungsten filament lamps are generally negligible with respect to
human health.
In applications where greater power (up to 5 kW) is required,
tungsten-halogen (often called quartz-halogen) lamps are often used.
These are quartz envelope tungsten filament lamps to which a halogen
vapour (usually iodine) is added. The presence of the vapour enables
operation at an increased gas pressure compared with conventional
incandescent lamps, and evaporation from the filament is minimized.
These features result in improved luminous efficacy and longer life.
In order to operate efficiently, the temperature of the wall of the
bulb must be maintained at not less than 260 °C. The majority of the
bulbs of tungsten-halogen lamps are made from silica (quartz) whose
thermal properties are most suitable. The combination of filament
temperatures which are likely to be in the range 2900 to 3450 K and
quartz bulbs results in a significantly higher level of emission of
potentially harmful UV compared with ordinary tungsten filament
lamps. The incorporation of suitable secondary filtration to reduce
UV emissions to an acceptable level is an important feature of the
design of any illumination system using tungsten halogen lamps
(McKinlay et al., 1989).
3.2.2 Gaseous discharge sources
The electrical excitation of a gas or vapour is a typical
mechanism for generating optical radiation. An electric current is
passed through a gas or a mixture of gases ionized to produce
electrons and positive ions. Emissions are the result of electronic
transitions in the atoms of a material from low to high energy
states (absorption and excitation) followed by transitions from the
high to low energy states (de-excitation and emission). This process
is often combined with the process of luminescence, whereby the
characteristic (line) photon emissions of the gas are absorbed by a
luminescent material (luminophor or phosphor) which in turn emits
optical radiation, typically as a continuum over a range of longer
wavelengths. The 253.7 nm UV emission from a low pressure mercury
vapour discharge is used as a source of excitation in low pressure
fluorescent lamps. By raising the pressure of the discharge to a few
atmospheres the emission lines increasingly broaden effectively
forming a continuum. In some cases the 253.7 nm line emission will
be self-absorbed by the vapour of the discharge.
Low pressure discharge lamps
Germicidal lamps
The low pressure mercury-discharge lamp is often used for the
purpose of germicide and disinfection. Such lamps are very efficient
emitters of UV. Approximately 50% of the electrical power is
converted to UV of which up to 95% is emitted at a wavelength of
253.7 nm (see figure 3.3). Germicidal type lamps are available in a
range of sizes, shapes and powers. Small, low-wattage (5-10 W)
germicidal lamps are often used as fluorescence-inducing lamps for
the purposes of, for example, chromatographic analysis and the
fluorescence identification and authentication of documents. The
quartz envelopes of some lamps in this category transmit 185 nm
wavelength radiation characteristic of mercury vapour.
Fluorescent lamps
The most common application of the low-pressure discharge is
fluorescent lamps. These operate by means of a discharge between two
electrodes through a mixture of mercury vapour and a rare gas,
usually argon. Light is produced by conversion of 253.7 nm mercury
emission to longer wavelength radiations by means of a phosphor
coating on the inside of the glass envelope of the lamp. Lamps are
available with many different phosphors and envelopes to produce a
wide range of spectral emissions covering the visible (light), UVA
and UVB regions. While the continuum emissions of fluorescent lamps
are characteristic of the phosphors the narrow peak, spectral
emissions are dominated by the characteristic line emission spectrum
of the low-pressure mercury vapour discharge.
General lighting fluorescent lamps
These lamps are available in a range of physical sizes, powers
and phosphors. The range of phosphors includes a large selection of
"near white" and "special colour" lamps. In relation to other light
sources the fluorescent lamp is particularly efficient, with about
20% of the input energy resulting in useful light. Detailed spectral
analysis of the UV emissions of different general lighting
fluorescent lamps have shown that in general UVB and UVC emissions
are extremely low due to the marked attenuation of wavelengths <
320 nm afforded by the glass envelope.
 Frequently in office and industrial environments where
fluorescent lamps are used, the luminaire assembly incorporates a
diffuser or controller. Three materials are commonly used in the
construction of diffusers; opal acrylic, opal styrene and opal
polycarbonate. Controllers are luminaire covers that are configured
with small prisms or lenses. Two commonly used materials are clear
acrylic and clear styrene. Some luminaires incorporate opal
(diffusing) sides and a clear figured [controller] base. The use of
diffusers and controllers results in the absorption and reflection
of the radiation emitted by the associated lamp. The UV-attenuating
properties of different diffusers is demonstrated by the measurement
data in table 3.5.
During the past few years the further development and improved
design of general lighting fluorescent lamps have been evident in
the production of compact fluorescent lamps. These lamps are
essentially low power small diameter fluorescent tubes folded in a
compact form. They are most readily available commercially with cool
white phosphors but other phosphors are also available. For a given
illuminance their spectral emissions of UV are essentially no
different from those of full size tubular fluorescent lamps as shown
in table 3.6.
Table 3.5 Measurements of UV irradiance from various diffusers/controllers
with a white fluorescent lamp as the source. Irradiance totals in each
waveband in mW m-2 (percentage in parentheses), (McKinlay et al., 1988).
Diffuser Type UVA UVB UVACGIH**
mW m-2 mW m-2 [mW m-2]effective
Bare lamp 22.32 (100d) 3.45 (100) 59 x 10-3 (100)
Clear acrylic+ 16.35 (73) 2.91 (84) 48 x 10-3 (81)
Clear styrene+ 2.87 (13) 0 (0) 0 (0)
Opal styrene* 0.92 (4) 3 x 10-3 (< 0.1) 0.02 x 10-3 (<0.1)
Opal polycarbonate* 0.20 (< 1) 12 x 10-3 (< 1) 0.09 x 10-3 (<1)
+ Surface figured with small prisms
* Reeded surface
** ACGIH occupational hazard weighted irradiance: 1 maximum permissible
exposure for an 8 h working day is equivalent to 10-3 [W m-2]effective ..
Table 3.6 Measurements of UV from compact fluorescent lamps,
normalised to an illuminance of 500 lux (Whillock et al., 1990)
Lamp type UVA UVB
mW m-2 µW m-2
Luma (7 W) LC7 47 x 103 0
Luma (7 W) LC7 with diffuser 197 0
Osram (11 W) Dulux EL 38 x 103 0.1
Philips (9 W) SL9 37 x 103 0
Sylvania (13 W) Lynx CFD 43 x 103 30.81
Thorn (16 W) 2D 54 x 103 2.48
Tungsram (16 W) Globulux 663 0
"Special" applications fluorescent lamps
Apart from a number of colour-rendering fluorescent lamps,
which are essentially variations of general lighting fluorescent
lamps, a number of special applications fluorescent lamps have been
developed.
A common example of a UVB emitting fluorescent lamp is the FS
type lamp with spectral emission shown in figure 3.4. Such lamps
were previously used for cosmetic tanning and are now often used as
a source of UVB in biological experiments. The blacklight lamp uses
a nickel/cobalt-oxide (Woods glass) envelope that is almost entirely
opaque to light. The phosphor chosen for this type of lamp emits
around 370 nm in the UVA. Such lamps are used for a number of
commercial, scientific and industrial fluorescence purposes as well
as for display and entertainment. Three types of fluorescent lamps
that emit UVA in printing and copying merit mention viz; lamps
suitable for diazo printing with a principal emission around 360 nm;
those intended for modern fast diazo printing with a main emission
around 420 nm; and those used for photocopying with predominantly
green emissions.
A medical treatment for hyperbilirubinaemia in neonates
(neonatal jaundice) consists of irradiating the newborn child with
phototherapy lamps emitting in the wavelength range approximately
400 to 470 nm. Some lamps used for this purpose have emission
spectra that extend into the UV region. Fluorescent lamps in
printing and copying include lamps for diazo printing with a
principal emission around 360 nm.
The development of a range of phosphors with enhanced UVA
emissions has led to the widespread use of fluorescent lamps in
sunbeds, solaria and PUVA (Psoralen + UVA) treatment cabinets.
High pressure discharge lamps
The designation "high pressure discharge" lamps is taken in
this monograph to include the families of lamps often called high
intensity discharge (HID), such as mercury vapour or metal halide
lamps, where an electric arc is not created. At still higher
pressures, arcs may be produced, e.g., the xenon or mercury compact
(short-arc) lamps.
Mercury and metal halide lamps
High-pressure mercury vapour lamps are widely used for lighting
in commerce, streets, displays, floodlighting and a large number of
printing, curing and other industrial applications. The spectral
emissions of the discharge are in the blue, green and yellow regions
of the spectrum and a large amount of UV is also generated. The
general construction of high-pressure mercury lamps is a fused
silica (quartz) discharge tube containing the mercury/argon vapour
discharge mounted inside an outer envelope of soda-lime or
borosilicate glass.
The outer glass envelope effectively absorbs most residual UV.
Consequently the quantity of potentially harmful UV emitted by such
lamps depends critically on the integrity of this envelope. In the
USA, but apparently not in Europe, it is a legally enforceable
manufacturing requirement that breakage of the outer envelope must
either cause the lamp to fail to operate, in which case the lamps
are described as "self-extinguishing" and are marked with the letter
"T", or if not "self-extinguishing" they should be marked with the
letter "R". In the latter case a warning notice must be included
with the packaging of the lamp (FDA., 1988). Data from measurements
made at 2 m from a mercury HID lamp with the outer envelope removed
illustrate the importance of this aspect of safety design, as shown
in table 3.7.
Frequently in office and industrial environments where
fluorescent lamps are used, the luminaire assembly incorporates a
diffuser or controller. Three materials are commonly used in the
construction of diffusers; opal acrylic, opal styrene and opal
polycarbonate. Controllers are luminaire covers that are configured
with small prisms or lenses. Two commonly used materials are clear
acrylic and clear styrene. Some luminaires incorporate opal
(diffusing) sides and a clear figured [controller] base. The use of
diffusers and controllers results in the absorption and reflection
of the radiation emitted by the associated lamp. The UV-attenuating
properties of different diffusers is demonstrated by the measurement
data in table 3.5.
During the past few years the further development and improved
design of general lighting fluorescent lamps have been evident in
the production of compact fluorescent lamps. These lamps are
essentially low power small diameter fluorescent tubes folded in a
compact form. They are most readily available commercially with cool
white phosphors but other phosphors are also available. For a given
illuminance their spectral emissions of UV are essentially no
different from those of full size tubular fluorescent lamps as shown
in table 3.6.
Table 3.5 Measurements of UV irradiance from various diffusers/controllers
with a white fluorescent lamp as the source. Irradiance totals in each
waveband in mW m-2 (percentage in parentheses), (McKinlay et al., 1988).
Diffuser Type UVA UVB UVACGIH**
mW m-2 mW m-2 [mW m-2]effective
Bare lamp 22.32 (100d) 3.45 (100) 59 x 10-3 (100)
Clear acrylic+ 16.35 (73) 2.91 (84) 48 x 10-3 (81)
Clear styrene+ 2.87 (13) 0 (0) 0 (0)
Opal styrene* 0.92 (4) 3 x 10-3 (< 0.1) 0.02 x 10-3 (<0.1)
Opal polycarbonate* 0.20 (< 1) 12 x 10-3 (< 1) 0.09 x 10-3 (<1)
+ Surface figured with small prisms
* Reeded surface
** ACGIH occupational hazard weighted irradiance: 1 maximum permissible
exposure for an 8 h working day is equivalent to 10-3 [W m-2]effective ..
Table 3.6 Measurements of UV from compact fluorescent lamps,
normalised to an illuminance of 500 lux (Whillock et al., 1990)
Lamp type UVA UVB
mW m-2 µW m-2
Luma (7 W) LC7 47 x 103 0
Luma (7 W) LC7 with diffuser 197 0
Osram (11 W) Dulux EL 38 x 103 0.1
Philips (9 W) SL9 37 x 103 0
Sylvania (13 W) Lynx CFD 43 x 103 30.81
Thorn (16 W) 2D 54 x 103 2.48
Tungsram (16 W) Globulux 663 0
"Special" applications fluorescent lamps
Apart from a number of colour-rendering fluorescent lamps,
which are essentially variations of general lighting fluorescent
lamps, a number of special applications fluorescent lamps have been
developed.
A common example of a UVB emitting fluorescent lamp is the FS
type lamp with spectral emission shown in figure 3.4. Such lamps
were previously used for cosmetic tanning and are now often used as
a source of UVB in biological experiments. The blacklight lamp uses
a nickel/cobalt-oxide (Woods glass) envelope that is almost entirely
opaque to light. The phosphor chosen for this type of lamp emits
around 370 nm in the UVA. Such lamps are used for a number of
commercial, scientific and industrial fluorescence purposes as well
as for display and entertainment. Three types of fluorescent lamps
that emit UVA in printing and copying merit mention viz; lamps
suitable for diazo printing with a principal emission around 360 nm;
those intended for modern fast diazo printing with a main emission
around 420 nm; and those used for photocopying with predominantly
green emissions.
A medical treatment for hyperbilirubinaemia in neonates
(neonatal jaundice) consists of irradiating the newborn child with
phototherapy lamps emitting in the wavelength range approximately
400 to 470 nm. Some lamps used for this purpose have emission
spectra that extend into the UV region. Fluorescent lamps in
printing and copying include lamps for diazo printing with a
principal emission around 360 nm.
The development of a range of phosphors with enhanced UVA
emissions has led to the widespread use of fluorescent lamps in
sunbeds, solaria and PUVA (Psoralen + UVA) treatment cabinets.
High pressure discharge lamps
The designation "high pressure discharge" lamps is taken in
this monograph to include the families of lamps often called high
intensity discharge (HID), such as mercury vapour or metal halide
lamps, where an electric arc is not created. At still higher
pressures, arcs may be produced, e.g., the xenon or mercury compact
(short-arc) lamps.
Mercury and metal halide lamps
High-pressure mercury vapour lamps are widely used for lighting
in commerce, streets, displays, floodlighting and a large number of
printing, curing and other industrial applications. The spectral
emissions of the discharge are in the blue, green and yellow regions
of the spectrum and a large amount of UV is also generated. The
general construction of high-pressure mercury lamps is a fused
silica (quartz) discharge tube containing the mercury/argon vapour
discharge mounted inside an outer envelope of soda-lime or
borosilicate glass.
The outer glass envelope effectively absorbs most residual UV.
Consequently the quantity of potentially harmful UV emitted by such
lamps depends critically on the integrity of this envelope. In the
USA, but apparently not in Europe, it is a legally enforceable
manufacturing requirement that breakage of the outer envelope must
either cause the lamp to fail to operate, in which case the lamps
are described as "self-extinguishing" and are marked with the letter
"T", or if not "self-extinguishing" they should be marked with the
letter "R". In the latter case a warning notice must be included
with the packaging of the lamp (FDA., 1988). Data from measurements
made at 2 m from a mercury HID lamp with the outer envelope removed
illustrate the importance of this aspect of safety design, as shown
in table 3.7.
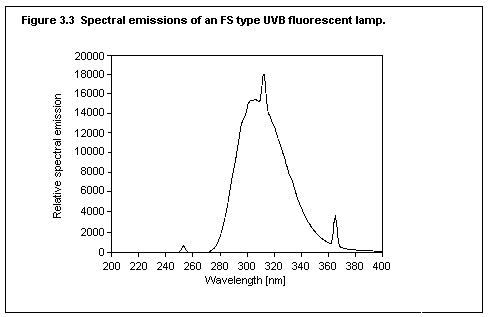 Table 3.7 UV emissions from HID mercury vapour general lighting (USA) lamps; effective irradiance
in mW m-2effective, (ACGIH), (Piltingsrud et al., 1978).
Test condition A B C D
Lamp type With outer Without With outer Without With outer Without With outer Without
bulb outer bulb bulb outer bulb bulb outer bulb bulb outer bulb
General Electric 2.5 110* < 0.1 160 12.5 3640 0.2 0.2
H400 A33-1
Clear
Westinghouse < 0.1 1040 < 0.1 190 0.2 3680 < 0.1 0.3
H33 GL 400/DX
White
General Electric < 0.2 75* < 0.1 180 0.5 3900* < 0.1 < 0.1*
H400D x 33-1
White
General Electric < 0.3 122* < 0.1 6* 1 510* < 0.1 < 0.1*
MV400/BUH
[Metal halide]
Test Conditions:
A: Lamp mounted vertically - measurements at 2 m on mid-line axis of lamp.
B: Lamp mounted horizontally - measurements at 2 m on central axis of lamp.
C: Lamp mounted horizontally in reflector shield with no face plate - measurements at 2 m on central axis of lamp.
D: Lamp mounted horizontally in reflector shield with glass face fitted - measurements
at 2 m on central axis of lamp.
* Lamps did not operate at normal intensity.
The family of metal halide lamps encompasses a number of
different types of high pressure mercury lamps whose discharges all
contain additives. The additives are most typically metal halides
chosen to produce either a strongly coloured emission (usually a
single halide), to produce a more broadly spectrally uniform
emission (multi-halide) or to enhance the UV (most often UVA)
emission. Compared with ordinary high pressure mercury lamps the
luminous efficacies of metal halide lamps are high. They are used
for a range of industrial and commercial applications that include
photochemical processing, graphic and photographic illumination,
studio lighting, reprography and are also used for UVA cosmetic
tanning equipment, for some medical applications and for solar
radiation simulation. The UV irradiances of some metal halide lamps
used in filtered industrial applications requiring an activating
range of wavelengths between 320 and 440 nm, e.g. lithographic
platemaking and printed circuit photo-resist etching, are shown in
table 3.8 (McKinlay et al., 1988).
Table 3.8 UV irradiances measured at 1 m from typical graphics arts
metal halide mercury lamps (McKinlay et al., 1988)
Lamp type Power UVC UVB UVA
[W] [W m-2] [W m-2] [W m-2]
HPA 400 400 0.5 3.2 9.0
HPA 1000 930 2.3 9.0 23.0
HPA 2000 1750 4.5 19.0 48.0
The emission of a device incorporating iron additive halide
lamps for phototherapy is illustrated in figure 3.5. The importance
of incorporating filtration in such devices in order to remove
unwanted components of the UV spectrum is illustrated.
Table 3.7 UV emissions from HID mercury vapour general lighting (USA) lamps; effective irradiance
in mW m-2effective, (ACGIH), (Piltingsrud et al., 1978).
Test condition A B C D
Lamp type With outer Without With outer Without With outer Without With outer Without
bulb outer bulb bulb outer bulb bulb outer bulb bulb outer bulb
General Electric 2.5 110* < 0.1 160 12.5 3640 0.2 0.2
H400 A33-1
Clear
Westinghouse < 0.1 1040 < 0.1 190 0.2 3680 < 0.1 0.3
H33 GL 400/DX
White
General Electric < 0.2 75* < 0.1 180 0.5 3900* < 0.1 < 0.1*
H400D x 33-1
White
General Electric < 0.3 122* < 0.1 6* 1 510* < 0.1 < 0.1*
MV400/BUH
[Metal halide]
Test Conditions:
A: Lamp mounted vertically - measurements at 2 m on mid-line axis of lamp.
B: Lamp mounted horizontally - measurements at 2 m on central axis of lamp.
C: Lamp mounted horizontally in reflector shield with no face plate - measurements at 2 m on central axis of lamp.
D: Lamp mounted horizontally in reflector shield with glass face fitted - measurements
at 2 m on central axis of lamp.
* Lamps did not operate at normal intensity.
The family of metal halide lamps encompasses a number of
different types of high pressure mercury lamps whose discharges all
contain additives. The additives are most typically metal halides
chosen to produce either a strongly coloured emission (usually a
single halide), to produce a more broadly spectrally uniform
emission (multi-halide) or to enhance the UV (most often UVA)
emission. Compared with ordinary high pressure mercury lamps the
luminous efficacies of metal halide lamps are high. They are used
for a range of industrial and commercial applications that include
photochemical processing, graphic and photographic illumination,
studio lighting, reprography and are also used for UVA cosmetic
tanning equipment, for some medical applications and for solar
radiation simulation. The UV irradiances of some metal halide lamps
used in filtered industrial applications requiring an activating
range of wavelengths between 320 and 440 nm, e.g. lithographic
platemaking and printed circuit photo-resist etching, are shown in
table 3.8 (McKinlay et al., 1988).
Table 3.8 UV irradiances measured at 1 m from typical graphics arts
metal halide mercury lamps (McKinlay et al., 1988)
Lamp type Power UVC UVB UVA
[W] [W m-2] [W m-2] [W m-2]
HPA 400 400 0.5 3.2 9.0
HPA 1000 930 2.3 9.0 23.0
HPA 2000 1750 4.5 19.0 48.0
The emission of a device incorporating iron additive halide
lamps for phototherapy is illustrated in figure 3.5. The importance
of incorporating filtration in such devices in order to remove
unwanted components of the UV spectrum is illustrated.
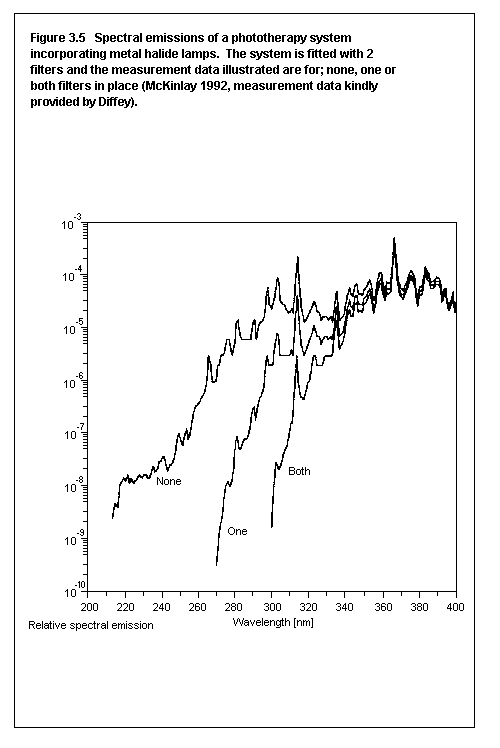 Xenon, compact and linear arcs
Where an optical source of very high radiance is required and
of small size a very high pressure arc lamp may be used. These have
a filling gas of mercury vapour, mercury vapour plus xenon gas or
xenon gas. Metal halide types are also available. Two physical types
are commonly used; the compact (short) arc and the linear arc. The
spectral emission of xenon lamps, which at wavelengths shorter than
infra-red, closely matches that of a black-body radiator at about
6000 K. This enables their use in photography and as solar radiation
simulators.
The spectral emission of xenon lamps, which at wavelengths
shorter than infrared closely matches that of a black body radiator
at about 6000 K, enables their use as solar radiation simulators.
Their emission spectrum is continuous from the UV through to the IR
regions. Large amounts of UVA, UVB and UVC are emitted by unfiltered
lamps to the extent that they can present a significant health
hazard if incorrectly used. The luminance of compact xenon arcs may
approach that of the sun and in some lamps with greater than 10 kW
rating the luminance may exceed that of the sun. They therefore
present a potentially severe retinal hazard if viewed.
3.2.3 Gas welding
Oil, coal and gas flames normally operate at temperatures below
about 2000 K and consequently emit virtually no UV. Oxyacetylene and
oxyhydrogen flames burn at much higher temperatures and emit UV
mostly in the UVA region.
3.2.4 Arc welding
By comparison with gas flame processes, the emissions of UV
from arc welding are very high (see figure 3.6) and many data on the
optical radiation emissions associated with a variety of electric
arc welding processes have been published (e.g., Sliney & Wolbarsht,
1980).
3.2.5 Lasers
All lasers have three basic components (see figure 3.7): (a) a
laser (active) medium; (b) an energy source (pumping system) and;
(c) a resonant optical cavity. A pumping system is necessary to
provide energy to electrons to raise them to excited states and
achieve population inversion. Optical pumping, using an intense
source of light such as a xenon flashtube; electron-collision
pumping, using an electrical discharge and; chemical pumping, using
the energy released from making and breaking chemical bonds, are all
used for this purpose. A resonant optical cavity is formed by
mirrors placed at each end of the laser medium. The construction is
such that the beam passes through the laser medium several times and
the number of emitted photons is amplified during each transit. One
of the mirrors is chosen to be partially transmitting thus enabling
part of the beam to be emitted from the cavity (figure 3.7).
Examples of lasers that emit UV are presented in table 3.9.
Table 3.9 UV laser emissions and characteristics,
(McKinlay, 1992)
Type Name Spectral emissions
Excimer Argon fluoride [(ArF) 193 nm
Krypton fluoride (KrF) 248 nm
Xenon chloride (XeCl) 308 nm
Xenon bromide (XeBr) 282 nm
Xenon fluoride (XeF) 351 nm
Krypton chloride (KrCl) 222 nm
Dye Excimer-, Nitrogen-, 300 nm-
Flash lamp pumped 340 nm-
Gas Nitrogen (N2) 337 nm
Helium cadmium (He-Cd) 325 nm
Gas ion Argon (Ar+0 333 nm-
357 nm
363 nm
Krypton (Kr+) 337.5 nm
350.7 nm
356.4 nm
A recent generation of UV lasers are the excimer lasers. The
active media used are rare gas halides such as ArF, KrCl, XeBr or
XeF. The molecules of those gases have only a short lifetime in the
ground state, however they are very stable in the excited state. As
a buffer medium, helium is often used. Population inversion is
easily achievable because the ground state is very unstable.
3.2.6 Sunbeds
Sunbeds are broadly used for cosmetic tanning purposes. The
expression "sunbed" includes tanning equipment consisting of a UV
emitting lamp or a number of such lamps incorporated in a bed,
canopy or panel, or any combination thereof. There are four distinct
types of lamps in use, each with different UV emission
characteristics. Those are UVA, low-pressure fluorescent tubes; UVA,
filtered high-intensity discharge lamps; UVB, low-pressure
fluorescent tubes; UVB, filtered high-intensity discharge lamps.
Xenon, compact and linear arcs
Where an optical source of very high radiance is required and
of small size a very high pressure arc lamp may be used. These have
a filling gas of mercury vapour, mercury vapour plus xenon gas or
xenon gas. Metal halide types are also available. Two physical types
are commonly used; the compact (short) arc and the linear arc. The
spectral emission of xenon lamps, which at wavelengths shorter than
infra-red, closely matches that of a black-body radiator at about
6000 K. This enables their use in photography and as solar radiation
simulators.
The spectral emission of xenon lamps, which at wavelengths
shorter than infrared closely matches that of a black body radiator
at about 6000 K, enables their use as solar radiation simulators.
Their emission spectrum is continuous from the UV through to the IR
regions. Large amounts of UVA, UVB and UVC are emitted by unfiltered
lamps to the extent that they can present a significant health
hazard if incorrectly used. The luminance of compact xenon arcs may
approach that of the sun and in some lamps with greater than 10 kW
rating the luminance may exceed that of the sun. They therefore
present a potentially severe retinal hazard if viewed.
3.2.3 Gas welding
Oil, coal and gas flames normally operate at temperatures below
about 2000 K and consequently emit virtually no UV. Oxyacetylene and
oxyhydrogen flames burn at much higher temperatures and emit UV
mostly in the UVA region.
3.2.4 Arc welding
By comparison with gas flame processes, the emissions of UV
from arc welding are very high (see figure 3.6) and many data on the
optical radiation emissions associated with a variety of electric
arc welding processes have been published (e.g., Sliney & Wolbarsht,
1980).
3.2.5 Lasers
All lasers have three basic components (see figure 3.7): (a) a
laser (active) medium; (b) an energy source (pumping system) and;
(c) a resonant optical cavity. A pumping system is necessary to
provide energy to electrons to raise them to excited states and
achieve population inversion. Optical pumping, using an intense
source of light such as a xenon flashtube; electron-collision
pumping, using an electrical discharge and; chemical pumping, using
the energy released from making and breaking chemical bonds, are all
used for this purpose. A resonant optical cavity is formed by
mirrors placed at each end of the laser medium. The construction is
such that the beam passes through the laser medium several times and
the number of emitted photons is amplified during each transit. One
of the mirrors is chosen to be partially transmitting thus enabling
part of the beam to be emitted from the cavity (figure 3.7).
Examples of lasers that emit UV are presented in table 3.9.
Table 3.9 UV laser emissions and characteristics,
(McKinlay, 1992)
Type Name Spectral emissions
Excimer Argon fluoride [(ArF) 193 nm
Krypton fluoride (KrF) 248 nm
Xenon chloride (XeCl) 308 nm
Xenon bromide (XeBr) 282 nm
Xenon fluoride (XeF) 351 nm
Krypton chloride (KrCl) 222 nm
Dye Excimer-, Nitrogen-, 300 nm-
Flash lamp pumped 340 nm-
Gas Nitrogen (N2) 337 nm
Helium cadmium (He-Cd) 325 nm
Gas ion Argon (Ar+0 333 nm-
357 nm
363 nm
Krypton (Kr+) 337.5 nm
350.7 nm
356.4 nm
A recent generation of UV lasers are the excimer lasers. The
active media used are rare gas halides such as ArF, KrCl, XeBr or
XeF. The molecules of those gases have only a short lifetime in the
ground state, however they are very stable in the excited state. As
a buffer medium, helium is often used. Population inversion is
easily achievable because the ground state is very unstable.
3.2.6 Sunbeds
Sunbeds are broadly used for cosmetic tanning purposes. The
expression "sunbed" includes tanning equipment consisting of a UV
emitting lamp or a number of such lamps incorporated in a bed,
canopy or panel, or any combination thereof. There are four distinct
types of lamps in use, each with different UV emission
characteristics. Those are UVA, low-pressure fluorescent tubes; UVA,
filtered high-intensity discharge lamps; UVB, low-pressure
fluorescent tubes; UVB, filtered high-intensity discharge lamps.
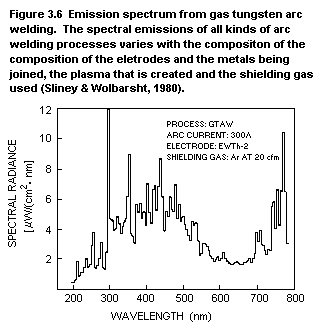
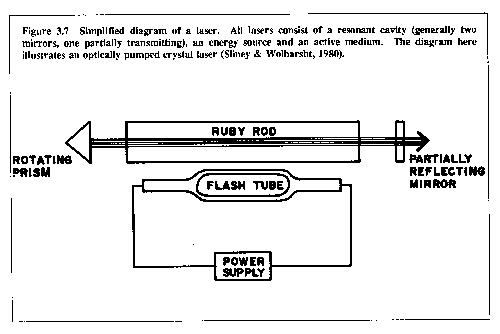 The emission characteristics and the health risks associated
with the use of each type of lamp are different. The last two lamp
types are associated with high levels of UVB and are now little
used. They have been almost universally replaced by the
predominantly UVA emitting lamp types.
4. HUMAN EXPOSURE
4.1 Sunlight
Outdoors, exposure to UV constantly changes during the day.
People are largely unaware of the degree of these changes. At noon,
when the sun is overhead, the level of UV at a wavelength of 300 nm
is ten times greater than at either three hours before (9 am) or
three hours after noon (3 pm). An untanned person with fair skin may
receive a mild sunburn in as little as 25 minutes at noon (depending
on the time of year and the latitude) but would have to lie in the
sun for at least two hours to receive the same dose after 3 pm. The
global biologically effective UV falling on a horizontal surface
occurs primarily during the midday hours, about 50% during the four
hours centred on noon-time zenith (Sliney, 1987).
Scattering of sunlight by air molecules (due to Raleigh
scattering) favours UV and blue light (hence the blue sky). For
longer pathlengths through the atmosphere when the sun is low in the
sky, more UV and sunlight is scattered. The sun which is white at
noonday becomes yellow and then orange as less UV and blue light are
present in the direct rays. When the sun is overhead staring at the
sun for 90 seconds would cause solar retinitis. A few hours later,
with the sun much lower in the sky, it would take many minutes to
reach a hazardous retinal dose, and it is virtually impossible to
cause any eye damage at sunset. Thus, the geometry of exposure as
well as the spectrum (hue) plays a major role in determining the
hazards from direct exposure from the sun (Sliney, 1983, 1986).
Estimation of an individual's lifetime UV exposure requires
knowledge of the ambient solar UV levels, history of outdoor
exposure and the relative exposures at the different anatomical
sites. Studies on the anatomical distribution of solar UV have been
reported (Diffey et al 1979; Rosenthal et al., 1985; Holman et
al., 1983; Gies et al 1992a; Roy et al., 1988). The relative
doses at various body sites have been determined using UV sensitive
polysulphone film on rotating manikins and headforms. It was found
that even though the relative doses to the face and eyes are higher
in winter, due to the lower solar elevation, the absolute doses are
higher in summer. The presence of a brimmed hat reduced the face
exposure by a factor of at least two and the eye exposure was
reduced by a factor of 4 to 5 (Diffey et al., 1979, Roy et al
1988).
Polysulphone film badges have also been used to quantify the
solar UV exposure received by different subjects and results
compared to those calculated from personal diaries and measured
ambient solar UVB (Gies et al., 1992a). In general, when UV
exposure activities took place under close supervision, good
correlations were obtained between the polysulphone badges and the
ambient/diaries approach. Results from a recent study (Roy and Gies,
1993), of indoor, outdoor and retired workers indicated that
exposures to badge locations of up to 30% of ambient were recorded.
Through studies of this type, knowledge is gained on the amount and
pattern of exposure from routine activities and this can then be
applied in the design of educational campaigns to modify outdoor
behaviour and reduce UV exposure. The use of polysulphone or CR39
plastics (Wong et al., 1989; Sydenham et al., 1991) as
contact-lens dosimeters have been proposed, but lack of sensitivity
and personal comfort has discouraged their use in field studies.
4.2 Skin Exposure Geometry
UV incident on human skin can follow one of three courses, it
can undergo absorption, reflection, or scattering. Thus, the actual
radiant exposure received by the various layers of the skin will be
lower than the incident exposure. Reflection not only occurs at the
surface of the stratum corneum, but at all interfaces changing in
refractive index. Scattering occurs because of the different
structural elements, such as hair follicles and sebaceous glands,
and also by cellular components, such as mitochondria and ribosomes.
The remaining UV can penetrate into deeper skin layers.
UV penetrates into the dermis exposing a variety of cells and
structures, depending in part on the thickness of the human stratum
corneum and epidermis. The depth of penetration is wavelength
dependent the longer the wavelength the deeper the penetration
(Bruls et al., 1984). From figure 4.1 it is seen that the same
incident exposure of UVA or UVB radiation will result in a higher
actual exposure of UVA than UVB at a given depth. For example, if
the incident UV exposure was 100 kJ m-2 then 50 kJ m-2 of 365 nm
radiation would be present at a depth of 30 µm and only 19 kJ m-2
at a depth of 70 µm. For 313 nm radiation only 33 kJ m-2 would be
present at 30 µm and 9.5 kJ m-2 at 70 µm. While less than 1% of
the UVC wavelengths can barely penetrate the epidermal layer (Bruls
et al., 1984), about 1% of the incident UVA dose can penetrate
into the subcutaneous tissue (Parrish et al., 1978). The
distribution and size of melanin particles also plays an important
role in protecting epidermal cells. Melanin particles have a
distribution within the stratum corneum and epidermal cells
depending upon skin type. In dark skin types (5 and 6) these
particles are positioned within cells to provide optimum optical
protection for the cell nuclei and in adequate size in the stratum
corneum (Kollias et al., 1991).
The emission characteristics and the health risks associated
with the use of each type of lamp are different. The last two lamp
types are associated with high levels of UVB and are now little
used. They have been almost universally replaced by the
predominantly UVA emitting lamp types.
4. HUMAN EXPOSURE
4.1 Sunlight
Outdoors, exposure to UV constantly changes during the day.
People are largely unaware of the degree of these changes. At noon,
when the sun is overhead, the level of UV at a wavelength of 300 nm
is ten times greater than at either three hours before (9 am) or
three hours after noon (3 pm). An untanned person with fair skin may
receive a mild sunburn in as little as 25 minutes at noon (depending
on the time of year and the latitude) but would have to lie in the
sun for at least two hours to receive the same dose after 3 pm. The
global biologically effective UV falling on a horizontal surface
occurs primarily during the midday hours, about 50% during the four
hours centred on noon-time zenith (Sliney, 1987).
Scattering of sunlight by air molecules (due to Raleigh
scattering) favours UV and blue light (hence the blue sky). For
longer pathlengths through the atmosphere when the sun is low in the
sky, more UV and sunlight is scattered. The sun which is white at
noonday becomes yellow and then orange as less UV and blue light are
present in the direct rays. When the sun is overhead staring at the
sun for 90 seconds would cause solar retinitis. A few hours later,
with the sun much lower in the sky, it would take many minutes to
reach a hazardous retinal dose, and it is virtually impossible to
cause any eye damage at sunset. Thus, the geometry of exposure as
well as the spectrum (hue) plays a major role in determining the
hazards from direct exposure from the sun (Sliney, 1983, 1986).
Estimation of an individual's lifetime UV exposure requires
knowledge of the ambient solar UV levels, history of outdoor
exposure and the relative exposures at the different anatomical
sites. Studies on the anatomical distribution of solar UV have been
reported (Diffey et al 1979; Rosenthal et al., 1985; Holman et
al., 1983; Gies et al 1992a; Roy et al., 1988). The relative
doses at various body sites have been determined using UV sensitive
polysulphone film on rotating manikins and headforms. It was found
that even though the relative doses to the face and eyes are higher
in winter, due to the lower solar elevation, the absolute doses are
higher in summer. The presence of a brimmed hat reduced the face
exposure by a factor of at least two and the eye exposure was
reduced by a factor of 4 to 5 (Diffey et al., 1979, Roy et al
1988).
Polysulphone film badges have also been used to quantify the
solar UV exposure received by different subjects and results
compared to those calculated from personal diaries and measured
ambient solar UVB (Gies et al., 1992a). In general, when UV
exposure activities took place under close supervision, good
correlations were obtained between the polysulphone badges and the
ambient/diaries approach. Results from a recent study (Roy and Gies,
1993), of indoor, outdoor and retired workers indicated that
exposures to badge locations of up to 30% of ambient were recorded.
Through studies of this type, knowledge is gained on the amount and
pattern of exposure from routine activities and this can then be
applied in the design of educational campaigns to modify outdoor
behaviour and reduce UV exposure. The use of polysulphone or CR39
plastics (Wong et al., 1989; Sydenham et al., 1991) as
contact-lens dosimeters have been proposed, but lack of sensitivity
and personal comfort has discouraged their use in field studies.
4.2 Skin Exposure Geometry
UV incident on human skin can follow one of three courses, it
can undergo absorption, reflection, or scattering. Thus, the actual
radiant exposure received by the various layers of the skin will be
lower than the incident exposure. Reflection not only occurs at the
surface of the stratum corneum, but at all interfaces changing in
refractive index. Scattering occurs because of the different
structural elements, such as hair follicles and sebaceous glands,
and also by cellular components, such as mitochondria and ribosomes.
The remaining UV can penetrate into deeper skin layers.
UV penetrates into the dermis exposing a variety of cells and
structures, depending in part on the thickness of the human stratum
corneum and epidermis. The depth of penetration is wavelength
dependent the longer the wavelength the deeper the penetration
(Bruls et al., 1984). From figure 4.1 it is seen that the same
incident exposure of UVA or UVB radiation will result in a higher
actual exposure of UVA than UVB at a given depth. For example, if
the incident UV exposure was 100 kJ m-2 then 50 kJ m-2 of 365 nm
radiation would be present at a depth of 30 µm and only 19 kJ m-2
at a depth of 70 µm. For 313 nm radiation only 33 kJ m-2 would be
present at 30 µm and 9.5 kJ m-2 at 70 µm. While less than 1% of
the UVC wavelengths can barely penetrate the epidermal layer (Bruls
et al., 1984), about 1% of the incident UVA dose can penetrate
into the subcutaneous tissue (Parrish et al., 1978). The
distribution and size of melanin particles also plays an important
role in protecting epidermal cells. Melanin particles have a
distribution within the stratum corneum and epidermal cells
depending upon skin type. In dark skin types (5 and 6) these
particles are positioned within cells to provide optimum optical
protection for the cell nuclei and in adequate size in the stratum
corneum (Kollias et al., 1991).
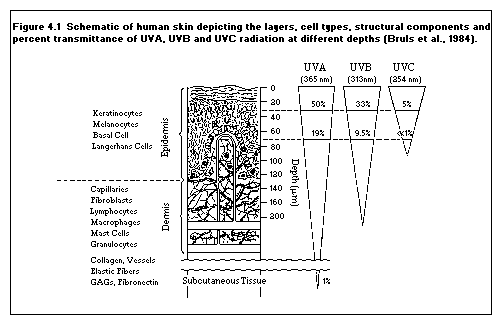 4.3 Ocular Exposure Geometry
People seldom look directly at the sun when it is overhead and
very hazardous to view. It is not very hazardous to view when the
sun is low in the sky and falls within the normal field-of-view.
When the sun is more than 10° above the horizon, the natural
tendency is to partially close the eyelids or squint (called squint
reflex), thus shielding the retina from direct exposure. These
factors reduce the exposure to the cornea to a maximum of about 5%
of that falling upon the exposed top of head (Sliney, 1986). If the
squint and other behavioural factors are not considered, the dose to
the eyelid would be approximately 20% of the dose falling on a
horizontal surface.
Although the cornea is more sensitive to UV injury than the
skin, people seldom experiences a corneal burn when in sunlight.
Using the action spectrum for human photokeratitis and
mathematically weighting this with the midday solar spectrum, the
time to achieve the threshold for photokeratitis is about 100
seconds (Sliney, 1987, Rosenthal et al., 1988). Again, the
geometry of exposure precludes photokeratitis except when ground
reflectance exceeds approximately 10%. When the sun is overhead and
UV exposure is most severe, the brow ridge and upper lid shield the
cornea, and if the eye is turned away from the sun, the more intense
scattered UV from overhead strikes the cornea at a grazing angle of
incidence where most is reflected and little is absorbed (Sliney,
1983). Only when the incident UV rays are parallel to the pupillary
axis are most rays (approx. 98%) absorbed.
When looking at snow, UV is reflected directly into the eye;
hence, the traditional eye protector of the Inuit or Eskimo, the
slit, in whalebone or in a seal-skin mask, provided geometrical
rather than spectral protection against UV exposure (Hedblom, 1961).
The lack of protection above and to the sides of sunglasses is a
serious shortcoming. However, to obtain a quantitative idea of this
component of exposure to the eye, measurements were made using a
simulated ocular geometry in sunlight (Sliney, 1986). The human eye
received 10 to 25 % of the UV dose when wearing glasses with lenses
opaque to UV compared to no lens in the glasses. Therefore, unless
goggles with side-shields are used, UV transmission factors in
lenses much less than 2-5 % do not provide the eye protection
suggested by the transmission factor (Sliney, 1986).
The strong dependence of reflectance with angle of incidence is
termed Fresnel's Law of Reflection. This law not only explains the
survival of the cornea exposed overhead to UV, but also the glare
experienced over water. When the sun is overhead, water reflects
about 2% of the UV upward (Sliney, 1986). When low in the sky, much
of the sunlight is reflected while the UV and blue light are
filtered by the atmosphere. Nevertheless, the strong reflections
from water at these low sun angles create discomfort glare and UV
exposure of the cornea is further reduced because of the squint.
If dark lenses are placed over the eyes, the natural aversion
to bright light, which leads to the squint reflex (that greatly
lowers retinal UV or exposure to the eye), would be disabled. This
may appear to be an unusual way to consider the comfort that shaded
lenses provide. However, poor sunglasses may actually lead to a
higher UV exposure (Sliney, 1983).
However, quantifying the protective value of the upper and
lower eye lids when they close to squint is difficult. In terms of
UV exposure, at least a twenty-fold reduction is likely. For
shielding the retina from the direct image of the sun, the upper lid
probably provides a protection factor exceeding a thousand (Sliney -
personal communication). If a brimmed hat is worn, the direct image
of the sun on the retina is rare and overhead UV exposure is
virtually eliminated. However, while using a hat the lid opens
further and ground reflection of UV could become important.
On an overcast day, the lids open wider, and although the UVB
irradiance is reduced by cloud cover, the actual UVB dose rate to
the eye from atmospheric scattering near the horizon may be reduced
by a factor of only two (Sliney, 1983). Hence, on a cloudy day the
eye may receive a greater UVB dose than on a bright sunny day.
However, a heavy overcast may attenuate the UVB sufficiently, that
this observation may not be true. As sunglasses are not typically
worn on an overcast day, one could argue that the concern over
sunglasses increasing total ocular exposure is unimportant. However,
sunglasses should have sufficient UV filtration so that ocular
exposure does not actually increase when they are worn on a sunny
day.
Eye and head movements can further reduce UV exposure. Most
humans in bright sunlight squint or avoid looking into the sun
sector of the sky. These behavioural and physiological factors are
not taken into account by simple UV measurements. There is an
obvious need to determine accurately the corneal and lenticular
exposure to ambient UV. Indeed, the results of previous
epidemiological studies of cataract may be questioned because of
inadequate dosimetry. Some epidemiological studies may have reached
incorrect conclusions regarding risk from UV or sunlight exposure by
assigning inaccurate exposure levels to population groups, because
they assumed that overhead UV exposure accurately predicts corneal
exposure.
4.4 Workplace
4.4.1 Outdoor work
Although humans have adapted and acclimatized to solar UV it
nevertheless represents the most hazardous source of optical
radiation likely to be encountered by the average person and with a
few exceptions by the worker. People who work outdoors will be
subjected to involuntary UV exposure. The highest exposure will
occur during the two hours period either side of noon. Workers must
be made aware of this and take appropriate precautions as discussed
in chapter 13.
In a study of various work and recreational situations
(Challoner et al., 1976; Diffey et al., 1982), it was found that
outdoor workers had the highest exposure, receiving approximately
10% of the ambient level, similar to that received during sailing
and sightseeing. Higher exposures were found for skiing (20%) and
the largest during sunbathing on a beach (80%). Office workers
received about 3% of the total ambient radiation with about half
that figure from weekend exposure (Leach et al., 1978). The actual
exposures vary depending on the time of day and year, duration and
frequency of exposure.
4.4.2 Indoor work
Examples of indoor workplace exposures to UV are given below.
Photoprocesses
Many industrial processes involve a photochemical component.
The large-scale nature of these processes often necessitates the use
of high-power (several kilowatts) lamps such as high pressure metal
halide lamps which emit significant amounts of UV (Diffey, 1990a).
The principal industrial applications of photopolymerisation include
the curing of protective coatings and inks and photoresists for
printed circuit boards. The curing of printing inks by exposure to
UV takes only a fraction of a second. UV drying units can be
installed between printing stations on a multicolour line, so that
each colour is dried before the next is applied. Another major use
of UV curing has been for metal decorating in the packaging industry
(Phillips, 1983). UVA is also used to inspect printed circuit boards
and integrated circuits in the electronics industry.
Sterilization
UV with wavelengths in the range 260-265 nm is the most
effective for sterilization and disinfection since it corresponds to
a maximum in the DNA absorption spectrum. Low-pressure mercury
discharge tubes are often used as the UV source as more than 90% of
the radiated energy lies in the 254 nm line, figure 3.2. These lamps
are often referred to as "germicidal lamps", "bactericidal lamps" or
simply "UVC lamps" (Diffey, 1990a).
UVC radiation has been used to disinfect sewage effluent,
drinking-water, water for the cosmetics industry and swimming pools.
Germicidal lamps are sometimes used inside microbiological safety
cabinets to inactivate airborne and surface microorganisms (Diffey,
1990a). The combination of UV and ozone has a very powerful
oxidizing action and can reduce the organic content of water to
extremely low levels (Phillips, 1983).
Welding
Welding equipment falls into two broad categories: gas welding
and electric arc welding. Only the latter process produces
significant levels of UV, the quality and quantity of which depend
primarily on the arc current, shielding gas and metals being welded
(Sliney & Wolbarsht, 1980). The levels of UV irradiance around
electric arc welding equipment are high; effective irradiance at 1 m
at an arc current of 400 A ranged from 1 to 50 W m-2 and the
unweighted UVA irradiance ranged from 3 to 70 W m-2, depending on
the type of welding and the metal being welded (Mariutti & Matzeu,
1987; Cox, 1987). It is not surprising therefore that most welders
at some time or another experience "arc eye" or "welder's flash"
(photokeratitis) and skin erythema. The effective irradiance at 0.3
m from many types of electric welding arcs operating at 150 A is
such that the maximum permissible exposure time for an 8-h working
period on unprotected eyes and skin varies from a few tenths of a
second to about 10 s, depending on the type of welding process and
the material used (Cox, 1987).
4.4.3 Research
Sources of UV are used by most experimental scientists engaged
in photobiology and photochemistry and in molecular biology. These
applications, in which the effect of UV irradiation on biological
and chemical species is of primary interest to the researcher, can
be differentiated from UV fluorescence by absorption techniques
where the effect is of secondary importance (Diffey, 1990a).
4.4.4 Commerce
Sunlamps emitting UV have been used for tanning, particularly
in northern Europe and North America. Prior to the mid-1970s, the
source of UV in sunlamps was usually an unfiltered mercury arc lamp
which emitted a broad spectrum of radiation, including large
quantities of UVB and UVC. There are four distinctly different types
of ultraviolet tanning lamps in use, each with different UV emission
characteristics viz.; low pressure UVA fluorescent lamps; filtered
high intensity discharge UVA lamps; low pressure UVB fluorescent
lamps and; filtered high intensity discharge UVB lamps (IRPA/INIRC
1991). The emission characteristics and the health risks associated
with each type of lamp are different (Diffey and McKinlay 1983, Gies
et al 1986, Diffey 1987). Tanning systems incorporating lamps that
emit predominantly UVB are now little used and have been almost
universally replaced by the low pressure UVA fluorescent and
filtered high intensity discharge UVA types. Tanning requires
deliberate exposure of the skin to UV; however, the eyes must be
protected. Similarly, staff working in tanning salons must ensure
their exposure is kept to a minimum.
Many contaminants of food products can be detected by UV
fluorescence techniques. For example, the bacterium Pseudomonas
aeruginosa, which causes rot in eggs, meat and fish, can be detected
by its yellow-green fluorescence under UVA irradiation. One of the
longest established uses of UVA fluorescence in public health is to
demonstrate contamination with rodent urine, which is highly
fluorescent.
Many flying insects are attracted by UVA radiation,
particularly in the region around 350 nm. This phenomenon is the
principle of electronic insect traps, in which a UVA fluorescent
lamp is mounted in a unit containing a high-voltage grid. The
insect, attracted by the UVA lamp, flies into the unit and is
electrocuted in the air gap between the high-voltage grid and a
grounded metal screen. Such units are commonly found in areas where
food is prepared and sold to the public (Diffey, 1990a).
UVA blacklight lamps are sometimes used in discotheques to
induce fluorescence in the skin and clothing of dancers. The levels
of UVA emitted are usually low (< 10 W m-2) (Diffey, 1990a).
Signatures, banknotes and other documents can be authenticated
by exposing them to UVA, under which they fluoresce. UVA exposure of
the user is normally to hands and irradiance is low (< 10 W m-2)
(Diffey, 1990a).
4.4.5 Medicine and dentistry
In both medical and dental applications of UV, the patient is
deliberately exposed to either treat or diagnose a disease or
disorder. While non-target areas of the patient must be covered,
care must also be taken to ensure staff are fully protected from UV
exposure (see chapter 13).
The diagnostic uses of UV are confined largely to fluorescing
of the skin and teeth. UV exposure is limited to small areas (< 15
cm in diameter) and the UVA radiation dose per examination is
probably no more than 5 x 104J m-2. Diagnostic techniques are
limited to the use of fluorescence to identify the presence of
various fungal and bacterial infectious agents on the skin or in
wounds. The source of UV most commonly used for this purpose is the
Woods lamp which emits predominantly UVA radiation (365 nm).
Exposure of the patient's skin to potentially
carcinogenically-effective UV is insignificant (Diffey, 1990a).
UV phototherapy is a well established method for the treatment
of a number of skin conditions and in particular for psoriasis
(Green et al., 1992). Two general types of UVB emitting lamps are
used in phototherapy, high intensity discharge (HID) mercury vapour
and mercury/metal halide vapour lamps and low pressure mercury
vapour fluorescent lamps. The spectral emissions of these different
types of lamps vary greatly and this, with other factors such as the
number and power of the lamps, the lamp to skin distance and the
individual sensitivity of the patient, determines the treatment
time. A treatment dose is chosen to cause erythema in a particular
patient. As treatment progresses and the skin becomes increasingly
acclimatised, individual treatment doses are increased accordingly.
Treatments may be given several times per week. Phototherapy has
also been used for the treatment of other skin conditions including
severe itching, acne, eczema, polymorphic light eruption (PLE),
pityriasis rosea and urticaria, and for renal failure (Green et
al., 1992).
Seasonal affective disorder (SAD) is frequently treated by
exposure to sources of high illumination. Most treatment regimes
have employed sources incorporating so-called "full spectrum
lighting" whose emissions contain small amounts of UVA and UVB
radiations (Terman et al., 1990).
PUVA
UV photochemotherapy is UV phototherapy used in conjunction
with the oral or topical application of a chemical
(photosensitising) agent to the patient. The most widely used
treatment is PUVA which involves the use of the photosensitising
agent 8-methoxypsoralen (8-MOP) in conjunction with UVA irradiation
for the treatment of psoriasis. The irradiation sources used for
such treatment incorporate either low pressure mercury vapour UVA
fluorescent lamps, or HID mercury vapour or mercury/metal halide
vapour lamps with an added filter to effectively attenuate the UVB.
The UVA irradiance on the skin from sources incorporating
fluorescent lamps is generally of the order of 60 W m-2 and for
the HID mercury systems, around 250 W m-2. Treatments consist of a
starting dose of between 500 and 40,000 J m-2, depending on skin
type, followed by further incremental doses two to three times
weekly. Total treatment radiant exposures of between 106 and 2.5
106 J m-2 have been reported for successful results. However,
studies have shown large uncertainties in the dosimetry associated
with PUVA treatment and point to equally large consequential
uncertainties about the actual treatment doses delivered (Diffey et
al 1980).
Hyperbilirubinaemia
Phototherapy is sometimes used in the treatment of neonatal
jaundice or hyperbilirubinaemia. The preferred method of treatment
is to irradiate the baby for several hours a day for up to one week
with visible light, particularly blue light. The lamps used for
phototherapy, although intended to emit only visible light, may also
have a UV component (Gies & Roy, 1990; Sliney & Wolbarsht, 1980).
Dentistry
Irradiation of the oral cavity with a Woods lamp can produce
fluorescence under certain conditions. This has been used in the
diagnosis of various dental disorders, such as early dental
cavities, the incorporation of tetracycline into bone and teeth,
dental plaque and calculus (Hefferren et al., 1971).
Pits and fissures in teeth have been treated using an adhesive
resin polymerized with UVA. The resin is applied with a fine brush
to the surfaces to be treated and hardened by exposure to UVA
radiation at a minimal irradiance of 100 W m-2 for about 30 s
(Eriksen, 1987; Diffey, 1990a).
4.5 Elective Exposure
By comparison to occupational exposure to UV, where control
measures are generally instituted to protect the eye and skin,
elective exposures can be much greater. Elective exposure results
from outdoor recreational activity, from intentional exposure to
sun-tanning equipment and to sunlight at the beach and elsewhere.
Actual dose estimates of elective exposure experienced by those who
attempt to maintain a tan may exceed 100 MED per year (Challoner et
al., 1976; Diffey, 1993a; Diffey et al., 1982).
5. DOSIMETRIC CONCEPTS
5.1 Interaction of UV with Matter
Attenuation of UV occurs due to absorption and scattering.
Reflection, refraction and diffraction are phenomena related to
boundaries between media. All of these interactions may change the
direction, intensity and the wavelength of UV. The quantities
describing interactions of UV with matter are dependent on the
wavelength.
The total energy absorbed in a material is influenced by
reflections from its surfaces. The attenuation coefficient describes
the attenuation of UV within tissue and is the sum of the absorption
and scattering coefficients. Penetration depth in a tissue is
inversely proportional to the exponential attenuation coefficient.
Absorption requires transfer of radiative energy to matter.
Apart from possible photomechanical effects, UV is absorbed as a
result of electron transitions at the atomic and molecular level.
Such molecular absorption can lead to photochemical reactions.
In most molecules the ground state or the singlet state (S0)
consists of two paired electrons. On absorption of radiant energy
one of the electrons can make a transition to an excited state
(S1), provided that the incident photon energy corresponds to an
existing level in the absorbing molecule. As shown in figure 5.1,
the molecule can release this absorbed energy from the excited state
by (a) transition directly back to the ground state, S0, or (b)
transition to the generally long-lived excited triplet state, T1,
and then discharging the remaining energy to return to S0. In
either case, transition back to S0 may be radiative (a photon
emitted) or non-radiative (energy dispersed by vibrational-
rotational relaxation). The radiative S1 -> S0 transition is
called "fluorescence" while radiative T1 ->S0 is called
"phosphorescence". In addition, molecules in the S1 or T1 states
can return to the ground state either by forming photoproducts or by
transferring the energy to an acceptor molecule. The effectiveness
of the photochemical process can be amplified by "photosensitizers"
or impeded by "quenchers".
4.3 Ocular Exposure Geometry
People seldom look directly at the sun when it is overhead and
very hazardous to view. It is not very hazardous to view when the
sun is low in the sky and falls within the normal field-of-view.
When the sun is more than 10° above the horizon, the natural
tendency is to partially close the eyelids or squint (called squint
reflex), thus shielding the retina from direct exposure. These
factors reduce the exposure to the cornea to a maximum of about 5%
of that falling upon the exposed top of head (Sliney, 1986). If the
squint and other behavioural factors are not considered, the dose to
the eyelid would be approximately 20% of the dose falling on a
horizontal surface.
Although the cornea is more sensitive to UV injury than the
skin, people seldom experiences a corneal burn when in sunlight.
Using the action spectrum for human photokeratitis and
mathematically weighting this with the midday solar spectrum, the
time to achieve the threshold for photokeratitis is about 100
seconds (Sliney, 1987, Rosenthal et al., 1988). Again, the
geometry of exposure precludes photokeratitis except when ground
reflectance exceeds approximately 10%. When the sun is overhead and
UV exposure is most severe, the brow ridge and upper lid shield the
cornea, and if the eye is turned away from the sun, the more intense
scattered UV from overhead strikes the cornea at a grazing angle of
incidence where most is reflected and little is absorbed (Sliney,
1983). Only when the incident UV rays are parallel to the pupillary
axis are most rays (approx. 98%) absorbed.
When looking at snow, UV is reflected directly into the eye;
hence, the traditional eye protector of the Inuit or Eskimo, the
slit, in whalebone or in a seal-skin mask, provided geometrical
rather than spectral protection against UV exposure (Hedblom, 1961).
The lack of protection above and to the sides of sunglasses is a
serious shortcoming. However, to obtain a quantitative idea of this
component of exposure to the eye, measurements were made using a
simulated ocular geometry in sunlight (Sliney, 1986). The human eye
received 10 to 25 % of the UV dose when wearing glasses with lenses
opaque to UV compared to no lens in the glasses. Therefore, unless
goggles with side-shields are used, UV transmission factors in
lenses much less than 2-5 % do not provide the eye protection
suggested by the transmission factor (Sliney, 1986).
The strong dependence of reflectance with angle of incidence is
termed Fresnel's Law of Reflection. This law not only explains the
survival of the cornea exposed overhead to UV, but also the glare
experienced over water. When the sun is overhead, water reflects
about 2% of the UV upward (Sliney, 1986). When low in the sky, much
of the sunlight is reflected while the UV and blue light are
filtered by the atmosphere. Nevertheless, the strong reflections
from water at these low sun angles create discomfort glare and UV
exposure of the cornea is further reduced because of the squint.
If dark lenses are placed over the eyes, the natural aversion
to bright light, which leads to the squint reflex (that greatly
lowers retinal UV or exposure to the eye), would be disabled. This
may appear to be an unusual way to consider the comfort that shaded
lenses provide. However, poor sunglasses may actually lead to a
higher UV exposure (Sliney, 1983).
However, quantifying the protective value of the upper and
lower eye lids when they close to squint is difficult. In terms of
UV exposure, at least a twenty-fold reduction is likely. For
shielding the retina from the direct image of the sun, the upper lid
probably provides a protection factor exceeding a thousand (Sliney -
personal communication). If a brimmed hat is worn, the direct image
of the sun on the retina is rare and overhead UV exposure is
virtually eliminated. However, while using a hat the lid opens
further and ground reflection of UV could become important.
On an overcast day, the lids open wider, and although the UVB
irradiance is reduced by cloud cover, the actual UVB dose rate to
the eye from atmospheric scattering near the horizon may be reduced
by a factor of only two (Sliney, 1983). Hence, on a cloudy day the
eye may receive a greater UVB dose than on a bright sunny day.
However, a heavy overcast may attenuate the UVB sufficiently, that
this observation may not be true. As sunglasses are not typically
worn on an overcast day, one could argue that the concern over
sunglasses increasing total ocular exposure is unimportant. However,
sunglasses should have sufficient UV filtration so that ocular
exposure does not actually increase when they are worn on a sunny
day.
Eye and head movements can further reduce UV exposure. Most
humans in bright sunlight squint or avoid looking into the sun
sector of the sky. These behavioural and physiological factors are
not taken into account by simple UV measurements. There is an
obvious need to determine accurately the corneal and lenticular
exposure to ambient UV. Indeed, the results of previous
epidemiological studies of cataract may be questioned because of
inadequate dosimetry. Some epidemiological studies may have reached
incorrect conclusions regarding risk from UV or sunlight exposure by
assigning inaccurate exposure levels to population groups, because
they assumed that overhead UV exposure accurately predicts corneal
exposure.
4.4 Workplace
4.4.1 Outdoor work
Although humans have adapted and acclimatized to solar UV it
nevertheless represents the most hazardous source of optical
radiation likely to be encountered by the average person and with a
few exceptions by the worker. People who work outdoors will be
subjected to involuntary UV exposure. The highest exposure will
occur during the two hours period either side of noon. Workers must
be made aware of this and take appropriate precautions as discussed
in chapter 13.
In a study of various work and recreational situations
(Challoner et al., 1976; Diffey et al., 1982), it was found that
outdoor workers had the highest exposure, receiving approximately
10% of the ambient level, similar to that received during sailing
and sightseeing. Higher exposures were found for skiing (20%) and
the largest during sunbathing on a beach (80%). Office workers
received about 3% of the total ambient radiation with about half
that figure from weekend exposure (Leach et al., 1978). The actual
exposures vary depending on the time of day and year, duration and
frequency of exposure.
4.4.2 Indoor work
Examples of indoor workplace exposures to UV are given below.
Photoprocesses
Many industrial processes involve a photochemical component.
The large-scale nature of these processes often necessitates the use
of high-power (several kilowatts) lamps such as high pressure metal
halide lamps which emit significant amounts of UV (Diffey, 1990a).
The principal industrial applications of photopolymerisation include
the curing of protective coatings and inks and photoresists for
printed circuit boards. The curing of printing inks by exposure to
UV takes only a fraction of a second. UV drying units can be
installed between printing stations on a multicolour line, so that
each colour is dried before the next is applied. Another major use
of UV curing has been for metal decorating in the packaging industry
(Phillips, 1983). UVA is also used to inspect printed circuit boards
and integrated circuits in the electronics industry.
Sterilization
UV with wavelengths in the range 260-265 nm is the most
effective for sterilization and disinfection since it corresponds to
a maximum in the DNA absorption spectrum. Low-pressure mercury
discharge tubes are often used as the UV source as more than 90% of
the radiated energy lies in the 254 nm line, figure 3.2. These lamps
are often referred to as "germicidal lamps", "bactericidal lamps" or
simply "UVC lamps" (Diffey, 1990a).
UVC radiation has been used to disinfect sewage effluent,
drinking-water, water for the cosmetics industry and swimming pools.
Germicidal lamps are sometimes used inside microbiological safety
cabinets to inactivate airborne and surface microorganisms (Diffey,
1990a). The combination of UV and ozone has a very powerful
oxidizing action and can reduce the organic content of water to
extremely low levels (Phillips, 1983).
Welding
Welding equipment falls into two broad categories: gas welding
and electric arc welding. Only the latter process produces
significant levels of UV, the quality and quantity of which depend
primarily on the arc current, shielding gas and metals being welded
(Sliney & Wolbarsht, 1980). The levels of UV irradiance around
electric arc welding equipment are high; effective irradiance at 1 m
at an arc current of 400 A ranged from 1 to 50 W m-2 and the
unweighted UVA irradiance ranged from 3 to 70 W m-2, depending on
the type of welding and the metal being welded (Mariutti & Matzeu,
1987; Cox, 1987). It is not surprising therefore that most welders
at some time or another experience "arc eye" or "welder's flash"
(photokeratitis) and skin erythema. The effective irradiance at 0.3
m from many types of electric welding arcs operating at 150 A is
such that the maximum permissible exposure time for an 8-h working
period on unprotected eyes and skin varies from a few tenths of a
second to about 10 s, depending on the type of welding process and
the material used (Cox, 1987).
4.4.3 Research
Sources of UV are used by most experimental scientists engaged
in photobiology and photochemistry and in molecular biology. These
applications, in which the effect of UV irradiation on biological
and chemical species is of primary interest to the researcher, can
be differentiated from UV fluorescence by absorption techniques
where the effect is of secondary importance (Diffey, 1990a).
4.4.4 Commerce
Sunlamps emitting UV have been used for tanning, particularly
in northern Europe and North America. Prior to the mid-1970s, the
source of UV in sunlamps was usually an unfiltered mercury arc lamp
which emitted a broad spectrum of radiation, including large
quantities of UVB and UVC. There are four distinctly different types
of ultraviolet tanning lamps in use, each with different UV emission
characteristics viz.; low pressure UVA fluorescent lamps; filtered
high intensity discharge UVA lamps; low pressure UVB fluorescent
lamps and; filtered high intensity discharge UVB lamps (IRPA/INIRC
1991). The emission characteristics and the health risks associated
with each type of lamp are different (Diffey and McKinlay 1983, Gies
et al 1986, Diffey 1987). Tanning systems incorporating lamps that
emit predominantly UVB are now little used and have been almost
universally replaced by the low pressure UVA fluorescent and
filtered high intensity discharge UVA types. Tanning requires
deliberate exposure of the skin to UV; however, the eyes must be
protected. Similarly, staff working in tanning salons must ensure
their exposure is kept to a minimum.
Many contaminants of food products can be detected by UV
fluorescence techniques. For example, the bacterium Pseudomonas
aeruginosa, which causes rot in eggs, meat and fish, can be detected
by its yellow-green fluorescence under UVA irradiation. One of the
longest established uses of UVA fluorescence in public health is to
demonstrate contamination with rodent urine, which is highly
fluorescent.
Many flying insects are attracted by UVA radiation,
particularly in the region around 350 nm. This phenomenon is the
principle of electronic insect traps, in which a UVA fluorescent
lamp is mounted in a unit containing a high-voltage grid. The
insect, attracted by the UVA lamp, flies into the unit and is
electrocuted in the air gap between the high-voltage grid and a
grounded metal screen. Such units are commonly found in areas where
food is prepared and sold to the public (Diffey, 1990a).
UVA blacklight lamps are sometimes used in discotheques to
induce fluorescence in the skin and clothing of dancers. The levels
of UVA emitted are usually low (< 10 W m-2) (Diffey, 1990a).
Signatures, banknotes and other documents can be authenticated
by exposing them to UVA, under which they fluoresce. UVA exposure of
the user is normally to hands and irradiance is low (< 10 W m-2)
(Diffey, 1990a).
4.4.5 Medicine and dentistry
In both medical and dental applications of UV, the patient is
deliberately exposed to either treat or diagnose a disease or
disorder. While non-target areas of the patient must be covered,
care must also be taken to ensure staff are fully protected from UV
exposure (see chapter 13).
The diagnostic uses of UV are confined largely to fluorescing
of the skin and teeth. UV exposure is limited to small areas (< 15
cm in diameter) and the UVA radiation dose per examination is
probably no more than 5 x 104J m-2. Diagnostic techniques are
limited to the use of fluorescence to identify the presence of
various fungal and bacterial infectious agents on the skin or in
wounds. The source of UV most commonly used for this purpose is the
Woods lamp which emits predominantly UVA radiation (365 nm).
Exposure of the patient's skin to potentially
carcinogenically-effective UV is insignificant (Diffey, 1990a).
UV phototherapy is a well established method for the treatment
of a number of skin conditions and in particular for psoriasis
(Green et al., 1992). Two general types of UVB emitting lamps are
used in phototherapy, high intensity discharge (HID) mercury vapour
and mercury/metal halide vapour lamps and low pressure mercury
vapour fluorescent lamps. The spectral emissions of these different
types of lamps vary greatly and this, with other factors such as the
number and power of the lamps, the lamp to skin distance and the
individual sensitivity of the patient, determines the treatment
time. A treatment dose is chosen to cause erythema in a particular
patient. As treatment progresses and the skin becomes increasingly
acclimatised, individual treatment doses are increased accordingly.
Treatments may be given several times per week. Phototherapy has
also been used for the treatment of other skin conditions including
severe itching, acne, eczema, polymorphic light eruption (PLE),
pityriasis rosea and urticaria, and for renal failure (Green et
al., 1992).
Seasonal affective disorder (SAD) is frequently treated by
exposure to sources of high illumination. Most treatment regimes
have employed sources incorporating so-called "full spectrum
lighting" whose emissions contain small amounts of UVA and UVB
radiations (Terman et al., 1990).
PUVA
UV photochemotherapy is UV phototherapy used in conjunction
with the oral or topical application of a chemical
(photosensitising) agent to the patient. The most widely used
treatment is PUVA which involves the use of the photosensitising
agent 8-methoxypsoralen (8-MOP) in conjunction with UVA irradiation
for the treatment of psoriasis. The irradiation sources used for
such treatment incorporate either low pressure mercury vapour UVA
fluorescent lamps, or HID mercury vapour or mercury/metal halide
vapour lamps with an added filter to effectively attenuate the UVB.
The UVA irradiance on the skin from sources incorporating
fluorescent lamps is generally of the order of 60 W m-2 and for
the HID mercury systems, around 250 W m-2. Treatments consist of a
starting dose of between 500 and 40,000 J m-2, depending on skin
type, followed by further incremental doses two to three times
weekly. Total treatment radiant exposures of between 106 and 2.5
106 J m-2 have been reported for successful results. However,
studies have shown large uncertainties in the dosimetry associated
with PUVA treatment and point to equally large consequential
uncertainties about the actual treatment doses delivered (Diffey et
al 1980).
Hyperbilirubinaemia
Phototherapy is sometimes used in the treatment of neonatal
jaundice or hyperbilirubinaemia. The preferred method of treatment
is to irradiate the baby for several hours a day for up to one week
with visible light, particularly blue light. The lamps used for
phototherapy, although intended to emit only visible light, may also
have a UV component (Gies & Roy, 1990; Sliney & Wolbarsht, 1980).
Dentistry
Irradiation of the oral cavity with a Woods lamp can produce
fluorescence under certain conditions. This has been used in the
diagnosis of various dental disorders, such as early dental
cavities, the incorporation of tetracycline into bone and teeth,
dental plaque and calculus (Hefferren et al., 1971).
Pits and fissures in teeth have been treated using an adhesive
resin polymerized with UVA. The resin is applied with a fine brush
to the surfaces to be treated and hardened by exposure to UVA
radiation at a minimal irradiance of 100 W m-2 for about 30 s
(Eriksen, 1987; Diffey, 1990a).
4.5 Elective Exposure
By comparison to occupational exposure to UV, where control
measures are generally instituted to protect the eye and skin,
elective exposures can be much greater. Elective exposure results
from outdoor recreational activity, from intentional exposure to
sun-tanning equipment and to sunlight at the beach and elsewhere.
Actual dose estimates of elective exposure experienced by those who
attempt to maintain a tan may exceed 100 MED per year (Challoner et
al., 1976; Diffey, 1993a; Diffey et al., 1982).
5. DOSIMETRIC CONCEPTS
5.1 Interaction of UV with Matter
Attenuation of UV occurs due to absorption and scattering.
Reflection, refraction and diffraction are phenomena related to
boundaries between media. All of these interactions may change the
direction, intensity and the wavelength of UV. The quantities
describing interactions of UV with matter are dependent on the
wavelength.
The total energy absorbed in a material is influenced by
reflections from its surfaces. The attenuation coefficient describes
the attenuation of UV within tissue and is the sum of the absorption
and scattering coefficients. Penetration depth in a tissue is
inversely proportional to the exponential attenuation coefficient.
Absorption requires transfer of radiative energy to matter.
Apart from possible photomechanical effects, UV is absorbed as a
result of electron transitions at the atomic and molecular level.
Such molecular absorption can lead to photochemical reactions.
In most molecules the ground state or the singlet state (S0)
consists of two paired electrons. On absorption of radiant energy
one of the electrons can make a transition to an excited state
(S1), provided that the incident photon energy corresponds to an
existing level in the absorbing molecule. As shown in figure 5.1,
the molecule can release this absorbed energy from the excited state
by (a) transition directly back to the ground state, S0, or (b)
transition to the generally long-lived excited triplet state, T1,
and then discharging the remaining energy to return to S0. In
either case, transition back to S0 may be radiative (a photon
emitted) or non-radiative (energy dispersed by vibrational-
rotational relaxation). The radiative S1 -> S0 transition is
called "fluorescence" while radiative T1 ->S0 is called
"phosphorescence". In addition, molecules in the S1 or T1 states
can return to the ground state either by forming photoproducts or by
transferring the energy to an acceptor molecule. The effectiveness
of the photochemical process can be amplified by "photosensitizers"
or impeded by "quenchers".
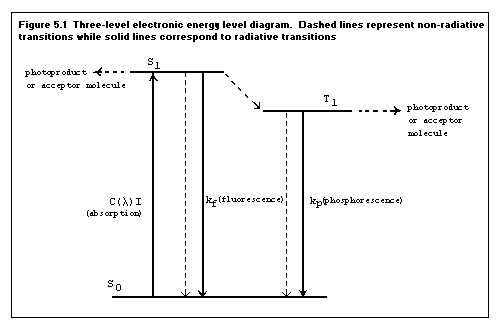 5.2 Biological Weighting Factors and Spectrally Weighted Quantities
Both radiant exposure H and irradiance E are quantities
integrated over the total spectrum of interest. To describe the
irradiance or radiant exposure in a very narrow spectral
(wavelength) interval (Delta lambda), the quantities of spectral
irradiance Elambda and spectral radiant exposure Hlambda are
employed. These have the units of: W m-2 nm and J m-2 nm,
respectively. If ElambdaDelta lambda is the irradiance in a narrow
interval Delta lambda around the wavelength lambda, then the
integrated irradiance E over the wavelength interval lamda1 to
lambda2 can be written as:
lambda2
E = Sigma Elambda Delta lambda
lambda1
Elambda and Hlambda are spectral functions. Other
spectroradiometric quantities exist with analogous definitions,
e.g., spectral radiant power, spectral radiant energy, etc, as shown
in table 2.2. The spectroradiometric quantities are important in
photobiology, and are critical in any discussion of the biological
effects of UV. The biological effects of UV are strongly wavelength
dependent. As a measure of these effects, "effective" or
"biologically active" quantities have been introduced (CIE, 1987).
The effective irradiance Eeff is defined as:
Eeff = Sigma Elambda Slambda Delta lambda
and similarly, the effective radiant exposure is
Heff = Sigma Hlambda Slambda Delta lambda
where Slambda is called the relative spectral effectiveness
function or action spectrum.
The action spectrum gives the relative biological response of
a tissue to irradiation at different wavelengths, and ideally will
correspond to the absorption spectrum of critical absorbing
molecules, or "chromophores." In biological systems, however, the
action spectrum function is modulated by the shielding (i.e.
thickness) of overlying tissue (e.g., stratum corneum of the skin),
energy transfer, and the action of sensitizers and quenchers. The
action spectrum is therefore specific for a certain effect arising
in a certain tissue layer. For example, the action spectrum for
erythema (skin) and photokeratitis (cornea) differ.
Based on a statistical analysis of the results of minimum
erythema dose studies carried out over the past 20 years or so, and
including data by Parrish et al. (1982) on the erythemal efficacy
of UVA, the CIE has promulgated a reference action curve (McKinlay
and Diffey, 1987), as shown in figure 5.2. This function consists of
three straight lines when plotted on a semi-logarithmic scale, and
although individual action spectra would not have the two inflection
points, the function can be readily expressed by three mathematical
functions. The function has been adopted internationally by the CIE
and the IEC and is being used by national organizations and
authorities for the determination of the erythemal potential of an
exposure to a given source of UV. A slightly different action
spectrum, which cannot be so readily expressed mathematically, has
been recommended by IRPA (1991) for risk assessment in occupational
health (see figure 5.2).
The erythemal response of the skin to UV is usually inferred
from the minimal erythema dose (MED). This value is determined by
exposing adjacent areas of skin to increasing doses of UV, and
recording the lowest dose to achieve erythema at a specified time,
usually 24 h after irradiation.
The visual detection of erythema is subjective and is affected
by unrelated factors such as viewing geometry, intensity and
spectral composition of ambient illumination, colour of unexposed
surrounding skin, (Chamberlain and Chamberlain 1980, Diffey and
Robson, 1992), and the experience and visual acuity of the observer.
The difficulty in judging accurately a minimal erythema response is
reflected by the varying definitions proposed for this value: these
range from the dose required to achieve a just perceptible erythema
(Everett et al., 1965, Kelfkens and van der Leun, 1989); to that
dose which will just produce a uniform redness with sharp borders
(Wucherpfenning, 1931).
The MED will vary according to the wavelength range over which
the effective UV is summed and for radiation protection purposes is
generally taken to lie in the range 200 to 300 J m-2 effective.
5.3 Measurement Techniques
There are three distinct types of measurement systems employed
in the detection of UV: radiometers, spectroradiometers and
dosimeters. Radiometers and spectroradiometers are direct-reading
instruments that use electro-optical (physical) detectors to convert
the incident radiation into an electrical signal. Radiometers
measure all incident radiant power over a wide spectral range;
whereas, spectroradiometers measure the radiant power distribution
over a wide spectral range. Either by electronic means or by
computer control, radiometers or spectroradiometers may be modified
and calibrated to operate as "dosimeters" by time-integration of the
output signal from the detector.
5.2 Biological Weighting Factors and Spectrally Weighted Quantities
Both radiant exposure H and irradiance E are quantities
integrated over the total spectrum of interest. To describe the
irradiance or radiant exposure in a very narrow spectral
(wavelength) interval (Delta lambda), the quantities of spectral
irradiance Elambda and spectral radiant exposure Hlambda are
employed. These have the units of: W m-2 nm and J m-2 nm,
respectively. If ElambdaDelta lambda is the irradiance in a narrow
interval Delta lambda around the wavelength lambda, then the
integrated irradiance E over the wavelength interval lamda1 to
lambda2 can be written as:
lambda2
E = Sigma Elambda Delta lambda
lambda1
Elambda and Hlambda are spectral functions. Other
spectroradiometric quantities exist with analogous definitions,
e.g., spectral radiant power, spectral radiant energy, etc, as shown
in table 2.2. The spectroradiometric quantities are important in
photobiology, and are critical in any discussion of the biological
effects of UV. The biological effects of UV are strongly wavelength
dependent. As a measure of these effects, "effective" or
"biologically active" quantities have been introduced (CIE, 1987).
The effective irradiance Eeff is defined as:
Eeff = Sigma Elambda Slambda Delta lambda
and similarly, the effective radiant exposure is
Heff = Sigma Hlambda Slambda Delta lambda
where Slambda is called the relative spectral effectiveness
function or action spectrum.
The action spectrum gives the relative biological response of
a tissue to irradiation at different wavelengths, and ideally will
correspond to the absorption spectrum of critical absorbing
molecules, or "chromophores." In biological systems, however, the
action spectrum function is modulated by the shielding (i.e.
thickness) of overlying tissue (e.g., stratum corneum of the skin),
energy transfer, and the action of sensitizers and quenchers. The
action spectrum is therefore specific for a certain effect arising
in a certain tissue layer. For example, the action spectrum for
erythema (skin) and photokeratitis (cornea) differ.
Based on a statistical analysis of the results of minimum
erythema dose studies carried out over the past 20 years or so, and
including data by Parrish et al. (1982) on the erythemal efficacy
of UVA, the CIE has promulgated a reference action curve (McKinlay
and Diffey, 1987), as shown in figure 5.2. This function consists of
three straight lines when plotted on a semi-logarithmic scale, and
although individual action spectra would not have the two inflection
points, the function can be readily expressed by three mathematical
functions. The function has been adopted internationally by the CIE
and the IEC and is being used by national organizations and
authorities for the determination of the erythemal potential of an
exposure to a given source of UV. A slightly different action
spectrum, which cannot be so readily expressed mathematically, has
been recommended by IRPA (1991) for risk assessment in occupational
health (see figure 5.2).
The erythemal response of the skin to UV is usually inferred
from the minimal erythema dose (MED). This value is determined by
exposing adjacent areas of skin to increasing doses of UV, and
recording the lowest dose to achieve erythema at a specified time,
usually 24 h after irradiation.
The visual detection of erythema is subjective and is affected
by unrelated factors such as viewing geometry, intensity and
spectral composition of ambient illumination, colour of unexposed
surrounding skin, (Chamberlain and Chamberlain 1980, Diffey and
Robson, 1992), and the experience and visual acuity of the observer.
The difficulty in judging accurately a minimal erythema response is
reflected by the varying definitions proposed for this value: these
range from the dose required to achieve a just perceptible erythema
(Everett et al., 1965, Kelfkens and van der Leun, 1989); to that
dose which will just produce a uniform redness with sharp borders
(Wucherpfenning, 1931).
The MED will vary according to the wavelength range over which
the effective UV is summed and for radiation protection purposes is
generally taken to lie in the range 200 to 300 J m-2 effective.
5.3 Measurement Techniques
There are three distinct types of measurement systems employed
in the detection of UV: radiometers, spectroradiometers and
dosimeters. Radiometers and spectroradiometers are direct-reading
instruments that use electro-optical (physical) detectors to convert
the incident radiation into an electrical signal. Radiometers
measure all incident radiant power over a wide spectral range;
whereas, spectroradiometers measure the radiant power distribution
over a wide spectral range. Either by electronic means or by
computer control, radiometers or spectroradiometers may be modified
and calibrated to operate as "dosimeters" by time-integration of the
output signal from the detector.
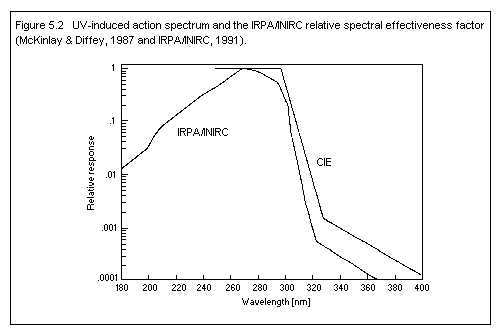 However, by dosimeters, one usually means devices that by
nature respond directly to incident dose, i.e. radiant exposure.
Dosimeters may be further optically modified and calibrated to
respond according to an action spectrum, thereby serving as a
direct-reading instrument for dose to a particular organ.
5.3.1 Detectors
The term detector normally refers to an electro-optical device
which converts an optical signal (e.g UV or light) into an
electronic signal which can be recorded. An important characteristic
of any detector is the responsivity, defined as the quotient of the
detector output (e.g amperes, A) and the radiant power incident upon
the detector (e.g. watts, W). Thus units for responsivity may vary.
The unit for irradiance-responsivity is often ampere-per-watt-per-
square-metre (A W-1 m-2). The spectral responsivity is the
responsivity as a function of wavelength. The term detectivity is
used to compare the detection capability (the smallest quantity of
radiation that can be detected) of different types of detectors, the
higher the detectivity the more sensitive the detector. Thermal
detectors, such as thermopiles and pyroelectric detectors, have a
much lower detectivity (less sensitive) than photodiodes, phototubes
and photomultipliers.
The most common types of detectors for UV are semiconductor
(junction) photodiodes, vacuum photodiodes (phototubes) and
photomultipliers. Junction photodiodes are usually silicon (Si)
photodiodes that may be enhanced to improve their UV responsivity
and with a spectral responsivity between 190 and 1100 nm.
Gallium-arsenide-phosphide (GaAsP) photodiodes have a spectral
responsivity between 190 and 670 nm, and gallium-phosphide (GaP)
photodiodes have a spectral responsivity between 190 and 520 nm.
Chemical detectors, such as photographic film emulsions or
polymer films of polysulphone or CR-39 resins, respond to incident
radiant exposure (J m-2). Their responsivity is generally strongly
wavelength dependant, and attempts are made to simulate a
photobiological action spectrum directly.
The choice of an optimum detector for a specific instrument or
measurement situation depends upon the requirements for ease of data
collection, portability, electrical power requirements, size and
accuracy. Each detector type has advantages and disadvantages.
Important parameters to consider for instrument requirements are:
spectral responsivity, noise-equivalent-power (NEP), linearity,
time/frequency response, stability over time, environmental
operating conditions, maintenance, ease of operation, the
requirements upon additional electronics, and cost.
Biological detectors are also used. For example, biofilms using
dried spores of bacillus subtilis, immobilized on transparent
polyester plastic sheets. After irradiation the biofilm is incubated
in a growth medium and the proteins synthesized after spore
germination, stained and evaluated by photometry. The biologically
effective dose is calculated using a calibration curve. The UV
response of this biofilm is additive and follows the reciprocity law
in the normal range of fluence rates investigated. The response is
independent of temperature (-20°C to 70°C) and humidity. The biofilm
can be stored for up to 9 months at room temperature without
significant influence on the viability of spores. These detectors
have been used in Antarctica and in space to measure the biological
consequences of ozone variations (Quintern et al., 1992).
5.3.2 Radiometers
A radiometer is a detection system that measures incident
radiation. A UV-radiometer usually measures irradiance in watts per
square metre (W m-2). The basic layout of a radiometer is shown in
figure 5.3.
The diffuser shown in figure 5.3 may not always be present.
When used, its purpose is to make the angular responsivity of the
radiometer proportional to the cosine of the angle of incidence
(measured from the normal to the diffuser surface). This arrangement
is often called a cosine-corrected radiometer. The diffuser may be
made from a flat or slightly curved piece of ground quartz or
teflon, or it may be an integrating sphere. The purpose of the
optical filter is to limit the spectral responsivity to a certain
band, having a lower and upper wavelength cutoff. The band-width can
vary upward from about 5 nm, but is often several tens or even
hundreds of nanometres wide. Hence, radiometers are called
broad-band meters as opposed to spectroradiometers. Ideally the
spectral responsivity is constant within the band and zero outside,
but in practice this is not possible. If the detector is sensitive
to radiation outside the passband of the filter, which is normally
the case, there will always be a non-zero responsivity outside the
band. Signals which are produced by radiation from outside the band
are called "out-of-band signals," "out-of-band leakage," or just
"stray light." This limitation of radiometers can often be a serious
problem, and is particularly troublesome in UV radiometers based on
Si photodiode detectors, because the spectral responsivities of Si
photodiodes extend to 1100 nm in the infrared. If one attempts to
use such radiometers to measure small quantities of in-band
radiation in the presence of large quantities of out-of-band
radiation, the results will be prone to large errors. This is
frequently encountered when attempting to measure the very small
component of biologically active UV present in a light source
spectrum.
However, by dosimeters, one usually means devices that by
nature respond directly to incident dose, i.e. radiant exposure.
Dosimeters may be further optically modified and calibrated to
respond according to an action spectrum, thereby serving as a
direct-reading instrument for dose to a particular organ.
5.3.1 Detectors
The term detector normally refers to an electro-optical device
which converts an optical signal (e.g UV or light) into an
electronic signal which can be recorded. An important characteristic
of any detector is the responsivity, defined as the quotient of the
detector output (e.g amperes, A) and the radiant power incident upon
the detector (e.g. watts, W). Thus units for responsivity may vary.
The unit for irradiance-responsivity is often ampere-per-watt-per-
square-metre (A W-1 m-2). The spectral responsivity is the
responsivity as a function of wavelength. The term detectivity is
used to compare the detection capability (the smallest quantity of
radiation that can be detected) of different types of detectors, the
higher the detectivity the more sensitive the detector. Thermal
detectors, such as thermopiles and pyroelectric detectors, have a
much lower detectivity (less sensitive) than photodiodes, phototubes
and photomultipliers.
The most common types of detectors for UV are semiconductor
(junction) photodiodes, vacuum photodiodes (phototubes) and
photomultipliers. Junction photodiodes are usually silicon (Si)
photodiodes that may be enhanced to improve their UV responsivity
and with a spectral responsivity between 190 and 1100 nm.
Gallium-arsenide-phosphide (GaAsP) photodiodes have a spectral
responsivity between 190 and 670 nm, and gallium-phosphide (GaP)
photodiodes have a spectral responsivity between 190 and 520 nm.
Chemical detectors, such as photographic film emulsions or
polymer films of polysulphone or CR-39 resins, respond to incident
radiant exposure (J m-2). Their responsivity is generally strongly
wavelength dependant, and attempts are made to simulate a
photobiological action spectrum directly.
The choice of an optimum detector for a specific instrument or
measurement situation depends upon the requirements for ease of data
collection, portability, electrical power requirements, size and
accuracy. Each detector type has advantages and disadvantages.
Important parameters to consider for instrument requirements are:
spectral responsivity, noise-equivalent-power (NEP), linearity,
time/frequency response, stability over time, environmental
operating conditions, maintenance, ease of operation, the
requirements upon additional electronics, and cost.
Biological detectors are also used. For example, biofilms using
dried spores of bacillus subtilis, immobilized on transparent
polyester plastic sheets. After irradiation the biofilm is incubated
in a growth medium and the proteins synthesized after spore
germination, stained and evaluated by photometry. The biologically
effective dose is calculated using a calibration curve. The UV
response of this biofilm is additive and follows the reciprocity law
in the normal range of fluence rates investigated. The response is
independent of temperature (-20°C to 70°C) and humidity. The biofilm
can be stored for up to 9 months at room temperature without
significant influence on the viability of spores. These detectors
have been used in Antarctica and in space to measure the biological
consequences of ozone variations (Quintern et al., 1992).
5.3.2 Radiometers
A radiometer is a detection system that measures incident
radiation. A UV-radiometer usually measures irradiance in watts per
square metre (W m-2). The basic layout of a radiometer is shown in
figure 5.3.
The diffuser shown in figure 5.3 may not always be present.
When used, its purpose is to make the angular responsivity of the
radiometer proportional to the cosine of the angle of incidence
(measured from the normal to the diffuser surface). This arrangement
is often called a cosine-corrected radiometer. The diffuser may be
made from a flat or slightly curved piece of ground quartz or
teflon, or it may be an integrating sphere. The purpose of the
optical filter is to limit the spectral responsivity to a certain
band, having a lower and upper wavelength cutoff. The band-width can
vary upward from about 5 nm, but is often several tens or even
hundreds of nanometres wide. Hence, radiometers are called
broad-band meters as opposed to spectroradiometers. Ideally the
spectral responsivity is constant within the band and zero outside,
but in practice this is not possible. If the detector is sensitive
to radiation outside the passband of the filter, which is normally
the case, there will always be a non-zero responsivity outside the
band. Signals which are produced by radiation from outside the band
are called "out-of-band signals," "out-of-band leakage," or just
"stray light." This limitation of radiometers can often be a serious
problem, and is particularly troublesome in UV radiometers based on
Si photodiode detectors, because the spectral responsivities of Si
photodiodes extend to 1100 nm in the infrared. If one attempts to
use such radiometers to measure small quantities of in-band
radiation in the presence of large quantities of out-of-band
radiation, the results will be prone to large errors. This is
frequently encountered when attempting to measure the very small
component of biologically active UV present in a light source
spectrum.
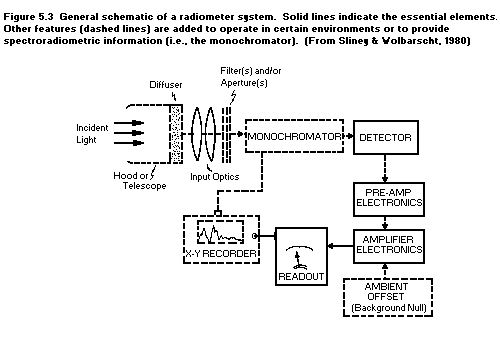 For many purposes it is desirable to have a UV radiometer which
has a spectral responsivity equal to or closely resembling a certain
action spectrum. If this is achieved, the radiometer signal is
directly proportional to the "effective" or "biologically active"
irradiance, because the radiometer will spectrally "weight" the
different wavelengths according to the action spectrum Slambda. A
well known example of such a radiometer is a photometer (lux-meter
or luminance-meter) which has a spectral responsiveness that closely
matches the photopic (visual) response of the human eye. Radiometers
are commercially available which have spectral responsivities that
match, for example, the UV erythema action spectrum (McKinlay and
Diffey, 1987) and UV hazard action spectrum adopted by the
IRPA/INIRC (1991) and ACGIH.
5.3.3 Spectroradiometers
A spectroradiometer is a radiometer that is capable of
measuring spectral radiometric quantities directly, such as spectral
irradiance or spectral radiance. The major difference between the
layout of a spectroradiometer and the layout of a radiometer is the
waveband selecting device which in a radiometer is usually a
broadband filter (see figure 5.3), whereas in a spectroradiometer it
is a monochromator or a spectrograph. Radiation entering a
monochromator or a spectrograph is dispersed by a grating or a prism
and only a small band of radiation is passed to the detector, the
so-called bandwidth. The spectroradiometer bandwidth can be selected
according to the application, but it is typically 1 - 5 nm. The
waveband passed to the detector can be changed manually or
automatically by rotating the grating or prism; the instrument is
scanned over the spectral range of interest. This type of instrument
is called a scanning spectroradiometer.
In a spectrograph, a portion of the spectrum is incident upon a
photographic film, or a linear photodiode array which can be read
diode by diode. This can occur very rapidly and a spectrum displayed
almost instantly. This type of spectroradiometer is therefore useful
for studies where time resolution is important. Another advantage is
that there are no moving parts compared to a scanning
spectroradiometer. However, spectrographs have disadvantages over
scanning systems, such as spectral resolution and detectivity.
Spectroradiometers are more complex to operate and maintain
than radiometers; they are considerably more expensive; and there
are many pitfalls for the inexperienced user. Spectroradiometers
generally employ the same kind of input optics as radiometers.
In an ideal spectroradiometer, the monochromator passes a small
band of wavelengths to the detector, and passes no radiation outside
this band. In practice the out-of-band radiation (leakage or
stray-light) that is passed to the detector is of the order of 0.1
percent of the amount of in-band radiation depending upon the
quality and size of the monochromator. In certain measurement
situations this may give rise to errors (just as in radiometers):
the signal caused by out-of-band radiation may be of the same order
of magnitude as the signal caused by in-band radiation. For example,
if one attempts to measure the solar UV spectrum in the 290-310 nm
region, this error occurs because the solar spectrum decreases about
five orders of magnitude from 310 nm to 290 nm. To overcome this
error, a double monochromator may be used. A double monochromator is
essentially two identical monochromators coupled in tandem, the
output of the first becomes the input of the second monochromator.
The stray light of a double monochromator is typically 0.01 percent.
5.3.4 Personal dosimetry
For personal monitoring of UV doses, radiometers are too bulky.
A thin-film polymer (eg polysulphone) dosimeter overcomes this
problem. It is a thin (0.04 mm) clear plastic film that may be worn
as a small badge comparable to those used to monitor ionizing
radiation. This allows monitoring of UVB doses on mobile subjects.
Polysulphone changes its absorbance (or transmittance) when
irradiated with UV (mainly by UVB). The received dose (radiant
exposure) is determined by measuring the change in absorbance of the
film before and after exposure (Diffey, 1989a). CR-39 plastic resins
have also been explored as UVB dosimeters; however, the low
sensitivity requires longer exposure periods (Wong et al., 1989).
5.4 Calibration
Improper or inadequate calibration of UV radiometers,
spectroradiometers and dosimeters is a serious and common source of
error. It is important to maintain a good calibration record for the
instruments, but only experience on instrument stability will
determine how often calibration is needed. The error caused by
calibration provides the minimum uncertainty that can be obtained in
measurement situations. Other sources of uncertainty include
geometry and spectrum of source emission, detector-source geometry
(angular errors), environmental influences and time factors.
Radiometers can be calibrated by using a source of known
irradiance. This may either be a line source, such as a mercury lamp
or a laser, or by a broadband source, such as a tungsten halogen
lamp. It must be realized that the irradiance-responsivity will
depend upon the source used for calibration. For example, a
radiometer which has been calibrated against a line source will give
erratic readings if used to measure a broadband source. In practice
it is advisable to have radiometers calibrated against a source
which emits a spectrum similar to those of the sources to be
measured.
Complete calibration of radiometers and spectral radiometers
include analysis of the cosine response of the instrument, the
azimuthal response and the temperature sensitivity of the
instrument. Recent work on instrument calibration and
intercomparison has shown that instruments can vary greatly in these
quantities even when the instruments agree on a simple spectral
calibration. In the field, instruments will measure different
quantities depending on the temperature and angle of incident
radiation.
Spectroradiometers are calibrated against standard lamps of
known spectral irradiance (or radiance). Such lamps can be obtained
from the standards institutes in various countries. Intercomparison
of lamp calibration from institute to institute can vary by as much
as eight percent in the UV region, while better agreement is common
in the visible region of standard lamps. This discrepancy should be
noted when comparing results based on instruments calibrated from
different lamp standards.
A tungsten halogen lamp is used as the standard lamp for
wavelengths between 250 and 2500 nm, whereas a deuterium lamp is
used in the region between 180 and 300-400 nm. It is good practice
to operate the standard lamp only when calibrating a
working-standard lamp and then use the working-standard for routine
calibration of a spectroradiometer. For very accurate work it is
recommended to maintain three calibrated lamps in order to find out
whether a change in response/calibration, at least of a
spectroradiometer, is caused by changes in the standard lamp used
for calibration or by changes in the spectroradiometer.
Dosimeters are calibrated in the same way as radiometers except
that exposure time is an integral part of the calibration process.
To obtain a reliable calibration for radiometers and dosimeters, it
is advisable to calibrate them against a source which emits a
spectrum similar to the one that is to be measured.
6. CELLULAR AND MOLECULAR STUDIES
6.1 Introduction
This chapter provides an overview of the evidence for cellular
and molecular effects of UV exposure on biological systems. Since UV
exposure has been associated with skin and eye cancers in humans,
emphasis will be given to the process of carcinogenesis. It is known
that UV exposure results in photochemical modification of the
genetic material (DNA), but most of this damage is accurately and
efficiently repaired by the cell. However if the amount of damage is
too great some of the alterations to the DNA may remain as permanent
mutations. It has been proposed that if unrepaired damage occurs to
regulatory genes this may be involved in the process of
carcinogenesis. In this context mutations to and activation of genes
may be important.
Other responses likely to result from UV exposure of cells
include increased cellular proliferation, which could have a tumour
promoting effect on genetically altered cells, as well as changes in
components of the immune system present in the skin. There is
evidence to suggest that UV exposure could elicit an
immunosuppressive effect which may compromise the body's ability to
identify and destroy tumour cells of the skin.
This review includes summary descriptions of UV action and
repair of damage to biomolecules, particularly in DNA, cellular
chromophores and other target molecules, as well as damage to the
cell membrane and proteins. Consequences of damage to the cell, its
membranes and activation of genes are also reviewed.
6.2 Interactions with Biomolecules
UV must be absorbed to produce a chemical change. At solar UV
intensities normally encountered, the first step in a photochemical
reaction is the absorption of a single photon by a molecule and the
production of an excited state in which one electron of the
absorbing molecule is raised to a higher energy level. Such
radiative transition can only occur efficiently when the photon
energy of the radiation is close to the energy difference of the
atom in the initial and final state (energy level). The
photochemistry that may then occur will therefore depend upon the
molecular structure and the wavelength of UV as well as the specific
reaction conditions. The primary products generated by UV absorption
are generally reactive species in a metastable excited state or free
radicals both of which form extremely fast. Dark chemical reactions
then occur often within microseconds but they may last for hours, as
is the case for the lipid peroxidation chain reaction. Finally these
relatively rapid processes are translated into photobiological
responses which may occur within seconds but can take years or even
decades to be manifested.
6.2.1 Cellular chromophores
Since chromophores (see chapter 5 for definition) are
characterized by the wavelengths at which they absorb, the nature of
the critical chromophores will change as a function of wavelength
throughout the UV range. The peak absorption of DNA is dictated by
its component nucleic acids and occurs at around 260 nm. There is a
sharp drop in absorption through the UVB range and absorption is
undetectable by conventional means at wavelengths longer than 320
nm. Using special detectors Sutherland and Griffin, (1981) have
measured DNA absorption at wavelengths as long as 360 nm. Although
overall protein absorption peaks in the UVC range, the aromatic
amino acids such as tryptophan (lambda max = 280 nm at pH7) and
tyrosine (lambda max = 275 nm at pH7) exhibit absorptions that
extend into the UVA range so that direct damage to proteins can
occur at much longer wavelengths than direct damage to DNA.
Several cellular components such as quinones, flavins, steroids
and porphyrins are important UVA chromophores. Porphyrins, which
exhibit an absorption with a peak around 405 nm, have been
implicated in the lethal action of UVA and near-visible light in
certain bacteria. Mutants in the bacterium Escherichia coli (E
coli) which are deficient in the synthesis of L-amino levulinic
acid, the first step in heme synthesis, are resistant to UVA
radiation (Tuveson and Sammartano, 1986) strongly suggesting that
porphyrin intermediates can be phototoxic. Porphyrin intermediates
evidently also arise during heme synthesis in humans. Indeed,
supplementation of human cells with amino levulinate (ALA) bypasses
the synthase step and leads to accumulation of protoporphyrin IX
(PPIX), the immediate precursor to heme and a strong
photosensitiser. ALA appears to be preferentially taken up by skin
cancer cells and the selective photosensitisation of such cells by
PPIX is the basis of a new type of phototherapy based on endogenous
sensitisers (Kennedy et al., 1990). Iron chelators can enhance the
sensitising effect by preventing the insertion of iron into the PPIX
macrocycle thus preventing the formation of the relatively
non-photoactive heme product. Accumulation of iron-free porphyrins
is the basis of the acute photosensitivity of skin in patients with
a variety of porphyrins. However, it is not clear to what extent
PPIX, for example, leads to UVA-mediated cytoxicity in normal human
skin cells.
Aside from DNA and proteins, the main chromophores in human
skin are urocanic acid and melanins (for review see Anderson &
Parrish, 1981). Trans-urocanic acid (4-amidazoleacrylic acid) is the
deamination product of histidine generated by histidase and because
of its broad absorption in the UV region has been considered as
contributing to a modest extent to the natural sunscreen properties
of skin. However, most notable is that the trans form can be
photoisomerised to the cis form and it has been proposed that this
conversion is a key factor in UV-induced immunosuppression (DeFabo
and Noonan, 1983). However, considerable controversy now surrounds
this original suggestion (eg. see Gibbs 1993).
Melanins are the major UV absorbing chromophores in skin,
exhibiting an extremely broad spectrum of absorption over the UVB,
UVA and visible ranges. Melanins are complex polymeric proteins that
are produced by melanocytes and transferred to keratinocytes.
Although often considered to be neutral density filters, this is not
strictly correct since melanins usually degrade upon UV exposure.
There is some evidence that melanin may function as a
photosensitizer of DNA damage.
In addition to iron-free porphyrins which generate singlet
oxygen upon exposure to UVA radiation, other small molecules also
have the potential to generate active oxygen intermediates upon UVA
exposure (Tyrell, 1992). For example, the photochemical degradation
of tryptophan by wavelengths which include the more energetic
portion of the UVA spectrum is able to generate hydrogen peroxide
and N-formyl kynurenin (McCormick et al., 1976). Although the
level of hydrogen peroxide generated in vivo by such a pathway
would appear to be in the low micromolar range it could nevertheless
be crucial to biological processes since iron complexes (such as
citrate) that are present in the cytoplasm will react with hydrogen
peroxide to generate the highly reactive hydroxyl radical in a
superoxide driven Fenton reaction (see Gutteridge, 1985; Imlay et
al., 1988). Since the reaction is driven by the continual
reduction of ferric iron to the ferrous state by superoxide anions,
a cellular source of superoxide anions is also required. In this
context it should be noted that both hydrogen peroxide and hydroxyl
radical are generated by UVA irradiation of NADH and NADPH
(Czochralska et al., 1984; Cunningham et al., 1985). However, it
is not at all clear whether this is really the key source of
superoxide anions or whether the main source is as a consequence of
normal cellular metabolism.
6.2.2 Cellular targets
The chromophores for UV effects are not necessarily the
critical targets which mediate the effects. The most important
cellular target for UV is considered to be DNA since the crucial
genetic material exists in unique and very low copy numbers in
cells. Radiation in the UVB range is absorbed by DNA and leads to
photochemical damage, so that DNA would certainly appear to be the
primary chromophore and site of damage for most of the biological
effects of short wavelength UV. DNA damage induced by UVB radiation
is the key factor leading to sunlight-induced mutations in
cancer-related genes and therefore in initiating the carcinogenic
process. At longer wavelengths, targets may change. For example, the
destruction of mitochondria may be a key factor in the breakdown of
cellular integrity following certain types of photosensitisation.
Membrane damage clearly takes on added significance when the UV
radiation employed (e.g.from sunlight) includes a strong component
of longer wavelengths. Breakdown of membranes can lead to aberrant
signal transduction as well as dramatic alterations in transport
pathways. Leakage of essential components or an influx of
extracellular molecules such as calcium can have severe cytostatic
and even lethal consequences for the cell and will clearly have an
influence on overall tissue/organ function.
6.3 Action Spectra
An action spectrum is a measure of the relative effectiveness
of different wavelengths, within the spectral region of study, to
produce a given response (see chapter 5 for definition). Clearly the
number of photochemical and photobiological endpoints that can be
measured is as large as the number of effects themselves and before
undertaking action spectroscopy for a given end-point, there should
exist a good a priori reason for the study. Numerous types of UV
induced DNA damage have now been recognized that include stand
breaks (single and double), cyclobutane-type pyrimidine dimers, 6-4
pyo photoproducts and the corresponding Dewar isomer, thymine
glycols, 8-hydroxy guanine, and many more. In addition, DNA-protein
cross-links are produced during UV exposure. Larger scale genetic
alterations include chromosome breakage, sister chromatid exchanges
and chromatid aberrations. Although partial UV action spectra are
now available for many of these lesions, the most studied have been
the different types of pyrimidine dimers. Since the indirect
oxidising component of radiation damage increases with increasing
wavelength, there is a dramatic shift in the type of lesion induced
as the wavelength increases. Pyrimidine dimers are characteristic of
the direct absorption that occurs at shorter UV wavelengths whereas
strand breaks and 8-hydroxy guanine type lesions become increasingly
important at longer wavelengths. At a higher level of complexity,
action spectra for cell death, mutation, in vitro transformation,
growth delays, cell permeability, etc, may also be measured.
One goal of determining action spectra has been to correlate
end-points with a specific type of initial damage. However, such
evaluations are complex partly because the absorbing chromophores
and crucial lesions will often change as a function of wavelength.
At the whole organism level, action spectra may be determined for
effects on entire organs, for example, various markers of leaf
damage in plants, erythema induction in skin and even tumour
induction.
Studies in cultured cells may be of value in predicting
responses in whole organs such as skin but various parameters must
be evaluated. In particular, the penetration of UV to the critical
chromophores as a function of wavelength must be taken into account
by considering the transmission through overlying tissue. As an
example, we may wish to calculate the relative cytotoxic action of
the different wavelengths in sunlight to cells at the basal layer of
the epidermis. In order to do this, an action spectrum must be
available for the cytotoxic action of individual wavelengths on
cultured epidermal keratinocytes. In addition, data is needed for
transmission through human skin to the target cells since this will
change as a function of wavelength. Finally if the effect of a
particular UV source is required (such as terrestrial sunlight under
defined conditions) then this must be determined by
spectroradiometry or from available data. The relative biological
effectiveness of individual wavelengths of sunlight in killing cells
at the basal level of the epidermis may then be predicted by
convoluting these three spectra. In practice, interactions
(synergistic, additive or antagonistic) exist between different UV
wavelength regions and these must also be taken into account in the
evaluation of the biological effects of broad spectrum sources.
Although action spectra may be of value for predictive evaluations,
the primary aim of studies with cultured cells is often to determine
chromophores. For this purpose, action spectra must be corrected in
order to express results as action per photon before comparison with
absorption spectra of critical biological molecules. Thus, DNA is
known to be an important chromophore in the UVB region. The results
are far more difficult to interpret in the longer wavelength UVA
region but at least for simpler organisms such as bacteria,
porphyrins have often been implicated. Action spectra, particularly
in plants, are often modified by protective absorbing molecules such
as carotenoids.
In response to questions posed by the threat to the ozone
layer, fairly detailed action spectra have now been determined for
squamous cell carcinoma in hairless mice and transmission
corrections made to estimate the spectrum in humans. DNA is clearly
the primary chromophore in the UVB region but a significant second
peak occurs in the long UVA wavelength region.
6.4 Biomolecular Damage
UV radiation can damage many cellular targets including the
nucleic acids, proteins and lipids. For the non-solar UVC
wavelengths, DNA is clearly the most important target and many
photochemical changes can occur as a result of direct absorption.
The genotoxic action of solar UVB radiation is also of critical
importance, although the spectrum of DNA damage begins to change as
oxidative events become more important. At longer UVA wavelengths,
indirect effects mediated by active oxygen intermediates are common
and except for events directly related to DNA modification (e.g.
mutation), it is difficult to discern the crucial targets.
6.4.1 Nucleic acids
Cyclobutane type pyrimidine dimers
Cyclobutane type pyrimidine dimers were the first type of
UV-induced base damage to be identified (Beukers & Berends, 1960)
and being the most frequent lesion induced by either UVC or UVB
radiation, they have been the most studied.
Cyclobutane-type pyrimidine dimer formation arises from the
production of reactive excited states (normally the forbidden but
long-lived triplet state) following absorption of UV radiation. The
action spectrum for dimer formation as shown in figure 6.1, closely
resembles that for the extinction coefficients of the appropriate
monomers, cytosine(C) or thymine(T) for wavelengths as long as 313
nm (Ellison & Childs, 1981) so that the mechanism of formation is
probably similar. Although pyrimidine dimer formation (e.g. thymine
lozenge thymine or T lozenge T) has been measured in the UVA range,
six orders of magnitude more energy is required at 365 nm as
compared to 254 nm (Tyrrell, 1973) and the mechanism of formation is
unclear. Most of the original observations concerning dimer
induction were made in isolated DNA, bacteriophage systems or in E
coli but essential results have since been confirmed in mammalian
cells including human skin fibroblasts. Action spectra determined in
human and mouse skin are sharply attenuated at shorter wavelengths,
but are otherwise basically similar to those obtained in vitro.
The action spectra for cyclobutane pyrimidine dimer formation in
naked DNA, cell cultures and epidermal DNA is given in figures 6.2
and 6.3. Based on studies with E coli, the ratio of T lozenge T, C
lozenge T to C lozenge C changes appreciably with wavelength. For
example, the ratio of T lozenge T to C lozenge T dimers is 0.63:1 at
313 nm (Ellison & Childs, 1981) but increases to 6:1 at 365 nm
(Tyrrell, 1973).
Although repair of DNA damage is dealt with separately below,
it is worth noting at this point that an extremely specific
light-dependant repair process, photoreactivation, exists for repair
of pyrimidine dimers in situ. Repair proceeds via photolyase which
has been shown to be present almost ubiquitously throughout the
animal world from E coli to non-placental mammals. Recent evidence
suggests that photoreactivation may be less important in human cells
(Li et al., 1993). Nevertheless, the photoenzymatic splitting of
pyrimidine dimers has provided a powerful technique since it has
provided a way of linking a specific type of DNA damage (the
pyrimidine dimer) with defined biological effects (e.g. cell death,
tumour formation).
For many purposes it is desirable to have a UV radiometer which
has a spectral responsivity equal to or closely resembling a certain
action spectrum. If this is achieved, the radiometer signal is
directly proportional to the "effective" or "biologically active"
irradiance, because the radiometer will spectrally "weight" the
different wavelengths according to the action spectrum Slambda. A
well known example of such a radiometer is a photometer (lux-meter
or luminance-meter) which has a spectral responsiveness that closely
matches the photopic (visual) response of the human eye. Radiometers
are commercially available which have spectral responsivities that
match, for example, the UV erythema action spectrum (McKinlay and
Diffey, 1987) and UV hazard action spectrum adopted by the
IRPA/INIRC (1991) and ACGIH.
5.3.3 Spectroradiometers
A spectroradiometer is a radiometer that is capable of
measuring spectral radiometric quantities directly, such as spectral
irradiance or spectral radiance. The major difference between the
layout of a spectroradiometer and the layout of a radiometer is the
waveband selecting device which in a radiometer is usually a
broadband filter (see figure 5.3), whereas in a spectroradiometer it
is a monochromator or a spectrograph. Radiation entering a
monochromator or a spectrograph is dispersed by a grating or a prism
and only a small band of radiation is passed to the detector, the
so-called bandwidth. The spectroradiometer bandwidth can be selected
according to the application, but it is typically 1 - 5 nm. The
waveband passed to the detector can be changed manually or
automatically by rotating the grating or prism; the instrument is
scanned over the spectral range of interest. This type of instrument
is called a scanning spectroradiometer.
In a spectrograph, a portion of the spectrum is incident upon a
photographic film, or a linear photodiode array which can be read
diode by diode. This can occur very rapidly and a spectrum displayed
almost instantly. This type of spectroradiometer is therefore useful
for studies where time resolution is important. Another advantage is
that there are no moving parts compared to a scanning
spectroradiometer. However, spectrographs have disadvantages over
scanning systems, such as spectral resolution and detectivity.
Spectroradiometers are more complex to operate and maintain
than radiometers; they are considerably more expensive; and there
are many pitfalls for the inexperienced user. Spectroradiometers
generally employ the same kind of input optics as radiometers.
In an ideal spectroradiometer, the monochromator passes a small
band of wavelengths to the detector, and passes no radiation outside
this band. In practice the out-of-band radiation (leakage or
stray-light) that is passed to the detector is of the order of 0.1
percent of the amount of in-band radiation depending upon the
quality and size of the monochromator. In certain measurement
situations this may give rise to errors (just as in radiometers):
the signal caused by out-of-band radiation may be of the same order
of magnitude as the signal caused by in-band radiation. For example,
if one attempts to measure the solar UV spectrum in the 290-310 nm
region, this error occurs because the solar spectrum decreases about
five orders of magnitude from 310 nm to 290 nm. To overcome this
error, a double monochromator may be used. A double monochromator is
essentially two identical monochromators coupled in tandem, the
output of the first becomes the input of the second monochromator.
The stray light of a double monochromator is typically 0.01 percent.
5.3.4 Personal dosimetry
For personal monitoring of UV doses, radiometers are too bulky.
A thin-film polymer (eg polysulphone) dosimeter overcomes this
problem. It is a thin (0.04 mm) clear plastic film that may be worn
as a small badge comparable to those used to monitor ionizing
radiation. This allows monitoring of UVB doses on mobile subjects.
Polysulphone changes its absorbance (or transmittance) when
irradiated with UV (mainly by UVB). The received dose (radiant
exposure) is determined by measuring the change in absorbance of the
film before and after exposure (Diffey, 1989a). CR-39 plastic resins
have also been explored as UVB dosimeters; however, the low
sensitivity requires longer exposure periods (Wong et al., 1989).
5.4 Calibration
Improper or inadequate calibration of UV radiometers,
spectroradiometers and dosimeters is a serious and common source of
error. It is important to maintain a good calibration record for the
instruments, but only experience on instrument stability will
determine how often calibration is needed. The error caused by
calibration provides the minimum uncertainty that can be obtained in
measurement situations. Other sources of uncertainty include
geometry and spectrum of source emission, detector-source geometry
(angular errors), environmental influences and time factors.
Radiometers can be calibrated by using a source of known
irradiance. This may either be a line source, such as a mercury lamp
or a laser, or by a broadband source, such as a tungsten halogen
lamp. It must be realized that the irradiance-responsivity will
depend upon the source used for calibration. For example, a
radiometer which has been calibrated against a line source will give
erratic readings if used to measure a broadband source. In practice
it is advisable to have radiometers calibrated against a source
which emits a spectrum similar to those of the sources to be
measured.
Complete calibration of radiometers and spectral radiometers
include analysis of the cosine response of the instrument, the
azimuthal response and the temperature sensitivity of the
instrument. Recent work on instrument calibration and
intercomparison has shown that instruments can vary greatly in these
quantities even when the instruments agree on a simple spectral
calibration. In the field, instruments will measure different
quantities depending on the temperature and angle of incident
radiation.
Spectroradiometers are calibrated against standard lamps of
known spectral irradiance (or radiance). Such lamps can be obtained
from the standards institutes in various countries. Intercomparison
of lamp calibration from institute to institute can vary by as much
as eight percent in the UV region, while better agreement is common
in the visible region of standard lamps. This discrepancy should be
noted when comparing results based on instruments calibrated from
different lamp standards.
A tungsten halogen lamp is used as the standard lamp for
wavelengths between 250 and 2500 nm, whereas a deuterium lamp is
used in the region between 180 and 300-400 nm. It is good practice
to operate the standard lamp only when calibrating a
working-standard lamp and then use the working-standard for routine
calibration of a spectroradiometer. For very accurate work it is
recommended to maintain three calibrated lamps in order to find out
whether a change in response/calibration, at least of a
spectroradiometer, is caused by changes in the standard lamp used
for calibration or by changes in the spectroradiometer.
Dosimeters are calibrated in the same way as radiometers except
that exposure time is an integral part of the calibration process.
To obtain a reliable calibration for radiometers and dosimeters, it
is advisable to calibrate them against a source which emits a
spectrum similar to the one that is to be measured.
6. CELLULAR AND MOLECULAR STUDIES
6.1 Introduction
This chapter provides an overview of the evidence for cellular
and molecular effects of UV exposure on biological systems. Since UV
exposure has been associated with skin and eye cancers in humans,
emphasis will be given to the process of carcinogenesis. It is known
that UV exposure results in photochemical modification of the
genetic material (DNA), but most of this damage is accurately and
efficiently repaired by the cell. However if the amount of damage is
too great some of the alterations to the DNA may remain as permanent
mutations. It has been proposed that if unrepaired damage occurs to
regulatory genes this may be involved in the process of
carcinogenesis. In this context mutations to and activation of genes
may be important.
Other responses likely to result from UV exposure of cells
include increased cellular proliferation, which could have a tumour
promoting effect on genetically altered cells, as well as changes in
components of the immune system present in the skin. There is
evidence to suggest that UV exposure could elicit an
immunosuppressive effect which may compromise the body's ability to
identify and destroy tumour cells of the skin.
This review includes summary descriptions of UV action and
repair of damage to biomolecules, particularly in DNA, cellular
chromophores and other target molecules, as well as damage to the
cell membrane and proteins. Consequences of damage to the cell, its
membranes and activation of genes are also reviewed.
6.2 Interactions with Biomolecules
UV must be absorbed to produce a chemical change. At solar UV
intensities normally encountered, the first step in a photochemical
reaction is the absorption of a single photon by a molecule and the
production of an excited state in which one electron of the
absorbing molecule is raised to a higher energy level. Such
radiative transition can only occur efficiently when the photon
energy of the radiation is close to the energy difference of the
atom in the initial and final state (energy level). The
photochemistry that may then occur will therefore depend upon the
molecular structure and the wavelength of UV as well as the specific
reaction conditions. The primary products generated by UV absorption
are generally reactive species in a metastable excited state or free
radicals both of which form extremely fast. Dark chemical reactions
then occur often within microseconds but they may last for hours, as
is the case for the lipid peroxidation chain reaction. Finally these
relatively rapid processes are translated into photobiological
responses which may occur within seconds but can take years or even
decades to be manifested.
6.2.1 Cellular chromophores
Since chromophores (see chapter 5 for definition) are
characterized by the wavelengths at which they absorb, the nature of
the critical chromophores will change as a function of wavelength
throughout the UV range. The peak absorption of DNA is dictated by
its component nucleic acids and occurs at around 260 nm. There is a
sharp drop in absorption through the UVB range and absorption is
undetectable by conventional means at wavelengths longer than 320
nm. Using special detectors Sutherland and Griffin, (1981) have
measured DNA absorption at wavelengths as long as 360 nm. Although
overall protein absorption peaks in the UVC range, the aromatic
amino acids such as tryptophan (lambda max = 280 nm at pH7) and
tyrosine (lambda max = 275 nm at pH7) exhibit absorptions that
extend into the UVA range so that direct damage to proteins can
occur at much longer wavelengths than direct damage to DNA.
Several cellular components such as quinones, flavins, steroids
and porphyrins are important UVA chromophores. Porphyrins, which
exhibit an absorption with a peak around 405 nm, have been
implicated in the lethal action of UVA and near-visible light in
certain bacteria. Mutants in the bacterium Escherichia coli (E
coli) which are deficient in the synthesis of L-amino levulinic
acid, the first step in heme synthesis, are resistant to UVA
radiation (Tuveson and Sammartano, 1986) strongly suggesting that
porphyrin intermediates can be phototoxic. Porphyrin intermediates
evidently also arise during heme synthesis in humans. Indeed,
supplementation of human cells with amino levulinate (ALA) bypasses
the synthase step and leads to accumulation of protoporphyrin IX
(PPIX), the immediate precursor to heme and a strong
photosensitiser. ALA appears to be preferentially taken up by skin
cancer cells and the selective photosensitisation of such cells by
PPIX is the basis of a new type of phototherapy based on endogenous
sensitisers (Kennedy et al., 1990). Iron chelators can enhance the
sensitising effect by preventing the insertion of iron into the PPIX
macrocycle thus preventing the formation of the relatively
non-photoactive heme product. Accumulation of iron-free porphyrins
is the basis of the acute photosensitivity of skin in patients with
a variety of porphyrins. However, it is not clear to what extent
PPIX, for example, leads to UVA-mediated cytoxicity in normal human
skin cells.
Aside from DNA and proteins, the main chromophores in human
skin are urocanic acid and melanins (for review see Anderson &
Parrish, 1981). Trans-urocanic acid (4-amidazoleacrylic acid) is the
deamination product of histidine generated by histidase and because
of its broad absorption in the UV region has been considered as
contributing to a modest extent to the natural sunscreen properties
of skin. However, most notable is that the trans form can be
photoisomerised to the cis form and it has been proposed that this
conversion is a key factor in UV-induced immunosuppression (DeFabo
and Noonan, 1983). However, considerable controversy now surrounds
this original suggestion (eg. see Gibbs 1993).
Melanins are the major UV absorbing chromophores in skin,
exhibiting an extremely broad spectrum of absorption over the UVB,
UVA and visible ranges. Melanins are complex polymeric proteins that
are produced by melanocytes and transferred to keratinocytes.
Although often considered to be neutral density filters, this is not
strictly correct since melanins usually degrade upon UV exposure.
There is some evidence that melanin may function as a
photosensitizer of DNA damage.
In addition to iron-free porphyrins which generate singlet
oxygen upon exposure to UVA radiation, other small molecules also
have the potential to generate active oxygen intermediates upon UVA
exposure (Tyrell, 1992). For example, the photochemical degradation
of tryptophan by wavelengths which include the more energetic
portion of the UVA spectrum is able to generate hydrogen peroxide
and N-formyl kynurenin (McCormick et al., 1976). Although the
level of hydrogen peroxide generated in vivo by such a pathway
would appear to be in the low micromolar range it could nevertheless
be crucial to biological processes since iron complexes (such as
citrate) that are present in the cytoplasm will react with hydrogen
peroxide to generate the highly reactive hydroxyl radical in a
superoxide driven Fenton reaction (see Gutteridge, 1985; Imlay et
al., 1988). Since the reaction is driven by the continual
reduction of ferric iron to the ferrous state by superoxide anions,
a cellular source of superoxide anions is also required. In this
context it should be noted that both hydrogen peroxide and hydroxyl
radical are generated by UVA irradiation of NADH and NADPH
(Czochralska et al., 1984; Cunningham et al., 1985). However, it
is not at all clear whether this is really the key source of
superoxide anions or whether the main source is as a consequence of
normal cellular metabolism.
6.2.2 Cellular targets
The chromophores for UV effects are not necessarily the
critical targets which mediate the effects. The most important
cellular target for UV is considered to be DNA since the crucial
genetic material exists in unique and very low copy numbers in
cells. Radiation in the UVB range is absorbed by DNA and leads to
photochemical damage, so that DNA would certainly appear to be the
primary chromophore and site of damage for most of the biological
effects of short wavelength UV. DNA damage induced by UVB radiation
is the key factor leading to sunlight-induced mutations in
cancer-related genes and therefore in initiating the carcinogenic
process. At longer wavelengths, targets may change. For example, the
destruction of mitochondria may be a key factor in the breakdown of
cellular integrity following certain types of photosensitisation.
Membrane damage clearly takes on added significance when the UV
radiation employed (e.g.from sunlight) includes a strong component
of longer wavelengths. Breakdown of membranes can lead to aberrant
signal transduction as well as dramatic alterations in transport
pathways. Leakage of essential components or an influx of
extracellular molecules such as calcium can have severe cytostatic
and even lethal consequences for the cell and will clearly have an
influence on overall tissue/organ function.
6.3 Action Spectra
An action spectrum is a measure of the relative effectiveness
of different wavelengths, within the spectral region of study, to
produce a given response (see chapter 5 for definition). Clearly the
number of photochemical and photobiological endpoints that can be
measured is as large as the number of effects themselves and before
undertaking action spectroscopy for a given end-point, there should
exist a good a priori reason for the study. Numerous types of UV
induced DNA damage have now been recognized that include stand
breaks (single and double), cyclobutane-type pyrimidine dimers, 6-4
pyo photoproducts and the corresponding Dewar isomer, thymine
glycols, 8-hydroxy guanine, and many more. In addition, DNA-protein
cross-links are produced during UV exposure. Larger scale genetic
alterations include chromosome breakage, sister chromatid exchanges
and chromatid aberrations. Although partial UV action spectra are
now available for many of these lesions, the most studied have been
the different types of pyrimidine dimers. Since the indirect
oxidising component of radiation damage increases with increasing
wavelength, there is a dramatic shift in the type of lesion induced
as the wavelength increases. Pyrimidine dimers are characteristic of
the direct absorption that occurs at shorter UV wavelengths whereas
strand breaks and 8-hydroxy guanine type lesions become increasingly
important at longer wavelengths. At a higher level of complexity,
action spectra for cell death, mutation, in vitro transformation,
growth delays, cell permeability, etc, may also be measured.
One goal of determining action spectra has been to correlate
end-points with a specific type of initial damage. However, such
evaluations are complex partly because the absorbing chromophores
and crucial lesions will often change as a function of wavelength.
At the whole organism level, action spectra may be determined for
effects on entire organs, for example, various markers of leaf
damage in plants, erythema induction in skin and even tumour
induction.
Studies in cultured cells may be of value in predicting
responses in whole organs such as skin but various parameters must
be evaluated. In particular, the penetration of UV to the critical
chromophores as a function of wavelength must be taken into account
by considering the transmission through overlying tissue. As an
example, we may wish to calculate the relative cytotoxic action of
the different wavelengths in sunlight to cells at the basal layer of
the epidermis. In order to do this, an action spectrum must be
available for the cytotoxic action of individual wavelengths on
cultured epidermal keratinocytes. In addition, data is needed for
transmission through human skin to the target cells since this will
change as a function of wavelength. Finally if the effect of a
particular UV source is required (such as terrestrial sunlight under
defined conditions) then this must be determined by
spectroradiometry or from available data. The relative biological
effectiveness of individual wavelengths of sunlight in killing cells
at the basal level of the epidermis may then be predicted by
convoluting these three spectra. In practice, interactions
(synergistic, additive or antagonistic) exist between different UV
wavelength regions and these must also be taken into account in the
evaluation of the biological effects of broad spectrum sources.
Although action spectra may be of value for predictive evaluations,
the primary aim of studies with cultured cells is often to determine
chromophores. For this purpose, action spectra must be corrected in
order to express results as action per photon before comparison with
absorption spectra of critical biological molecules. Thus, DNA is
known to be an important chromophore in the UVB region. The results
are far more difficult to interpret in the longer wavelength UVA
region but at least for simpler organisms such as bacteria,
porphyrins have often been implicated. Action spectra, particularly
in plants, are often modified by protective absorbing molecules such
as carotenoids.
In response to questions posed by the threat to the ozone
layer, fairly detailed action spectra have now been determined for
squamous cell carcinoma in hairless mice and transmission
corrections made to estimate the spectrum in humans. DNA is clearly
the primary chromophore in the UVB region but a significant second
peak occurs in the long UVA wavelength region.
6.4 Biomolecular Damage
UV radiation can damage many cellular targets including the
nucleic acids, proteins and lipids. For the non-solar UVC
wavelengths, DNA is clearly the most important target and many
photochemical changes can occur as a result of direct absorption.
The genotoxic action of solar UVB radiation is also of critical
importance, although the spectrum of DNA damage begins to change as
oxidative events become more important. At longer UVA wavelengths,
indirect effects mediated by active oxygen intermediates are common
and except for events directly related to DNA modification (e.g.
mutation), it is difficult to discern the crucial targets.
6.4.1 Nucleic acids
Cyclobutane type pyrimidine dimers
Cyclobutane type pyrimidine dimers were the first type of
UV-induced base damage to be identified (Beukers & Berends, 1960)
and being the most frequent lesion induced by either UVC or UVB
radiation, they have been the most studied.
Cyclobutane-type pyrimidine dimer formation arises from the
production of reactive excited states (normally the forbidden but
long-lived triplet state) following absorption of UV radiation. The
action spectrum for dimer formation as shown in figure 6.1, closely
resembles that for the extinction coefficients of the appropriate
monomers, cytosine(C) or thymine(T) for wavelengths as long as 313
nm (Ellison & Childs, 1981) so that the mechanism of formation is
probably similar. Although pyrimidine dimer formation (e.g. thymine
lozenge thymine or T lozenge T) has been measured in the UVA range,
six orders of magnitude more energy is required at 365 nm as
compared to 254 nm (Tyrrell, 1973) and the mechanism of formation is
unclear. Most of the original observations concerning dimer
induction were made in isolated DNA, bacteriophage systems or in E
coli but essential results have since been confirmed in mammalian
cells including human skin fibroblasts. Action spectra determined in
human and mouse skin are sharply attenuated at shorter wavelengths,
but are otherwise basically similar to those obtained in vitro.
The action spectra for cyclobutane pyrimidine dimer formation in
naked DNA, cell cultures and epidermal DNA is given in figures 6.2
and 6.3. Based on studies with E coli, the ratio of T lozenge T, C
lozenge T to C lozenge C changes appreciably with wavelength. For
example, the ratio of T lozenge T to C lozenge T dimers is 0.63:1 at
313 nm (Ellison & Childs, 1981) but increases to 6:1 at 365 nm
(Tyrrell, 1973).
Although repair of DNA damage is dealt with separately below,
it is worth noting at this point that an extremely specific
light-dependant repair process, photoreactivation, exists for repair
of pyrimidine dimers in situ. Repair proceeds via photolyase which
has been shown to be present almost ubiquitously throughout the
animal world from E coli to non-placental mammals. Recent evidence
suggests that photoreactivation may be less important in human cells
(Li et al., 1993). Nevertheless, the photoenzymatic splitting of
pyrimidine dimers has provided a powerful technique since it has
provided a way of linking a specific type of DNA damage (the
pyrimidine dimer) with defined biological effects (e.g. cell death,
tumour formation).
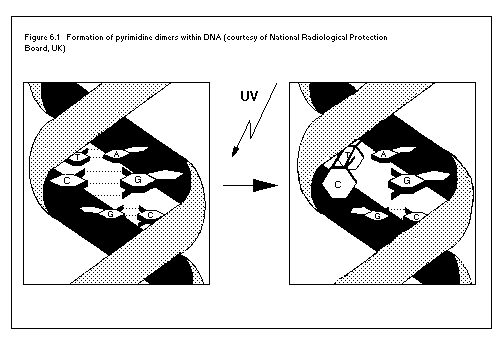
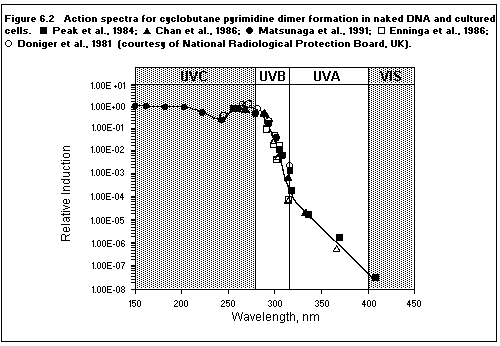
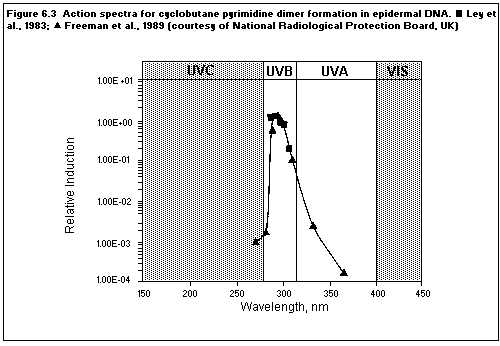 Thy (6-4) pyo photoproducts
A second type of pyrimidine dimer can be formed by UV (Varghese
& Wang, 1967) which was originally termed a pyrimidine adduct and is
now more commonly referred to as the thy (6-4) pyo photoproduct.
More recently a pyrimidine nucleoside-cytidine lesion was recognized
in highly reiterated sequences of human DNA (Lippke et al., 1981)
which is almost certainly the precursor of the thy (6-4) pyo
photoproduct (Brash & Haseltine, 1982; Franklin, et al., 1982).
The (6-4) photoproduct is formed with much greater frequency between
cytosines located 5' of adjacent pyrimidines. Action spectra that
compare induction of cyclobutane-type pyrimidine dimers are all
similar between 265 nm and 302 nm (Patrick, 1977; Chan et al.,
1986; Matsunaga, et al., 1991; Rosenstein & Mitchell, 1987) as
shown in figure 6.4. However, at longer wavelengths not all the
spectra agree. At least for human skin fibroblasts the relative
level of (6-4) photoproduct induction is only half that for
cyclobutane-type dimers. The reason for this appears to be that the
(6-4) photoproducts are converted to a Dewar pyrimidine isomer by UV
radiation at wavelengths peaking in the UVB range. This has been
confirmed in studies by Mitchell & Rosenstein (1987) using
radio-immuno assays for the (6-4) photoproduct and its Dewar isomer.
Although (6-4) photoproducts form with 5-10 fold lower
efficiency than cyclobutane type dimers, they may be formed with
equal efficiency at certain sites (Kraemer et al., 1988).
Furthermore, in the UVB region, a significant level of Dewar
pyrimidine isomers will be formed and the precise ratio and level of
these three major types of base damage will depend on the
wavelength(s) of radiation employed, as well as irradiation
conditions.
Other types of dimeric base damage which include purines have
been isolated from DNA heavily irradiated with UVC. These include
the thymine-adenine dimer (Bose & Davies, 1984) and adenine
dehydrodimer (Gasparro & Fresco, 1986). Such photoproducts occur at
fairly low frequency (approximately 1 percent of that of
pyrimidine-type dimers. The relative levels and significance of
these photoproducts in the UVB or longer wavelength regions has not
yet been determined.
Thy (6-4) pyo photoproducts
A second type of pyrimidine dimer can be formed by UV (Varghese
& Wang, 1967) which was originally termed a pyrimidine adduct and is
now more commonly referred to as the thy (6-4) pyo photoproduct.
More recently a pyrimidine nucleoside-cytidine lesion was recognized
in highly reiterated sequences of human DNA (Lippke et al., 1981)
which is almost certainly the precursor of the thy (6-4) pyo
photoproduct (Brash & Haseltine, 1982; Franklin, et al., 1982).
The (6-4) photoproduct is formed with much greater frequency between
cytosines located 5' of adjacent pyrimidines. Action spectra that
compare induction of cyclobutane-type pyrimidine dimers are all
similar between 265 nm and 302 nm (Patrick, 1977; Chan et al.,
1986; Matsunaga, et al., 1991; Rosenstein & Mitchell, 1987) as
shown in figure 6.4. However, at longer wavelengths not all the
spectra agree. At least for human skin fibroblasts the relative
level of (6-4) photoproduct induction is only half that for
cyclobutane-type dimers. The reason for this appears to be that the
(6-4) photoproducts are converted to a Dewar pyrimidine isomer by UV
radiation at wavelengths peaking in the UVB range. This has been
confirmed in studies by Mitchell & Rosenstein (1987) using
radio-immuno assays for the (6-4) photoproduct and its Dewar isomer.
Although (6-4) photoproducts form with 5-10 fold lower
efficiency than cyclobutane type dimers, they may be formed with
equal efficiency at certain sites (Kraemer et al., 1988).
Furthermore, in the UVB region, a significant level of Dewar
pyrimidine isomers will be formed and the precise ratio and level of
these three major types of base damage will depend on the
wavelength(s) of radiation employed, as well as irradiation
conditions.
Other types of dimeric base damage which include purines have
been isolated from DNA heavily irradiated with UVC. These include
the thymine-adenine dimer (Bose & Davies, 1984) and adenine
dehydrodimer (Gasparro & Fresco, 1986). Such photoproducts occur at
fairly low frequency (approximately 1 percent of that of
pyrimidine-type dimers. The relative levels and significance of
these photoproducts in the UVB or longer wavelength regions has not
yet been determined.
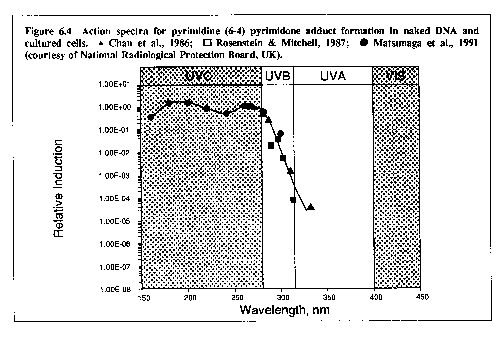 Monobasic DNA damage
Damage to a single base is a relatively low frequency event in
UV damaged DNA. Simple hydrates of cytosine and thymine can
certainly be formed (Fisher & Johns, 1976) but these low frequency
events are unreliable and therefore extremely difficult to study
from a biological viewpoint. They may be related to the low
frequency cytosine photoproduct identified by sequencing techniques
by Gallagher et al. (1989). Sequencing techniques in combination
with endonuclease V treatment have also led to the detection of a
class of rare and unidentified purine or purine-pyrimidine sites
after broad spectrum UV irradiation (Gallagher & Duker, 1986).
A group of lesions induced by both UV and ionizing radiation
are ring-saturated thymines of the 5, 6- dihyroxydihydrothymine type
(thymine glycols). Although original measurements indicated that
they were induced almost as frequently as pyrimidine dimers at 313
nm as against 21:1 at 254 nm (Cerutti & Netrawali, 1979), it has
been concluded from more recent data using sequencing analysis
(Mitchell et al., 1993) that they do not occur at a significant
rate in UVC or UVB irradiated DNA.
Increasingly sophisticated chemical methods are now becoming
available to measure oxidative DNA damage (Cadet et al., Methods
in Enzymology, in press). Since the proportion of this type of
damage will increase with increasing wavelength, such techniques
will soon be applied to consolidate the picture of UV-induced based
damage throughout the entire solar UV spectrum. Furthermore, an
interesting enzyme has been isolated (Boiteux et al., 1987) termed
the Fapy glycosylase because of its ability to excise the
ring-opened form of N7-methylguanine from DNA. The enzyme will also
recognize DNA products generated in photosensitisation reactions
involving singlet oxygen (Epe et al., in press) and
8-hydroxyguanine may be a major component. Since the longer UV
wavelength in sunlight clearly generate biologically relevant levels
of singlet oxygen and other active intermediates (Tyrrell, 1991;
Basu-Modak & Tyrrell, 1993), this type of analysis may also be
usefully applied to the solar UV spectrum. Certainly, near-visible
light alone is able to generate Fapy glycosylase-sensitive lesions
in the DNA of cultured mammalian cells and these, at least in part,
are probably 8-hydroxyguanine (Epe et al., 1993).
DNA strand breaks
Few DNA strand breaks are induced by UVC radiation but they
constitute an increasing proportion of the total lesions as
wavelength is increased. For example, in E. coli the ratio of DNA
strand breaks to pyrimidine dimers is 1:44 at 313 nm (Miguel &
Tyrrell, 1983) whereas at 365 nm one strand break is formed for
every two pyrimidine dimers (Tyrrell et al., 1974). Action spectra
have now been determined for the induction of DNA single strand
breaks in human skin cells which show that breaks occur throughout
the UVA and into the near-visible range (Peak et al., 1987). Since
break measurements involve alkaline denaturation, 10-20 percent of
the so-called breaks are due to the fragility of chemical bonds at
apurinic and apyrimidinic sites. It is important to note that unlike
most of the common forms of base damage, induction of strand breaks
is strongly dependent on oxygen throughout the UVB and UVA ranges
(Tyrrell, et al., 1974; Peak & Peak, 1982; Miguel and Tyrrell, 1983)
and at the longest wavelengths may involve generation of singlet
oxygen (Peak et al., 1987). Figure 6.5 gives the action spectra
for single strand breaks and figure 6.6 for double strand breaks in
cultured cells.
DNA-protein cross links
Photochemically induced DNA-protein cross links, mostly
involving cysteine are clearly formed in vitro (Smith, 1976;
Shetlar, 1980) and appear to be formed in vivo, particularly in
the UVA range (Bradley, et al., 1979; Peak & Peak, 1991). Despite
the obvious importance of this type of damage, there is little known
at the molecular level concerning the nature of the damage formed
in vivo. Action spectra for DNA-protein cross links in cultured
cells is given in figure 6.7 (from Peak et al., 1985a, b).
Ribonucleic acid
Messenger RNA is readily susceptible to modification by UV
radiation. However, given the fairly rapid turnover of most
molecules of this type (half-lives generally of the order of minutes
and hours) and the capacity for de novo synthesis in the absence
of DNA damage, mRNA is not generally considered a critical target of
radiation damage in mammalian cells. Bacterial transfer RNA is often
extremely photosensitive because of the presence of an unusual
nucleoside 4-thiouridine and this leads to some fascinating
photobiological phenomena of ecological significance (Jagger, 1985).
However, such findings are of little significance to eukaryotic
cells.
Monobasic DNA damage
Damage to a single base is a relatively low frequency event in
UV damaged DNA. Simple hydrates of cytosine and thymine can
certainly be formed (Fisher & Johns, 1976) but these low frequency
events are unreliable and therefore extremely difficult to study
from a biological viewpoint. They may be related to the low
frequency cytosine photoproduct identified by sequencing techniques
by Gallagher et al. (1989). Sequencing techniques in combination
with endonuclease V treatment have also led to the detection of a
class of rare and unidentified purine or purine-pyrimidine sites
after broad spectrum UV irradiation (Gallagher & Duker, 1986).
A group of lesions induced by both UV and ionizing radiation
are ring-saturated thymines of the 5, 6- dihyroxydihydrothymine type
(thymine glycols). Although original measurements indicated that
they were induced almost as frequently as pyrimidine dimers at 313
nm as against 21:1 at 254 nm (Cerutti & Netrawali, 1979), it has
been concluded from more recent data using sequencing analysis
(Mitchell et al., 1993) that they do not occur at a significant
rate in UVC or UVB irradiated DNA.
Increasingly sophisticated chemical methods are now becoming
available to measure oxidative DNA damage (Cadet et al., Methods
in Enzymology, in press). Since the proportion of this type of
damage will increase with increasing wavelength, such techniques
will soon be applied to consolidate the picture of UV-induced based
damage throughout the entire solar UV spectrum. Furthermore, an
interesting enzyme has been isolated (Boiteux et al., 1987) termed
the Fapy glycosylase because of its ability to excise the
ring-opened form of N7-methylguanine from DNA. The enzyme will also
recognize DNA products generated in photosensitisation reactions
involving singlet oxygen (Epe et al., in press) and
8-hydroxyguanine may be a major component. Since the longer UV
wavelength in sunlight clearly generate biologically relevant levels
of singlet oxygen and other active intermediates (Tyrrell, 1991;
Basu-Modak & Tyrrell, 1993), this type of analysis may also be
usefully applied to the solar UV spectrum. Certainly, near-visible
light alone is able to generate Fapy glycosylase-sensitive lesions
in the DNA of cultured mammalian cells and these, at least in part,
are probably 8-hydroxyguanine (Epe et al., 1993).
DNA strand breaks
Few DNA strand breaks are induced by UVC radiation but they
constitute an increasing proportion of the total lesions as
wavelength is increased. For example, in E. coli the ratio of DNA
strand breaks to pyrimidine dimers is 1:44 at 313 nm (Miguel &
Tyrrell, 1983) whereas at 365 nm one strand break is formed for
every two pyrimidine dimers (Tyrrell et al., 1974). Action spectra
have now been determined for the induction of DNA single strand
breaks in human skin cells which show that breaks occur throughout
the UVA and into the near-visible range (Peak et al., 1987). Since
break measurements involve alkaline denaturation, 10-20 percent of
the so-called breaks are due to the fragility of chemical bonds at
apurinic and apyrimidinic sites. It is important to note that unlike
most of the common forms of base damage, induction of strand breaks
is strongly dependent on oxygen throughout the UVB and UVA ranges
(Tyrrell, et al., 1974; Peak & Peak, 1982; Miguel and Tyrrell, 1983)
and at the longest wavelengths may involve generation of singlet
oxygen (Peak et al., 1987). Figure 6.5 gives the action spectra
for single strand breaks and figure 6.6 for double strand breaks in
cultured cells.
DNA-protein cross links
Photochemically induced DNA-protein cross links, mostly
involving cysteine are clearly formed in vitro (Smith, 1976;
Shetlar, 1980) and appear to be formed in vivo, particularly in
the UVA range (Bradley, et al., 1979; Peak & Peak, 1991). Despite
the obvious importance of this type of damage, there is little known
at the molecular level concerning the nature of the damage formed
in vivo. Action spectra for DNA-protein cross links in cultured
cells is given in figure 6.7 (from Peak et al., 1985a, b).
Ribonucleic acid
Messenger RNA is readily susceptible to modification by UV
radiation. However, given the fairly rapid turnover of most
molecules of this type (half-lives generally of the order of minutes
and hours) and the capacity for de novo synthesis in the absence
of DNA damage, mRNA is not generally considered a critical target of
radiation damage in mammalian cells. Bacterial transfer RNA is often
extremely photosensitive because of the presence of an unusual
nucleoside 4-thiouridine and this leads to some fascinating
photobiological phenomena of ecological significance (Jagger, 1985).
However, such findings are of little significance to eukaryotic
cells.
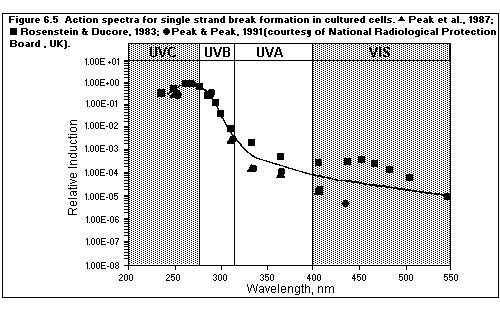
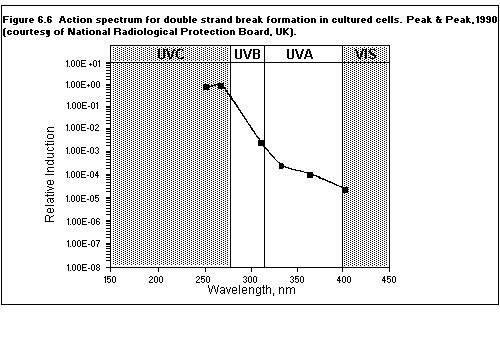
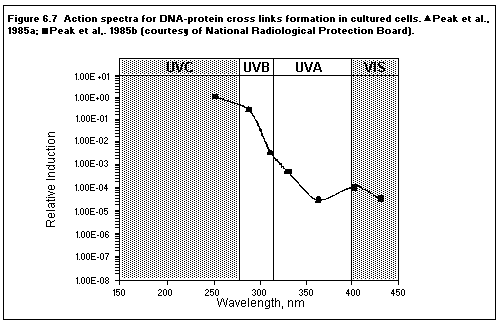 6.4.2 Membranes
UV-induced changes in membrane permeability and membrane
transport systems would be expected to have fairly dramatic
consequences for human skin and eyes. Unsaturated fatty acids are
readily oxidised to hydroperoxides. Several reports have now shown
that UV radiation can peroxidise membrane lipids (Desai, et al.,
1964; Roshchupkin et al., 1975; Putvinsky et al., 1979; Azizova,
et al., 1980). Using liposomal models (Mandal & Chatterjee, 1980;
Bose et al., 1989), it has been shown that UVA radiation causes a
dose-dependent increase in lipid peroxidation as measured by various
techniques and that this can be largely inhibited by membrane
antioxidants such as butylated hydroxytoluene. In E coli,
sensitivity to UVA correlates with the levels of unsaturated fat in
membranes (Klamen & Tuveson, 1982; Chamberlain & Moss, 1987). An
agent which enhances singlet oxygen lifetime, deuterium oxide,
enhances the level of membrane damage, sensitivity to UVA and lipid
peroxidation. There are now reports that both UVB and UVA (Morlière
et al., 1991; Punnonen et al., 1991; Vile et al., 1994)
radiation can cause lipid peroxidation at biologically relevant
fluences in the membranes of human fibroblasts and keratinocytes.
Cell leakage experiments have been used to assess membrane
damage in yeast (Ito and Ito, 1983) and a similar technique has now
been used to show UVA-mediated enhancement of membrane leakage in
human skin fibroblasts (Gaboriau et al., 1993; Vile et al.,
1994). UVA also causes changes in membrane fluidity as assessed
using a lipophilic fluorescent probe. UVA-induced alteration of
membrane transport systems received considerable attention in
prokaryotes (see Jagger, 1985) but little information is available
in mammalian cells. A recent study has demonstrated that UVA
radiation inhibits both receptor-mediated (low density lipoprotein)
and nonspecific (sucrose) uptake of exogenous molecules
(Djavuheri-Mergny et al., 1993). These findings may be related to
other studies in human fibroblasts which have shown that broad
spectrum UV sources cause cytoskeletal damage as manifested by
dose-dependent microtubule disassembly (Zamansky & Chou, 1987).
6.4.3 Proteins
As the wavelength is increased through the UVB and UVA regions
damage to proteins becomes increasingly important because of the
absorption properties of the aromatic amino acids relative to
nucleic acids. In addition, many proteins that include the
antioxidant enzymes catalases and peroxidases contain heme groups
thus making them UVA chromophores and potentially photosensitisers.
Indeed, catalase is sensitive to sunlight (Mitchell & Anderson,
1965), probably as a result of UVA absorption (Kramer & Ames, 1987).
In bacteria there is evidence that endogenous catalase and possibly
alkyl hydroperoxide reductase are actually photosensitisers
(Eisenstark & Perrot, 1987; Kramer & Ames, 1987) but no similar
experiments to have been carried out in mammalian cells. Repair
enzymes are also sensitive to UVA radiation and there is evidence
that UV-induced repair disruption plays a role in cell death and
mutagenesis (Haynes, 1966; Webb, 1977; Tyrrell, 1982). Considerable
attention has been given recently to metal ion catalysed oxidation
of protein since this is clearly a physiologically relevant process
(for review see Stadtman, 1990). Numerous proteins have been shown
to be modified by a free radical generating, system modelling those
that cells are exposed to during normal metabolism or exogenous
insult. It appears that active oxygen/free radical species are
generated at specific metal binding sites on proteins and that this
leads to reactions with amino acid residues at specific steric
locations.
6.5 Cellular Defences
6.5.1 DNA
Even one or two pyrimidine dimers in the entire genome of a
bacterium such as E coli may be lethal if DNA repair processes are
defective. However, DNA repair processes are not only crucial for
cell survival but also for the maintenance of genetic stability
since DNA damage is continuously generated as a by-product of
metabolism even in the absence of exogenous insults. Because of the
availability of mutants, most of the original studies concerning
removal of DNA damage employed bacteria and bacteriophage. The first
process to be identified was photoreactivation later known as
photoenzymatic repair. In this process, a DNA photolyase forms a
specific complex with a pyrimidine dimer which can be split in the
presence of UVA and/or visible light to leave the original
pyrimidines in the DNA. The process has been widely used to
correlate a specific type of DNA damage (the pyrimidine dimer) with
a given biological effect (from cell death in bacteria to cancer in
fish), although recent evidence for an enzyme which recognizes (6-4)
photoproducts may cast doubt on the validity of this approach.
Photoenzymatic repair is entirely error free (i.e. it does not lead
to mutations) and occurs in a wide variety of single and
multiple-celled organisms. Although photoenzymatic repair and
associated photolyases occur in non-placental mammals such as the
marsupials, evidence for photoreactivation in placental mammals,
including humans, has been fragmentary and controversial. Recent
evidence suggesting that photoreactivation of UV damage may not be a
significant process in human skin (Li et al., 1993) could be of
crucial importance when considering potential interactions between
wavelengths.
Evidence for two broad categories of DNA repair, originally
denoted as dark repair processes, were revealed by bacterial studies
(see Friedberg, 1984 for review). Post-replicational repair, as the
name implies, can occur only after DNA synthesis and includes many
of the complex pathways of genetic recombination. Humans with the
genetic disease Xeroderma pigmentosum (XP) of the variant
complementation group are deficient in this type of repair. However,
the SOS repair process in bacteria which involves the inducible
activation of a post-replication error-prone (mutagenic) repair
pathway, has no equivalent in humans. On the other hand, excision
repair is the most widespread of DNA repair processes and occurs
from bacteria to man. In excision repair, damaged or incorrect bases
are excised from the genome and replaced by the correct nucleotides.
The first step in the process involves recognition of the damage and
incision at or close to the damaged site. This can occur by two
distinct mechanisms. Damaged bases may be recognised by a series of
specific glycosylases. In this case, incision occurs by a two-step
reaction which involves the sequential activities of the DNA
glycosylase and an apyrimidinic/apurinic endonuclease. However, a
second mechanism of incision involves the direct action of a
damage-specific DNA incising activity without the need for
glycosylase action. At least in E Coli, incisions are made at
either side of the damaged base so that exonucleolytic removal of
bases is unnecessary in this case. Repolymerization of the gap
created by endonuclease/exonuclease activity is an essential step
and can take place using the undamaged strand as a template. The
action of DNA ligase restores the original integrity of the
double-stranded DNA.
DNA excision repair in humans is also an extremely complex
process as evidenced by the considerable genetic heterogeneity in XP
which is a rare but extensively studied disease whose cellular basis
involves a defect in DNA repair. Genomic clones of several human DNA
repair genes have now been derived by transfecting DNA from repair
proficient human cells into a series of UV-sensitive mutant rodent
cell lines (Westerveld et al., 1984). A considerable amount of
information is now available concerning the molecular genetics of
eukaryotic DNA excision repair (Hoeijmakers & Bootsma, 1990). The
repair genes isolated to date often have DNA binding and/or nucleic
acid helicase domains. Most recently, a repair gene product has been
identified as one of the components of a transcription factor
complex involved in the transcription of polymerase II genes
(Schaeffer et al., 1993). This is currently an extremely active
area of research and further genes are expected to be isolated.
In vitro repair systems are now available (Wood et al.,
1988). In addition, animal models are now being developed using
homologous recombination techniques and genetic manipulation of
embryonic stem cells.
6.5.2 Human excision repair disorders
Several human diseases involve defects in DNA repair. The most
studied example is the genetic disorder XP which actually involves
several types of excision repair defect as well as the variant form
which is defective in post-replication repair. These cell lines are
crucial to basic studies of DNA repair in humans. Cell lines derived
from the excision-defective individuals are many times more
sensitive to inactivation and mutation by UVC and UVB radiation
(Arlett et al., 1992). This appears to be directly correlated with
the fact that individuals with this disease are extremely prone to
tumours of the skin, eye and lips (Kraemer et al., 1987). Indeed,
it is the study of cancer incidence in these individuals which
provides the strongest evidence that the induction of photoproducts
in skin by UV is the first event in a sequence which eventually
leads to basal cell carcinoma, squamous cell carcinoma and melanomas
in man.
The above conclusion is now complicated by studies with cells
from patients with the rare disease, Trichothiodystrophy. Although,
cells from most of the patients studied so far have a marked defect
in DNA excision repair, the patients do not show elevated evidence
of skin tumours. They are, nevertheless, sun-sensitive in terms of
their erythemal response (Bridges 1990). A partial explanation may
be that XP patients but not Trichothiodystrophy patients have
defects in both DNA repair and oxidative metabolism and both these
processes are involved in carcinogenesis (Vuillaume et al., 1992).
Fibroblasts from patients with Cockayne's syndrome are
specifically defective in excision of dimers from DNA undergoing
active transcription (Mayne et al., 1988) and are sensitive to
killing and mutation by UVC radiation. (Arlett & Harcourt 1983).
However, these individuals are not exceptionally cancer-prone
(Barret et al., 1991).
6.5.3 Antioxidant pathways
Small antioxidant molecules
Recent reviews have evaluated the nature and importance of
small antioxidant molecules in human blood plasma (Stocker & Frei,
1991), the eye (Spector, 1991) and human skin (Fuchs & Packer 1991;
Tyrrell 1991).
Glutathione is a major constituent of lens epithelial cells
(Rathbun 1989) and these cells have a high capacity for maintaining
glutathione in the reduced state because of extremely active hexose
monophosphate shunt activity. This compound may be involved in a
number of antioxidant reactions that are relevant to UV-mediated
oxidative stress including detoxification of hydrogen peroxide (as a
cofactor of GSH peroxidase), detoxification of free radicals,
reduction of protein disulphides, and competition with protein
thiols for oxidising species. The role of ascorbate as an effective
antioxidant in lens is less clear because of the potential of
ascorbate to be involved in the generation of deleterious components
(Spector, 1991). Vitamin E is also a potentially useful antioxidant
but there is no general agreement as to the usefulness of vitamin E
therapy for cataract.
Free radical intermediates have been implicated in UV- induced
carcinogenesis in investigations originally stimulated by the
isolation of the putative carcinogen cholesterol epoxide from human
skin (Black 1987). Various components present in skin are potent
antioxidants including ascorbate, uric acid, carotenoids and
sulphydrils. Carotenoids have been shown to inhibit UV-induced
epidermal damage and tumour formation in mouse models (Mathews-Roth
& Krinsky, 1987). In cell culture models using human skin cells, it
has been clearly shown that glutathione depletion leads to a large
sensitization to UVA (334 nm, 365 nm) and near-visible (405 nm)
wavelengths as well as to radiation in the UVB (302 nm, 313 nm)
(Tyrrell & Pidoux, 1986,1988). There is a direct correlation between
the levels of sensitisation and cellular glutathione content.
Additional evidence that glutathione is a photoprotective agent in
skin cells is derived from experiments which have demonstrated that
glutathione levels in both dermis and epidermis are depleted by UVA
treatment (Connor & Wheeler, 1987).
Water-soluble antioxidants in plasma include glucose, pyruvate,
uric acid, ascorbic acid, bilirubin and glutathione. Lipid soluble
anti-oxidants include alpha-tocopherol, ubiquinol-10, lycopene,
ß-carotene, lutein, zeaxanthin and alpha-carotene. Since, the long
wavelengths in sunlight can penetrate through tissue and into blood
at the longer wavelengths, these defenses may be critical under
certain circumstances.
Antioxidant proteins including enzymes
The major classes of antioxidant enzymes characterized to date
in eukaryotic cells are superoxide dismutase (which converts
superoxide anion to hydrogen peroxide), catalase (which destroys
hydrogen peroxide) and glutathione peroxide and associated enzymes
(which in addition to metabolizing hydrogen peroxide can also reduce
hydroperoxides such as those that result from lipid peroxidation).
Both glutathione peroxide and catalase are present in the lens,
although the latter is present at low levels and concentrated in the
epithelial cells (Bhuyan & Bhuyan 1983). Spector (1991) claims that
these enzymes are present at sufficient concentrations to handle the
normal levels of hydrogen peroxide generated in the lens. This
author suggests that the thioredoxin/thioredoxin-reductase system
may also be involved in the defense of lens against oxidative
stress. This system can quench free radicals and also reduce some
protein disulphides.
All the major antioxidant enzymes are present in skin but their
role in protecting cells against oxidative damage generated by UV
radiation has not been elucidated. Acute UV exposures lead to
changes in glutathione reductase and catalase activity in mouse skin
but insignificant changes in superoxide dismutase and glutathione
peroxidase (Fuchs et al., 1989). Consistent with original studies
in bacteria, neither endogenous catalase nor superoxide dismutase
play a major role in protecting cells against the lethal effects of
UVA irradiation (Tyrrell & Pidoux 1989). Iron plays a critical role
in oxidative reactions as a catalyst in the Fenton reaction so that
cellular levels of free iron need to be kept low. The intracellular
storage protein, ferritin may therefore play a critical role in
cellular antioxidant defense. UVA radiation (and other oxidant
stress) leads to high levels of expression of the heme oxygenase 1
gene (HO1) (Keyse & Tyrrell 1989) which in turn leads to the
catabolism of heme and release of free iron. The increased ferritin
levels that result appear to be directly responsible for a
UVA-mediated adaptive response involving the protection of human
fibroblast membranes against subsequent UVA radiation damage (Vile
et al., 1994).
6.5.4 Summary
Repair of UV-induced DNA damage is crucial in removing
potentially mutagenic damage from cells although errors in repair
can themselves lead to mutations. The repair capacity of human skin
cells therefore directly relates to the probability of initiation of
the carcinogenesis process and eventually tumour formation. At
longer UV wavelengths, an increasing component of oxidative damage
to DNA, membranes and proteins influences the biological effects.
Both endogenous and exogenous photosensitisers normally generate
active oxygen intermediates. Cellular antioxidant defense mechanisms
are therefore crucial for the prevention or removal of the damage
caused by the oxidising component of UV radiation.
6.6 Cellular Consequences of Damage
6.6.1 Membrane disruption
Lipid and protein damage by UV associated with cytoskeletal
damage may lead to severe disruption of plasma membrane functions
including a breakdown in the permeability barrier and interruptions
in active transport functions. Crucial signalling molecules such as
cytokines may be inappropriately released leading to aberrant cell
to cell communication and toxins present in the external cellular
environment may gain free access to the interior of the cell.
Critical ion pumps may be damaged, thereby influencing a wide range
of processes that rely on ion homeostasis. Breakdown of internal
lipid membranes in eukaryotic cells will also be highly disruptive
leading to many pathological consequences including mitochondrial
damage, leakage of proteases from disrupted lysosomes and breakdown
of the nuclear membrane permeability barrier. Details in this
important area of UV effects are sparse but it is clear that
consequences of UV damage to membrane components need to be better
understood and incorporated into current models for cellular and
organ damage.
6.6.2 Activation of genes
UV inducible defense pathways in bacteria and human cells
In bacteria, UVC radiation induces a large set of genes under
the regulatory control of the rec A gene which lead to enhanced
DNA repair, mutagenesis, prophage induction and inhibition of cell
division (for review see Walker, 1987). Many groups have sought a
similar SOS response in eukaryotic cells. Reactivation of several
different UVC damaged viruses (human cytomegalovirus or Herpes
Simplex virus) has been observed upon infection into UVC treated
cultured human skin fibroblasts. The effects are not generally large
(Dion & Hammelin, 1987; Abrahams et al., 1984). In one study, a
parallel phenomenon has been observed using split doses of UV in
arrested cultures of human fibroblasts (Tyrrell, 1984). Cell
populations irradiated with low doses of UV developed enhanced
resistance to a second UV challenge dose with a maximum response
occurring between 2-4 days. The level of reactivation was much
higher in a repair deficient (XP variant cell line). As for viral
reactivation, protein synthesis is necessary. No mutation studies
have been carried out under similar conditions. Furthermore, these
phenomena have not been investigated in the solar UVB region
although it is likely that the response will be similar. Although
these adaptation responses appear to reflect enhanced gene
expression, there are few clues as to the molecular mechanism
underlying these observations.
Bacteria also respond strongly to oxidative stress by either
the oxyR pathway (Christman et al., 1985) or the soxRS pathway
(Greenberg et al., 1990) which is induced by superoxide-
generating redox cycling agents such as menadione and paraquat. The
oxyR pathway is induced by agents such as hydrogen peroxide and the
regulatory protein encoded by this gene controls expression of at
least 9 proteins including the antioxidant enzymes, catalase,
glutathione reductase and alkyl hydroperoxide reductase. This
pathway is emphasised here because it also appears to be involved in
protection of bacterial cells against the cytotoxic action of UVA
radiation (Kramer & Ames 1987). However, although several eukaryotic
genes are induced by both UVA radiation and hydrogen peroxide (see
below), none of them involves such a clear-cut direct activation of
antioxidant enzymes.
Gene activation in mammalian cells by UVC and UVB radiations
A large number of genes have been shown to be activated by UVC
radiation (listed in Table 1 of a review by Keyse 1993). However, in
many cases the doses required are very high, so that by the
criterion of colony-forming ability, the majority of the population
is dead. Although it is a reasonable assumption that the same genes
will be induced by UVB radiation, this has only been shown in
certain cases. Damage to DNA may be a critical intermediate in
triggering the response since induction of certain genes has been
shown to occur at much lower doses in mutant cell strains lacking
DNA repair. (Stein et al., 1989, Miskin & Ben-Ishai, 1981).
Furthermore, crude action spectra for induction of several genes by
UV radiation correspond to the action spectra for DNA damage
induction (Stein et al., 1989). However, at least for the
induction of the fos gene (see action spectrum in figure 6.8),
events at the membrane also appear to be involved (Devary et al.,
1992). There is still a controversy as to whether or not the initial
signal occurs in the nucleus and is then transduced to the membrane
or whether the crucial initiating events occur at the membrane
itself. UVC activation of the HIV-1 promoter also appears to involve
membrane events (Devary et al., 1993).
Metallothioneins are also induced by both UVC and UVB radiation
(Angel et al., 1986). However, in general, the relationship
between UVC/UVB activation of genes and cellular defense against
short wavelength UV radiations remains obscure. The synthesis of a
constitutive damage specific DNA-binding protein has been shown to
be stimulated by UV radiation in both monkey and human cells
(Hirschfield et al., 1990). Although the binding activity was
shown to be absent in certain strains of XP (group E) (Chu & Chang
1988), subsequent studies showed that not all strains from this
group lacked the damage specific protein (Kataoka & Fujiwara 1991).
Gene activation in mammalian cells by UVA radiation
The eukaryotic genes induced by UVA radiation are, for the most
part, distinct from those induced by the shorter wavelengths. This
appears to be largely due to the fact that UVA radiation acts
indirectly by generating active oxygen intermediates rather than
being directly absorbed by biomolecules. Although UVA radiation can
activate antioxidant enzymes such as catalase in prokaryotic cells,
there is no evidence that UVA radiation can activate any of the
common antioxidant pathways in mammalian cells. In contrast to the
studies with UVB/UVC radiations, UVA radiation can activate genes at
doses which kill only a small percentage of the total population in
cultured cell models. This is crucial since it indicates that gene
activation by UVA radiation may actually relate to events in living
tissue and could reasonably be expected to be involved in protective
pathways.
6.4.2 Membranes
UV-induced changes in membrane permeability and membrane
transport systems would be expected to have fairly dramatic
consequences for human skin and eyes. Unsaturated fatty acids are
readily oxidised to hydroperoxides. Several reports have now shown
that UV radiation can peroxidise membrane lipids (Desai, et al.,
1964; Roshchupkin et al., 1975; Putvinsky et al., 1979; Azizova,
et al., 1980). Using liposomal models (Mandal & Chatterjee, 1980;
Bose et al., 1989), it has been shown that UVA radiation causes a
dose-dependent increase in lipid peroxidation as measured by various
techniques and that this can be largely inhibited by membrane
antioxidants such as butylated hydroxytoluene. In E coli,
sensitivity to UVA correlates with the levels of unsaturated fat in
membranes (Klamen & Tuveson, 1982; Chamberlain & Moss, 1987). An
agent which enhances singlet oxygen lifetime, deuterium oxide,
enhances the level of membrane damage, sensitivity to UVA and lipid
peroxidation. There are now reports that both UVB and UVA (Morlière
et al., 1991; Punnonen et al., 1991; Vile et al., 1994)
radiation can cause lipid peroxidation at biologically relevant
fluences in the membranes of human fibroblasts and keratinocytes.
Cell leakage experiments have been used to assess membrane
damage in yeast (Ito and Ito, 1983) and a similar technique has now
been used to show UVA-mediated enhancement of membrane leakage in
human skin fibroblasts (Gaboriau et al., 1993; Vile et al.,
1994). UVA also causes changes in membrane fluidity as assessed
using a lipophilic fluorescent probe. UVA-induced alteration of
membrane transport systems received considerable attention in
prokaryotes (see Jagger, 1985) but little information is available
in mammalian cells. A recent study has demonstrated that UVA
radiation inhibits both receptor-mediated (low density lipoprotein)
and nonspecific (sucrose) uptake of exogenous molecules
(Djavuheri-Mergny et al., 1993). These findings may be related to
other studies in human fibroblasts which have shown that broad
spectrum UV sources cause cytoskeletal damage as manifested by
dose-dependent microtubule disassembly (Zamansky & Chou, 1987).
6.4.3 Proteins
As the wavelength is increased through the UVB and UVA regions
damage to proteins becomes increasingly important because of the
absorption properties of the aromatic amino acids relative to
nucleic acids. In addition, many proteins that include the
antioxidant enzymes catalases and peroxidases contain heme groups
thus making them UVA chromophores and potentially photosensitisers.
Indeed, catalase is sensitive to sunlight (Mitchell & Anderson,
1965), probably as a result of UVA absorption (Kramer & Ames, 1987).
In bacteria there is evidence that endogenous catalase and possibly
alkyl hydroperoxide reductase are actually photosensitisers
(Eisenstark & Perrot, 1987; Kramer & Ames, 1987) but no similar
experiments to have been carried out in mammalian cells. Repair
enzymes are also sensitive to UVA radiation and there is evidence
that UV-induced repair disruption plays a role in cell death and
mutagenesis (Haynes, 1966; Webb, 1977; Tyrrell, 1982). Considerable
attention has been given recently to metal ion catalysed oxidation
of protein since this is clearly a physiologically relevant process
(for review see Stadtman, 1990). Numerous proteins have been shown
to be modified by a free radical generating, system modelling those
that cells are exposed to during normal metabolism or exogenous
insult. It appears that active oxygen/free radical species are
generated at specific metal binding sites on proteins and that this
leads to reactions with amino acid residues at specific steric
locations.
6.5 Cellular Defences
6.5.1 DNA
Even one or two pyrimidine dimers in the entire genome of a
bacterium such as E coli may be lethal if DNA repair processes are
defective. However, DNA repair processes are not only crucial for
cell survival but also for the maintenance of genetic stability
since DNA damage is continuously generated as a by-product of
metabolism even in the absence of exogenous insults. Because of the
availability of mutants, most of the original studies concerning
removal of DNA damage employed bacteria and bacteriophage. The first
process to be identified was photoreactivation later known as
photoenzymatic repair. In this process, a DNA photolyase forms a
specific complex with a pyrimidine dimer which can be split in the
presence of UVA and/or visible light to leave the original
pyrimidines in the DNA. The process has been widely used to
correlate a specific type of DNA damage (the pyrimidine dimer) with
a given biological effect (from cell death in bacteria to cancer in
fish), although recent evidence for an enzyme which recognizes (6-4)
photoproducts may cast doubt on the validity of this approach.
Photoenzymatic repair is entirely error free (i.e. it does not lead
to mutations) and occurs in a wide variety of single and
multiple-celled organisms. Although photoenzymatic repair and
associated photolyases occur in non-placental mammals such as the
marsupials, evidence for photoreactivation in placental mammals,
including humans, has been fragmentary and controversial. Recent
evidence suggesting that photoreactivation of UV damage may not be a
significant process in human skin (Li et al., 1993) could be of
crucial importance when considering potential interactions between
wavelengths.
Evidence for two broad categories of DNA repair, originally
denoted as dark repair processes, were revealed by bacterial studies
(see Friedberg, 1984 for review). Post-replicational repair, as the
name implies, can occur only after DNA synthesis and includes many
of the complex pathways of genetic recombination. Humans with the
genetic disease Xeroderma pigmentosum (XP) of the variant
complementation group are deficient in this type of repair. However,
the SOS repair process in bacteria which involves the inducible
activation of a post-replication error-prone (mutagenic) repair
pathway, has no equivalent in humans. On the other hand, excision
repair is the most widespread of DNA repair processes and occurs
from bacteria to man. In excision repair, damaged or incorrect bases
are excised from the genome and replaced by the correct nucleotides.
The first step in the process involves recognition of the damage and
incision at or close to the damaged site. This can occur by two
distinct mechanisms. Damaged bases may be recognised by a series of
specific glycosylases. In this case, incision occurs by a two-step
reaction which involves the sequential activities of the DNA
glycosylase and an apyrimidinic/apurinic endonuclease. However, a
second mechanism of incision involves the direct action of a
damage-specific DNA incising activity without the need for
glycosylase action. At least in E Coli, incisions are made at
either side of the damaged base so that exonucleolytic removal of
bases is unnecessary in this case. Repolymerization of the gap
created by endonuclease/exonuclease activity is an essential step
and can take place using the undamaged strand as a template. The
action of DNA ligase restores the original integrity of the
double-stranded DNA.
DNA excision repair in humans is also an extremely complex
process as evidenced by the considerable genetic heterogeneity in XP
which is a rare but extensively studied disease whose cellular basis
involves a defect in DNA repair. Genomic clones of several human DNA
repair genes have now been derived by transfecting DNA from repair
proficient human cells into a series of UV-sensitive mutant rodent
cell lines (Westerveld et al., 1984). A considerable amount of
information is now available concerning the molecular genetics of
eukaryotic DNA excision repair (Hoeijmakers & Bootsma, 1990). The
repair genes isolated to date often have DNA binding and/or nucleic
acid helicase domains. Most recently, a repair gene product has been
identified as one of the components of a transcription factor
complex involved in the transcription of polymerase II genes
(Schaeffer et al., 1993). This is currently an extremely active
area of research and further genes are expected to be isolated.
In vitro repair systems are now available (Wood et al.,
1988). In addition, animal models are now being developed using
homologous recombination techniques and genetic manipulation of
embryonic stem cells.
6.5.2 Human excision repair disorders
Several human diseases involve defects in DNA repair. The most
studied example is the genetic disorder XP which actually involves
several types of excision repair defect as well as the variant form
which is defective in post-replication repair. These cell lines are
crucial to basic studies of DNA repair in humans. Cell lines derived
from the excision-defective individuals are many times more
sensitive to inactivation and mutation by UVC and UVB radiation
(Arlett et al., 1992). This appears to be directly correlated with
the fact that individuals with this disease are extremely prone to
tumours of the skin, eye and lips (Kraemer et al., 1987). Indeed,
it is the study of cancer incidence in these individuals which
provides the strongest evidence that the induction of photoproducts
in skin by UV is the first event in a sequence which eventually
leads to basal cell carcinoma, squamous cell carcinoma and melanomas
in man.
The above conclusion is now complicated by studies with cells
from patients with the rare disease, Trichothiodystrophy. Although,
cells from most of the patients studied so far have a marked defect
in DNA excision repair, the patients do not show elevated evidence
of skin tumours. They are, nevertheless, sun-sensitive in terms of
their erythemal response (Bridges 1990). A partial explanation may
be that XP patients but not Trichothiodystrophy patients have
defects in both DNA repair and oxidative metabolism and both these
processes are involved in carcinogenesis (Vuillaume et al., 1992).
Fibroblasts from patients with Cockayne's syndrome are
specifically defective in excision of dimers from DNA undergoing
active transcription (Mayne et al., 1988) and are sensitive to
killing and mutation by UVC radiation. (Arlett & Harcourt 1983).
However, these individuals are not exceptionally cancer-prone
(Barret et al., 1991).
6.5.3 Antioxidant pathways
Small antioxidant molecules
Recent reviews have evaluated the nature and importance of
small antioxidant molecules in human blood plasma (Stocker & Frei,
1991), the eye (Spector, 1991) and human skin (Fuchs & Packer 1991;
Tyrrell 1991).
Glutathione is a major constituent of lens epithelial cells
(Rathbun 1989) and these cells have a high capacity for maintaining
glutathione in the reduced state because of extremely active hexose
monophosphate shunt activity. This compound may be involved in a
number of antioxidant reactions that are relevant to UV-mediated
oxidative stress including detoxification of hydrogen peroxide (as a
cofactor of GSH peroxidase), detoxification of free radicals,
reduction of protein disulphides, and competition with protein
thiols for oxidising species. The role of ascorbate as an effective
antioxidant in lens is less clear because of the potential of
ascorbate to be involved in the generation of deleterious components
(Spector, 1991). Vitamin E is also a potentially useful antioxidant
but there is no general agreement as to the usefulness of vitamin E
therapy for cataract.
Free radical intermediates have been implicated in UV- induced
carcinogenesis in investigations originally stimulated by the
isolation of the putative carcinogen cholesterol epoxide from human
skin (Black 1987). Various components present in skin are potent
antioxidants including ascorbate, uric acid, carotenoids and
sulphydrils. Carotenoids have been shown to inhibit UV-induced
epidermal damage and tumour formation in mouse models (Mathews-Roth
& Krinsky, 1987). In cell culture models using human skin cells, it
has been clearly shown that glutathione depletion leads to a large
sensitization to UVA (334 nm, 365 nm) and near-visible (405 nm)
wavelengths as well as to radiation in the UVB (302 nm, 313 nm)
(Tyrrell & Pidoux, 1986,1988). There is a direct correlation between
the levels of sensitisation and cellular glutathione content.
Additional evidence that glutathione is a photoprotective agent in
skin cells is derived from experiments which have demonstrated that
glutathione levels in both dermis and epidermis are depleted by UVA
treatment (Connor & Wheeler, 1987).
Water-soluble antioxidants in plasma include glucose, pyruvate,
uric acid, ascorbic acid, bilirubin and glutathione. Lipid soluble
anti-oxidants include alpha-tocopherol, ubiquinol-10, lycopene,
ß-carotene, lutein, zeaxanthin and alpha-carotene. Since, the long
wavelengths in sunlight can penetrate through tissue and into blood
at the longer wavelengths, these defenses may be critical under
certain circumstances.
Antioxidant proteins including enzymes
The major classes of antioxidant enzymes characterized to date
in eukaryotic cells are superoxide dismutase (which converts
superoxide anion to hydrogen peroxide), catalase (which destroys
hydrogen peroxide) and glutathione peroxide and associated enzymes
(which in addition to metabolizing hydrogen peroxide can also reduce
hydroperoxides such as those that result from lipid peroxidation).
Both glutathione peroxide and catalase are present in the lens,
although the latter is present at low levels and concentrated in the
epithelial cells (Bhuyan & Bhuyan 1983). Spector (1991) claims that
these enzymes are present at sufficient concentrations to handle the
normal levels of hydrogen peroxide generated in the lens. This
author suggests that the thioredoxin/thioredoxin-reductase system
may also be involved in the defense of lens against oxidative
stress. This system can quench free radicals and also reduce some
protein disulphides.
All the major antioxidant enzymes are present in skin but their
role in protecting cells against oxidative damage generated by UV
radiation has not been elucidated. Acute UV exposures lead to
changes in glutathione reductase and catalase activity in mouse skin
but insignificant changes in superoxide dismutase and glutathione
peroxidase (Fuchs et al., 1989). Consistent with original studies
in bacteria, neither endogenous catalase nor superoxide dismutase
play a major role in protecting cells against the lethal effects of
UVA irradiation (Tyrrell & Pidoux 1989). Iron plays a critical role
in oxidative reactions as a catalyst in the Fenton reaction so that
cellular levels of free iron need to be kept low. The intracellular
storage protein, ferritin may therefore play a critical role in
cellular antioxidant defense. UVA radiation (and other oxidant
stress) leads to high levels of expression of the heme oxygenase 1
gene (HO1) (Keyse & Tyrrell 1989) which in turn leads to the
catabolism of heme and release of free iron. The increased ferritin
levels that result appear to be directly responsible for a
UVA-mediated adaptive response involving the protection of human
fibroblast membranes against subsequent UVA radiation damage (Vile
et al., 1994).
6.5.4 Summary
Repair of UV-induced DNA damage is crucial in removing
potentially mutagenic damage from cells although errors in repair
can themselves lead to mutations. The repair capacity of human skin
cells therefore directly relates to the probability of initiation of
the carcinogenesis process and eventually tumour formation. At
longer UV wavelengths, an increasing component of oxidative damage
to DNA, membranes and proteins influences the biological effects.
Both endogenous and exogenous photosensitisers normally generate
active oxygen intermediates. Cellular antioxidant defense mechanisms
are therefore crucial for the prevention or removal of the damage
caused by the oxidising component of UV radiation.
6.6 Cellular Consequences of Damage
6.6.1 Membrane disruption
Lipid and protein damage by UV associated with cytoskeletal
damage may lead to severe disruption of plasma membrane functions
including a breakdown in the permeability barrier and interruptions
in active transport functions. Crucial signalling molecules such as
cytokines may be inappropriately released leading to aberrant cell
to cell communication and toxins present in the external cellular
environment may gain free access to the interior of the cell.
Critical ion pumps may be damaged, thereby influencing a wide range
of processes that rely on ion homeostasis. Breakdown of internal
lipid membranes in eukaryotic cells will also be highly disruptive
leading to many pathological consequences including mitochondrial
damage, leakage of proteases from disrupted lysosomes and breakdown
of the nuclear membrane permeability barrier. Details in this
important area of UV effects are sparse but it is clear that
consequences of UV damage to membrane components need to be better
understood and incorporated into current models for cellular and
organ damage.
6.6.2 Activation of genes
UV inducible defense pathways in bacteria and human cells
In bacteria, UVC radiation induces a large set of genes under
the regulatory control of the rec A gene which lead to enhanced
DNA repair, mutagenesis, prophage induction and inhibition of cell
division (for review see Walker, 1987). Many groups have sought a
similar SOS response in eukaryotic cells. Reactivation of several
different UVC damaged viruses (human cytomegalovirus or Herpes
Simplex virus) has been observed upon infection into UVC treated
cultured human skin fibroblasts. The effects are not generally large
(Dion & Hammelin, 1987; Abrahams et al., 1984). In one study, a
parallel phenomenon has been observed using split doses of UV in
arrested cultures of human fibroblasts (Tyrrell, 1984). Cell
populations irradiated with low doses of UV developed enhanced
resistance to a second UV challenge dose with a maximum response
occurring between 2-4 days. The level of reactivation was much
higher in a repair deficient (XP variant cell line). As for viral
reactivation, protein synthesis is necessary. No mutation studies
have been carried out under similar conditions. Furthermore, these
phenomena have not been investigated in the solar UVB region
although it is likely that the response will be similar. Although
these adaptation responses appear to reflect enhanced gene
expression, there are few clues as to the molecular mechanism
underlying these observations.
Bacteria also respond strongly to oxidative stress by either
the oxyR pathway (Christman et al., 1985) or the soxRS pathway
(Greenberg et al., 1990) which is induced by superoxide-
generating redox cycling agents such as menadione and paraquat. The
oxyR pathway is induced by agents such as hydrogen peroxide and the
regulatory protein encoded by this gene controls expression of at
least 9 proteins including the antioxidant enzymes, catalase,
glutathione reductase and alkyl hydroperoxide reductase. This
pathway is emphasised here because it also appears to be involved in
protection of bacterial cells against the cytotoxic action of UVA
radiation (Kramer & Ames 1987). However, although several eukaryotic
genes are induced by both UVA radiation and hydrogen peroxide (see
below), none of them involves such a clear-cut direct activation of
antioxidant enzymes.
Gene activation in mammalian cells by UVC and UVB radiations
A large number of genes have been shown to be activated by UVC
radiation (listed in Table 1 of a review by Keyse 1993). However, in
many cases the doses required are very high, so that by the
criterion of colony-forming ability, the majority of the population
is dead. Although it is a reasonable assumption that the same genes
will be induced by UVB radiation, this has only been shown in
certain cases. Damage to DNA may be a critical intermediate in
triggering the response since induction of certain genes has been
shown to occur at much lower doses in mutant cell strains lacking
DNA repair. (Stein et al., 1989, Miskin & Ben-Ishai, 1981).
Furthermore, crude action spectra for induction of several genes by
UV radiation correspond to the action spectra for DNA damage
induction (Stein et al., 1989). However, at least for the
induction of the fos gene (see action spectrum in figure 6.8),
events at the membrane also appear to be involved (Devary et al.,
1992). There is still a controversy as to whether or not the initial
signal occurs in the nucleus and is then transduced to the membrane
or whether the crucial initiating events occur at the membrane
itself. UVC activation of the HIV-1 promoter also appears to involve
membrane events (Devary et al., 1993).
Metallothioneins are also induced by both UVC and UVB radiation
(Angel et al., 1986). However, in general, the relationship
between UVC/UVB activation of genes and cellular defense against
short wavelength UV radiations remains obscure. The synthesis of a
constitutive damage specific DNA-binding protein has been shown to
be stimulated by UV radiation in both monkey and human cells
(Hirschfield et al., 1990). Although the binding activity was
shown to be absent in certain strains of XP (group E) (Chu & Chang
1988), subsequent studies showed that not all strains from this
group lacked the damage specific protein (Kataoka & Fujiwara 1991).
Gene activation in mammalian cells by UVA radiation
The eukaryotic genes induced by UVA radiation are, for the most
part, distinct from those induced by the shorter wavelengths. This
appears to be largely due to the fact that UVA radiation acts
indirectly by generating active oxygen intermediates rather than
being directly absorbed by biomolecules. Although UVA radiation can
activate antioxidant enzymes such as catalase in prokaryotic cells,
there is no evidence that UVA radiation can activate any of the
common antioxidant pathways in mammalian cells. In contrast to the
studies with UVB/UVC radiations, UVA radiation can activate genes at
doses which kill only a small percentage of the total population in
cultured cell models. This is crucial since it indicates that gene
activation by UVA radiation may actually relate to events in living
tissue and could reasonably be expected to be involved in protective
pathways.
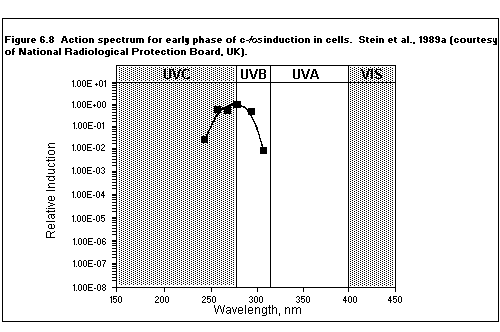 For simplicity, we can distinguish three groups of genes that
are activated in cultured mammalian cells by UVA radiation. These
may involve secreted molecules in cell to cell interactions:
collagenase (Scharffetter et al., 1991) and intracellular adhesion
molecule 1; proteins involved in early cellular events and signal
transduction (protein kinase C, phospholipase A2, and a phosphatase,
see Keyse, 1993) or catabolic enzymes (currently only represented by
heme oxygenase 1). To date only heme oxygenase 1 has been implicated
in a cellular defense pathway in which human fibroblasts adapt to
oxidative damage to membranes (Vile et al., 1994).
UV activation of viruses
UV radiation has long been known to activate Herpes Simplex
virus. Experimental evidence now indicates that UVC and UVB
radiation can activate the promoter of the human immune deficiency
virus (see figure 6.9) as well as complete HIV (Valerie et al.,
1988). This activation may contribute to the faster development of
AIDS in seropositive individuals. Current evidence in both cell
systems and transgenic animal models have led to the conclusion that
UVA radiation cannot activate the virus (see Beer & Smudzka, 1991).
However UVA at solar radiation levels can clearly activate binding
of the NFKB transcription factors to the corresponding DNA binding
site which is present in several promoters including the long
terminal repeat of the HIV promoter (Tyrrell, R.M Personal
communication). This issue clearly merits further study in view of
the importance to human health.
6.6.3 Cell death
Cells irradiated with UV may show changes in permeability,
inhibition of macromolecular synthesis, loss of ability to divide
and total metabolic disruption eventually leading to cellular
disruption. A discussion of how to define and measure cell death is
beyond the scope of this overview, but the most commonly accepted
parameter employed is the loss of the ability of a cell to divide
and form colonies. Many classical studies of UV effects were based
on these parameters and survival curves and their interpretation
have been extensively discussed (Jagger, 1985). Inactivation rates
can be derived from survival curves and used to construct action
spectra for inactivation of cultured organisms ranging from viruses
to human cells. Such survival curves are particularly useful for
defining sensitive and resistant cell populations, often an
indication of genetic defects in repair processes. Action spectra
for inactivation of cell populations generally follow DNA absorption
at wavelengths as long as 320 nm. However cells are killed more
efficiently than predicted by DNA damage at longer wavelengths as a
result of multiple effects of the longer wavelength including
disruption of DNA repair and the increasing importance of targets
such as the membrane.
For simplicity, we can distinguish three groups of genes that
are activated in cultured mammalian cells by UVA radiation. These
may involve secreted molecules in cell to cell interactions:
collagenase (Scharffetter et al., 1991) and intracellular adhesion
molecule 1; proteins involved in early cellular events and signal
transduction (protein kinase C, phospholipase A2, and a phosphatase,
see Keyse, 1993) or catabolic enzymes (currently only represented by
heme oxygenase 1). To date only heme oxygenase 1 has been implicated
in a cellular defense pathway in which human fibroblasts adapt to
oxidative damage to membranes (Vile et al., 1994).
UV activation of viruses
UV radiation has long been known to activate Herpes Simplex
virus. Experimental evidence now indicates that UVC and UVB
radiation can activate the promoter of the human immune deficiency
virus (see figure 6.9) as well as complete HIV (Valerie et al.,
1988). This activation may contribute to the faster development of
AIDS in seropositive individuals. Current evidence in both cell
systems and transgenic animal models have led to the conclusion that
UVA radiation cannot activate the virus (see Beer & Smudzka, 1991).
However UVA at solar radiation levels can clearly activate binding
of the NFKB transcription factors to the corresponding DNA binding
site which is present in several promoters including the long
terminal repeat of the HIV promoter (Tyrrell, R.M Personal
communication). This issue clearly merits further study in view of
the importance to human health.
6.6.3 Cell death
Cells irradiated with UV may show changes in permeability,
inhibition of macromolecular synthesis, loss of ability to divide
and total metabolic disruption eventually leading to cellular
disruption. A discussion of how to define and measure cell death is
beyond the scope of this overview, but the most commonly accepted
parameter employed is the loss of the ability of a cell to divide
and form colonies. Many classical studies of UV effects were based
on these parameters and survival curves and their interpretation
have been extensively discussed (Jagger, 1985). Inactivation rates
can be derived from survival curves and used to construct action
spectra for inactivation of cultured organisms ranging from viruses
to human cells. Such survival curves are particularly useful for
defining sensitive and resistant cell populations, often an
indication of genetic defects in repair processes. Action spectra
for inactivation of cell populations generally follow DNA absorption
at wavelengths as long as 320 nm. However cells are killed more
efficiently than predicted by DNA damage at longer wavelengths as a
result of multiple effects of the longer wavelength including
disruption of DNA repair and the increasing importance of targets
such as the membrane.
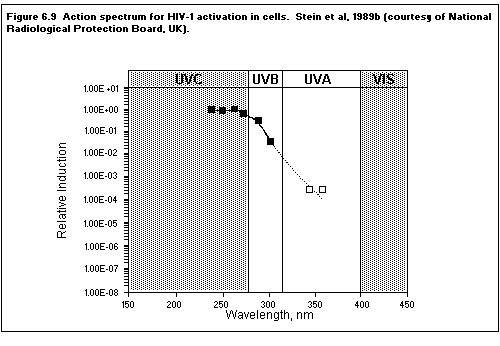 At solar UV levels that lead to erythema and acute skin burn in
human beings, extensive cell death may occur. This may be
particularly important to the eventual appearance of melanomas which
have been linked to severe sunburn early on in life in
epidemiological studies. It should also be noted that action spectra
data taken together with solar spectroradiometric measurements and
the known transmission of human skin have led to the conclusion that
the UVA component of sunlight is a major factor in the cytotoxic
action of sunlight at the basal layer of the epidermis (Tyrrell and
Pidoux 1987).
6.6.4 Mutation, chromosomal damage and transformation
Studies in microorganisms have unambiguously shown that damage
induced by UV throughout the whole range is mutagenic (Webb, 1977).
Until the present decade, UVB and UVA studies in mammalian cells
were much less conclusive because most of them employed broad
spectrum lamps and mutagenic effects could be partially or entirely
attributed to wavelengths at the short end of the emitted spectrum.
Nevertheless, a large body of work now supports the conclusion that
both UVC and UVB radiations are mutagenic to cultured human cells.
Studies using predominently UVA sources or monochromatic radiations
in three different cultured human cell systems have reported
positive (Enninga et al., 1986), ambiguous (Jones et al., 1987)
or negative (Tyrrell 1984) results. These results are clearly a
function of the system employed and merely underline the fact that
although pre-mutagenic lesions are induced by wavelengths longer
than 320 nm in human cells, the efficiency of mutation induction is
several orders of magnitude lower than that induced by the short
wavelengths. It should be stressed at this point that UVA radiation
is clearly carcinogenic in animal models (De Gruijl et al., 1993).
Most DNA photoproducts are likely to be pre-mutagenic lesions
because of the potential for error occurring during repair. Recent
attention has focused on the relative mutagenicity of the
cyclobutane type pyrimidine dimers compared with the second most
common DNA lesion formed by UVB and UVC radiation, the (6-4) pyo
photoproduct (reviewed by Mitchell et al., 1993). Most of the
conclusions are based on the specific type of sequence changes
(transitions, tandem transitions) that occur in a variety of test
systems. In certain cases, clear evidence has emerged that the minor
(6-4) pyo photoproduct may be highly mutagenic relative to the
pyrimidine dimer (Leclerc et al., 1991). Much additional study
will be required to resolve the mutagenic potential of each
photoproduct, especially considering the rapid expansion in numbers
of additional photoproducts that are now being recognised.
Except for UVC radiation, most of the studies of the
chromosomal effects of UV radiation have been carried out with broad
spectrum sources. The data has been extensively tabulated in a
recent evaluation of UV effects (IARC 1992). A similar comment
applies to the studies on morphological transformation.
6.7 Conclusions
DNA is the most critical chromophore and target for damage by
UVB and UVC radiation. The fraction of oxidative type damage
involving other chromophores and additional targets increases with
increasing wavelength. The determination of accurate action spectra
in cultured cells and animal models is critical to obtaining clues
as to the nature of these chromophores and for predictive evaluation
in the many cases in which human data is lacking.
A considerable amount of knowledge is available concerning the
interaction of UV radiation with nucleic acids. Controversy still
exists as to which type of lesion constitutes the most important
type of pre-mutagenic damage, although (6-4) pyo photoproducts and
cyclobutane type pyrimidine dimers may both be relevant. Damage to
membranes and other organelles is also being given increasing
attention since non-DNA events may also be involved in UV mediated
biological effects.
Studies of DNA repair defective disorders in humans have
clearly established a link between UV induced DNA damage in skin and
various types of cancer. A clear understanding of endogenous defense
pathways including antioxidant defense is essential for
understanding the origins of UV-related human disease and to the
elaboration of adequate protective measures.
Cell death, chromosome changes, mutation and morphological
transformations are observed after irradiation of prokaryotic and
eukaryotic cells with UV. Many different genes and several viruses
(including HIV) are activated by UV radiation. However, the genes
activated by UVC and UVB are different from those activated by UVA
radiation.
7. ANIMAL STUDIES
7.1 Skin Carcinogenesis
7.1.1 Domestic animals
Skin tumours have been reported in some domestic and food
animals including cats, dogs, cows, sheep and goats (Dorn et al.,
1971; Emmett, 1973; Madewell et al., 1981). That the tumours
observed often develop in sparsely haired, light-coloured skin
suggests that sunlight was involved. Cancers of the external
membranes of the eye are also observed, particularly in cattle, and
are thought to be related to sun exposure (Russell et al., 1956).
7.1.2 Experimental animals
The experimental induction of skin cancers in mice following
exposure to a mercury arc lamp was first reported by Findlay (1928).
Since then, carcinogenicity of UV has been investigated in many
experiments, mostly in mice, less often in rats, and infrequently in
other species (Blum et al, 1959; Urbach et al., 1974; Kripke &
Sass, 1978; WHO, 1979; van der Leun, 1984; Epstein, 1985).
Spontaneous skin tumours are rare in rodents but have been
consistently observed following experimental UV exposure with a
clear dose-response relationship in well-conducted studies. A recent
review concluded that there was sufficient evidence that UV caused
skin cancer in experimental animals (IARC, 1992).
Broad Spectrum UV
The carcinogenicity of sunlight was tested in two studies in
mice and rats (Roffo, 1934; 1939). In the first study of 600 rats,
165 (70%) of 235 which survived the acute heat load of exposure to
sunlight for 5 hours a day developed tumours of the skin (mainly
squamous cell carcinomas on the ears) or conjunctiva (spindle cell
sarcomas). No tumours developed when sunlight was filtered through
glass. Similar results were obtained in a second study in 2 000 rats
and mice.
"Solar-simulated radiation" has been studied in a number of
experiments (Forbes et al., 1982; Staberg et al., 1983; Young
et al., 1990; Menzies et al., 1991). In the study by Forbes et
al. (1982), 1 000 hairless mice were exposed for up to 80 weeks to
radiation from a xenon arc lamp passed through various filters to
simulate sunlight. More than 90% of the mice developed tumours,
particularly squamous cell carcinomas.
UVB
Many studies have been conducted with sources emitting mainly
UVB radiation. A few of the more informative studies in different
species are summarised below.
Mouse: Freeman (1975) exposed one ear of each haired albino
mouse in groups of 30, three times a week, to one of four
wavelengths of narrow band UVB (290, 300, 310 and 320 nm), produced
by a high-intensity diffraction grating monochromator with a
half-power band-width of 5 nm, at doses proportional to the MED of
each wavelength for untanned human skin. Tumours were produced in
about 50% of surviving animals at each wavelength except 290 nm
where no tumours were produced although all animals survived. Most
tumours were squamous cell carcinomas (SCC).
De Gruijl et al. (1983) exposed six groups of 22 to 44 (total
199) male and female Skh-hr 1 hairless albino mice to daily doses of
from 57 to 1 900 J m-2 of mainly UVB from Westinghouse FS40TL12
sunlamps. Most of the mice developed skin tumours, mainly squamous
cell carcinomas, even though the highest daily dose was
sub-erythemal. A power relationship (linear on log-log scales) was
observed between the daily dose and the time required for 50% of the
animals to develop tumours (figure 7.1). SCC developed in 71% of
mice in the lowest dose group and two skin tumours of different
types were observed late in the lives of 24 control mice (received
no UVB radiation).
Other Species: Stenbäck (1975a) exposed groups of shaven NMR
rats, Syrian golden hamsters and guinea pigs to mainly UVB from
Westinghouse FS40T12 sunlamps for 60 weeks at weekly doses of 5.4 to
10.8 J m-2. Tumours developed in 16 of 40 rats (mainly papillomas
on the ears), 14 of 40 hamsters (mainly papillomas of skin not on
the ears) and 2 of 25 guinea pigs.
A number of experiments were carried out in which groups of a
South American opossum, Monodelphis domestica, were exposed to
mainly UVB from Westinghouse FS40 sunlamps. M domestica is unusual
in showing photoreactivation of cyclobutylpyrimidine dimers (Ley,
1985). Regular exposures of 250 mJ m-2 produced melanomas in 5 of
13 surviving animals by 100 weeks from commencement of the
experiment. No melanomas were seen in a much larger colony not
exposed to artificial UV (Ley et al., 1989). In other experiments,
nonmelanocytic skin tumours (mainly fibrosarcomas and squamous cell
carcinomas) and fibrosarcomas of the corneal stroma were produced
(Ley et al., 1987, 1989). The latter were delayed in appearance
and reduced in number when UV exposure was followed immediately by
photoreactivating light.
At solar UV levels that lead to erythema and acute skin burn in
human beings, extensive cell death may occur. This may be
particularly important to the eventual appearance of melanomas which
have been linked to severe sunburn early on in life in
epidemiological studies. It should also be noted that action spectra
data taken together with solar spectroradiometric measurements and
the known transmission of human skin have led to the conclusion that
the UVA component of sunlight is a major factor in the cytotoxic
action of sunlight at the basal layer of the epidermis (Tyrrell and
Pidoux 1987).
6.6.4 Mutation, chromosomal damage and transformation
Studies in microorganisms have unambiguously shown that damage
induced by UV throughout the whole range is mutagenic (Webb, 1977).
Until the present decade, UVB and UVA studies in mammalian cells
were much less conclusive because most of them employed broad
spectrum lamps and mutagenic effects could be partially or entirely
attributed to wavelengths at the short end of the emitted spectrum.
Nevertheless, a large body of work now supports the conclusion that
both UVC and UVB radiations are mutagenic to cultured human cells.
Studies using predominently UVA sources or monochromatic radiations
in three different cultured human cell systems have reported
positive (Enninga et al., 1986), ambiguous (Jones et al., 1987)
or negative (Tyrrell 1984) results. These results are clearly a
function of the system employed and merely underline the fact that
although pre-mutagenic lesions are induced by wavelengths longer
than 320 nm in human cells, the efficiency of mutation induction is
several orders of magnitude lower than that induced by the short
wavelengths. It should be stressed at this point that UVA radiation
is clearly carcinogenic in animal models (De Gruijl et al., 1993).
Most DNA photoproducts are likely to be pre-mutagenic lesions
because of the potential for error occurring during repair. Recent
attention has focused on the relative mutagenicity of the
cyclobutane type pyrimidine dimers compared with the second most
common DNA lesion formed by UVB and UVC radiation, the (6-4) pyo
photoproduct (reviewed by Mitchell et al., 1993). Most of the
conclusions are based on the specific type of sequence changes
(transitions, tandem transitions) that occur in a variety of test
systems. In certain cases, clear evidence has emerged that the minor
(6-4) pyo photoproduct may be highly mutagenic relative to the
pyrimidine dimer (Leclerc et al., 1991). Much additional study
will be required to resolve the mutagenic potential of each
photoproduct, especially considering the rapid expansion in numbers
of additional photoproducts that are now being recognised.
Except for UVC radiation, most of the studies of the
chromosomal effects of UV radiation have been carried out with broad
spectrum sources. The data has been extensively tabulated in a
recent evaluation of UV effects (IARC 1992). A similar comment
applies to the studies on morphological transformation.
6.7 Conclusions
DNA is the most critical chromophore and target for damage by
UVB and UVC radiation. The fraction of oxidative type damage
involving other chromophores and additional targets increases with
increasing wavelength. The determination of accurate action spectra
in cultured cells and animal models is critical to obtaining clues
as to the nature of these chromophores and for predictive evaluation
in the many cases in which human data is lacking.
A considerable amount of knowledge is available concerning the
interaction of UV radiation with nucleic acids. Controversy still
exists as to which type of lesion constitutes the most important
type of pre-mutagenic damage, although (6-4) pyo photoproducts and
cyclobutane type pyrimidine dimers may both be relevant. Damage to
membranes and other organelles is also being given increasing
attention since non-DNA events may also be involved in UV mediated
biological effects.
Studies of DNA repair defective disorders in humans have
clearly established a link between UV induced DNA damage in skin and
various types of cancer. A clear understanding of endogenous defense
pathways including antioxidant defense is essential for
understanding the origins of UV-related human disease and to the
elaboration of adequate protective measures.
Cell death, chromosome changes, mutation and morphological
transformations are observed after irradiation of prokaryotic and
eukaryotic cells with UV. Many different genes and several viruses
(including HIV) are activated by UV radiation. However, the genes
activated by UVC and UVB are different from those activated by UVA
radiation.
7. ANIMAL STUDIES
7.1 Skin Carcinogenesis
7.1.1 Domestic animals
Skin tumours have been reported in some domestic and food
animals including cats, dogs, cows, sheep and goats (Dorn et al.,
1971; Emmett, 1973; Madewell et al., 1981). That the tumours
observed often develop in sparsely haired, light-coloured skin
suggests that sunlight was involved. Cancers of the external
membranes of the eye are also observed, particularly in cattle, and
are thought to be related to sun exposure (Russell et al., 1956).
7.1.2 Experimental animals
The experimental induction of skin cancers in mice following
exposure to a mercury arc lamp was first reported by Findlay (1928).
Since then, carcinogenicity of UV has been investigated in many
experiments, mostly in mice, less often in rats, and infrequently in
other species (Blum et al, 1959; Urbach et al., 1974; Kripke &
Sass, 1978; WHO, 1979; van der Leun, 1984; Epstein, 1985).
Spontaneous skin tumours are rare in rodents but have been
consistently observed following experimental UV exposure with a
clear dose-response relationship in well-conducted studies. A recent
review concluded that there was sufficient evidence that UV caused
skin cancer in experimental animals (IARC, 1992).
Broad Spectrum UV
The carcinogenicity of sunlight was tested in two studies in
mice and rats (Roffo, 1934; 1939). In the first study of 600 rats,
165 (70%) of 235 which survived the acute heat load of exposure to
sunlight for 5 hours a day developed tumours of the skin (mainly
squamous cell carcinomas on the ears) or conjunctiva (spindle cell
sarcomas). No tumours developed when sunlight was filtered through
glass. Similar results were obtained in a second study in 2 000 rats
and mice.
"Solar-simulated radiation" has been studied in a number of
experiments (Forbes et al., 1982; Staberg et al., 1983; Young
et al., 1990; Menzies et al., 1991). In the study by Forbes et
al. (1982), 1 000 hairless mice were exposed for up to 80 weeks to
radiation from a xenon arc lamp passed through various filters to
simulate sunlight. More than 90% of the mice developed tumours,
particularly squamous cell carcinomas.
UVB
Many studies have been conducted with sources emitting mainly
UVB radiation. A few of the more informative studies in different
species are summarised below.
Mouse: Freeman (1975) exposed one ear of each haired albino
mouse in groups of 30, three times a week, to one of four
wavelengths of narrow band UVB (290, 300, 310 and 320 nm), produced
by a high-intensity diffraction grating monochromator with a
half-power band-width of 5 nm, at doses proportional to the MED of
each wavelength for untanned human skin. Tumours were produced in
about 50% of surviving animals at each wavelength except 290 nm
where no tumours were produced although all animals survived. Most
tumours were squamous cell carcinomas (SCC).
De Gruijl et al. (1983) exposed six groups of 22 to 44 (total
199) male and female Skh-hr 1 hairless albino mice to daily doses of
from 57 to 1 900 J m-2 of mainly UVB from Westinghouse FS40TL12
sunlamps. Most of the mice developed skin tumours, mainly squamous
cell carcinomas, even though the highest daily dose was
sub-erythemal. A power relationship (linear on log-log scales) was
observed between the daily dose and the time required for 50% of the
animals to develop tumours (figure 7.1). SCC developed in 71% of
mice in the lowest dose group and two skin tumours of different
types were observed late in the lives of 24 control mice (received
no UVB radiation).
Other Species: Stenbäck (1975a) exposed groups of shaven NMR
rats, Syrian golden hamsters and guinea pigs to mainly UVB from
Westinghouse FS40T12 sunlamps for 60 weeks at weekly doses of 5.4 to
10.8 J m-2. Tumours developed in 16 of 40 rats (mainly papillomas
on the ears), 14 of 40 hamsters (mainly papillomas of skin not on
the ears) and 2 of 25 guinea pigs.
A number of experiments were carried out in which groups of a
South American opossum, Monodelphis domestica, were exposed to
mainly UVB from Westinghouse FS40 sunlamps. M domestica is unusual
in showing photoreactivation of cyclobutylpyrimidine dimers (Ley,
1985). Regular exposures of 250 mJ m-2 produced melanomas in 5 of
13 surviving animals by 100 weeks from commencement of the
experiment. No melanomas were seen in a much larger colony not
exposed to artificial UV (Ley et al., 1989). In other experiments,
nonmelanocytic skin tumours (mainly fibrosarcomas and squamous cell
carcinomas) and fibrosarcomas of the corneal stroma were produced
(Ley et al., 1987, 1989). The latter were delayed in appearance
and reduced in number when UV exposure was followed immediately by
photoreactivating light.
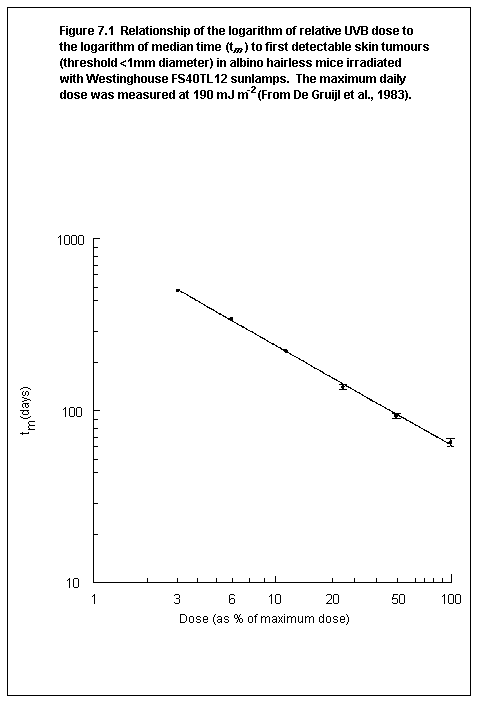 A group of 460 hybrid fish of two strains developed by crossing
platyfish ( Xiphophorus maculatus) and swordtails ( Xiphophorus
helleri) were exposed for 5 to 20 days to mainly UVB from
Westinghouse FS40 sunlamps filtered through acetate sheets
transmitting > 290 nm (150 or 300 mJ m-2) and >340 nm (850 and
1700 J m-2). Between 19% and 40% of exposed fish developed
melanocytic tumours compared with 12% and 2% in control fish (Setlow
et al., 1989).
In an attempt to confirm a suspected association between
sunlight and cancer of the eye in cattle, four Hereford cattle were
exposed to mainly UVB from Westinghouse FS40 sunlamps. Visible
tumours developed in the eyes of three of them and one was confirmed
histopathologically as a preneoplastic growth (Kopecky et al.,
1979).
UVC
Experiments have been carried out in which animals have been
exposed to low-pressure mercury discharge germicidal lamps which
emit most of their radiation at 254 nm with weaker spectral lines in
the UVB, UVA and visible spectra (IARC, 1992). Examples are given
below.
A group of 40 mice was irradiated with germicidal lamps at
weekly doses of 3 x 104 J m-2. By 52 weeks, 97% of mice had
developed skin tumours, the majority of which were squamous cell
carcinomas (Lill, 1983). In another study, groups of 24 hairless
albino mice were irradiated daily at 230, 1460 or 7000 J m-2. The
prevalence of tumour-bearing mice (with mainly squamous cell
carcinomas) increased with time to over 75% in each exposure group
(Sterenborg et al., 1988). A comparison with other studies showing
response to exposure to UVB led the authors to conclude that the
small amounts of UVB emitted by the germicidal lamps could not
explain the response observed. Interaction between UVC and UVB could
not be excluded. Yields of keratoacanthoma like skin tumours,
proportional to dose, were observed in small groups of rats exposed
to radiation from Westinghouse G36T6L germicidal lamps (Strickland
et al., 1979).
UVA
A number of studies have been conducted in which skin tumours
have been produced in hairless mice by UVA under conditions in which
the exclusion of UVB from the exposure was adequately documented
(IARC, 1992). For example, groups of 24 albino hairless mice were
exposed to UVA from Philips TL40W/09 fluorescent tubes filtered
through 10mm of glass, highly absorbent for UVB, for 12 h/day, seven
days a week, for one year. The daily dose was 220 kJ m-2. Skin
tumours appeared in all animals and histopathological examination of
the larger lesions showed the majority to be SCC. On the basis of
the known dose-response to UVB, it was estimated that 100,000 times
more UVB than was residually present in the exposure would have been
necessary to produce the incidence of tumours observed (van Weelden
et al., 1986, 1988). Similar findings were obtained in a study in
which UVA at > 340 nm was produced by passing radiation from
Philips HPA 400 W lamps through liquid filters (Sterenborg and van
der Leun, 1990).
7.1.3 Interactions between radiations of different wavelengths
Simple photoaddition is commonly assumed for the interaction of
different wavelengths of UV in producing cancer. That is, exposure
at each wavelength contributes to the effective dose in an additive
way. Several studies, however, suggest that the true position is
more complex and the subject has been reviewed in detail by van der
Leun (1987, 1992).
For exposures to UV at different wavelength ranges administered
simultaneously, or in close temporal proximity, both reductions and
increases in the carcinogenic effect have been reported by
comparison with what would have been expected on the basis of simple
addition. Following detailed review, these effects were described as
"nonexistent, unproven or small" (IARC, 1992) and this conclusion is
consistent with the results of a recent study (Berg et al., 1993).
Such interactions play only a small role in the evaluation of risks
of UV (Health Council of the Netherlands, 1986).
There is a well-established protective effect of visible light
against UV carcinogenesis (and other effects) in M. domestica
(Ley, 1993) and lower animals (see, for example, Setlow et al.,
1993) which possesses the photoreactivating enzyme, photolyase.
Whether or not humans possess a DNA photolyase or show
photoreactivation is controversial (Ley, 1993, Li et al., 1993).
Several experiments, with somewhat conflicting results, have
been carried out in which exposures to UV to one wavelength range
have been separated in time from exposures to another (Forbes et
al., 1978; Staberg et al., 1983; Bech-Thomsen et al., 1988a,
b; Slaper, 1987). On review of these studies, it was concluded that
the combined effects tend to be slightly less than what would be
expected from simple photoaddition (IARC, 1992).
7.1.4 Dose-response
The accurate and quantitative description of the relationship
between UV and the occurrence of skin tumours, usually SCC, has been
based on experiments in which mice have been exposed regularly,
usually daily, to UV from standard sources. In most relevant
experiments, a UV dose, usually much lower than that of the outdoors
environment, was delivered daily or several times per week until
skin tumours developed. The UVB dose which induces tumours in mice
is lower than for acute reactions such as erythema or oedema and up
to 33 times lower in one experiment which produced an abundance of
skin cancers (De Gruijl et al., 1983). The higher the dose given,
the less time it takes for tumours to appear. In most experiments,
the time taken for 50% of mice to develop tumours has ranged between
a few months and one year but can be brought down to as low as 18
days (IARC, 1992; Willis et al., 1981).
Quantitative dose-effect relationships have been derived for
mice exposed regularly (usually daily) to UV. The median time to
first tumour (tm) has been used as the measure of effect.
Dose-effect relationships of the following form have been proposed
(IARC, 1992):
tm = k1 D-r
or, equivalently,
log tm = -r log D + log k1 .
In these expressions, k1 is a constant representing both the
susceptibility of the mouse strain and the effectiveness of the
radiation spectrum administered, D is the daily dose of radiation
and r is a numerical exponent giving the slope of the dose response
curve. Estimates of r vary between 0.5 and 0.6 in most experiments,
with broad-spectrum UV and broadband UVB; the value of 0.5 is
typical for large tumours, and 0.6 for small tumours (Blum et al.,
1959; de Gruiji, 1983). In one experiment with UVC a value r=0.2 was
found (Sterenborg, 1988). The relationship found by de Gruijl et
al. (1983) for the induction of skin tumours less than 1 mm in
diameter by UVB in albino hairless mice is shown in figure 7.1; the
value of r in this relationship is 0.6.
A corresponding expression for the dose-response relationship
in albino hairless mice is given by:
Y = k2 Dc td
where Y is the average number of tumours per mouse (the yield), k2
is a constant, D is the daily dose of UV radiation, t is the number
of days of exposure to UV and c and d are numerical exponents
describing the dose response relationship (van der Leun & der
Gruijl, 1993). By describing the response in terms of a measure of
incidence of tumours rather than time to tumour development, this
expression may be more useful for risk assessment in humans.
7.1.5 Effect of pattern of exposure
Tumours can be induced by a single dose of UV (Hsu et al.,
1975; Strickland et al., 1979). In mice, this has required a dose
that caused ulceration of the skin which is known to be a tumour
promoter.
Two experiments have reported on the effects of pattern of UV
(mainly UVB) exposure in albino hairless mice. At constant
instantaneous intensity and constant weekly dose of UV, increasing
fractionation of the dose from once weekly to three times weekly to
five times weekly increased the incidence of tumours (Forbes et
al., 1981). Similarly, with a constant daily dose of UV, incidence
of tumours increased with extension of the period of delivery (and
corresponding reduction in instantaneous intensity) from 1.25 to 4
or 12 hours (Kelfkens et al., 1991); there was no appreciable
difference between exposures of 4 and 12 hours. Similar results had
been obtained by Forbes and Davies (cited in Kelfkens et al.,
1991).
7.1.6 Action spectrum
Ideally, to determine the action spectrum of UV carcinogenesis,
experiments would be carried out with monochromatic radiation to
determine the relationship between dose and median time to first
tumour for each wavelength. However, narrow-band monochromatic
sources suitable for experiments of these types are difficult to
achieve. An alternative approach is to conduct experiments with a
number of overlapping broad band sources and to derive an action
spectrum by appropriate mathematical analysis of the results.
Recently two large collections of data on carcinogenicity of UV
in albino hairless mice following exposure to multiple overlapping
sources in the UVC, UVB and UVA ranges, supplemented with results of
experiments using highly filtered UVA sources, have been combined to
produce what is probably the best estimate to date of the action
spectrum for skin carcinogenesis in any strain of animals (de Gruijl
et al., 1993). As far as possible, the end point in the
experimental results used was the appearance of SCC.
The action spectrum produced is shown in figure 7.2. The upper
and lower dashed ( - - ) curves result from a sensitivity analysis
showing the boundaries of the effects of displacements of 5 nm
segments of the action spectrum that do not increase the chi2 for
the fit of the spectrum to the data by more than 1.0. Effectiveness
rises as wavelength increases to a peak at about 295 nm, falls
steeply to an initial minimum at about 350 nm, rises again to 380 nm
and then appears to fall away sharply. The definition of this action
spectrum is very good in the UVB range, there is substantial
uncertainty in the UVA range and essentially no information below
280 nm except at 254 nm. Even with the uncertainty, if there is not
actually a second peak in the UVA range, there is at least a plateau
from about 340 to 380 nm. This action spectrum is very similar to
the action spectra for erythema in humans (McKinlay and Diffey,
1987) and the induction of oedema in the skin of mice (Cole et al.,
1986).
Recently, results have been reported on the action spectrum for
the production of melanomas in hybrid fish (Setlow et al., 1993).
Groups of between 20 and 124 fish were exposed to single doses, at
two to six exposure levels, of UV or visible light at wavelengths of
302 nm, 313 nm, 365 nm, 405 nm and 436 nm. Narrow band radiation was
produced by use of a grating monochromator and, for the higher
wavelengths, various filters to eliminate any radiation at
substantially lower wavelengths. The experiment was terminated after
four months and all fish examined for melanomas which occurred in 5%
to 24% of control fish and 24% to 45% of irradiated fish. The
estimated action spectrum obtained is shown in figure 7.3
superimposed on published action spectra for mammalian cell
mutagenicity and cytotoxicity. Relative to an effectiveness in
producing melanomas of 1.0 for UV at 302 nm, the effectiveness at
313 nm was 0.16 and that at 365 nm, 0.32; for visible light, the
relative effectiveness were 0.017 at 405 nm and 0.023 at 436 nm. The
effectiveness of UVA in producing melanoma in this model relative to
the effectiveness of 300 nm radiation was some 3 orders of magnitude
greater than the effectiveness of UVA, relative to the same
baseline, in producing SCC in hairless albino mice (figure 7.2; de
Gruijl et al., 1993). An action spectrum has not yet been
determined for production of melanoma in M. domestica.
A group of 460 hybrid fish of two strains developed by crossing
platyfish ( Xiphophorus maculatus) and swordtails ( Xiphophorus
helleri) were exposed for 5 to 20 days to mainly UVB from
Westinghouse FS40 sunlamps filtered through acetate sheets
transmitting > 290 nm (150 or 300 mJ m-2) and >340 nm (850 and
1700 J m-2). Between 19% and 40% of exposed fish developed
melanocytic tumours compared with 12% and 2% in control fish (Setlow
et al., 1989).
In an attempt to confirm a suspected association between
sunlight and cancer of the eye in cattle, four Hereford cattle were
exposed to mainly UVB from Westinghouse FS40 sunlamps. Visible
tumours developed in the eyes of three of them and one was confirmed
histopathologically as a preneoplastic growth (Kopecky et al.,
1979).
UVC
Experiments have been carried out in which animals have been
exposed to low-pressure mercury discharge germicidal lamps which
emit most of their radiation at 254 nm with weaker spectral lines in
the UVB, UVA and visible spectra (IARC, 1992). Examples are given
below.
A group of 40 mice was irradiated with germicidal lamps at
weekly doses of 3 x 104 J m-2. By 52 weeks, 97% of mice had
developed skin tumours, the majority of which were squamous cell
carcinomas (Lill, 1983). In another study, groups of 24 hairless
albino mice were irradiated daily at 230, 1460 or 7000 J m-2. The
prevalence of tumour-bearing mice (with mainly squamous cell
carcinomas) increased with time to over 75% in each exposure group
(Sterenborg et al., 1988). A comparison with other studies showing
response to exposure to UVB led the authors to conclude that the
small amounts of UVB emitted by the germicidal lamps could not
explain the response observed. Interaction between UVC and UVB could
not be excluded. Yields of keratoacanthoma like skin tumours,
proportional to dose, were observed in small groups of rats exposed
to radiation from Westinghouse G36T6L germicidal lamps (Strickland
et al., 1979).
UVA
A number of studies have been conducted in which skin tumours
have been produced in hairless mice by UVA under conditions in which
the exclusion of UVB from the exposure was adequately documented
(IARC, 1992). For example, groups of 24 albino hairless mice were
exposed to UVA from Philips TL40W/09 fluorescent tubes filtered
through 10mm of glass, highly absorbent for UVB, for 12 h/day, seven
days a week, for one year. The daily dose was 220 kJ m-2. Skin
tumours appeared in all animals and histopathological examination of
the larger lesions showed the majority to be SCC. On the basis of
the known dose-response to UVB, it was estimated that 100,000 times
more UVB than was residually present in the exposure would have been
necessary to produce the incidence of tumours observed (van Weelden
et al., 1986, 1988). Similar findings were obtained in a study in
which UVA at > 340 nm was produced by passing radiation from
Philips HPA 400 W lamps through liquid filters (Sterenborg and van
der Leun, 1990).
7.1.3 Interactions between radiations of different wavelengths
Simple photoaddition is commonly assumed for the interaction of
different wavelengths of UV in producing cancer. That is, exposure
at each wavelength contributes to the effective dose in an additive
way. Several studies, however, suggest that the true position is
more complex and the subject has been reviewed in detail by van der
Leun (1987, 1992).
For exposures to UV at different wavelength ranges administered
simultaneously, or in close temporal proximity, both reductions and
increases in the carcinogenic effect have been reported by
comparison with what would have been expected on the basis of simple
addition. Following detailed review, these effects were described as
"nonexistent, unproven or small" (IARC, 1992) and this conclusion is
consistent with the results of a recent study (Berg et al., 1993).
Such interactions play only a small role in the evaluation of risks
of UV (Health Council of the Netherlands, 1986).
There is a well-established protective effect of visible light
against UV carcinogenesis (and other effects) in M. domestica
(Ley, 1993) and lower animals (see, for example, Setlow et al.,
1993) which possesses the photoreactivating enzyme, photolyase.
Whether or not humans possess a DNA photolyase or show
photoreactivation is controversial (Ley, 1993, Li et al., 1993).
Several experiments, with somewhat conflicting results, have
been carried out in which exposures to UV to one wavelength range
have been separated in time from exposures to another (Forbes et
al., 1978; Staberg et al., 1983; Bech-Thomsen et al., 1988a,
b; Slaper, 1987). On review of these studies, it was concluded that
the combined effects tend to be slightly less than what would be
expected from simple photoaddition (IARC, 1992).
7.1.4 Dose-response
The accurate and quantitative description of the relationship
between UV and the occurrence of skin tumours, usually SCC, has been
based on experiments in which mice have been exposed regularly,
usually daily, to UV from standard sources. In most relevant
experiments, a UV dose, usually much lower than that of the outdoors
environment, was delivered daily or several times per week until
skin tumours developed. The UVB dose which induces tumours in mice
is lower than for acute reactions such as erythema or oedema and up
to 33 times lower in one experiment which produced an abundance of
skin cancers (De Gruijl et al., 1983). The higher the dose given,
the less time it takes for tumours to appear. In most experiments,
the time taken for 50% of mice to develop tumours has ranged between
a few months and one year but can be brought down to as low as 18
days (IARC, 1992; Willis et al., 1981).
Quantitative dose-effect relationships have been derived for
mice exposed regularly (usually daily) to UV. The median time to
first tumour (tm) has been used as the measure of effect.
Dose-effect relationships of the following form have been proposed
(IARC, 1992):
tm = k1 D-r
or, equivalently,
log tm = -r log D + log k1 .
In these expressions, k1 is a constant representing both the
susceptibility of the mouse strain and the effectiveness of the
radiation spectrum administered, D is the daily dose of radiation
and r is a numerical exponent giving the slope of the dose response
curve. Estimates of r vary between 0.5 and 0.6 in most experiments,
with broad-spectrum UV and broadband UVB; the value of 0.5 is
typical for large tumours, and 0.6 for small tumours (Blum et al.,
1959; de Gruiji, 1983). In one experiment with UVC a value r=0.2 was
found (Sterenborg, 1988). The relationship found by de Gruijl et
al. (1983) for the induction of skin tumours less than 1 mm in
diameter by UVB in albino hairless mice is shown in figure 7.1; the
value of r in this relationship is 0.6.
A corresponding expression for the dose-response relationship
in albino hairless mice is given by:
Y = k2 Dc td
where Y is the average number of tumours per mouse (the yield), k2
is a constant, D is the daily dose of UV radiation, t is the number
of days of exposure to UV and c and d are numerical exponents
describing the dose response relationship (van der Leun & der
Gruijl, 1993). By describing the response in terms of a measure of
incidence of tumours rather than time to tumour development, this
expression may be more useful for risk assessment in humans.
7.1.5 Effect of pattern of exposure
Tumours can be induced by a single dose of UV (Hsu et al.,
1975; Strickland et al., 1979). In mice, this has required a dose
that caused ulceration of the skin which is known to be a tumour
promoter.
Two experiments have reported on the effects of pattern of UV
(mainly UVB) exposure in albino hairless mice. At constant
instantaneous intensity and constant weekly dose of UV, increasing
fractionation of the dose from once weekly to three times weekly to
five times weekly increased the incidence of tumours (Forbes et
al., 1981). Similarly, with a constant daily dose of UV, incidence
of tumours increased with extension of the period of delivery (and
corresponding reduction in instantaneous intensity) from 1.25 to 4
or 12 hours (Kelfkens et al., 1991); there was no appreciable
difference between exposures of 4 and 12 hours. Similar results had
been obtained by Forbes and Davies (cited in Kelfkens et al.,
1991).
7.1.6 Action spectrum
Ideally, to determine the action spectrum of UV carcinogenesis,
experiments would be carried out with monochromatic radiation to
determine the relationship between dose and median time to first
tumour for each wavelength. However, narrow-band monochromatic
sources suitable for experiments of these types are difficult to
achieve. An alternative approach is to conduct experiments with a
number of overlapping broad band sources and to derive an action
spectrum by appropriate mathematical analysis of the results.
Recently two large collections of data on carcinogenicity of UV
in albino hairless mice following exposure to multiple overlapping
sources in the UVC, UVB and UVA ranges, supplemented with results of
experiments using highly filtered UVA sources, have been combined to
produce what is probably the best estimate to date of the action
spectrum for skin carcinogenesis in any strain of animals (de Gruijl
et al., 1993). As far as possible, the end point in the
experimental results used was the appearance of SCC.
The action spectrum produced is shown in figure 7.2. The upper
and lower dashed ( - - ) curves result from a sensitivity analysis
showing the boundaries of the effects of displacements of 5 nm
segments of the action spectrum that do not increase the chi2 for
the fit of the spectrum to the data by more than 1.0. Effectiveness
rises as wavelength increases to a peak at about 295 nm, falls
steeply to an initial minimum at about 350 nm, rises again to 380 nm
and then appears to fall away sharply. The definition of this action
spectrum is very good in the UVB range, there is substantial
uncertainty in the UVA range and essentially no information below
280 nm except at 254 nm. Even with the uncertainty, if there is not
actually a second peak in the UVA range, there is at least a plateau
from about 340 to 380 nm. This action spectrum is very similar to
the action spectra for erythema in humans (McKinlay and Diffey,
1987) and the induction of oedema in the skin of mice (Cole et al.,
1986).
Recently, results have been reported on the action spectrum for
the production of melanomas in hybrid fish (Setlow et al., 1993).
Groups of between 20 and 124 fish were exposed to single doses, at
two to six exposure levels, of UV or visible light at wavelengths of
302 nm, 313 nm, 365 nm, 405 nm and 436 nm. Narrow band radiation was
produced by use of a grating monochromator and, for the higher
wavelengths, various filters to eliminate any radiation at
substantially lower wavelengths. The experiment was terminated after
four months and all fish examined for melanomas which occurred in 5%
to 24% of control fish and 24% to 45% of irradiated fish. The
estimated action spectrum obtained is shown in figure 7.3
superimposed on published action spectra for mammalian cell
mutagenicity and cytotoxicity. Relative to an effectiveness in
producing melanomas of 1.0 for UV at 302 nm, the effectiveness at
313 nm was 0.16 and that at 365 nm, 0.32; for visible light, the
relative effectiveness were 0.017 at 405 nm and 0.023 at 436 nm. The
effectiveness of UVA in producing melanoma in this model relative to
the effectiveness of 300 nm radiation was some 3 orders of magnitude
greater than the effectiveness of UVA, relative to the same
baseline, in producing SCC in hairless albino mice (figure 7.2; de
Gruijl et al., 1993). An action spectrum has not yet been
determined for production of melanoma in M. domestica.
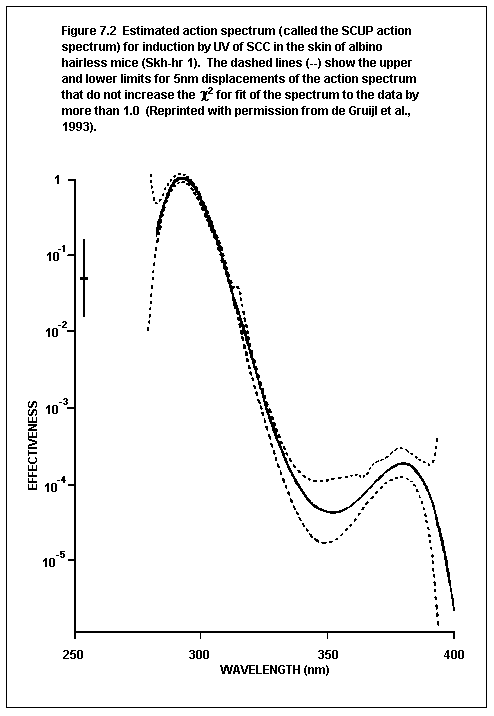
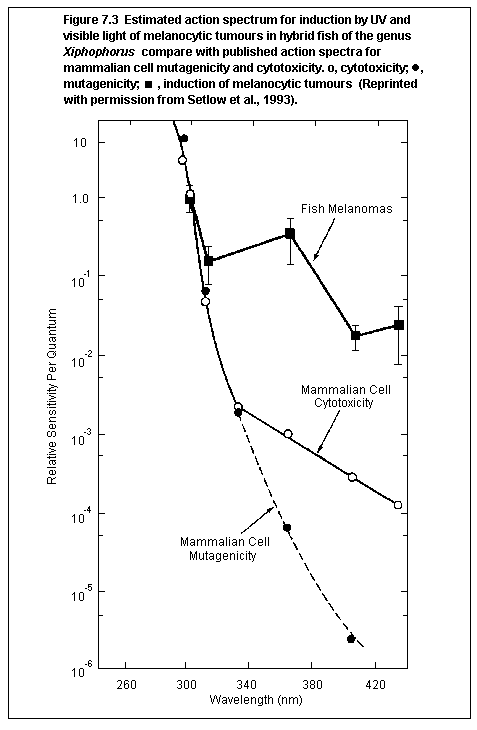 7.1.7 Interaction between UV and chemicals
Interaction with chemical carcinogens
A number of studies have been carried out in which UV has been
administered before or after administration of a known chemical
carcinogen. A period of irradiation with UVB before application of
3,4-Benzo[a]pyrene to the skin of mice increased the carcinogenic
response to high-dose 3,4-Benzo[a]pyrene (Gensler & Bowden, 1987;
Gensler, 1988). A number of studies in which mice were irradiated
with UV after application of 7,12-Dimethylbenz[a]anthracene (DMBA)
to the skin showed an increase in tumour production over that
produced by DMBA alone (Epstein & Epstein, 1962; Epstein, 1965;
Reeve et al., 1990; Husain et al., 1991). Of particular interest
in this regard is the production of melanoma-like lesions in mice by
the combination of DMBA and UV. In a number of experiments, DMBA
alone has produced benign pigmented naevi. Subsequent administration
of UVB, UVB + UVA, and UVA alone caused these naevi to grow into
lesions that had the appearance of malignant melanoma; these lesions
did not develop in mice treated with DMBA alone (Epstein, 1965;
Epstein et al., 1967; Husain et al., 1991). Irradiation with UVB
from Westinghouse FS40 sunlamps before application of DMBA and the
promoter 12-O-tetradecanoylphorbol 13-acetate (TPA) reduced the
incidence and number of tumours produced in CDF1 mice both when
the site of subsequent application of the chemicals was irradiated
and when a distant site was irradiated (Gensler and Welch, 1992).
This effect, therefore, appeared to be mediated systemically.
Both croton oil and TPA, applied to skin after irradiation with
UV, have been shown to increase the carcinogenic response to UV
(Epstein & Roth, 1968; Pound, 1970; Stenbäck, 1975b; Strickland et
al., 1985). Of two other, suspected cancer promoters, one, methyl
ethyl ketone peroxide increased tumour production (Logani et al.,
1984) and the other, benzoyl peroxide, did not (Epstein, 1988;
Iversen, 1988).
Interaction with other chemicals
In several experiments, mice exposed to either solar- simulated
radiation or UVA and topical application of 5- methoxypsoralen
showed higher incidences of skin papillomas or SCC than did mice
exposed to UV or 5-methoxypsoralen alone (Zadjela & Bisagni, 1981;
Cartwright & Walter, 1983; Young et al., 1983). Methoxsalen
(8-methoxypsoralen) shows the same effects when given with UV (IARC,
1980).
7.1.8 Mechanisms of UV carcinogenesis
DNA damage
There is abundant evidence that UV causes DNA damage, both by
direct photochemical effects (for example, the production of
cyclobutylpyrimidine dimers and 6-4 photoproduct) and by oxidative
effects leading to DNA strand breakage and DNA-protein cross-linkage
(see Chapter 6).
There is evidence that UV-induced DNA damage, and particularly
the formation of pyrimidine dimers, is one step in the mechanism
whereby UV can cause cancer in experimental animals. First, Hart et
al. (1977) showed, in the clonal fish, Poecilia formosa, that
the transplantation of UVC irradiated thyroid cells from one fish to
another induced thyroid tumours in the host fish. If irradiation of
the thyroid of the donor fish was followed immediately by
application of photoreactivating light (which monomerizes the
pyrimidine dimers), the development of thyroid tumours was almost
completely prevented. Second, and similarly, Ley et al. (1991)
showed that exposure to photoreactivating light, after exposure to
UV, delayed and reduced the yield of skin tumours produced in M.
domestica by irradiation with Westinghouse FS40 sunlamps. The same
result was obtained for corneal tumours but, unexpectedly, exposure
to photoreactivating light was as effective in reducing
carcinogenesis when given immediately before UV as when given after.
There is evidence that photoreactivating light may also inhibit the
production by UV of melanocytic lesions in the skin of M. domestica
(Ley et al., 1989). Third, Yarosh et al. (1992) applied
liposomes containing T4 endonuclease V, an enzyme that specifically
repairs cyclobutyl pyrimidine dimers, to the skin of albino hairless
mice, three times a week, after they had been irradiated with
Westinghouse FS20 sunlamps. The incidence of skin tumours observed
in the mice fell in proportion to the quantity of active liposomes
applied.
There is also evidence that activated oncogenes and a mutated
tumour-suppressor gene are present in some skin cancers induced
experimentally by UV. Husain et al. (1990) reported studies of
expression of the cHa-ras oncogene in three papillomas and three
carcinomas from among lesions that developed in 17 of 90 Sencar mice
following a single exposure, at 7 x 104 J m-2, to UVB from
Westinghouse FS20 sunlamps. RNA preparations from all six tumours
showed elevated levels of Cha- ras-specific messenger RNA
sequences, suggesting overexpression of this oncogene. DNA from the
carcinomas, but not the papillomas, was able to induce
transformation of NIH-3T3 cells which demonstrated overexpression
and amplification of the Cha-ras oncogene. Subsequently, in their
studies of melanocytic tumours produced by DMBA and UV in hairless
mice (Skh-hr 2), Husain et al. (1991) found mutations in codon 61
of the N- ras oncogene in three of eight precursor naevi and one
melanoma. The base transitions, however, were not of a type to
suggest that they had been caused by UV. Kress et al. (1992)
screened exons 5 to 8 of the p53 tumour-suppressor gene for mutation
in 35 epidermal tumours induced in four strains of mice by UV from
Westinghouse FS40 sunlamps. Mutations were found in seven tumours.
All mutations occurred at dipyrimidine sequences. C -> T and CC ->
TT transitions were present in five of the seven tumours with
mutations, strongly suggesting that these mutations were due to UV.
Oxidative processes
Oxidative processes in the skin may also mediate the
carcinogenicity of UV, particularly UVA (Morlière et al., 1992),
in skin, either by direct oxidative damage to DNA or by way of
potentially carcinogenic intermediates, such as cholesterol-5,6-
epoxide (Black, 1987; Morin et al., 1991). A number of studies
have shown that relative increases in dietary polyunsaturated fatty
acids, which may lead to lipid peroxidation, can increase the tumour
response to exposure to UV in experimental animals and that this
effect can be inhibited by the simultaneous feeding of antioxidants
(Reeve et al., 1988; Black et al., 1992). In addition, the
feeding of antioxidants, such as tocopherol, can reduce the tumour
response to UV in the absence of any manipulation of dietary fat
(Black & Chan, 1975; Gerrish & Gensler, 1993).
Immune-suppression
UV-induced immune-suppression appears to play a major role in
UV carcinogenesis in mice. A review of the literature on the
association between UV-induced immune suppression and increased
susceptibility to tumours may be found in section 7.3.2.
There is evidence also that administration of immunosuppressive
drugs will increase the carcinogenic response to UV in experimental
animals (IARC, 1992). Experiments have been carried out with
anti-lymphocyte serum, azathioprine, cyclophosphamide, cyclosporine
and 6-mercaptopurine. Consistent increases in the carcinogenic
response to UV over two or more experiments were seen with
azathioprine and cyclosporine (Reeve et al., 1985; Nelson et al.,
1987; Servilla et al., 1987; Kelly et al., 1987, 1989).
7.1.9 Conclusions
From a review of the animal studies one can conclude the
following about UV exposure of the skin:
Carcinogenesis by sunlight is a widespread phenomenon among
domestic and food animals. Most tumours found are squamous cell
carcinomas (SCC). In experimental animals skin cancer is mainly
caused by UV radiation. Again, most tumours induced are SCC. For the
induction of SCC in albino hairless mice, the effectiveness is a
function of wavelength:
- the effectiveness is found to peak in the UVB range;
- UVA is also carcinogenic, but at a much lower level of
effectiveness - similar to what is found for erythema and
tanning;
- the effectiveness in the UVC range is unknown, except for one
wavelength, 254 nm; at that wavelength the effectiveness is
lower than that in the UVB peak
- it is still unknown if there is any effectiveness of visible
light.
For the induction of SCC in hairless mice by daily exposures to
UVB the dose effect relationship was found to be a power function.
It showed no indication of a threshold dose, even at doses as low as
3% of that required for an acute reaction, such as oedema.
Radiations of different wavelengths can cooperate in the induction
of SCC.
Melanomas are much less common among animals. Only two animal
models have been found where exposure to UV induced melanomas:
- the South American opossum M. domestica, and
- a hybrid fish, derived from the swordtail and platyfish.
An initial action spectrum was determined for the hybrid fish.
It peaks in the UVB range but also shows a high effectiveness in the
UVA. An action spectrum for the induction of melanoma in M.
domestica has not been determined.
Basal cell carcinomas are uncommon in animals. Suitable animal
models for the induction of BCC are not available.
7.2 Immune Responses
7.2.1 Immune function assays
The immune system depicted in figure 7.4 is composed of primary
lymphoid organs (bone marrow and thymus), secondary lymphoid organs
including spleen and lymph nodes, and several cell types. In
addition a number of mediators including cytokines, antibodies and
complement regulate and/or are produced by the immune system. A
number of studies have shown that exposure to UV suppresses contact
hypersensitivity (CHS) and other delayed-type hypersensitivity (DTH)
responses of the immune system. Both of these responses provide a
measure the competence of T lymphocytes, suppression of which would
be expected to compromise host resistance to infectious agents such
as viruses and mycobacterium. Unless otherwise indicated animal
studies have utilized FS sunlamps (see Chapter 3 for spectral
characterization) and exposed shaved skin.
Two types of experiments have been described in the literature.
In experiments designed to investigate local immune suppression,
mice were exposed to UV and a hapten chemical such as
dinitrofluorobenzene (DNFB) was applied to the site of irradiation.
After an incubation period of several days, animals were challenged
by painting the chemical on the ear, and the immune response was
assessed by measuring ear thickness, which is an expression of CHS.
Mice exposed (usually for 4 successive days prior to chemical
application) to doses of UV, which might cause a minimal erythemal
response in an untanned human, failed to show ear swelling (Toews
et al., 1980; Noonan & De Fabo, 1990; Jeevan et al., 1992a). The
failure to respond appeared to be due to the development of
suppressor T cells which rendered the mouse tolerant to the
particular antigen. This concept was supported by the observation
that the same mice could not be sensitized with the same chemical
through unirradiated skin 14 days later. Moreover, antigen-specific
unresponsiveness could be adoptively transferred to naive mice by
spleen and lymph node cells obtained from mice skin-painted with
DNFB through UV-irradiated skin (Elmets et al., 1983). It should
be noted that the precise nature of the activity of these suppressor
cells remains the subject of some debate.
In a second type of experiment the effects of UV on systemic
immune responses were demonstrated by irradiating mice at one site
and subsequently (3-5 days later) exposing them to a contact
allergen (CHS) or injecting a protein antigen (DTH) at an
unirradiated site. These mice failed to respond when subsequently
challenged on the ear (CHS) or in the footpad (DTH) with the same
antigen (Noonan et al., 1981a, Kim et al., 1990; Ullrich et
al., 1986a; Ullrich, 1986; Jeevan & Kripke, 1990; Howie et al.,
1986; Denkins et al., 1989; Giannini, 1986a,b). Again
unresponsiveness was attributed to the development of antigen
specific "suppressor T cells" (Noonan et al., 1981b; Ullrich
1985). These experiments suggest that immune suppression could occur
even when the site of entry for the antigen was not the same as the
site of UV exposure.
7.1.7 Interaction between UV and chemicals
Interaction with chemical carcinogens
A number of studies have been carried out in which UV has been
administered before or after administration of a known chemical
carcinogen. A period of irradiation with UVB before application of
3,4-Benzo[a]pyrene to the skin of mice increased the carcinogenic
response to high-dose 3,4-Benzo[a]pyrene (Gensler & Bowden, 1987;
Gensler, 1988). A number of studies in which mice were irradiated
with UV after application of 7,12-Dimethylbenz[a]anthracene (DMBA)
to the skin showed an increase in tumour production over that
produced by DMBA alone (Epstein & Epstein, 1962; Epstein, 1965;
Reeve et al., 1990; Husain et al., 1991). Of particular interest
in this regard is the production of melanoma-like lesions in mice by
the combination of DMBA and UV. In a number of experiments, DMBA
alone has produced benign pigmented naevi. Subsequent administration
of UVB, UVB + UVA, and UVA alone caused these naevi to grow into
lesions that had the appearance of malignant melanoma; these lesions
did not develop in mice treated with DMBA alone (Epstein, 1965;
Epstein et al., 1967; Husain et al., 1991). Irradiation with UVB
from Westinghouse FS40 sunlamps before application of DMBA and the
promoter 12-O-tetradecanoylphorbol 13-acetate (TPA) reduced the
incidence and number of tumours produced in CDF1 mice both when
the site of subsequent application of the chemicals was irradiated
and when a distant site was irradiated (Gensler and Welch, 1992).
This effect, therefore, appeared to be mediated systemically.
Both croton oil and TPA, applied to skin after irradiation with
UV, have been shown to increase the carcinogenic response to UV
(Epstein & Roth, 1968; Pound, 1970; Stenbäck, 1975b; Strickland et
al., 1985). Of two other, suspected cancer promoters, one, methyl
ethyl ketone peroxide increased tumour production (Logani et al.,
1984) and the other, benzoyl peroxide, did not (Epstein, 1988;
Iversen, 1988).
Interaction with other chemicals
In several experiments, mice exposed to either solar- simulated
radiation or UVA and topical application of 5- methoxypsoralen
showed higher incidences of skin papillomas or SCC than did mice
exposed to UV or 5-methoxypsoralen alone (Zadjela & Bisagni, 1981;
Cartwright & Walter, 1983; Young et al., 1983). Methoxsalen
(8-methoxypsoralen) shows the same effects when given with UV (IARC,
1980).
7.1.8 Mechanisms of UV carcinogenesis
DNA damage
There is abundant evidence that UV causes DNA damage, both by
direct photochemical effects (for example, the production of
cyclobutylpyrimidine dimers and 6-4 photoproduct) and by oxidative
effects leading to DNA strand breakage and DNA-protein cross-linkage
(see Chapter 6).
There is evidence that UV-induced DNA damage, and particularly
the formation of pyrimidine dimers, is one step in the mechanism
whereby UV can cause cancer in experimental animals. First, Hart et
al. (1977) showed, in the clonal fish, Poecilia formosa, that
the transplantation of UVC irradiated thyroid cells from one fish to
another induced thyroid tumours in the host fish. If irradiation of
the thyroid of the donor fish was followed immediately by
application of photoreactivating light (which monomerizes the
pyrimidine dimers), the development of thyroid tumours was almost
completely prevented. Second, and similarly, Ley et al. (1991)
showed that exposure to photoreactivating light, after exposure to
UV, delayed and reduced the yield of skin tumours produced in M.
domestica by irradiation with Westinghouse FS40 sunlamps. The same
result was obtained for corneal tumours but, unexpectedly, exposure
to photoreactivating light was as effective in reducing
carcinogenesis when given immediately before UV as when given after.
There is evidence that photoreactivating light may also inhibit the
production by UV of melanocytic lesions in the skin of M. domestica
(Ley et al., 1989). Third, Yarosh et al. (1992) applied
liposomes containing T4 endonuclease V, an enzyme that specifically
repairs cyclobutyl pyrimidine dimers, to the skin of albino hairless
mice, three times a week, after they had been irradiated with
Westinghouse FS20 sunlamps. The incidence of skin tumours observed
in the mice fell in proportion to the quantity of active liposomes
applied.
There is also evidence that activated oncogenes and a mutated
tumour-suppressor gene are present in some skin cancers induced
experimentally by UV. Husain et al. (1990) reported studies of
expression of the cHa-ras oncogene in three papillomas and three
carcinomas from among lesions that developed in 17 of 90 Sencar mice
following a single exposure, at 7 x 104 J m-2, to UVB from
Westinghouse FS20 sunlamps. RNA preparations from all six tumours
showed elevated levels of Cha- ras-specific messenger RNA
sequences, suggesting overexpression of this oncogene. DNA from the
carcinomas, but not the papillomas, was able to induce
transformation of NIH-3T3 cells which demonstrated overexpression
and amplification of the Cha-ras oncogene. Subsequently, in their
studies of melanocytic tumours produced by DMBA and UV in hairless
mice (Skh-hr 2), Husain et al. (1991) found mutations in codon 61
of the N- ras oncogene in three of eight precursor naevi and one
melanoma. The base transitions, however, were not of a type to
suggest that they had been caused by UV. Kress et al. (1992)
screened exons 5 to 8 of the p53 tumour-suppressor gene for mutation
in 35 epidermal tumours induced in four strains of mice by UV from
Westinghouse FS40 sunlamps. Mutations were found in seven tumours.
All mutations occurred at dipyrimidine sequences. C -> T and CC ->
TT transitions were present in five of the seven tumours with
mutations, strongly suggesting that these mutations were due to UV.
Oxidative processes
Oxidative processes in the skin may also mediate the
carcinogenicity of UV, particularly UVA (Morlière et al., 1992),
in skin, either by direct oxidative damage to DNA or by way of
potentially carcinogenic intermediates, such as cholesterol-5,6-
epoxide (Black, 1987; Morin et al., 1991). A number of studies
have shown that relative increases in dietary polyunsaturated fatty
acids, which may lead to lipid peroxidation, can increase the tumour
response to exposure to UV in experimental animals and that this
effect can be inhibited by the simultaneous feeding of antioxidants
(Reeve et al., 1988; Black et al., 1992). In addition, the
feeding of antioxidants, such as tocopherol, can reduce the tumour
response to UV in the absence of any manipulation of dietary fat
(Black & Chan, 1975; Gerrish & Gensler, 1993).
Immune-suppression
UV-induced immune-suppression appears to play a major role in
UV carcinogenesis in mice. A review of the literature on the
association between UV-induced immune suppression and increased
susceptibility to tumours may be found in section 7.3.2.
There is evidence also that administration of immunosuppressive
drugs will increase the carcinogenic response to UV in experimental
animals (IARC, 1992). Experiments have been carried out with
anti-lymphocyte serum, azathioprine, cyclophosphamide, cyclosporine
and 6-mercaptopurine. Consistent increases in the carcinogenic
response to UV over two or more experiments were seen with
azathioprine and cyclosporine (Reeve et al., 1985; Nelson et al.,
1987; Servilla et al., 1987; Kelly et al., 1987, 1989).
7.1.9 Conclusions
From a review of the animal studies one can conclude the
following about UV exposure of the skin:
Carcinogenesis by sunlight is a widespread phenomenon among
domestic and food animals. Most tumours found are squamous cell
carcinomas (SCC). In experimental animals skin cancer is mainly
caused by UV radiation. Again, most tumours induced are SCC. For the
induction of SCC in albino hairless mice, the effectiveness is a
function of wavelength:
- the effectiveness is found to peak in the UVB range;
- UVA is also carcinogenic, but at a much lower level of
effectiveness - similar to what is found for erythema and
tanning;
- the effectiveness in the UVC range is unknown, except for one
wavelength, 254 nm; at that wavelength the effectiveness is
lower than that in the UVB peak
- it is still unknown if there is any effectiveness of visible
light.
For the induction of SCC in hairless mice by daily exposures to
UVB the dose effect relationship was found to be a power function.
It showed no indication of a threshold dose, even at doses as low as
3% of that required for an acute reaction, such as oedema.
Radiations of different wavelengths can cooperate in the induction
of SCC.
Melanomas are much less common among animals. Only two animal
models have been found where exposure to UV induced melanomas:
- the South American opossum M. domestica, and
- a hybrid fish, derived from the swordtail and platyfish.
An initial action spectrum was determined for the hybrid fish.
It peaks in the UVB range but also shows a high effectiveness in the
UVA. An action spectrum for the induction of melanoma in M.
domestica has not been determined.
Basal cell carcinomas are uncommon in animals. Suitable animal
models for the induction of BCC are not available.
7.2 Immune Responses
7.2.1 Immune function assays
The immune system depicted in figure 7.4 is composed of primary
lymphoid organs (bone marrow and thymus), secondary lymphoid organs
including spleen and lymph nodes, and several cell types. In
addition a number of mediators including cytokines, antibodies and
complement regulate and/or are produced by the immune system. A
number of studies have shown that exposure to UV suppresses contact
hypersensitivity (CHS) and other delayed-type hypersensitivity (DTH)
responses of the immune system. Both of these responses provide a
measure the competence of T lymphocytes, suppression of which would
be expected to compromise host resistance to infectious agents such
as viruses and mycobacterium. Unless otherwise indicated animal
studies have utilized FS sunlamps (see Chapter 3 for spectral
characterization) and exposed shaved skin.
Two types of experiments have been described in the literature.
In experiments designed to investigate local immune suppression,
mice were exposed to UV and a hapten chemical such as
dinitrofluorobenzene (DNFB) was applied to the site of irradiation.
After an incubation period of several days, animals were challenged
by painting the chemical on the ear, and the immune response was
assessed by measuring ear thickness, which is an expression of CHS.
Mice exposed (usually for 4 successive days prior to chemical
application) to doses of UV, which might cause a minimal erythemal
response in an untanned human, failed to show ear swelling (Toews
et al., 1980; Noonan & De Fabo, 1990; Jeevan et al., 1992a). The
failure to respond appeared to be due to the development of
suppressor T cells which rendered the mouse tolerant to the
particular antigen. This concept was supported by the observation
that the same mice could not be sensitized with the same chemical
through unirradiated skin 14 days later. Moreover, antigen-specific
unresponsiveness could be adoptively transferred to naive mice by
spleen and lymph node cells obtained from mice skin-painted with
DNFB through UV-irradiated skin (Elmets et al., 1983). It should
be noted that the precise nature of the activity of these suppressor
cells remains the subject of some debate.
In a second type of experiment the effects of UV on systemic
immune responses were demonstrated by irradiating mice at one site
and subsequently (3-5 days later) exposing them to a contact
allergen (CHS) or injecting a protein antigen (DTH) at an
unirradiated site. These mice failed to respond when subsequently
challenged on the ear (CHS) or in the footpad (DTH) with the same
antigen (Noonan et al., 1981a, Kim et al., 1990; Ullrich et
al., 1986a; Ullrich, 1986; Jeevan & Kripke, 1990; Howie et al.,
1986; Denkins et al., 1989; Giannini, 1986a,b). Again
unresponsiveness was attributed to the development of antigen
specific "suppressor T cells" (Noonan et al., 1981b; Ullrich
1985). These experiments suggest that immune suppression could occur
even when the site of entry for the antigen was not the same as the
site of UV exposure.
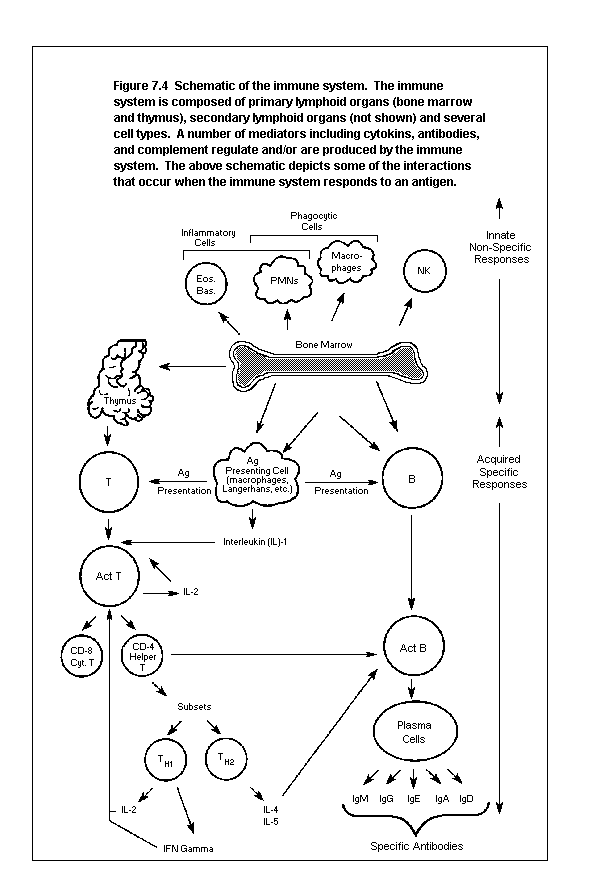 Noonan et al. (1981a) reported a 50% suppression of CHS in
BALB/c mice after a single dose of 2 KJ m-2 UV. She also reported
that there was no difference in the dose response curves for
UV-induced local and systemic immunosuppression of CHS responses;
however, the dose of UV required to suppress CHS in C57BL/6 mice was
6.4 times less than that required to produce similar immune
suppression in BALB/c mice (Noonan & De Fabo, 1990). Also, while
local suppression could be demonstrated in mice sensitized
immediately after irradiation, systemic suppression occurred only if
the sensitizer was applied 3 or more days after radiation (Noonan &
De Fabo, 1990).
Fifty percent suppression of CHS and DTH has been reported at
doses ranging from 2.3 kJ m-2 to 20 kJ m-2, (Kim et al., 1990;
Jeevan & Kripke, 1990; Jeevan et al., 1992b) levels which produce
minimal or no edema. The dose needed to produce 50% suppression
varies depending on the strain of mouse and type of antigen used,
and on the laboratory producing the data. Suppression of
virus-specific DTH was observed in mice exposed to a suberythemal
dose of UVB 3-7 days prior to infection with Herpes simplex virus
(HSV) but not 14 days and persisted for at least 3 months after
irradiation (Howie et al., 1986). Elimination of wavelengths below
315 nm with a mylar filter either eliminated or greatly reduced the
suppression of CHS following exposure to UV indicating that most if
not all the suppression is due to UVB (Noonan et al., 1981b;
Noonan and De Fabo, 1990). Using narrow bands of UV at 10
wavelengths from 250 to 320 nm, it was reported that maximum
suppression of CHS occurred between 260 and 270nm, there was a
shoulder in the action spectrum from 280-290 nm and then a steady
decline to 3% of maximum at 320 nm (De Fabo and Noonan, 1983).
There are two subpopulations of T helper cells, designated Th1
and Th2 which appear to be differentially affected by UV exposure.
These two populations are thought to regulate different sets of
immune responses. Th1 cells produce interleukin (IL) 2 and gamma
interferon (gamma IFN) as well as other cytokines, promote
delayed-type hypersensitivity (cell mediated, type IV) responses
such as CHS, provide help for certain antibody subtype responses
including complement-fixing antibodies, activate macrophages, and
may be particularly important for dealing with antigens expressed on
cell surfaces, such as viral and tumour antigens (Coffman et al.,
1988). Th2 cells produce a different array of cytokines including
IL-4 and IL-5 which promotes antibody responses. Only Th2 cells can
stimulate a primary IgE response which is mediated by IL-4 and
inhibited by gamma IFN (Coffman et al., 1988). Thus, Th2 cells may
be particularly important in responding to certain parasitic
infections, and also play an important role in immediate-type
hypersensitivity including reactions to common allergens such as
pollen and dust mite.
Recent studies have suggested that both local and systemic
immune suppression induced by UV may be the result of an inability
to present antigen to and hence activate Th1 cells (Simon et al.,
1990, 1991; Araneo et al., 1989).
Suppression of systemic CHS and DTH responses similar to that
observed following UV exposure have been obtained by injecting mice
with supernatant from UV-irradiated keratinocytes (Kim et al.,
1990; Rivas & Ullrich, 1992; Jeevan et al., 1992c). Among factors
derived from keratinocytes, the cytokines tumour necrosis factor
alpha (TNF alpha) (Yoshikawa & Streilein, 1990; Vincek et al.,
1993) and IL-10 (Rivas & Ullrich, 1992) are thought to be important
in UV-induced immunosuppression. Also, cis-urocanic acid, the
product of UV isomerization of urocanic acid located in the stratum
corneum, when injected subcutaneously or painted on the epidermis,
produces immunosuppressive effects which mimic UVB-induced
immunosuppression (Ross et al., 1986), and the action spectrum of
UV-induced immune suppression closely follows the absorption
spectrum of urocanic acid (De Fabo and Noonan, 1983). In all three
cases it is thought that these mediators act by modifying antigen
presentation (Vermeer & Streilein, 1990; Rivas & Ullrich, 1992;
Noonan et al., 1988).
In summary, it appears that UVB causes the release of mediators
from the skin which alter the antigen presenting capability of
Langerhans cells as well as antigen presenting cells at other sites,
resulting in the development of "suppressor T-cells". It may be that
these suppressor T cells are Th2 cells. The net result is the
failure to activate Th1 cells and suppression of DTH responses
thought to play an important role in host defences against certain
types of tumours and microbial infections. The immune suppression is
antigen specific, (i.e. only responses to antigens administered
within 7 days after irradiation are affected) and is long lasting
(at least 3 months).
In addition to DTH and CHS responses, UV affects several other
immune functions frequently included in the standard protocols for
immunotoxicity testing. Spleen cells taken from UV-exposed mice
(single exposure, 54 kJ m-2) failed to respond in a mixed
lymphocyte response assay in vitro (Ullrich, 1985), and spleen
cells from tumour-implanted UV-treated (10 kJ m-2, 3 times/wk, for
3 months) mice were not cytotoxic in vitro against UV-induced
tumours (Fisher & Kripke, 1977). In contrast, exposure to
approximately 2-3 kJ m-2 day-1, for 23 days did not affect
responses of spleen or inguinal lymph node cells to the mitogens,
phytohaemagglutinin, concanavalin A or bacterial lipopolysaccharide.
Similarly, lymphocyte responses to these mitogens and peritoneal
macrophage phagocytic and tumouricidal activities were unaffected by
exposure to 10 kJ m-2, 3 times per week, for up to 6 months
(Funnell & Keast 1985; Norbury et al., 1977).
Kripke et al., (1977) reported no effect of UV (in mice
exposed to 10 kJ m-2, 3 times per week for up to 6 months) on the
primary haemagglutonin antibody response to sheep red blood cells;
however in another study, the IgM and IgG plaque forming cell
responses of lymph node cells to sheep red blood cells given
intradermally (through irradiated skin) were suppressed (Funnell &
Keast, 1985). Also, suppressor cells generated by irradiating mice
with a single exposure of 30-40 kJ m-2, 5 days prior to
sensitization with trinitrochlorobenzene prevented the development
of hapten specific antibody-forming cells when injected
intravenously along with hapten conjugated sheep red blood cells
into syngeneic recipients (Ullrich et al., 1986b).
UV exposure enhanced poly I.C augmented natural killer cell
(NK) activity in mice (Lynch & Daynes, 1983); however, suppression
of NK activity in rats has been reported following exposure to
suberythemal doses of UV (Garssen et al., 1993). Hence, in
laboratory animals UV exposure appears to affect mixed lymphocyte
responses, cytotoxic T cell activity, possibly NK activity, and in
some circumstances antibody responses. With the exception of NK
activity all of these responses are antigen specific responses. In
contrast UV does not appear to alter several non-specific responses
including macrophage phagocytic or tumouricidal activities, or T or
B cell responses to mitogens. However, unlike CHS and DTH, none of
these responses has been studied in detail in rodents, and dose
response information is not available.
7.2.2 Susceptibility to tumours
Interest in the immunosuppressive properties of UVB was first
sparked by the observations of Kripke and associates that UV
exposure induced highly antigenic tumours which did not grow when
transplanted into syngeneic, immunocompetent mice but did grow when
transplanted into immunosuppressed or UV treated mice (Kripke 1974;
Kripke & Fisher, 1976). The precise nature of the tumour antigens
has not been defined. Mice treated with 1.8 kJ m-2, 3 times per
week, for 3 months were unable to reject UV-induced syngeneic
tumours when challenged subcutaneously at any time from 2 weeks into
the irradiation exposure regimen to as late as 5 months after the
end of UV treatment (Kripke & Fisher, 1976). Hence susceptibility to
tumour challenge was detectable well before the appearance of
primary skin cancers induced by the UV and persisted long after
exposure ended. Similarly, more UV-induced fibrosarcoma tumour
colonies were detected in the lungs following i.v injection of
tumour cells in mice treated with 7 kJ m-2, 3 times per week, for
5 weeks (Kripke & Fidler 1980) as compared to unirradiated controls.
Susceptibility to tumour challenge was directly proportional to dose
of UV and a dose fractionated over time was no more effective than
the same total dose given as a single treatment (Kripke & Fidler
1980; De Fabo and Kripke 1979).
In BALB/cAnN mice 50% tumour incidence following subcutaneous
tumour challenge at an unirradiated site occurred at a dose of
approximately 40 kJ m-2. Enhanced tumour incidence was observed in
mice exposed to 21.6 kJ m-2 as much as 32 weeks before tumour
challenge. A number of studies (Fisher & Kripke, 1977; Daynes &
Spellman, 1977; Daynes et al., 1977; Spellman & Daynes, 1978;
Spellman et al., 1977; Ullrich & Kripke, 1984, de Gruijl and van
der Leun 1982, 1983) have demonstrated that susceptibility to
challenge with UV-induced tumours following UV exposure, like
suppression of CHS and DTH, is mediated by antigen specific
"suppressor T cells" which are present in spleen and lymph nodes of
UV-irradiated mice. As with suppression of DTH and CHS responses,
enhanced susceptibility to UV-induced tumours is antigen specific
and long lasting.
While it is very clear that UV increases the susceptibility of
mice to UV-induced tumours, much less work has been done on the
effects of UV on susceptibility to other types of tumours. UV
treated mice that were unable to reject syngeneic UV- induced
tumours were able to reject 2 types of non-UV-induced tumours, B16
melanoma tumours and a spontaneous mouse leukaemia tumour (Kripke
et al., 1977). Roberts and Daynes (1980) reported that mice
irradiated with subcarcinogenic doses of UV for 3 weeks prior to
treatment with either benz[a]pyrene or methycholanthrene at an
unirradiated site had reduced latency periods for the development of
these chemically-induced tumours. Also, chemically induced tumours
from these UV-treated mice appeared to be more antigenic than those
induced in untreated mice in that they were incapable of progressive
growth when transplanted into normal (immunocompetent) syngeneic
mice but were capable of progressive growth in UV-treated mice. The
authors suggested that decreased selective pressures exerted by the
host (due to UV-induced suppressor cells) at the time of tumour
induction allowed more immunogenic tumours to emerge and progress.
7.2.3 Susceptibility to infectious disease
Several types of infectious disease models have been developed
to study the effects of UV exposure: 1) the infectious agent was
injected through irradiated skin, 2) exposure to the infectious
agent occurred at a site distant from the site of irradiation, or 3)
in the case of some herpes simplex virus (HSV) studies, exposure to
UV occurred after infection. The first exposure scenario is
representative of vector-borne infections and some vaccinations
while the second exposure regimen is representative of infections
which do not necessarily enter the host via the skin. The third
scenario was designed to mimic reactivation of HSV in humans
following sun exposure. In all cases the focus has been on microbial
agents that are controlled, at least in part, by DTH responses.
An example of the first type of model is that of Leishmania
major infection in mice (Giannini 1986a, 1986b, 1987, 1992). Mice
were irradiated only on the tail with suberythemal doses (0.06-6 kJ
m-2), 3 times/week, for 1 month and were infected intradermally
through the irradiated surface of the tail 24 hr after the first UV
exposure. In UV-treated mice the DTH response to L. major antigens
2 and 6 weeks post infection was suppressed, the number of organisms
recovered from skin at the injection site was (Giannini, 1986a) and,
a larger number of parasites was observed in the draining lymph node
(Giannini, 1987, 1992). Finally, mice which were infected through
irradiated skin failed to develop protective immunity such that
lesions following reinfection at an unirradiated site were
significantly larger when compared to lesions of previously infected
but unirradiated mice (Giannini, 1986b).
In a similar model mice treated with 13 or 33 kJ m-2 for 4
consecutive days and infected intradermally with HSV at the site of
irradiation had a higher incidence of zosteriform lesions than
unirradiated mice, and at the higher exposure level 100% mortality
was observed as opposed to no mortality in unirradiated mice
(Yasumoto et al., 1987). A decreased DTH response to viral antigen
was observed and appeared to result from the induction of suppressor
T cells (Yasumoto et al., 1987, Aurelian et al., 1988). Also,
following in vitro exposure to UV, a defect in the ability of
Langerhans cells to present HSV antigen to lymphocytes was
demonstrated (Hayashi & Aurelian, 1986).
In contrast, no effect of UV on either parasite specific DTH
responses or recovery of parasite from internal organs was observed
following percutaneous injection of Schistosoma mansoni through
UV-irradiated (0.4 kJ m-2, 4 consecutive days) skin (Jeevan et
al., 1992a). In this same study, suppression of microbe specific
DTH responses were not observed following intradermal injection of
Mycobacterium bovis (BCG) or subcutaneous injection of Candida
albicans at the site of irradiation; however, the number of viable
mycobacteria recovered from the lymphoid organs of BCG-infected mice
was increased significantly in the UV treated mice for a period of
more than 2 months post infection.
BCG has also been used as an infectious disease model to
illustrate systemic effects of UV immunosuppression. In this model
mice were irradiated on the back and injected with BCG
subcutaneously in the footpad. Mice exposed from 1 to 15 times (3
times/week for up to 5 weeks) to one minimal erythemal dose (2.25 kJ
m-2) showed significant suppression in their DTH response to
tuberculin (PPD) and increased numbers of live bacteria in the
spleen and lymph node compared to unirradiated controls (Jeevan &
Kripke, 1990). However, when exposures were continued beyond 5
weeks, the DTH response recovered and mice challenged with bacteria
at that point did not exhibit increased numbers of organisms in the
spleen and lymph node, suggesting that eventually some sort of
adaptation to exposure occurs. Significant suppression of DTH was
observed in mice which received a single dose as low as 1.4 kJ m-2
3 days prior to infection and significant increases in bacteria in
spleen and lymph node were observed in mice which received as little
as 0.7 kJ m-2. Similar effects were observed when supernatants
from keratinocyte cultures exposed to UV in vitro were injected
intravenously 3 days prior to infection (Jeevan et al., 1992c).
In a similar model, mice treated with a single high UV dose (45
kJ m-2) 3 days before infection with Mycobacterium lepraemurium
exhibited significant suppression of DTH responses to mycobacterial
antigen 3 and 6 months after infection and had significantly more
bacteria in the infected footpad, lymph node, and spleen 3-6 months
post infection (Jeevan et al., 1992b). This high dose also reduced
the median survival time in mice infected i.v. With a lower exposure
dose (2.3 kJ m-2) 50% suppression of the DTH response to
mycobacterial antigen was observed 3 months post infection, and
increased numbers of bacteria were observed in the footpad, spleen
and lymph node of mice exposed to UV doses greater than or equal to
5.6 kJ m-2. When mice were treated with 2.25 kJ m-2, 3
times/week from 3-15 times, the DTH response to M. lepraemurium
was suppressed, but as with BCG, the DTH response was normal in mice
which received more than 15 exposures.
Effects of systemic UV-induced immune suppression have been
demonstrated in mice infected with HSV, 3 days after irradiation
with 1 kJ m-2, at a site distant from the site of infection. In
this model suppression of DTH was mediated by modulation of
epidermal antigen presenting cells and the development of suppressor
T cells (Howie et al., 1986, 1987). Subcutaneous injection or
epidermal application of UV-treated urocanic acid produced similar
immune suppression (Ross et al., 1986). Effects of UV on the
actual progression of disease in this model have not been reported.
While most of the work to date has been done in mice, recent
work in the rat suggests that suberythemal doses of UV also suppress
immune responses. In addition rats exposed to suberythemal doses of
UV had higher levels of microorganisms in target organs following
oral infection with Trichinella spiralis or intraperitoneal
infection with cytomegalovirus (Garssen et al., 1993).
In summary, exposure to suberythemal doses of UV has been shown
to exacerbate a variety of infections in rodent models. Both
infections which are initiated at the site of UV exposure and
infections initiated at distant sites have been affected. While most
of the infections test, have evolved in some manner more recent work
indicates that systemic infections without skin involvement may also
be affected. Enhanced susceptibility appears to result from
suppression of T-helper-1 cell activity. The mechanisms associated
with this suppression appear to be the same as those identified in
association with suppression to CHS responses.
Reactivation of latent HSV infections can be induced in mouse
and guinea pig by exposure of the previously infected site to sub-
or minimal erythemal doses of UV. However, the role that immune
suppression plays in this reactivation has not been established
(Blyth et al., 1976; Norval et al., 1987; Laycock et al.,
1991).
7.2.4 Susceptibility to immunologically-mediated
diseases
Several studies have suggested that stimulation of Th2 cells
may remain intact or be enhanced in UV-exposed mice (Simon et al.,
1990; 1991; Araneo et al., 1989). Since Th2 cells are the only
cells that can stimulate a primary IgE response (Coffman et al.,
1988), it is possible that UV exposure may increase the risk of
immediate-type hypersensitivity. This possibility has not yet been
studied, but deserves attention in light of the increasing incidence
of morbidity and mortality due to asthma, which is often triggered
by allergic responses (NIH, 1991; Danielle, 1988).
Acceleration of autoimmunity following UV exposure has also
been reported in mice. Both acute (2 hr/day, for 7 days) and chronic
(3 hr/wk, for 4 wks) exposure to 20 J m-2 s-1 enhanced mortality
in an autoimmune strain of mice (NZB X BZW F1). These exposures
also caused increased serum antibodies to single stranded DNA,
enhanced polyclonal B-cell activity in the spleen, and caused more
severe renal glomerular inflammatory changes (Ansel et al., 1985),
all hallmarks of autoimmunity. It is unclear whether there is a
relationship between these reactions and the effects of UV on
immunological processes described above.
7.2.5 Conclusions
Exposure of mice to UV radiation impairs certain immune
responses, enhances susceptibility to a variety of infectious agents
and UV-induced tumours. While fewer studies have been done in rats,
similar effects have been observed. Suppression of contact and
delayed type hypersensitivity responses have been studied most
extensively; however, several other cell mediated responses are also
affected. Suppression of these immune responses appears to be
mediated by release of soluble mediators from UB exposed skin which
alter antigen presentation by Langerhans (and other) cells such that
they fail to activate TH 1 cells. The resulting immune suppression
is antigen specific, can occur regardless of whether antigen is
applied at the site of exposure or not, and is relatively long
lasting.
Enhanced susceptibility to a variety of infections in mice and
rats corresponds to suppression of DTH responses to microbe specific
antigens. UV exposure also prevents the development of protective
immunity to these infections.
7.3 Ocular Studies
7.3.1 Introduction
The large number of animal studies enhances our understanding
of both acute and delayed ocular effect from UV exposure (Zigman
1993; Andley 1987; Dillon et al., 1990). Studies of the action
spectrum of delayed or permanent effects such as cataract or
retinopathy and UV damage mechanisms in the cornea, lens and retina
are only possible in animal models.
7.3.2 General effects
Photokeratitis is an acute reversible radiation-induced injury
of the corneal epithelium. It is analogous to acute sunburn of the
skin. Animal studies have clearly demonstrated that exposure to an
artificial source of UVB can lead to acute photokeratitis, and
action spectra have been determined (Cogan & Kinsey, 1946; Pitts et
al., 1977; Zuclich and Kurtin, 1977). Pitts (1974; 1978) estimated
the mean threshold of UVB (290-315 nm) for photokeratitis was 35 J
m-2 at 270 nm.
Damage to corneal endothelium has been reported in rabbits
(Doughty & Cullen, 1990). The effects on the corneal stroma have
also been observed in rabbits, where reversible damage to the
stromal keratocytes has followed exposure to UV (Ringvold &
Davangar, 1985). This histological study showed that keratocytes
disappear following UV exposure and later reappear. More recent
animal studies have reported the use of a UVB filtering contact lens
or the application of UVB absorbing chromophores to the cornea of
rabbit eyes protects against photokeratitis (Bergmanson et al.,
1988; Oldenburg et al., 1990).
7.3.3 Cataractogenesis
There is substantial evidence that exposure to UV induces
discrete opacities in the anterior lens of experimental animals.
Several studies have shown that UVB, but not UVA, induces anterior
opacities in animals (Bachem, 1956; Pitts et al., 1977; Jose &
Pitts, 1985; Söderberg 1990). Supra threshold doses of UVB at 300 nm
cause a Na-K shift between the lens and the surrounding (Söderberg
(1991). This leads to swelling and disruption of lens cells,
probably causing zones of deviating refractive index within the lens
observed as opacities (Söderberg 1989). Anterior lens opacities have
developed in albino mice after daily exposure for 1-2 months to
mixed UVA and UVB source (290-400 nm), but not when the source was
filtered to remove radiation <320 nm (Jose & Pitts, 1985). Anterior
cataract have also been produced in young albino mice exposed to
black light (predominantly UVA, with some UVB) (Zigman et al.,
1974). With more prolonged exposure of mice, cortical and posterior
subcapsular opacities have been induced (Zigman et al., 1975;
Jose, 1986). By contrast, a dose of 2 kJ m-2 applied every day
produced anterior polar cataract within 8 weeks. Extension of the
irradiation to 5 months created an opacity in the deeper cortex
(Weager et al., 1989). In testing possible cataractogenic efficacy
of UVA and UVB, UVB alone was cataractogenic, whereas, UVA showed
the same effect only when other cataractogenic factors were added
(e.g. X-ray) (Schmitt et al., 1988).
Ham et al. (1989) irradiated rhesus monkeys to 1 mW cm-2
daily for three years to UVA radiation (UVB radiation was carefully
excluded) and was unable to detect lenticular opacities.
7.3.4 Retinal effects
It is well accepted that short wavelength visible non-coherent
and coherent light, as well as UVA, can cause photochemical,
mechanical and thermal damage to the retina and the pigment
epithelium (review: Ham et al., 1984; Organisciak & Winkler,
1993). Aphakic monkeys revealed retinal lesions after exposure to
UVA (Guerry et al., 1985). Exposure of aphakic albino rats to UVA
created acute retinal lesions with 50-80% more effectiveness than
blue light (Rapp & Smith 1992). Mechanical and thermal lesions are
almost entirely limited to exposure to high energy laser light.
Photochemical damage can occur through two different light exposure
regimes: short term, high irradiance (minutes to hours) and
long-term (days to weeks) low irradiance (Kremers & van Norren,
1988; Remé et al., 1991; Rapp & Smith, 1992). Whereas damage
thresholds differ depending on the animal species, basic mechanisms
at the molecular and structural level are essentially the same (for
review: see Organisciak and Winkler, 1993). Photochemical lesions
are strongly dependent on wavelength, i.e. the absorbing chromophore
and its quantum efficiency, obey, at least within a certain time
frame, the reciprocity of intensity and exposure duration (e.g.
Kremers & van Norren, 1988).
Action spectra for high irradiance short exposure duration
lesions have been obtained in several studies (e.g. Ham et al.,
1984). Action spectra for prolonged exposure to relatively low light
levels are less clear and were first evaluated by Noell et al.
(1966). Prolonged exposure to low light levels may elicit secondary
tissue responses that may not be directly related to the initial
action spectrum. These tissues responses include acute cellular
necrosis and apoptotic cell death, an inflammatory response with
oedematous changes and macrophage invasion, and tissue proliferation
with scar formation and neovascularization (e.g. Hoppeler et al.,
1988; Yoshida et al., 1993). Action spectra, tissue responses,
preventive measures as well as enhancing factors, which are all
intensively investigated in many laboratories, are likely to be
similar to the human.
Studies of photochemical retinal injury in aphakic rhesus
monkeys have extended the action spectrum for short-wavelength light
damage down to 310 nm (Ham et al., 1984). These studies showed
that the retina, if exposed, is up to six times more vulnerable to
photochemical damage in the UV than in the visible. These results
are therefore of direct relevance to pseudophakic individuals
without a UV-protective lens, and may also indicate a contributory
role to light damage from the small fraction of UV that reaches the
retina of the normal eye.
8. HUMAN STUDIES: THE SKIN
8.1 Characteristics
8.1.1 Structure and optical properties
The skin is a large organ with an area of more than 1.5 m2 in
adults. It provides the first stage of protection for chemicals,
radiations, xenobiotics and also prevents the evaporation of water
and the loss of ions and proteins. The skin has developed specific
mechanisms for photoprotection and biological responses to UV as
discussed below.
Skin is composed of three very different parts (as shown in
figure 4.1): the epidermis, dermis and subcutaneous tissue. The
epidermis is the outermost layer of the skin and varies in thickness
from 50 µm to 600 µm (palmoplantor skin). The fibrous proteins known
as keratin are produced in the keratinocytes of the epidermis. It is
keratin that serves as a major protective tough substance in the
skin; hair and nails are composed almost entirely of keratin. The
daughter cells of the keratinocytes in the basal layer (stratum
basale) of the epidermis differentiate and become "prickle cells" of
the malpighian layer of the epidermis - the stratum malpighii. As
these cells migrate outward, changes continue to occur; granules
appear in the cytoplasm of each cell; the cells tend to flatten and
form the stratum granulosum. Still later, the cells lose their
nuclei, die, dehydrate and flatten out to form the tough stratum
corneum (horny layer). It is generally agreed that the entire
process of cell migration from the basal layer of the epidermis to
final shedding from the surface of the stratum corneum takes 28 days
in normal skin. Of this 28-day period, the cell spends about 14 days
in the epidermis and 14 days in the stratum corneum.
The skin contains millions of tiny glands, including: apocrine
(sweat) glands which discharge sweat into hair follicles, eccrine
(sweat) glands which carry saline from the dermis and subcutaneous
layer directly to the skin surface, and sebaceous glands which
secrete sebum - an oily substance which lubricates hairshafts and
maintains a slightly acidic, oily film over the stratum corneum. The
collection of eccrine sweat glands plays a major role in the body's
thermoregulatory mechanism, since evaporative cooling is the most
efficient means the body has for removing excess heat, as long as
the humidity is not 100%. More than two litres of sweat can be
discharged in one day. On average, the skin contains about as many
sebaceous glands as eccrine sweat glands, except that there are few
sebaceous glands on the palms of the hands and soles of the feet.
The dermis, or corium, is much thicker than the epidermis, but
consists of much larger cells. The dermis is largely connective
tissue which gives the skin its elasticity and supportive strength.
Nerve cells, blood vessels and lymphatic glands are found in the
outermost dermal layer - the papillary dermis. Unlike the epidermis,
the thickness of the dermis is not at all uniform throughout the
body; it varies from 1 mm to 4 mm.
The basal layer of the epidermis is delineated from the dermis
by a complex basement membrane. The keratinocytes are anchored to
the basement membrane by semi desmosomes. This is the only layer
where cell division takes place. The division of keratinocytes,
under normal conditions, occurs every 17 to 38 days. The division
rate may vary in different parts of the body.
Melanocytes are formed in the basal layer of the epidermis
loosely attached to the neighbouring keratinocytes (ratio 1
melanocyte for 46 keratinocytes). The melanocytes synthesize the
pigment melanin which matures into melanosomes. Then the melanosomes
are transferred to the keratinocytes where they are digested if
their size is below 1 cubic micron. Large melanosomes, as in dark
skin, are not digested but transferred intact to the stratum corneum
with the normal shedding of the epidermis.
Melanocytes are cells of nervous origin which have migrated
during the 10th week of embryonic life in the epidermis and in the
hair roots. These cells divide very slowly (one division every 3-5
years). There is a boost of division when new hairs are growing or
after exposure to UV light.
Chemically, melanins are bipolymers: red melanins contain
sulphurs and are soluble at PH 7.2, black melanins are insoluble.
The ratio of red and black melanins within a melanocyte is
genetically determined. Mixed types of melanins are deposited on a
protein matrix contained in the melanosomes. Both types absorb UV
and participate in the screening effect of the whole epidermis. Two
major steps in the melanin synthesis take place: the enzymatic
(tyrosinase) oxidation of the tyrosine to produce dopa
(di-hydroxyphenylalanine) and the spontaneous oxidation of dopa in
quinones.
Black melanin absorbs UV. In the presence of oxygen, it
produces a free radical which, in sufficient quantities, can be
deleterious to the melanocytes and the cellular environment,
including keratinocytes or in the dermis, fibroblasts and fibres.
A third type of cell is present in the upper layers of the
stratum Malpighi: the Langerhans cell. It is a migrating dendritic
cell which is able to recognise foreign or abnormal structures. This
cell plays a major role in immunological recognition and its
activity is very sensitive to UV. Its function is impaired by a UV
dose as low as one half of a minimal erythema dose.
The innermost layer of the skin is generally known as the
subcutaneous layer. It is composed largely of fatty tissue that
serves a shock-absorbing and insulating role. The thickness of this
layer varies considerably from one body region to another and from
one person to another.
Skin optics are governed by two basic processes, the absorption
and scattering of light. Absorption is the loss of a photon when its
energy is reduced within the atom or molecule, and more specifically
in a target species called a chromophore. UV absorption occurs in
wavelength bands where the molecules have characteristic absorption
spectra. The absorbed photon energy is dissipated as heat or
reemission of light when the excited molecules return to their
ground state, or the energy is spent on photochemical reactions.
Scattering is a process where the direction of propagation of UV is
altered, especially at boundaries of refractive index. Scattering
and absorption of the photons limit the depth of penetration of UV
in the skin (see section 4.2 and figure 4.1).
In the white skin, the change in the refractive index causes 5%
of the normally incident light to be reflected. The remaining 95% is
absorbed or reflected at the dermis. The back-scattered light is
reflected again at different interfaces. This phenomenon explains
why, in the epidermis and in the upper papillary dermis, the
strength of the light becomes several times that of the incident
light itself.
The transmission of UV through isolated epidermis is strongly
dependent on the chromophores contained in the structure. Aromatic
amino acids (tryptophan, tyrosin, phenyl-alanine) absorb strongly
near 275 nm, urocanic acid and melanins play the role of endogenous
sunscreens. The DNA absorbs 260 nm wavelengths. Melanin is the
unique chromophore with an absorption extending into the UVA and
visible regions.
8.1.2 Skin types
The sensitivity of skin to UV has been defined by six
phototypes: types I to IV are characteristics of caucasoid
populations; type V represents mongoloid Middle Eastern populations
(Fitzpatrick et al., 1974); and type VI represents African and
American negroid populations. The capacities to acquire natural tan
or to present naturally a deep pigmentation are keys to the response
to UV exposure. Among caucasians, there is a general correlation
between skin type and resistance to sunburn and capacity to tan.
8.2 Beneficial Effects
8.2.1 Vitamin D3
An established beneficial effect of UV exposure is the
synthesis of vitamin D3 (Adams et al., 1982). In adults, the
epidermis contains nearly 50% of the total concentration of
7-dihydrocholesterol in the skin. UVB exposure causes the
provitamins D3 to be isomerized to pre-vitamin D3. During continual
exposure to sunlight, the pre-vitamin D3 forms by
photo-isomerization the biologically inert photo-products lumisterol
and tachysterol. Once formed, the pre-vitamin D3 spontaneously
isomerizes into vitamin D3 (reaction with a maximum efficacy at
37°C), a more stable form. After crossing the basal membrane of the
epidermis, vitamin D3 is linked to a circulating alpha1-globulin:
vitamin D-binding protein. The protein linked vitamin D3 is
transformed to 25-OH-T3 which can be measured in the blood.
Transported to the kidney, it is metabolised to 1,25 dihydroxy
vitamin D3 which is the biologically active form.
Vitamin D3 is required for the intestinal absoprtion of calcium
(Davies 1985). After a single whole body exposure to one MED the
circulating vitamin D3 level increases by an order of magnitude (2
ng/ml to 24 ng/ml within 24 hours) and returns to normal levels
within a week (Holick 1985). Since active vitamin D3 is metabolised
by the kidney, it was expected that in the circulating blood it does
not change dramatically after repeated solar exposures. Chronically
reduced vitamin D3 synthesis may lead to a deficit in active vitamin
D3, as was found in elderly subjects (Omdahl et al., 1982).
The use of sunscreens was found to suppress cutaneous vitamin
D3 synthesis (Matsuoka et al., 1988). This was also found in
children, pregnant or lactating women and debilitated patients with
poor intestinal absorption. Widespread use of sunscreens could lead
to vitamin D deficiency in some groups and inadequate fixation of
calcium (Prystowsky 1988). More recently, receptors for the active
form for vitamin D3 were found in the keratinocytes of the
epidermis. Vitamin D3 inhibits the proliferation of cultured
keratinocytes and induces them to terminally differentiate (Smith
et al., 1986). The topical or oral administration of 1, 25-OH-D3
has proved to be effective for the treatment of psoriasis, in
replacement or in addition to the classical PUVA treatment. This is
a new approach for the treatment of this condition and a possible
explanation for the success of the heliotherapy (Morimoto and
Kunihiko, 1989).
For the entire system of vitamin D3 production the amount of UV
radiation reaching the skin is critical. The doses needed are small,
and daily exposures of the face and hands to sun and light for 15
minutes is considered sufficient. The minimum dose requirement was
estimated to be equivalent to 55 MED per year (Health Council of the
Netherlands, 1986). When too little UVB reaches the skin,
deficiencies of vitamin D may occur, resulting in a weakening of the
bones. Groups at risk are particularly dark-skinned children in high
latitude cities and elderly people living entirely indoors.
Supplementation of vitamin D3 in the diet is then recommended.
8.2.2 Skin adaptation
Another beneficial effect of modest exposure to UVB radiation
is the maintenance of the ability of the skin to sustain further UV
exposures. Loss of this adaptation forms an important component in
photodermatosis, skin diseases where the lesions are caused by
light. These patients can be treated by regular exposures to UVB.
The doses required are in the same range as that needed for the
synthesis of vitamin D3.
8.2.3 Other benefits
It has been suggested that beneficial effects of UV exposures
may occur such as: improvement of cardiopathy and functions, and
better microorganism defense. These effects have not been confirmed
in well designed studies. Bright light therapy for winter depression
is most likely to be the consequence of visible light stimulating
the ocular system. Because of the high illumination needed for this
treatment special care is needed to avoid the emission of UV light
by these sources (Terman et al., 1990).
Treatment of portwine stains and pigment dyschromia by lasers
are applications of selective absorption of some wavelengths by
specific chromophores contained in the lesion. Phototherapy by
specific dyes absorbed by tumour cells or specific structures, is
currently a growing field.
8.3 Acute Effects
8.3.1 Erythema and sunburn
In its mildest form, sunburn consists of a reddening of the
skin (erythema) that appears up to about 8 h after exposure to UV
and gradually fades after a few days. In its most severe form, it
results in inflammation, blistering, and peeling of the skin. The
degree to which a person will experience sunburn depends critically
on skin type. For fair-skinned people, the relative effectiveness of
UV for tanning and for erythema are approximately the same over the
entire UVB and UVA ranges of wavelengths (Parrish et al., 1982).
The most important factors that define if a dose of UV will
induce erythema are the wavelength of the radiation, the skin type,
and the pigmentation of the subject. UVA, UVB and UVC are all able
to induce the erythema.
The relative effectiveness of the different wavelengths to
induce erythema is expressed as an erythema action spectrum
(McKinlay and Diffey 1987). For minimal erythema, the most
erythemogenic wavelengths are in the 250-290 nm range and a decrease
in effectiveness is observed as the wavelengths increase. Erythema
occurs 3-5 hours after UV exposure and reaches a maximum intensity
between 8 and 24 hours, fading over 3 days. The vasodilatation of
capillary vessels within the papillary dermis can be observed before
the erythema becomes visible, and occurs in the same way for
children, adults and elderly. However, the exposure time required to
produce UVB erythema increases after about 60 years of age.
Histologic alterations from erythema are observed in the
photodyskeratotic keratinocytes as well as intercellular edema with
exocytosis (lymphocytes in the epidermis). Superficial vascular
plexus, endothelial cell enlargement, perivenular edema, red blood
cells in the capillary are characteristic features observed between
3 hours and 72 hours after UV exposure. Dermal neutrophils appear
immediately after irradiation reaching a peak level at 24 hours.
DNA may be the primary chromophore involved in the induction of
erythema (Ley, 1985, Wolf et al., 1993). Subsequently, a number of
inflammatory mediators are induced. Cutaneous blisters have been
used extensively to study these mediators. Prostaglandin (E2 and
F2) levels were elevated in these blisters within 6 hours, peaked
at 24 hours and returned to control levels by 48 hours after UV
exposure, although erythema persisted beyond that time. Indomethacin
suppressed prostaglandin formation, however, blood flow was only
slightly altered by this treatment suggesting that other factors
must play an important role in UVB induced inflammation (Greaves et
al., 1978). Similarly while elevated histamine levels have been
observed, antihistamines have little effect in diminishing UV
induced erythema (Gilchrest et al., 1981).
Finally, UV exposure causes keratinocytes, to release the
cytokines interleukin-1 (IL-1) and tumour necrosis factor alpha (TNF
alpha) both of which are potent mediators of inflammation (Oxholm
et al., 1988; Rasanen et al., 1989). UVB exposure also causes
up-regulation of adhesion molecule such as ELAM-1 which facilitates
inflammatory cell infiltration (Murphy et al., 1991).
Irradiation of human skin with 3 MED is associated with
increased levels of transforming growth factor, suggesting a role of
this molecule in keratinocyte proliferation, epidermal hyperplasia
and angiogenesis.
8.3.2 Skin pigmentation and tanning
When skin is exposed to UV, two distinct tanning reactions
ensue. Immediate pigment darkening (IPD) begins immediately on
exposure to UV and is caused by the darkening of the pigment melanin
that is already present in the skin; it is normally seen only in
people who have at least a moderate constitutive tan. Such
pigmentation begins to fade within a few hours after cessation of
exposure. UVA is regarded as being most effective for IPD.
Delayed tanning (melanogenesis) takes about three days to
develop and is more effectively produced by UVB than by UVA (Parrish
et al., 1982; Gange et al., 1985). Delayed tanning is more
persistent than IPD and results from an increase in the number, size
and pigmentation of melanin granules. Exposure to UVB results also
in an increase in the thickness and scattering properties of the
epidermis (outer layer of the skin). Because UVA does not produce
thickening of the epidermis, the tan obtained from it, while perhaps
cosmetically acceptable, is not as effective in protecting against
further exposure to UV as the equivalent pigmentation induced by
exposure to UVB or solar radiation.
8.3.3 Photosensitization
The use of certain medicines may produce a photosensitizing
effect on exposure to UVA as may the topical application of certain
products, including some perfumes, body lotions, etc. Many
medications and other agents contain ingredients that may cause
photosensitivity, which is defined as a chemically induced change
that makes an individual unusually sensitive to light. An individual
who has been photosensitized may develop a rash, sunburn, or other
adverse effect from exposure to light of an intensity or duration
that would normally not affect that individual.
Reactions to photosensitizing agents involve both photoallergy
(allergic reaction of the skin) and phototoxicity (irritation of the
skin) after exposure to ultraviolet radiation from natural sunlight
or artificial lighting (particularly from tanning booths). This
photosensitization of the skin may be caused by creams or ointments
applied to the skin, by medications taken orally or by injection, or
by the use of prescription inhalers.
In addition to an exaggerated skin burn, itching, scaling,
rash, or swelling, exposure to UV in combination with certain
medications may result in (FDA 1992); Skin cancer, Premature skin
aging, Skin and eye burns, Allergic reactions, Cataracts, Reduced
immunity and Blood vessel damage.
This can result in photoallergic or phototoxic reactions that
are accelerated by UV exposure. Phototoxic contact dermatitis often
occurs clinically as exaggerated sunburn but occasionally blisters
may also occur on the erythematous areas. Most phototoxic
sensitizers have an action spectrum in the UV from 280-430 nm.
Window glass which absorbs UV below 320 nm will protect patients
from phototoxic compounds with absorption below 320 nm, but fails to
protect against photosensitizers such as tar and psoralens which are
efficient at longer wavelengths. Examples of photocontact drugs and
substances are given below in table 8.1.
Table 8.1 Some photosensitizing substances
(CIE, 1990, FDA, 1992)
Sulphonamides
Salicylanilides
Coal-Tar derivatives
- acridine
- anthracene
- phenanthrene
Dyes
- anthraquinone
- eosin
- methylene blue
- rose bengal
Psoralens
Some fragrances
Cyclamate (artificial sweetener)
Non-steroidal anti-inflammatory drugs (pain reliever,
antiathritics)
Deodorant and bacteriostatic agents in soaps
Fluorescent brightening agent for cellulose, nylon or wool
fibres
Phenothiazines (major tranquilizers, anti-emetics)
Sulfonylureas (oral anti-diabetics, hypoglycemics)
Sunscreen ingredients
- 6-Acetoxy-2,4,-dimethly-m-dioxane (preservative)
- Benzophenones
- Cinnamates
- Para-aminobenzoic acid (PABA)
- Paba esters
Tetracyclines (antibiotics, anti-infectives)
Tricyclic antidepressants
The concentration of drugs needed to elicit a photoallergic
reaction is much lower than that needed to cause a phototoxic
reaction. On the other hand, photoallergic reactions occur only in a
small proportion of exposed individuals while phototoxic reactions
may occur in anyone, given sufficient exposure. Generally, no
clinical reaction occurs on first exposure to an agent causing
subsequent photoallergy. Even sunscreen agents used to protect
against photocontact dermatitis may be photoallergenic. A small
number of individuals who develop photocontact dermatitis may retain
a persistent reactivity to light (including UV) long after exposure
to the photosensitizing compound.
8.4 Chronic Effects on the Skin Other than Cancer
UV radiation causes a number of chronic degenerative changes in
the skin, mainly in caucasian populations, as a result of its action
on keratinocytes, melanocytes and components of the dermal stroma
including fibrous tissue (collagen and elastin) and blood vessels.
These changes include freckles (ephelides), melanocytic naevi,
lentigines, telangiectasia, skin wrinkling and atrophy, yellow
papules and plaques on the face, colloid milium (firm, small,
yellow, translucent papules on the face, forearms and hands),
diffuse erythema, diffuse brown pigmentation and ecchymoses
(Goldberg & Altman, 1984). These or related changes are sometimes
grouped into syndromes such as cutis rhomboidalis nuchae (thick,
yellow, furrowed skin, particularly on the back of the neck),
Favre-Racouchot syndrome (yellow, thick comedones and follicular
cysts of the periorbital, malar and nasal areas) and reticulated
poikiloderma (reddish brown reticulated pigmentation with
telangiectasia and atrophy and prominent hair follicles on the
exposed chest and neck). With the exception of freckles and
melanocytic naevi, these changes are also referred to collectively
as "photoageing" (Gilchrest, 1990) because of their association with
increasing age but presumed correlation with total accumulated
exposure to the sun rather than with age per se.
In the US National Health and Nutrition Examination Survey
(Engel et al., 1988), age-adjusted prevalence proportions of
senile elastosis, actinic (solar) keratosis, fine telangiectasia,
localized hypermelanism, senile (solar) lentigines and freckles, in
whites 1 to 74 years of age, were associated with lifetime exposure
to the sun as estimated by dermatologists.
Freckles and solar lentigines
Freckles and solar (also called senile) lentigines are
pigmented macules occurring on the sun exposed skin of caucasians.
Their prevalence is increased in those with highly sun-sensitive
skin (Azizi et al., 1988). Freckles occur most commonly in
children while the frequency of solar lentigines increases with age
and is greatest in those over 60 years of age (estimated at 75% in
the USA: Rhodes et al., 1991). They show similar histological
patterns: their are increased numbers of melanocytes and an
increased concentration of melanin in the basal layer of the
epidermis (Rhodes et al., 1991). Melanocytic atypia has been
observed in both. An increased risk of melanoma has been observed in
relation to freckling in childhood and an increased risk of
non-melanocytic skin cancer in relation to both freckling and
prevalence of solar lentigines (see sections 8.4.1 and 8.4.2).
Melanocytic naevi
Melanocytic naevi are benign proliferations of melanocytes
usually beginning in the basal layer of the epidermis and later
extending into the dermis. They are common in white populations and
rare in black and Asian populations (Armstrong & English, 1988;
Gallagher et al., 1991), are associated, in white populations,
with phenotypic indicators of constitutional sensitivity to the sun,
particularly fair skin colour (Green et al., 1988b; Gallagher et
al., 1990b), occur mainly on body sites that are maximally or
intermittently exposed to the sun (Kopf et al., 1978, 1985;
Augustsson et al., 1990; Gallagher et al., 1990a), occur more
commonly in Australian than British children (Green et al., 1988b)
and in persons born in Australia than immigrants who arrived in
Australia after about 15 years of age (Armstrong et al., 1986),
and somewhat inconsistently with measures of sun exposure, including
sunburn in early life (Armstrong et al., 1986; Gallagher et al.,
1990c; Coombs et al., 1992). They are associated with an increased
risk of cutaneous melanoma (see section 8.4.2).
Solar keratoses
Solar keratoses are benign proliferations of epidermal
keratinocytes. They are very common on exposed body sites in older
people in caucasian populations living in areas of high ambient
solar irradiance (see for example, Marks et al., 1983 and Holman
et al., 1984a). Solar keratoses have been reported to be
associated with phenotypic indicators of cutaneous sun sensitivity
(Vitasa et al., 1990), to be more common in people born in
Australia than in migrants to Australia (Goodman et al., 1984), to
be associated with estimates of total and occupational sun exposure
(Goodman et al., 1985; Vitasa et al., 1990), and to be
associated with other benign indicators of cutaneous sun damage
(Holman et al., 1984b; Green 1991). Their number on the skin is
strongly associated with risk of non-melanocytic skin cancer (see
section 8.4.1).
8.5 Cancer
Epidemiological evidence relevant to the effects of UV on risk
of cancer in humans derives mainly from study of the effects of sun
exposure (presumably solar UV but not separable from other solar
radiation) on cancer risk. There are four general lines of evidence
available from which it may be inferred that sun exposure causes any
particular cancer. They are that: the cancer in question occurs more
frequently in people who are sensitive to the sun, occurs mainly at
sun-exposed body sites, is increased in residents of areas of high
ambient solar irradiance, and is increased in people with high
personal sun exposure.These lines of evidence will not apply to all
cancers that may be linked to sun exposure. They form nonetheless, a
useful framework within which most of the relevant evidence can be
described. To this framework will be added a consideration of
evidence relating to artificial sources of UV when it is available.
The most direct evidence of the carcinogenicity of UV in humans
should come, in principle, from observation of the effects of
personal exposure to the sun. In practice, it is very difficult to
make measurements of personal sun exposure accurately. Most often
they are made by questionnaire and require recall of rather
non-salient details of life over 60 or more years. This is a very
difficult task (Kricker et al., 1993). A difficulty is presented
by the fact that people who have sun sensitive skin and are at
higher risk of skin cancer will tend to expose themselves less to
the sun. To obtain an accurate measure of the effects of personal
sun exposure, this confounding with sun sensitivity should be
controlled - this has not always been done. It may not be
surprising, therefore, that measures of personal exposure to the sun
have not been consistently associated with risk of cancers thought
to be related to the sun and that more indirect evidence has proved
to be stronger.
8.5.1 Nonmelanocytic skin cancer
Introduction
There are two major histopathological types of nonmelanocytic
skin cancer: basal cell carcinoma (BCC) and squamous cell carcinoma
(SCC). BCC is the commoner type in white populations.
The epidemiology of nonmelanocytic skin cancer is difficult to
describe accurately. Its routine recording is often not attempted by
cancer registries because of the large numbers of cases involved
and, if attempted, is invariably incomplete because of the rarity
with which primary nonmelanocytic skin cancers require hospital
treatment and the frequency with which probable nonmelanocytic skin
cancers are not sent for histopathological verification of the
diagnosis (Muir et al., 1987).
These difficulties have influenced the quality of the
epidemiological evidence relating nonmelanocytic skin cancer to sun
exposure. In addition, most of the earlier cross-sectional and
case-control studies of nonmelanocytic skin cancer , and some of the
more recent ones, (see, for example, table 11 in IARC, 1992) are
deficient in that the control series were rarely population-based,
appropriate effect measures or p values were often not estimated,
and confounding by age and sex were not controlled in the analysis.
In the narrative that follows, little reliance will be placed on
these studies which include those reported by Lancaster & Nelson
(1957), Gellin et al. (1965), O'Beirn et al. (1970), Urbach et
al. (1974), Aubry & MacGibbon (1985), O'Loughlin et al. (1985),
Herity et al. (1989), Hogan et al. (1989) and Gafa et al.
(1991).
The results of most of the studies referred to below have been
described and their results tabulated in detail in other
publications (e.g., IARC, 1992) thus only the immediately salient
features will be described here.
Sun-sensitivity
Race
Nonmelanocytic skin cancer is much less frequent in populations
with dark skins than those with light skins (Fitzpatrick & Sober,
1985; Hoffman, 1987; Urbach, 1987). Data from cancer registries
represented in Cancer Incidence in Five Continents Volumes II-VI
(Doll et al., 1970; Waterhouse et al., 1976, 1982; Muir et al.,
1987; and Parkin et al., 1992) in which direct comparisons of
nonmelanocytic skin cancer incidence in different ethnic groups can
be made within a single geographical area show that the incidence
rates in the light skinned populations are consistently the highest.
Similarly, in the 1977-78 US survey of nonmelanocytic skin cancer;
rates in whites were 232.6 per 100 000 person years compared with
3.4 in blacks (Scotto et al., 1983).
Available evidence suggests that BCC occurs less frequently
than SCC among dark skinned populations. The opposite is the case in
light skinned populations. BCC was the most common nonmelanocytic
skin cancer reported in South African whites in 1949-1975 but was
rare among black Africans, occurring mainly in albinos (Oettlé,
1963; Oluwasanmi et al., 1969; Rippey and Schmaman, 1972; Isaacson
et al., 1978). SCC, principally on the lower limb and associated
with previous trauma, was the commoner of the two in blacks (Oettlé,
1963; Oluwasanmi et al., 1969; Rippey and Schmaman, 1972; Isaacson
et al., 1978; Rose, 1973). Melanesians (Foster & Webb, 1988) and
Polynesians (Paksoy et al., 1991) had less BCC than SCC, while no
BCCs were reported in the Melanesians of North Samoa, a particularly
heavily pigmented people (Foster & Webb, 1988).
Ethnic background is an important determinant of risk of
nonmelanocytic skin cancer in Caucasians. In people of southern
European ethnic origin born in Australia, relative to other people
born in Australia, risk of BCC was 0.56 (95% CI 0.14-1.65) for those
with one southern European grandparent, 0.17 (0.00-1.05) for two and
0.00 (0.00-0.86) for three or four (p for trend 0.002; Kricker et
al., 1991a). No subject with SCC in this study had any southern
European grandparents (RR 0.00, 0.00-1.23).
Individual sun sensitivity
Among recent cross-sectional, case-control and cohort studies
several have reported significantly elevated relative risk (RR)
estimates for red or light hair colour with BCC (RRs between 1.5 and
2.9; Hunter et al., 1990; Green & Battistutta, 1990; Kricker et
al., 1991a) and SCC (RR=2.4; Kricker et al., 1991a). However, a
light complexion was significantly associated only with SCC (RR=3.3;
Kricker et al., 1991a), and high RRs (of 3.4) for light hair
colour with BCC and SCC were reported in only one study (Vitasa et
al., 1990).
Sensitivity of the skin to the sun, as measured by ability to
tan and susceptibility to sunburn, was more consistently related to
risk of BCC and SCC than hair, skin and eye colour. RRs of 2.0 or
more for BCC were found with a skin that burns rather than tans
(Marks et al., 1989; Vitasa et al., 1990; Hunter et al., 1990;
Kricker et al., 1991a). For SCC, the evidence of an increased risk
with sun-sensitive skin was somewhat weaker, with RRs around 2.0 and
95% confidence intervals (CI) that were wide and included 1.0, in
the two studies that reported RRs adjusted for age, sex, and other
relevant confounders (Vitasa et al., 1990; Kricker et al.,
1991a).
Xeroderma pigmentosum and albinism
Xeroderma pigmentosum (XP) is a recessively inherited syndrome
characterised by clinical and cellular hypersensitivity to solar
radiation and a defect in the capacity to repair UV-induced damage
in DNA (Fitzpatrick et al., 1963, Cleaver, 1973). Evidence of
cutaneous sun damage may appear as early as 1-2 years of age in the
absence of specific protection from the sun, and skin cancers are
very frequent (Kraemer et al., 1987). In a survey of 830 cases of
XP located through published case reports, 45% were reported to have
had skin cancers (Kraemer et al., 1987). The median age of
diagnosis of the first skin cancer was 8 years. Ninety seven per
cent of BCC and SCC were on constantly exposed sites, that is, the
face, head, and neck, compared with an estimated 80% in the general
US population. In 220 XP patients in whom the number of skin cancers
was stated, half had more than two cancers and 5% had more than 30;
79 patients were described as having BCC and 112 SCC (Kraemer et
al., 1987).
Albinism is an inherited disorder of melanin metabolism with a
decrease in or complete absence of melanin; as a result, the skin of
albinos is highly sensitive to the sun. The most common type of
albinism occurs in 1 in 15,000 American blacks, 1 in 40,000 European
or American Caucasians, and has estimated frequencies as high as
1:3,900 in Soweto, South Africa (Kromberg and Jenkins, 1982), and 1
in 1,000 in Nigeria (Cervenka et al., 1979). African albinos have
been reported to have a high incidence of SCC and a somewhat lower
rate of BCC (Cervenka et al., 1979; Luande et al., 1985;
Kromberg et al., 1989).
Body-site distribution
It is commonly stated that the site distributions of
non-melanocytic skin cancers, particularly SCC, correspond well to
what would be expected from the exposure of different body sites to
the sun. They do generally conform to this pattern with more than
60% of lesions occurring on the head and neck (Kricker et al.,
1993). It is consistently observed, however, that the proportion of
SCC on the upper limbs is higher than that of BCC while BCC has the
higher proportion (some 10% or more) on the trunk (Scotto et al.,
1983; Osterlind et al., 1988a; Giles et al., 1988; Glass &
Hoover, 1989; Levi et al., 1988; Karjalainen et al., 1989;
Kricker et al., 1990; Gallagher et al., 1990a; Roberts, 1990;
Magnus, 1991; Serrano et al., 1991; Kricker et al., 1993). At
the sub-site level, BCCs are almost completely absent on the heavily
exposed backs of hands, and infrequent on the forearms compared with
the upper arms (Brodkin et al., 1969; Scotto et al., 1983;
Goodman et al., 1984; Kricker et al., 1990); in addition, this
subsite distribution on the face is not highly correlated with the
distribution of UV erythema on the face (Diffey et al., 1979).
Residence in areas of high ambient solar irradiance
Geographical variation
Annual incidence rates of nonmelanocytic skin cancer in 29
populations of mainly Western European origin in Cancer Incidence in
Five Continents, volume 6 (Parkin et al., 1992) show little
evidence of any consistent relationship between incidence and
latitude. The highest incidence rates were in the populations of
Tasmania, Australia (42°S) (213.2 per 100,000 in men and 113.1 per
100,000 in women) and British Columbia, Canada (49°N) (134.1 in men
and 91.2 in women), and were also comparatively high in Switzerland
(47°N) (78.7 in men and 50.2 in women) and Southern Ireland (53°N)
(71.5 in men and 48.0 in women).
In contrast to these international patterns, incidence rates of
nonmelanocytic skin cancer within countries do appear to increase
with proximity to the equator as indicated by broad place of
residence, latitude, or measures of solar irradiance. Geographical
variation in nonmelanocytic skin cancer incidence in the USA has
been described in three National Cancer Surveys (Mountin & Dorn,
1939; Dorn, 1944a, 1944b; Auerbach, 1961; Haenszel, 1963; Scotto et
al., 1974) and several related studies (Scotto et al., 1982,
1983; Serrano et al., 1991). Incidence of all types increased with
increasing proximity to the equator, with similar gradients for men,
women and all ages. The same pattern is seen in Australia (Marks et
al., 1993)
Migrants
The nonmelanocytic skin cancer experience of light-skinned
migrants from areas of low to areas of high ambient solar irradiance
has generally been consistent with an effect of sun exposure on skin
cancer incidence. In Australia, incidence and mortality were found
to be lower in migrants, most of whom had come from the UK, an area
of lower sun exposure, than in those born in Australia (Armstrong
et al., 1983; Giles et al., 1988). Kricker et al. (1991a)
examined in some detail the relationship between BCC and SCC and
migration to Australia. Migrants (excluding those from southern
Europe who may be at lower constitutional risk for skin cancer) had
a lower risk of BCC and SCC (RRs around 0.3) than did those born in
Australia. In addition, for BCC (which had sufficient cases to
analyze) the RR in those who migrated in the first 10 years of life
(1.05) was the same as that in those born in Australia (1.0) but
then fell to low levels in those who migrated later in life (RR
about 0.2). Hunter et al., (1990) observed an association between
risk of BCC and residence in southern parts relative to elsewhere in
the USA (RRs of 1.6 for residence in California and 2.1 for
Florida).
Personal sun exposure
Total sun exposure
Reported measurements of total current or accumulated sun
exposure in studies of nonmelanocytic skin cancer are likely to be
subject to substantial error not only because of the difficulties in
recalling sun exposure over periods of 60 years or more (Kricker et
al., 1993) but also because of the use of broad summary variables
such as "estimated average daily outdoor exposure" (Gellin et al.,
1965). The two studies which attempted a more quantitative measure
found no evidence of a positive association between estimates of
total sun exposure and risk of BCC (RRs of 1.0 or less) (Hunter et
al., 1990; Vitasa et al., 1990). Only one of these studies
included SCC (with only about 50% confirmed histopathologically);
for sun exposure above the 75th centile the estimated RR was 2.5
(95% CI 1.2-5.4) (Vitasa et al., 1990).
Occupational sun exposure
Early clinical reports associated skin cancer with outdoor
occupation (Molesworth, 1927; Blum, 1948; Emmett, 1973). However,
the relationship between occupation and skin cancer has been
examined adequately in relatively few studies and the evidence for
an association is weak. At the population level, the best conducted
studies which attempted to classify occupational sun exposure on the
basis of occupational title found only small differences in skin
cancer incidence between outdoor and indoor workers. Several
population studies reported associations with employment in
agriculture (Atkin et al., 1949; Whitaker et al., 1979; Teppo
et al., 1980) and outdoor employment generally (Beral & Robinson,
1981; Vågerö et al., 1986) but their interpretation is complicated
by incomplete and potentially biased case ascertainment (Vågerö et
al., 1986; Teppo et al., 1980), inclusion of other cancers
(Atkin et al., 1949), and confounding between occupation and
social class (Beral & Robinson, 1981).
There have been few well-conducted studies of occupational sun
exposure at the individual level. They show a generally consistent
but not strong association between BCC and SCC and various crude
measures of outdoor employment, e.g., "indoors", or "outdoors"
occupation. The RRs have generally been under 2.0 for BCC (Hogan et
al., 1989; Marks et al., 1989; Green and Battistutta, 1990; Gafa
et al., 1991). One study reported a high RR for SCC with an
"outdoors" occupation but with an extremely wide confidence interval
(Green and Battistutta, 1990). Other studies found RRs close to 1.0
(Aubry and MacGibbon, 1985, Hogan et al., 1989; Marks et al.,
1989; Gafa et al., 1991) and only one appeared to be statistically
significant (Hogan et al., 1989).
Recreational exposure
The relationship of exposure to sunlight in non-working hours
(largely recreational sun exposure) with nonmelanocytic skin cancer
has been described in only two studies. One study reported an RR
below 1.0 for BCC in those with "mainly outdoor leisure" and a
positive but not statistically significant association for SCC: RR
of 3.9 (95% CI 0.5-30.9) (Green & Battistutta, 1990). In the study
of Aubry & MacGibbon (1985), the highest category of a
"non-occupational exposure score" showed an elevated risk of SCC (RR
1.6; p-value=0.07).
Sunburn
Risk of BCC was significantly increased in subjects with a
history of sunburn in one of two recent studies. Hunter et al.
(1990) observed an increasing risk with increasing frequency of
painful sunburns (RR 2.91, 95% CI 2.37-3.58, for 6+ occasions).
Green and Battistutta (1990), on the other hand, found no
discernible association between BCC and number of painful sunburns.
The risk of SCC, however, was increased with any history of painful
sunburn in this study: RRs 3.3 (95% CI 0.9-12.3) for 1-5 sunburns
and 3.0 (0.7-12.2) for 6+.
Other sun-related skin conditions
Cutaneous microtopography (a measure of loss of the fine skin
markings on the backs of the hands presumed due to loss of dermal
collagen; Holman et al., 1984a), prevalence of solar elastosis of
the neck, solar telangiectasia and solar lentigines and a history of
solar keratoses have been taken as indicators of a high level of
total accumulated exposure to the sun in those who have BCC or SCC.
The evidence that sun exposure causes these conditions, however, is
no stronger, and may be weaker, than the evidence that it causes
BCC, SCC or cutaneous melanoma.
Increasingly severe sun-related skin damage as measured by
cutaneous microtopography was associated with increasing risks of
both BCC (RR 3.1, 95% CI 1.5-6.4 for the highest grade) and SCC (RR
1.8, 95% CI 0.8-4.2 for the highest grade) (Kricker et al.,
1991a). Other indicators of sun damage to the skin (e.g. freckles,
telangiectasia, and elastosis) were also strongly related to risk of
both BCC and SCC, whether considered separately (Green and
Battistutta, 1990; Kricker et al., 1991a) or together (Holman et
al., 1984a; Green et al., 1988a), and including when adjusted
for cutaneous sun sensitivity (Kricker et al., 1991a). The RRs
associated with the presence of 40 or more solar keratoses were 10.4
(95% CI 5.8-18.8) for BCC and 34.3 (95% CI, 14.0-84.0) for SCC
(Kricker et al., 1991a).
Exposure to artificial sources of UV
Any use of a sunlamp was associated with a statistically
significantly increased risk of SCC in the study of Aubry & McGibbon
(1985) but three other studies (O'Loughlin et al., 1985; Herity
et al., 1989; Hogan et al., 1989) found no association with
"artificial sunlight" or use of sunlamps or sunbeds.
8.5.2 Cutaneous Melanoma
Introduction
Cutaneous melanoma began to be studied epidemiologically in
1948 when it was separated from other primary malignancies of the
skin in the 6th Revision of the International Statistical
Classification of Diseases (WHO, 1948), and Eleanor MacDonald
published the first population-based study, covering 272 incident
cases of melanoma in Connecticut (MacDonald, 1948). Because of its
likely fatality if not treated, melanoma is much more likely to come
to medical attention and to be diagnosed histopathologically than
nonmelanocytic skin cancer. It has, therefore, been more readily
recorded on a population basis than nonmelanocytic skin cancer and
better studied epidemiologically. Thus, while generally considered
less clearly associated with sun exposure, a series of large and
well-conducted studies in different countries over the last 10 years
have served to clarify this issue.
Sun-sensitivity
Race
Melanoma is a disease primarily of light-skinned populations
and occurs much less frequently in people with darker skins. In the
United States, the incidence in whites is ten fold or more higher
than in blacks living in the same areas (Parkin et al., 1992).
Rates are lowest (less than 0.7 per 100 000 person years) in parts
of Asia (eg Japan, Singapore, India, China, Philippines) while in
Los Angeles, USA, incidence was less than 1 per 100 000 in Japanese
and Chinese people compared with slightly higher rates, around 2.0,
in blacks (Parkin et al., 1992).
Among Caucasians, ethnic background is an important determinant
of melanoma incidence. The incidence is substantially lower among
Hispanics than among other whites in the United States. For example,
the incidence among Hispanics in New Mexico is less than 2 per
100,000 person years, but in other whites it is about 11 per 100,000
(Muir et al., 1987). In several case-control studies, subjects
with a southern or eastern European background had substantially
lower risks than those of northern European or United Kingdom
origins (Elwood et al., 1984; Holman and Armstrong, 1984a; Graham
et al., 1985).
Individual sun sensitivity
Blue eyes, fair or red hair, and pale complexion in people of
European origin are well established risk factors for melanoma.
These pigmentary characteristics were documented in most melanoma
case-control studies (see table 16 in IARC, 1992 and, as examples,
Elwood et al., 1984, Holman & Armstrong, 1984a, and Green et al.,
1985). Relative risk for light skin colour ranged from a little more
than 1.0 to about 3. Compared with those with dark brown to black
hair, those with fair hair generally had a less than two-fold
increase in risk, but those with red hair usually had a 2 to 4-fold
increase in risk. Eye colour was generally a weak risk factor with
relative risks less than 2, and what increased risk there was
generally disappeared after adjustment for the other traits.
Increased risks of melanoma in those with a reduced ability to
tan and an increased tendency to sunburn were observed in all
case-control studies in which these sun-sensitivity measures were
examined (see, for example, Elwood et al., 1984, Holman &
Armstrong, 1984a, and Green et al., 1985).
Xeroderma pigmentosum and albinism
In a review of reports of 378 patients with XP in which cancer
was mentioned, 37 patients had a melanoma (Kraemer et al., 1987).
African albinos have been reported to have a low rate of
melanoma (Cervenka et al., 1979; Luande et al., 1985; Kromberg
et al., 1989). Levine et al. (1992) state that there have been
only 16 documented cases of melanoma in albino patients reported in
the English language literature. Since albinos generally have a
normal number of melanocytes (Dargent et al., 1992), the rarity of
melanoma in them may indicate that melanin plays an important role
in the genesis of this cancer.
Body site distribution
The distribution of melanoma appears to favour sites which are
less heavily sun exposed, that is the back and face in Caucasian men
and the lower limbs in women (Crombie, 1981). However, when
whole-population series of cancer cases have been examined and body
surface area taken into account, sites that were usually covered by
clothing had lower rates than those usually exposed, with the
exception of the forearms and hands for which the rates per unit
area were low (Elwood & Gallagher, 1983; Green et al., 1993). The
anatomic site distribution of melanoma in blacks is quite different
from that in Caucasians with the majority of melanomas on the soles
of the feet (Higginson and Oettlé, 1959; Lewis, 1967; Fleming et
al., 1975).
Residence in areas of high ambient solar irradiance
Geographical variation
Internationally, the incidence of melanoma varies over
100-fold. Among countries included in Cancer Incidence in Five
Continents, Volume VI, the lowest rates reported around 1983-87 were
0.1-0.2 per 100 000 person years in China (Qidong), Japan (Osaka),
India (Bombay), and in Kuwaitis in Kuwait, while the highest were
about 25 per 100 000 person years in parts of Australia (Parkin et
al., 1992). In the USA and Australia, which have reasonably
homogeneous populations of mainly European origin distributed across
a wide latitude range, melanoma incidence increases with increasing
proximity to the equator or with increasing measured, annual ambient
UV irradiance (Scotto & Fears, 1987; Jones et al., 1992).
A simple latitude gradient for melanoma is not evident in
Europe. Armstrong (1984) showed that the incidence of melanoma in
Europe decreased from about latitude 35°N to a minimum around 55o N
and then rose with increasing latitude because of high rates in
Scandinavian and Scottish populations. The gradient of risk for
melanoma from north to south in northern Europe matches the
increasing natural pigmentation of the skin and may also, in part,
be due to differing patterns of sun exposure, particularly
recreational and vacation sun exposure.
Migrants
If a person migrates in childhood to a country of high ambient
solar radiation such as Australia, Israel, and New Zealand, their
risk of melanoma is observed to be similar to that in those born in
the country to which they migrate, whereas if they migrate after
this age their risk is substantially less than that in the
locally-born population (Katz et al., 1982; Cooke and Fraser,
1985; McCredie & Coates 1989; Steinitz et al., 1989; Khlat et
al., 1992). Overall, British immigrants to Australia and New
Zealand, where the populations are of predominantly British origin,
had incidence and mortality rates of melanoma of about a half the
levels in those born in these countries (Cooke and Fraser, 1985;
Khlat et al., 1992).
Residence history
Studies of individual lifetime ambient solar radiance, based on
latitude, location (eg, tropical, near coast), or average sunshine
hours of all places of residence are consistent in showing an
increased risk of melanoma with increased average radiance (Green &
Siskind, 1983; Holman et al., 1984b; Graham et al., 1985;
Osterlind et al., 1988b; MacKie et al., 1989; Weinstock et al.,
1989; Beitner et al., 1990). RRs for the highest category of
ambient solar radiance were 2.8 for mean annual hours of bright
sunlight at places of residence in Australia (Holman et al.,
1984b), between 1.4 and 2.6 for a southerly latitude in the USA
(Graham et al., 1985; Weinstock et al., 1989), or a tropical or
Mediterranean residence (MacKie et al., 1989; Beitner et al.,
1990). RRs differed greatly for residence near the coast in Denmark
(RR=1.7; Osterlind et al., 1988b) and Australia (RR=5.0; Green &
Siskind, 1983).
Personal sun exposure
Lifetime total exposure
Inconsistent results have been obtained in studies in which
lifetime total sun exposure were assessed by questionnaires. RRs for
the highest exposure category were between 0.6 and 5.3 (see table 18
in IARC, 1992). Two of five studies showed a statistically
significant positive association with RRs of 3.4 and 5.4 (Grob et
al., 1990; Lê et al., 1992) while one study showed a significant
negative association (RR 0.6; Graham et al., 1985). The other two
RRs were 1.1 (Dubin et al., 1986) and 5.3 (95% CI 0.8-30.8; Green,
1984).
Usual or recent total exposure
In this context, total means exposure from all sources (i.e.
occupational and non-occupational) rather than the total over some
period of time. This concept has been measured in a variety of ways.
For example, it has been common to enquire about present "usual"
exposure to the sun or usual exposure in some time period in the
fairly recent past. The evidence of any effect of such exposures on
risk of melanoma is weak. Of five studies, four had relative risks
for the highest categories of exposure ranging from 0.7 to 1.2
(Elwood et al., 1985b; Dubin et al., 1986; Holman et al.,
1986; Cristofolini et al., 1987). The remaining study found a
relative risk of 2.5 (p < 0.001) for the highest category of
exposure of average daily sun exposure 10-20 years ago (Rigel et
al., 1983).
Occupational exposure
Four studies of melanoma have shown statistically significant
positive associations with estimated lifetime occupational exposure
to the sun (see table 20 in IARC, 1992) with RRs ranging up to 6.0
(95% CI, 2.1 to 17.4) for outdoor employment (Paffenbarger et al.,
1978; Dubin et al., 1986; Garbe et al., 1989; Grob et al.,
1990). On the other hand, four studies showed statistically
significant results in which the RR for the highest category of
exposure was < 1.0 (Elwood et al., 1985b; Holman et al., 1986;
Osterlind et al., 1988b; Beitner et al., 1990). The remaining
studies have shown RRs around 1.0.
Recreational exposure
Recreational exposure to the sun has generally been measured by
either the type of recreational activity or the frequency or
duration of outdoor recreation (see table 22: in IARC, 1992). Some
studies have also recorded sun exposure during vacations separately
from other recreational sun exposure. At least one statistically
significant positive association was found in 11 of 16 studies in
which recreational exposure was considered (Adam et al., 1981; Lew
et al., 1983; Rigel et al., 1983; Elwood et al., 1985b; Dubin
et al., 1986; Holman et al., 1986; Osterlind et al., 1988b;
Zanetti et al., 1988; Beitner et al., 1990; Grob et al., 1990;
Nelemans et al. 1993). The RRs in the highest categories of
exposure were generally between 1.5 and 2.5. Only one study has
shown a negative association with recreational exposure: MacKie &
Aitchison (1982), in Scotland, observed an RR of 0.4 (95% CI
0.2-0.9) for the highest category of hours a week in outdoor
recreation. Their statistical model, however, included socioeconomic
status and history of sunburn, both of which may measure sun
exposure; thus overadjustment of the estimate of RR is likely. All
measures of number or frequency of vacations in sunny places were
positively associated with risk of melanoma. The RRs for the highest
category of this variable ranged from 1.2 to 5 and the finding was
statistically significant in six of eight studies (Lew et al.,
1983; Elwood et al., 1985b, Osterlind et al., 1988b, Zanetti et
al., 1988; Beitner et al., 1990; Nelemans et al., 1993).
Sunburn
Fifteen of seventeen studies of melanoma in which sunburn was
recorded (see table 23 in IARC, 1992) showed a statistically
significant, moderately to strongly positive association with a
history of sunburn (MacKie & Aitchison, 1982; Lew et al., 1983;
Elwood et al., 1985a; Green et al., 1985; Sorahan & Grimley,
1985; Elwood et al., 1986; Holman et al., 1986; Holly et al.,
1987; Osterlind et al., 1988b; Zanetti et al., 1988; MacKie et
al., 1989; Weinstock et al., 1989; Beitner et al., 1990;
Elwood et al., 1990; Nelemans et al., 1993) (see table 23: IARC,
1992). Twelve of these studies showed RRs greater than 2.0 for the
highest category of sunburn. Positive associations were obtained for
both sunburn in childhood and sunburn at any age. The greater
consistency of the relationship of melanoma with sunburn compared to
that with other exposure variables may indicate a specific
association with sunburn per se or simply that sunburn is a more
accurately measured indicator of sun exposure.
Presence of other sun-related skin conditions
Cutaneous microtopography
Severity of sun damage to the skin as measured by cutaneous
microtopography was strongly associated with risk of melanoma in
Western Australia. The RR for the most severe grade of damage was
2.7 (95% CI 1.4-5.0; p-value for trend=0.003; Holman et al.,
1984b). There was no similar relationship, however, in a study in
Denmark, in which similar methods were used (Osterlind et al.,
1988b).
Freckling of the skin has been shown to be associated with an
increased risk of melanoma in several studies (see for example,
Elwood et al., 1986, 1990, Osterlind et al., 1988b; Dubin et
al., 1986).
Solar keratoses and other skin cancers
The risk of melanoma was significantly increased in association
with a past history of nonmelanocytic skin cancer with RRs of 3.7
(95% CI 2.1-6.6) in Australia (Holman et al., 1984b) and 3.8
(1.2-12.4) in the USA (Holly et al., 1987). Relative risks for a
history of solar keratoses or "actinic tumours on the face" were
similarly high (Green & O'Rourke, 1985; Dubin et al., 1986).
Exposure to artificial sources of UV
Several studies have reported increased risks of cutaneous
melanoma in users of sunlamps or sunbeds (IARC, 1992). In the most
recent of these studies (Walter et al., 1990), the relative risks
for any use of sunbeds or sunlamps were 1.9 (95% CI 1.2-3.0) in men
and 1.5 (95% CI 1.0-2.1) in women. Relative risk increased with
increasing duration of use: for more than 12 months use the relative
risks were 2.1 (0.9-5.3) in men and 3.0 (1.1-9.6) in women. Positive
associations were found in two of four other case-control studies in
which these exposures were studied (Swerdlow et al., 1988; MacKie
et al., 1989; see also IARC, 1992). Sun exposure is a potential
confounding variable in studies of sunlamps and sunbeds but has not
been taken into account. Specifically it was considered by Walter
et al. (1990). Exposure to a group of other artificial sources of
UV, including plan printers, laboratory equipment emitting UV,
insect tubes, black lights and photocopiers, was not associated with
melanoma in one case control study in Australia (Holman et al.,
1986) but for a similar group plus welding the RR was 2.2 (95% CI
1.0-4.9) in Canada (Elwood et al., 1986). Siemiatycki (1991) found
no association between arc welding or other occupational exposure to
UV in a study of occupation and cancer in Canada.
A number of case-control studies have examined the association
between melanoma and exposure to fluorescent lighting. Only the
first such study found a statistically significant positive result
(Beral et al., 1982). Six subsequent studies found little or no
evidence of a positive association (IARC, 1992).
8.5.3 Cancer of the lip
Cancer of the lip is defined as cancer of the vermilion border
and adjacent mucous membranes and thus excludes cancers of the skin
of the lip (WHO, 1977). Most are SCC and occur on the lower lip
(Keller, 1970; Lindqvist, 1979), a site more heavily exposed to the
sun than the upper lip (Urbach et al., 1966). The inclusion of
adjacent oral mucosa in the definition of cancer of the lip raises
the possibility of confounding of tobacco and alcohol use (well
established causes of cancer of the mouth; Tomatis, 1990).
The incidence of cancer of the lip is much more common in men
than women. Among men in the USA, it is some 20 times more common in
whites than blacks and it is rare in black and Asian populations
worldwide (Parkin et al., 1992). Incidence of and mortality from
cancer of the lip are substantially lower in migrants to Australia
and Israel, who come from places with potentially lower sun
exposure, than those born in these countries (Armstrong et al.,
1983; McCredie & Coates, 1989; Steinitz et al., 1989).
The incidence of cancer of the lip is higher in rural than
urban areas (Doll, 1991) and descriptive studies have consistently
reported higher incidence or mortality rates of this cancer in men
with outdoor occupations such as farmers, agricultural labourers and
fishermen. (Atkin et al., 1949; Clemmesen, 1965; Gallagher et
al., 1984; Olsen & Jensen, 1987; Lynge & Thygesen, 1990). Four
case-control studies of varying quality have examined the
association between outdoor work and cancer of the lip. Keller
(1970) compared 301 men with cancer of the lip with 301 oral cancer
controls. Crude RRs for employment as a farmer or in any outdoor
work were 4.0 and 2.6 respectively. The use of patients with oral
cancer as controls allowed a rough adjustment for confounding with
tobacco and alcohol use in this study. Spitzer et al. (1975)
compared 339 men with cancer of the lip with 199 matched population
controls. The relative risk for any outdoor work was 1.52 and, for
employment as a fisherman, 1.50 (p < 0.05 in both cases). Pipe
smoking but not alcohol drinking was controlled in the analysis. The
other two studies also showed a positive association between outdoor
work and cancer of the lip (Lindqvist, 1979; Dardanoni et al.,
1984) but did not control for smoking and alcohol use and showed
other methodological difficulties (IARC, 1992).
8.5.4 Ocular cancers
Ocular melanoma and other ocular cancers are dealt with in
Chapter 10.
8.5.5 Other cancers
Observations of increasing mortality with increasing latitude
have formed the basis of suggestions that cancers of the breast,
colon and prostate may be prevented by increasing exposure to the
sun (Garland & Garland, 1980; Gorham et al., 1989, 1990; Garland
et al., 1989, 1990; Hanchette & Schwartz, 1992), perhaps through
proposed anti-carcinogenic actions of vitamin D (Eisman et al.,
1980; Colston et al., 1989; Schwartz & Hulka, 1990; Ainsleigh,
1993). Variation with latitude in medical care, death certification
practices, diet and other lifestyle factors are alternative
explanations for the latitude gradients in these cancers. These
observations, therefore, can be taken only to raise hypotheses about
indirect anti-carcinogenic effects of UV which require testing by
other means.
8.5.6 Action spectrum
There are few data from which the action spectrum of UV
carcinogenesis in humans can be inferred. For practical purposes,
therefore, reliance is placed on action spectra which have been
determined in experimental animals (see chapter 7).
If, as is argued below, it is accepted that the formation of
cyclobutylthymidine dimers is a step in the production of mutations
that are associated at least with BCC and SCC, then it would be
reasonable to accept the action spectrum for production of thymidine
dimers in humans as possibly indicative of the action spectrum for
production of nonmelanocytic skin cancer (figure 8.1). This action
spectrum was determined by Freeman et al. (1989) who irradiated
untanned gluteal skin of 30 caucasian volunteers with 1 or 2 MED of
narrow band UV at 275 nm, 282 nm, 290 nm, 296 nm, 304 nm, 334 nm and
365 nm. At least five subjects were irradiated at each wavelength.
Following irradiation, shave biopsies were taken of the irradiated
skin, DNA extracted and incubated with Micrococcus luteus
endonuclease to determine the frequency of endonuclease sensitive
sites which indicate the presence of thymidine dimers. Figure 8.1
shows that the effectiveness of UV in producing dimers increased
from 275 nm to a peak at 296 to 304 nm and then fell by four orders
of magnitude to the lowest measured level at 365 nm. This fall is
quite consistent with that observed in the relative effectiveness of
UV for carcinogenesis in mouse skin over the same wavelength range.
The authors reported, also, that observations had been made at 385
nm and 405 nm and that these wavelengths were "largely ineffective"
in net production of dimers.
Some indication of the effectiveness of UVA in causing skin
cancer could be obtained from the experience of people exposed to
high doses of UVA from sunbeds or in tanning salons. There is some
evidence to suggest that these exposures are associated with an
increase in risk of nonmelanocytic skin cancer, cutaneous melanoma
and ocular melanoma (see above). In none of the studies carried out,
however, has exposure to mainly UVA sources been distinguished from
sources that emit both UVA and UVB. In addition, confounding with
sun exposure is a possibility in all these studies.
8.5.7 Dose-response
In theory, the best way to estimate the dose-response
relationship between UV and cancer in humans would be to measure the
lifetime exposure to the sun of each member of a population and
relate this measurement, by way of a cohort or a case-control study,
to their probability of developing skin cancer. In practice this is
very difficult. First, measurement of lifetime exposure to the sun
generally requires recall of amount and pattern of sun exposure and
use of protective measures against the sun for as long as 40 or more
years in the past.
Second, exposure in early life may be particularly important in
determining risk of skin cancer, at least, and this is to a large
extent outside the range of accurate recall. Third, while problems
of recall could be solved by conduct of a cohort study, such a study
would require measurements of sun exposure beginning in the first
few years of life and repeated periodically throughout life to the
age at which UV-related cancers become common. Such a study would be
logistically and economically impracticable. The whole process is
complicated by the fact that a simple measure of lifetime exposure
may be insufficient for adequate description of dose-response. The
evidence that period and pattern of exposure may be important
determinants of some UV-related cancers (see below) means that to
model incidence correctly as a function of exposure it would be
necessary to collect data on dose-rate of UV and its variation
throughout life.
Noonan et al. (1981a) reported a 50% suppression of CHS in
BALB/c mice after a single dose of 2 KJ m-2 UV. She also reported
that there was no difference in the dose response curves for
UV-induced local and systemic immunosuppression of CHS responses;
however, the dose of UV required to suppress CHS in C57BL/6 mice was
6.4 times less than that required to produce similar immune
suppression in BALB/c mice (Noonan & De Fabo, 1990). Also, while
local suppression could be demonstrated in mice sensitized
immediately after irradiation, systemic suppression occurred only if
the sensitizer was applied 3 or more days after radiation (Noonan &
De Fabo, 1990).
Fifty percent suppression of CHS and DTH has been reported at
doses ranging from 2.3 kJ m-2 to 20 kJ m-2, (Kim et al., 1990;
Jeevan & Kripke, 1990; Jeevan et al., 1992b) levels which produce
minimal or no edema. The dose needed to produce 50% suppression
varies depending on the strain of mouse and type of antigen used,
and on the laboratory producing the data. Suppression of
virus-specific DTH was observed in mice exposed to a suberythemal
dose of UVB 3-7 days prior to infection with Herpes simplex virus
(HSV) but not 14 days and persisted for at least 3 months after
irradiation (Howie et al., 1986). Elimination of wavelengths below
315 nm with a mylar filter either eliminated or greatly reduced the
suppression of CHS following exposure to UV indicating that most if
not all the suppression is due to UVB (Noonan et al., 1981b;
Noonan and De Fabo, 1990). Using narrow bands of UV at 10
wavelengths from 250 to 320 nm, it was reported that maximum
suppression of CHS occurred between 260 and 270nm, there was a
shoulder in the action spectrum from 280-290 nm and then a steady
decline to 3% of maximum at 320 nm (De Fabo and Noonan, 1983).
There are two subpopulations of T helper cells, designated Th1
and Th2 which appear to be differentially affected by UV exposure.
These two populations are thought to regulate different sets of
immune responses. Th1 cells produce interleukin (IL) 2 and gamma
interferon (gamma IFN) as well as other cytokines, promote
delayed-type hypersensitivity (cell mediated, type IV) responses
such as CHS, provide help for certain antibody subtype responses
including complement-fixing antibodies, activate macrophages, and
may be particularly important for dealing with antigens expressed on
cell surfaces, such as viral and tumour antigens (Coffman et al.,
1988). Th2 cells produce a different array of cytokines including
IL-4 and IL-5 which promotes antibody responses. Only Th2 cells can
stimulate a primary IgE response which is mediated by IL-4 and
inhibited by gamma IFN (Coffman et al., 1988). Thus, Th2 cells may
be particularly important in responding to certain parasitic
infections, and also play an important role in immediate-type
hypersensitivity including reactions to common allergens such as
pollen and dust mite.
Recent studies have suggested that both local and systemic
immune suppression induced by UV may be the result of an inability
to present antigen to and hence activate Th1 cells (Simon et al.,
1990, 1991; Araneo et al., 1989).
Suppression of systemic CHS and DTH responses similar to that
observed following UV exposure have been obtained by injecting mice
with supernatant from UV-irradiated keratinocytes (Kim et al.,
1990; Rivas & Ullrich, 1992; Jeevan et al., 1992c). Among factors
derived from keratinocytes, the cytokines tumour necrosis factor
alpha (TNF alpha) (Yoshikawa & Streilein, 1990; Vincek et al.,
1993) and IL-10 (Rivas & Ullrich, 1992) are thought to be important
in UV-induced immunosuppression. Also, cis-urocanic acid, the
product of UV isomerization of urocanic acid located in the stratum
corneum, when injected subcutaneously or painted on the epidermis,
produces immunosuppressive effects which mimic UVB-induced
immunosuppression (Ross et al., 1986), and the action spectrum of
UV-induced immune suppression closely follows the absorption
spectrum of urocanic acid (De Fabo and Noonan, 1983). In all three
cases it is thought that these mediators act by modifying antigen
presentation (Vermeer & Streilein, 1990; Rivas & Ullrich, 1992;
Noonan et al., 1988).
In summary, it appears that UVB causes the release of mediators
from the skin which alter the antigen presenting capability of
Langerhans cells as well as antigen presenting cells at other sites,
resulting in the development of "suppressor T-cells". It may be that
these suppressor T cells are Th2 cells. The net result is the
failure to activate Th1 cells and suppression of DTH responses
thought to play an important role in host defences against certain
types of tumours and microbial infections. The immune suppression is
antigen specific, (i.e. only responses to antigens administered
within 7 days after irradiation are affected) and is long lasting
(at least 3 months).
In addition to DTH and CHS responses, UV affects several other
immune functions frequently included in the standard protocols for
immunotoxicity testing. Spleen cells taken from UV-exposed mice
(single exposure, 54 kJ m-2) failed to respond in a mixed
lymphocyte response assay in vitro (Ullrich, 1985), and spleen
cells from tumour-implanted UV-treated (10 kJ m-2, 3 times/wk, for
3 months) mice were not cytotoxic in vitro against UV-induced
tumours (Fisher & Kripke, 1977). In contrast, exposure to
approximately 2-3 kJ m-2 day-1, for 23 days did not affect
responses of spleen or inguinal lymph node cells to the mitogens,
phytohaemagglutinin, concanavalin A or bacterial lipopolysaccharide.
Similarly, lymphocyte responses to these mitogens and peritoneal
macrophage phagocytic and tumouricidal activities were unaffected by
exposure to 10 kJ m-2, 3 times per week, for up to 6 months
(Funnell & Keast 1985; Norbury et al., 1977).
Kripke et al., (1977) reported no effect of UV (in mice
exposed to 10 kJ m-2, 3 times per week for up to 6 months) on the
primary haemagglutonin antibody response to sheep red blood cells;
however in another study, the IgM and IgG plaque forming cell
responses of lymph node cells to sheep red blood cells given
intradermally (through irradiated skin) were suppressed (Funnell &
Keast, 1985). Also, suppressor cells generated by irradiating mice
with a single exposure of 30-40 kJ m-2, 5 days prior to
sensitization with trinitrochlorobenzene prevented the development
of hapten specific antibody-forming cells when injected
intravenously along with hapten conjugated sheep red blood cells
into syngeneic recipients (Ullrich et al., 1986b).
UV exposure enhanced poly I.C augmented natural killer cell
(NK) activity in mice (Lynch & Daynes, 1983); however, suppression
of NK activity in rats has been reported following exposure to
suberythemal doses of UV (Garssen et al., 1993). Hence, in
laboratory animals UV exposure appears to affect mixed lymphocyte
responses, cytotoxic T cell activity, possibly NK activity, and in
some circumstances antibody responses. With the exception of NK
activity all of these responses are antigen specific responses. In
contrast UV does not appear to alter several non-specific responses
including macrophage phagocytic or tumouricidal activities, or T or
B cell responses to mitogens. However, unlike CHS and DTH, none of
these responses has been studied in detail in rodents, and dose
response information is not available.
7.2.2 Susceptibility to tumours
Interest in the immunosuppressive properties of UVB was first
sparked by the observations of Kripke and associates that UV
exposure induced highly antigenic tumours which did not grow when
transplanted into syngeneic, immunocompetent mice but did grow when
transplanted into immunosuppressed or UV treated mice (Kripke 1974;
Kripke & Fisher, 1976). The precise nature of the tumour antigens
has not been defined. Mice treated with 1.8 kJ m-2, 3 times per
week, for 3 months were unable to reject UV-induced syngeneic
tumours when challenged subcutaneously at any time from 2 weeks into
the irradiation exposure regimen to as late as 5 months after the
end of UV treatment (Kripke & Fisher, 1976). Hence susceptibility to
tumour challenge was detectable well before the appearance of
primary skin cancers induced by the UV and persisted long after
exposure ended. Similarly, more UV-induced fibrosarcoma tumour
colonies were detected in the lungs following i.v injection of
tumour cells in mice treated with 7 kJ m-2, 3 times per week, for
5 weeks (Kripke & Fidler 1980) as compared to unirradiated controls.
Susceptibility to tumour challenge was directly proportional to dose
of UV and a dose fractionated over time was no more effective than
the same total dose given as a single treatment (Kripke & Fidler
1980; De Fabo and Kripke 1979).
In BALB/cAnN mice 50% tumour incidence following subcutaneous
tumour challenge at an unirradiated site occurred at a dose of
approximately 40 kJ m-2. Enhanced tumour incidence was observed in
mice exposed to 21.6 kJ m-2 as much as 32 weeks before tumour
challenge. A number of studies (Fisher & Kripke, 1977; Daynes &
Spellman, 1977; Daynes et al., 1977; Spellman & Daynes, 1978;
Spellman et al., 1977; Ullrich & Kripke, 1984, de Gruijl and van
der Leun 1982, 1983) have demonstrated that susceptibility to
challenge with UV-induced tumours following UV exposure, like
suppression of CHS and DTH, is mediated by antigen specific
"suppressor T cells" which are present in spleen and lymph nodes of
UV-irradiated mice. As with suppression of DTH and CHS responses,
enhanced susceptibility to UV-induced tumours is antigen specific
and long lasting.
While it is very clear that UV increases the susceptibility of
mice to UV-induced tumours, much less work has been done on the
effects of UV on susceptibility to other types of tumours. UV
treated mice that were unable to reject syngeneic UV- induced
tumours were able to reject 2 types of non-UV-induced tumours, B16
melanoma tumours and a spontaneous mouse leukaemia tumour (Kripke
et al., 1977). Roberts and Daynes (1980) reported that mice
irradiated with subcarcinogenic doses of UV for 3 weeks prior to
treatment with either benz[a]pyrene or methycholanthrene at an
unirradiated site had reduced latency periods for the development of
these chemically-induced tumours. Also, chemically induced tumours
from these UV-treated mice appeared to be more antigenic than those
induced in untreated mice in that they were incapable of progressive
growth when transplanted into normal (immunocompetent) syngeneic
mice but were capable of progressive growth in UV-treated mice. The
authors suggested that decreased selective pressures exerted by the
host (due to UV-induced suppressor cells) at the time of tumour
induction allowed more immunogenic tumours to emerge and progress.
7.2.3 Susceptibility to infectious disease
Several types of infectious disease models have been developed
to study the effects of UV exposure: 1) the infectious agent was
injected through irradiated skin, 2) exposure to the infectious
agent occurred at a site distant from the site of irradiation, or 3)
in the case of some herpes simplex virus (HSV) studies, exposure to
UV occurred after infection. The first exposure scenario is
representative of vector-borne infections and some vaccinations
while the second exposure regimen is representative of infections
which do not necessarily enter the host via the skin. The third
scenario was designed to mimic reactivation of HSV in humans
following sun exposure. In all cases the focus has been on microbial
agents that are controlled, at least in part, by DTH responses.
An example of the first type of model is that of Leishmania
major infection in mice (Giannini 1986a, 1986b, 1987, 1992). Mice
were irradiated only on the tail with suberythemal doses (0.06-6 kJ
m-2), 3 times/week, for 1 month and were infected intradermally
through the irradiated surface of the tail 24 hr after the first UV
exposure. In UV-treated mice the DTH response to L. major antigens
2 and 6 weeks post infection was suppressed, the number of organisms
recovered from skin at the injection site was (Giannini, 1986a) and,
a larger number of parasites was observed in the draining lymph node
(Giannini, 1987, 1992). Finally, mice which were infected through
irradiated skin failed to develop protective immunity such that
lesions following reinfection at an unirradiated site were
significantly larger when compared to lesions of previously infected
but unirradiated mice (Giannini, 1986b).
In a similar model mice treated with 13 or 33 kJ m-2 for 4
consecutive days and infected intradermally with HSV at the site of
irradiation had a higher incidence of zosteriform lesions than
unirradiated mice, and at the higher exposure level 100% mortality
was observed as opposed to no mortality in unirradiated mice
(Yasumoto et al., 1987). A decreased DTH response to viral antigen
was observed and appeared to result from the induction of suppressor
T cells (Yasumoto et al., 1987, Aurelian et al., 1988). Also,
following in vitro exposure to UV, a defect in the ability of
Langerhans cells to present HSV antigen to lymphocytes was
demonstrated (Hayashi & Aurelian, 1986).
In contrast, no effect of UV on either parasite specific DTH
responses or recovery of parasite from internal organs was observed
following percutaneous injection of Schistosoma mansoni through
UV-irradiated (0.4 kJ m-2, 4 consecutive days) skin (Jeevan et
al., 1992a). In this same study, suppression of microbe specific
DTH responses were not observed following intradermal injection of
Mycobacterium bovis (BCG) or subcutaneous injection of Candida
albicans at the site of irradiation; however, the number of viable
mycobacteria recovered from the lymphoid organs of BCG-infected mice
was increased significantly in the UV treated mice for a period of
more than 2 months post infection.
BCG has also been used as an infectious disease model to
illustrate systemic effects of UV immunosuppression. In this model
mice were irradiated on the back and injected with BCG
subcutaneously in the footpad. Mice exposed from 1 to 15 times (3
times/week for up to 5 weeks) to one minimal erythemal dose (2.25 kJ
m-2) showed significant suppression in their DTH response to
tuberculin (PPD) and increased numbers of live bacteria in the
spleen and lymph node compared to unirradiated controls (Jeevan &
Kripke, 1990). However, when exposures were continued beyond 5
weeks, the DTH response recovered and mice challenged with bacteria
at that point did not exhibit increased numbers of organisms in the
spleen and lymph node, suggesting that eventually some sort of
adaptation to exposure occurs. Significant suppression of DTH was
observed in mice which received a single dose as low as 1.4 kJ m-2
3 days prior to infection and significant increases in bacteria in
spleen and lymph node were observed in mice which received as little
as 0.7 kJ m-2. Similar effects were observed when supernatants
from keratinocyte cultures exposed to UV in vitro were injected
intravenously 3 days prior to infection (Jeevan et al., 1992c).
In a similar model, mice treated with a single high UV dose (45
kJ m-2) 3 days before infection with Mycobacterium lepraemurium
exhibited significant suppression of DTH responses to mycobacterial
antigen 3 and 6 months after infection and had significantly more
bacteria in the infected footpad, lymph node, and spleen 3-6 months
post infection (Jeevan et al., 1992b). This high dose also reduced
the median survival time in mice infected i.v. With a lower exposure
dose (2.3 kJ m-2) 50% suppression of the DTH response to
mycobacterial antigen was observed 3 months post infection, and
increased numbers of bacteria were observed in the footpad, spleen
and lymph node of mice exposed to UV doses greater than or equal to
5.6 kJ m-2. When mice were treated with 2.25 kJ m-2, 3
times/week from 3-15 times, the DTH response to M. lepraemurium
was suppressed, but as with BCG, the DTH response was normal in mice
which received more than 15 exposures.
Effects of systemic UV-induced immune suppression have been
demonstrated in mice infected with HSV, 3 days after irradiation
with 1 kJ m-2, at a site distant from the site of infection. In
this model suppression of DTH was mediated by modulation of
epidermal antigen presenting cells and the development of suppressor
T cells (Howie et al., 1986, 1987). Subcutaneous injection or
epidermal application of UV-treated urocanic acid produced similar
immune suppression (Ross et al., 1986). Effects of UV on the
actual progression of disease in this model have not been reported.
While most of the work to date has been done in mice, recent
work in the rat suggests that suberythemal doses of UV also suppress
immune responses. In addition rats exposed to suberythemal doses of
UV had higher levels of microorganisms in target organs following
oral infection with Trichinella spiralis or intraperitoneal
infection with cytomegalovirus (Garssen et al., 1993).
In summary, exposure to suberythemal doses of UV has been shown
to exacerbate a variety of infections in rodent models. Both
infections which are initiated at the site of UV exposure and
infections initiated at distant sites have been affected. While most
of the infections test, have evolved in some manner more recent work
indicates that systemic infections without skin involvement may also
be affected. Enhanced susceptibility appears to result from
suppression of T-helper-1 cell activity. The mechanisms associated
with this suppression appear to be the same as those identified in
association with suppression to CHS responses.
Reactivation of latent HSV infections can be induced in mouse
and guinea pig by exposure of the previously infected site to sub-
or minimal erythemal doses of UV. However, the role that immune
suppression plays in this reactivation has not been established
(Blyth et al., 1976; Norval et al., 1987; Laycock et al.,
1991).
7.2.4 Susceptibility to immunologically-mediated
diseases
Several studies have suggested that stimulation of Th2 cells
may remain intact or be enhanced in UV-exposed mice (Simon et al.,
1990; 1991; Araneo et al., 1989). Since Th2 cells are the only
cells that can stimulate a primary IgE response (Coffman et al.,
1988), it is possible that UV exposure may increase the risk of
immediate-type hypersensitivity. This possibility has not yet been
studied, but deserves attention in light of the increasing incidence
of morbidity and mortality due to asthma, which is often triggered
by allergic responses (NIH, 1991; Danielle, 1988).
Acceleration of autoimmunity following UV exposure has also
been reported in mice. Both acute (2 hr/day, for 7 days) and chronic
(3 hr/wk, for 4 wks) exposure to 20 J m-2 s-1 enhanced mortality
in an autoimmune strain of mice (NZB X BZW F1). These exposures
also caused increased serum antibodies to single stranded DNA,
enhanced polyclonal B-cell activity in the spleen, and caused more
severe renal glomerular inflammatory changes (Ansel et al., 1985),
all hallmarks of autoimmunity. It is unclear whether there is a
relationship between these reactions and the effects of UV on
immunological processes described above.
7.2.5 Conclusions
Exposure of mice to UV radiation impairs certain immune
responses, enhances susceptibility to a variety of infectious agents
and UV-induced tumours. While fewer studies have been done in rats,
similar effects have been observed. Suppression of contact and
delayed type hypersensitivity responses have been studied most
extensively; however, several other cell mediated responses are also
affected. Suppression of these immune responses appears to be
mediated by release of soluble mediators from UB exposed skin which
alter antigen presentation by Langerhans (and other) cells such that
they fail to activate TH 1 cells. The resulting immune suppression
is antigen specific, can occur regardless of whether antigen is
applied at the site of exposure or not, and is relatively long
lasting.
Enhanced susceptibility to a variety of infections in mice and
rats corresponds to suppression of DTH responses to microbe specific
antigens. UV exposure also prevents the development of protective
immunity to these infections.
7.3 Ocular Studies
7.3.1 Introduction
The large number of animal studies enhances our understanding
of both acute and delayed ocular effect from UV exposure (Zigman
1993; Andley 1987; Dillon et al., 1990). Studies of the action
spectrum of delayed or permanent effects such as cataract or
retinopathy and UV damage mechanisms in the cornea, lens and retina
are only possible in animal models.
7.3.2 General effects
Photokeratitis is an acute reversible radiation-induced injury
of the corneal epithelium. It is analogous to acute sunburn of the
skin. Animal studies have clearly demonstrated that exposure to an
artificial source of UVB can lead to acute photokeratitis, and
action spectra have been determined (Cogan & Kinsey, 1946; Pitts et
al., 1977; Zuclich and Kurtin, 1977). Pitts (1974; 1978) estimated
the mean threshold of UVB (290-315 nm) for photokeratitis was 35 J
m-2 at 270 nm.
Damage to corneal endothelium has been reported in rabbits
(Doughty & Cullen, 1990). The effects on the corneal stroma have
also been observed in rabbits, where reversible damage to the
stromal keratocytes has followed exposure to UV (Ringvold &
Davangar, 1985). This histological study showed that keratocytes
disappear following UV exposure and later reappear. More recent
animal studies have reported the use of a UVB filtering contact lens
or the application of UVB absorbing chromophores to the cornea of
rabbit eyes protects against photokeratitis (Bergmanson et al.,
1988; Oldenburg et al., 1990).
7.3.3 Cataractogenesis
There is substantial evidence that exposure to UV induces
discrete opacities in the anterior lens of experimental animals.
Several studies have shown that UVB, but not UVA, induces anterior
opacities in animals (Bachem, 1956; Pitts et al., 1977; Jose &
Pitts, 1985; Söderberg 1990). Supra threshold doses of UVB at 300 nm
cause a Na-K shift between the lens and the surrounding (Söderberg
(1991). This leads to swelling and disruption of lens cells,
probably causing zones of deviating refractive index within the lens
observed as opacities (Söderberg 1989). Anterior lens opacities have
developed in albino mice after daily exposure for 1-2 months to
mixed UVA and UVB source (290-400 nm), but not when the source was
filtered to remove radiation <320 nm (Jose & Pitts, 1985). Anterior
cataract have also been produced in young albino mice exposed to
black light (predominantly UVA, with some UVB) (Zigman et al.,
1974). With more prolonged exposure of mice, cortical and posterior
subcapsular opacities have been induced (Zigman et al., 1975;
Jose, 1986). By contrast, a dose of 2 kJ m-2 applied every day
produced anterior polar cataract within 8 weeks. Extension of the
irradiation to 5 months created an opacity in the deeper cortex
(Weager et al., 1989). In testing possible cataractogenic efficacy
of UVA and UVB, UVB alone was cataractogenic, whereas, UVA showed
the same effect only when other cataractogenic factors were added
(e.g. X-ray) (Schmitt et al., 1988).
Ham et al. (1989) irradiated rhesus monkeys to 1 mW cm-2
daily for three years to UVA radiation (UVB radiation was carefully
excluded) and was unable to detect lenticular opacities.
7.3.4 Retinal effects
It is well accepted that short wavelength visible non-coherent
and coherent light, as well as UVA, can cause photochemical,
mechanical and thermal damage to the retina and the pigment
epithelium (review: Ham et al., 1984; Organisciak & Winkler,
1993). Aphakic monkeys revealed retinal lesions after exposure to
UVA (Guerry et al., 1985). Exposure of aphakic albino rats to UVA
created acute retinal lesions with 50-80% more effectiveness than
blue light (Rapp & Smith 1992). Mechanical and thermal lesions are
almost entirely limited to exposure to high energy laser light.
Photochemical damage can occur through two different light exposure
regimes: short term, high irradiance (minutes to hours) and
long-term (days to weeks) low irradiance (Kremers & van Norren,
1988; Remé et al., 1991; Rapp & Smith, 1992). Whereas damage
thresholds differ depending on the animal species, basic mechanisms
at the molecular and structural level are essentially the same (for
review: see Organisciak and Winkler, 1993). Photochemical lesions
are strongly dependent on wavelength, i.e. the absorbing chromophore
and its quantum efficiency, obey, at least within a certain time
frame, the reciprocity of intensity and exposure duration (e.g.
Kremers & van Norren, 1988).
Action spectra for high irradiance short exposure duration
lesions have been obtained in several studies (e.g. Ham et al.,
1984). Action spectra for prolonged exposure to relatively low light
levels are less clear and were first evaluated by Noell et al.
(1966). Prolonged exposure to low light levels may elicit secondary
tissue responses that may not be directly related to the initial
action spectrum. These tissues responses include acute cellular
necrosis and apoptotic cell death, an inflammatory response with
oedematous changes and macrophage invasion, and tissue proliferation
with scar formation and neovascularization (e.g. Hoppeler et al.,
1988; Yoshida et al., 1993). Action spectra, tissue responses,
preventive measures as well as enhancing factors, which are all
intensively investigated in many laboratories, are likely to be
similar to the human.
Studies of photochemical retinal injury in aphakic rhesus
monkeys have extended the action spectrum for short-wavelength light
damage down to 310 nm (Ham et al., 1984). These studies showed
that the retina, if exposed, is up to six times more vulnerable to
photochemical damage in the UV than in the visible. These results
are therefore of direct relevance to pseudophakic individuals
without a UV-protective lens, and may also indicate a contributory
role to light damage from the small fraction of UV that reaches the
retina of the normal eye.
8. HUMAN STUDIES: THE SKIN
8.1 Characteristics
8.1.1 Structure and optical properties
The skin is a large organ with an area of more than 1.5 m2 in
adults. It provides the first stage of protection for chemicals,
radiations, xenobiotics and also prevents the evaporation of water
and the loss of ions and proteins. The skin has developed specific
mechanisms for photoprotection and biological responses to UV as
discussed below.
Skin is composed of three very different parts (as shown in
figure 4.1): the epidermis, dermis and subcutaneous tissue. The
epidermis is the outermost layer of the skin and varies in thickness
from 50 µm to 600 µm (palmoplantor skin). The fibrous proteins known
as keratin are produced in the keratinocytes of the epidermis. It is
keratin that serves as a major protective tough substance in the
skin; hair and nails are composed almost entirely of keratin. The
daughter cells of the keratinocytes in the basal layer (stratum
basale) of the epidermis differentiate and become "prickle cells" of
the malpighian layer of the epidermis - the stratum malpighii. As
these cells migrate outward, changes continue to occur; granules
appear in the cytoplasm of each cell; the cells tend to flatten and
form the stratum granulosum. Still later, the cells lose their
nuclei, die, dehydrate and flatten out to form the tough stratum
corneum (horny layer). It is generally agreed that the entire
process of cell migration from the basal layer of the epidermis to
final shedding from the surface of the stratum corneum takes 28 days
in normal skin. Of this 28-day period, the cell spends about 14 days
in the epidermis and 14 days in the stratum corneum.
The skin contains millions of tiny glands, including: apocrine
(sweat) glands which discharge sweat into hair follicles, eccrine
(sweat) glands which carry saline from the dermis and subcutaneous
layer directly to the skin surface, and sebaceous glands which
secrete sebum - an oily substance which lubricates hairshafts and
maintains a slightly acidic, oily film over the stratum corneum. The
collection of eccrine sweat glands plays a major role in the body's
thermoregulatory mechanism, since evaporative cooling is the most
efficient means the body has for removing excess heat, as long as
the humidity is not 100%. More than two litres of sweat can be
discharged in one day. On average, the skin contains about as many
sebaceous glands as eccrine sweat glands, except that there are few
sebaceous glands on the palms of the hands and soles of the feet.
The dermis, or corium, is much thicker than the epidermis, but
consists of much larger cells. The dermis is largely connective
tissue which gives the skin its elasticity and supportive strength.
Nerve cells, blood vessels and lymphatic glands are found in the
outermost dermal layer - the papillary dermis. Unlike the epidermis,
the thickness of the dermis is not at all uniform throughout the
body; it varies from 1 mm to 4 mm.
The basal layer of the epidermis is delineated from the dermis
by a complex basement membrane. The keratinocytes are anchored to
the basement membrane by semi desmosomes. This is the only layer
where cell division takes place. The division of keratinocytes,
under normal conditions, occurs every 17 to 38 days. The division
rate may vary in different parts of the body.
Melanocytes are formed in the basal layer of the epidermis
loosely attached to the neighbouring keratinocytes (ratio 1
melanocyte for 46 keratinocytes). The melanocytes synthesize the
pigment melanin which matures into melanosomes. Then the melanosomes
are transferred to the keratinocytes where they are digested if
their size is below 1 cubic micron. Large melanosomes, as in dark
skin, are not digested but transferred intact to the stratum corneum
with the normal shedding of the epidermis.
Melanocytes are cells of nervous origin which have migrated
during the 10th week of embryonic life in the epidermis and in the
hair roots. These cells divide very slowly (one division every 3-5
years). There is a boost of division when new hairs are growing or
after exposure to UV light.
Chemically, melanins are bipolymers: red melanins contain
sulphurs and are soluble at PH 7.2, black melanins are insoluble.
The ratio of red and black melanins within a melanocyte is
genetically determined. Mixed types of melanins are deposited on a
protein matrix contained in the melanosomes. Both types absorb UV
and participate in the screening effect of the whole epidermis. Two
major steps in the melanin synthesis take place: the enzymatic
(tyrosinase) oxidation of the tyrosine to produce dopa
(di-hydroxyphenylalanine) and the spontaneous oxidation of dopa in
quinones.
Black melanin absorbs UV. In the presence of oxygen, it
produces a free radical which, in sufficient quantities, can be
deleterious to the melanocytes and the cellular environment,
including keratinocytes or in the dermis, fibroblasts and fibres.
A third type of cell is present in the upper layers of the
stratum Malpighi: the Langerhans cell. It is a migrating dendritic
cell which is able to recognise foreign or abnormal structures. This
cell plays a major role in immunological recognition and its
activity is very sensitive to UV. Its function is impaired by a UV
dose as low as one half of a minimal erythema dose.
The innermost layer of the skin is generally known as the
subcutaneous layer. It is composed largely of fatty tissue that
serves a shock-absorbing and insulating role. The thickness of this
layer varies considerably from one body region to another and from
one person to another.
Skin optics are governed by two basic processes, the absorption
and scattering of light. Absorption is the loss of a photon when its
energy is reduced within the atom or molecule, and more specifically
in a target species called a chromophore. UV absorption occurs in
wavelength bands where the molecules have characteristic absorption
spectra. The absorbed photon energy is dissipated as heat or
reemission of light when the excited molecules return to their
ground state, or the energy is spent on photochemical reactions.
Scattering is a process where the direction of propagation of UV is
altered, especially at boundaries of refractive index. Scattering
and absorption of the photons limit the depth of penetration of UV
in the skin (see section 4.2 and figure 4.1).
In the white skin, the change in the refractive index causes 5%
of the normally incident light to be reflected. The remaining 95% is
absorbed or reflected at the dermis. The back-scattered light is
reflected again at different interfaces. This phenomenon explains
why, in the epidermis and in the upper papillary dermis, the
strength of the light becomes several times that of the incident
light itself.
The transmission of UV through isolated epidermis is strongly
dependent on the chromophores contained in the structure. Aromatic
amino acids (tryptophan, tyrosin, phenyl-alanine) absorb strongly
near 275 nm, urocanic acid and melanins play the role of endogenous
sunscreens. The DNA absorbs 260 nm wavelengths. Melanin is the
unique chromophore with an absorption extending into the UVA and
visible regions.
8.1.2 Skin types
The sensitivity of skin to UV has been defined by six
phototypes: types I to IV are characteristics of caucasoid
populations; type V represents mongoloid Middle Eastern populations
(Fitzpatrick et al., 1974); and type VI represents African and
American negroid populations. The capacities to acquire natural tan
or to present naturally a deep pigmentation are keys to the response
to UV exposure. Among caucasians, there is a general correlation
between skin type and resistance to sunburn and capacity to tan.
8.2 Beneficial Effects
8.2.1 Vitamin D3
An established beneficial effect of UV exposure is the
synthesis of vitamin D3 (Adams et al., 1982). In adults, the
epidermis contains nearly 50% of the total concentration of
7-dihydrocholesterol in the skin. UVB exposure causes the
provitamins D3 to be isomerized to pre-vitamin D3. During continual
exposure to sunlight, the pre-vitamin D3 forms by
photo-isomerization the biologically inert photo-products lumisterol
and tachysterol. Once formed, the pre-vitamin D3 spontaneously
isomerizes into vitamin D3 (reaction with a maximum efficacy at
37°C), a more stable form. After crossing the basal membrane of the
epidermis, vitamin D3 is linked to a circulating alpha1-globulin:
vitamin D-binding protein. The protein linked vitamin D3 is
transformed to 25-OH-T3 which can be measured in the blood.
Transported to the kidney, it is metabolised to 1,25 dihydroxy
vitamin D3 which is the biologically active form.
Vitamin D3 is required for the intestinal absoprtion of calcium
(Davies 1985). After a single whole body exposure to one MED the
circulating vitamin D3 level increases by an order of magnitude (2
ng/ml to 24 ng/ml within 24 hours) and returns to normal levels
within a week (Holick 1985). Since active vitamin D3 is metabolised
by the kidney, it was expected that in the circulating blood it does
not change dramatically after repeated solar exposures. Chronically
reduced vitamin D3 synthesis may lead to a deficit in active vitamin
D3, as was found in elderly subjects (Omdahl et al., 1982).
The use of sunscreens was found to suppress cutaneous vitamin
D3 synthesis (Matsuoka et al., 1988). This was also found in
children, pregnant or lactating women and debilitated patients with
poor intestinal absorption. Widespread use of sunscreens could lead
to vitamin D deficiency in some groups and inadequate fixation of
calcium (Prystowsky 1988). More recently, receptors for the active
form for vitamin D3 were found in the keratinocytes of the
epidermis. Vitamin D3 inhibits the proliferation of cultured
keratinocytes and induces them to terminally differentiate (Smith
et al., 1986). The topical or oral administration of 1, 25-OH-D3
has proved to be effective for the treatment of psoriasis, in
replacement or in addition to the classical PUVA treatment. This is
a new approach for the treatment of this condition and a possible
explanation for the success of the heliotherapy (Morimoto and
Kunihiko, 1989).
For the entire system of vitamin D3 production the amount of UV
radiation reaching the skin is critical. The doses needed are small,
and daily exposures of the face and hands to sun and light for 15
minutes is considered sufficient. The minimum dose requirement was
estimated to be equivalent to 55 MED per year (Health Council of the
Netherlands, 1986). When too little UVB reaches the skin,
deficiencies of vitamin D may occur, resulting in a weakening of the
bones. Groups at risk are particularly dark-skinned children in high
latitude cities and elderly people living entirely indoors.
Supplementation of vitamin D3 in the diet is then recommended.
8.2.2 Skin adaptation
Another beneficial effect of modest exposure to UVB radiation
is the maintenance of the ability of the skin to sustain further UV
exposures. Loss of this adaptation forms an important component in
photodermatosis, skin diseases where the lesions are caused by
light. These patients can be treated by regular exposures to UVB.
The doses required are in the same range as that needed for the
synthesis of vitamin D3.
8.2.3 Other benefits
It has been suggested that beneficial effects of UV exposures
may occur such as: improvement of cardiopathy and functions, and
better microorganism defense. These effects have not been confirmed
in well designed studies. Bright light therapy for winter depression
is most likely to be the consequence of visible light stimulating
the ocular system. Because of the high illumination needed for this
treatment special care is needed to avoid the emission of UV light
by these sources (Terman et al., 1990).
Treatment of portwine stains and pigment dyschromia by lasers
are applications of selective absorption of some wavelengths by
specific chromophores contained in the lesion. Phototherapy by
specific dyes absorbed by tumour cells or specific structures, is
currently a growing field.
8.3 Acute Effects
8.3.1 Erythema and sunburn
In its mildest form, sunburn consists of a reddening of the
skin (erythema) that appears up to about 8 h after exposure to UV
and gradually fades after a few days. In its most severe form, it
results in inflammation, blistering, and peeling of the skin. The
degree to which a person will experience sunburn depends critically
on skin type. For fair-skinned people, the relative effectiveness of
UV for tanning and for erythema are approximately the same over the
entire UVB and UVA ranges of wavelengths (Parrish et al., 1982).
The most important factors that define if a dose of UV will
induce erythema are the wavelength of the radiation, the skin type,
and the pigmentation of the subject. UVA, UVB and UVC are all able
to induce the erythema.
The relative effectiveness of the different wavelengths to
induce erythema is expressed as an erythema action spectrum
(McKinlay and Diffey 1987). For minimal erythema, the most
erythemogenic wavelengths are in the 250-290 nm range and a decrease
in effectiveness is observed as the wavelengths increase. Erythema
occurs 3-5 hours after UV exposure and reaches a maximum intensity
between 8 and 24 hours, fading over 3 days. The vasodilatation of
capillary vessels within the papillary dermis can be observed before
the erythema becomes visible, and occurs in the same way for
children, adults and elderly. However, the exposure time required to
produce UVB erythema increases after about 60 years of age.
Histologic alterations from erythema are observed in the
photodyskeratotic keratinocytes as well as intercellular edema with
exocytosis (lymphocytes in the epidermis). Superficial vascular
plexus, endothelial cell enlargement, perivenular edema, red blood
cells in the capillary are characteristic features observed between
3 hours and 72 hours after UV exposure. Dermal neutrophils appear
immediately after irradiation reaching a peak level at 24 hours.
DNA may be the primary chromophore involved in the induction of
erythema (Ley, 1985, Wolf et al., 1993). Subsequently, a number of
inflammatory mediators are induced. Cutaneous blisters have been
used extensively to study these mediators. Prostaglandin (E2 and
F2) levels were elevated in these blisters within 6 hours, peaked
at 24 hours and returned to control levels by 48 hours after UV
exposure, although erythema persisted beyond that time. Indomethacin
suppressed prostaglandin formation, however, blood flow was only
slightly altered by this treatment suggesting that other factors
must play an important role in UVB induced inflammation (Greaves et
al., 1978). Similarly while elevated histamine levels have been
observed, antihistamines have little effect in diminishing UV
induced erythema (Gilchrest et al., 1981).
Finally, UV exposure causes keratinocytes, to release the
cytokines interleukin-1 (IL-1) and tumour necrosis factor alpha (TNF
alpha) both of which are potent mediators of inflammation (Oxholm
et al., 1988; Rasanen et al., 1989). UVB exposure also causes
up-regulation of adhesion molecule such as ELAM-1 which facilitates
inflammatory cell infiltration (Murphy et al., 1991).
Irradiation of human skin with 3 MED is associated with
increased levels of transforming growth factor, suggesting a role of
this molecule in keratinocyte proliferation, epidermal hyperplasia
and angiogenesis.
8.3.2 Skin pigmentation and tanning
When skin is exposed to UV, two distinct tanning reactions
ensue. Immediate pigment darkening (IPD) begins immediately on
exposure to UV and is caused by the darkening of the pigment melanin
that is already present in the skin; it is normally seen only in
people who have at least a moderate constitutive tan. Such
pigmentation begins to fade within a few hours after cessation of
exposure. UVA is regarded as being most effective for IPD.
Delayed tanning (melanogenesis) takes about three days to
develop and is more effectively produced by UVB than by UVA (Parrish
et al., 1982; Gange et al., 1985). Delayed tanning is more
persistent than IPD and results from an increase in the number, size
and pigmentation of melanin granules. Exposure to UVB results also
in an increase in the thickness and scattering properties of the
epidermis (outer layer of the skin). Because UVA does not produce
thickening of the epidermis, the tan obtained from it, while perhaps
cosmetically acceptable, is not as effective in protecting against
further exposure to UV as the equivalent pigmentation induced by
exposure to UVB or solar radiation.
8.3.3 Photosensitization
The use of certain medicines may produce a photosensitizing
effect on exposure to UVA as may the topical application of certain
products, including some perfumes, body lotions, etc. Many
medications and other agents contain ingredients that may cause
photosensitivity, which is defined as a chemically induced change
that makes an individual unusually sensitive to light. An individual
who has been photosensitized may develop a rash, sunburn, or other
adverse effect from exposure to light of an intensity or duration
that would normally not affect that individual.
Reactions to photosensitizing agents involve both photoallergy
(allergic reaction of the skin) and phototoxicity (irritation of the
skin) after exposure to ultraviolet radiation from natural sunlight
or artificial lighting (particularly from tanning booths). This
photosensitization of the skin may be caused by creams or ointments
applied to the skin, by medications taken orally or by injection, or
by the use of prescription inhalers.
In addition to an exaggerated skin burn, itching, scaling,
rash, or swelling, exposure to UV in combination with certain
medications may result in (FDA 1992); Skin cancer, Premature skin
aging, Skin and eye burns, Allergic reactions, Cataracts, Reduced
immunity and Blood vessel damage.
This can result in photoallergic or phototoxic reactions that
are accelerated by UV exposure. Phototoxic contact dermatitis often
occurs clinically as exaggerated sunburn but occasionally blisters
may also occur on the erythematous areas. Most phototoxic
sensitizers have an action spectrum in the UV from 280-430 nm.
Window glass which absorbs UV below 320 nm will protect patients
from phototoxic compounds with absorption below 320 nm, but fails to
protect against photosensitizers such as tar and psoralens which are
efficient at longer wavelengths. Examples of photocontact drugs and
substances are given below in table 8.1.
Table 8.1 Some photosensitizing substances
(CIE, 1990, FDA, 1992)
Sulphonamides
Salicylanilides
Coal-Tar derivatives
- acridine
- anthracene
- phenanthrene
Dyes
- anthraquinone
- eosin
- methylene blue
- rose bengal
Psoralens
Some fragrances
Cyclamate (artificial sweetener)
Non-steroidal anti-inflammatory drugs (pain reliever,
antiathritics)
Deodorant and bacteriostatic agents in soaps
Fluorescent brightening agent for cellulose, nylon or wool
fibres
Phenothiazines (major tranquilizers, anti-emetics)
Sulfonylureas (oral anti-diabetics, hypoglycemics)
Sunscreen ingredients
- 6-Acetoxy-2,4,-dimethly-m-dioxane (preservative)
- Benzophenones
- Cinnamates
- Para-aminobenzoic acid (PABA)
- Paba esters
Tetracyclines (antibiotics, anti-infectives)
Tricyclic antidepressants
The concentration of drugs needed to elicit a photoallergic
reaction is much lower than that needed to cause a phototoxic
reaction. On the other hand, photoallergic reactions occur only in a
small proportion of exposed individuals while phototoxic reactions
may occur in anyone, given sufficient exposure. Generally, no
clinical reaction occurs on first exposure to an agent causing
subsequent photoallergy. Even sunscreen agents used to protect
against photocontact dermatitis may be photoallergenic. A small
number of individuals who develop photocontact dermatitis may retain
a persistent reactivity to light (including UV) long after exposure
to the photosensitizing compound.
8.4 Chronic Effects on the Skin Other than Cancer
UV radiation causes a number of chronic degenerative changes in
the skin, mainly in caucasian populations, as a result of its action
on keratinocytes, melanocytes and components of the dermal stroma
including fibrous tissue (collagen and elastin) and blood vessels.
These changes include freckles (ephelides), melanocytic naevi,
lentigines, telangiectasia, skin wrinkling and atrophy, yellow
papules and plaques on the face, colloid milium (firm, small,
yellow, translucent papules on the face, forearms and hands),
diffuse erythema, diffuse brown pigmentation and ecchymoses
(Goldberg & Altman, 1984). These or related changes are sometimes
grouped into syndromes such as cutis rhomboidalis nuchae (thick,
yellow, furrowed skin, particularly on the back of the neck),
Favre-Racouchot syndrome (yellow, thick comedones and follicular
cysts of the periorbital, malar and nasal areas) and reticulated
poikiloderma (reddish brown reticulated pigmentation with
telangiectasia and atrophy and prominent hair follicles on the
exposed chest and neck). With the exception of freckles and
melanocytic naevi, these changes are also referred to collectively
as "photoageing" (Gilchrest, 1990) because of their association with
increasing age but presumed correlation with total accumulated
exposure to the sun rather than with age per se.
In the US National Health and Nutrition Examination Survey
(Engel et al., 1988), age-adjusted prevalence proportions of
senile elastosis, actinic (solar) keratosis, fine telangiectasia,
localized hypermelanism, senile (solar) lentigines and freckles, in
whites 1 to 74 years of age, were associated with lifetime exposure
to the sun as estimated by dermatologists.
Freckles and solar lentigines
Freckles and solar (also called senile) lentigines are
pigmented macules occurring on the sun exposed skin of caucasians.
Their prevalence is increased in those with highly sun-sensitive
skin (Azizi et al., 1988). Freckles occur most commonly in
children while the frequency of solar lentigines increases with age
and is greatest in those over 60 years of age (estimated at 75% in
the USA: Rhodes et al., 1991). They show similar histological
patterns: their are increased numbers of melanocytes and an
increased concentration of melanin in the basal layer of the
epidermis (Rhodes et al., 1991). Melanocytic atypia has been
observed in both. An increased risk of melanoma has been observed in
relation to freckling in childhood and an increased risk of
non-melanocytic skin cancer in relation to both freckling and
prevalence of solar lentigines (see sections 8.4.1 and 8.4.2).
Melanocytic naevi
Melanocytic naevi are benign proliferations of melanocytes
usually beginning in the basal layer of the epidermis and later
extending into the dermis. They are common in white populations and
rare in black and Asian populations (Armstrong & English, 1988;
Gallagher et al., 1991), are associated, in white populations,
with phenotypic indicators of constitutional sensitivity to the sun,
particularly fair skin colour (Green et al., 1988b; Gallagher et
al., 1990b), occur mainly on body sites that are maximally or
intermittently exposed to the sun (Kopf et al., 1978, 1985;
Augustsson et al., 1990; Gallagher et al., 1990a), occur more
commonly in Australian than British children (Green et al., 1988b)
and in persons born in Australia than immigrants who arrived in
Australia after about 15 years of age (Armstrong et al., 1986),
and somewhat inconsistently with measures of sun exposure, including
sunburn in early life (Armstrong et al., 1986; Gallagher et al.,
1990c; Coombs et al., 1992). They are associated with an increased
risk of cutaneous melanoma (see section 8.4.2).
Solar keratoses
Solar keratoses are benign proliferations of epidermal
keratinocytes. They are very common on exposed body sites in older
people in caucasian populations living in areas of high ambient
solar irradiance (see for example, Marks et al., 1983 and Holman
et al., 1984a). Solar keratoses have been reported to be
associated with phenotypic indicators of cutaneous sun sensitivity
(Vitasa et al., 1990), to be more common in people born in
Australia than in migrants to Australia (Goodman et al., 1984), to
be associated with estimates of total and occupational sun exposure
(Goodman et al., 1985; Vitasa et al., 1990), and to be
associated with other benign indicators of cutaneous sun damage
(Holman et al., 1984b; Green 1991). Their number on the skin is
strongly associated with risk of non-melanocytic skin cancer (see
section 8.4.1).
8.5 Cancer
Epidemiological evidence relevant to the effects of UV on risk
of cancer in humans derives mainly from study of the effects of sun
exposure (presumably solar UV but not separable from other solar
radiation) on cancer risk. There are four general lines of evidence
available from which it may be inferred that sun exposure causes any
particular cancer. They are that: the cancer in question occurs more
frequently in people who are sensitive to the sun, occurs mainly at
sun-exposed body sites, is increased in residents of areas of high
ambient solar irradiance, and is increased in people with high
personal sun exposure.These lines of evidence will not apply to all
cancers that may be linked to sun exposure. They form nonetheless, a
useful framework within which most of the relevant evidence can be
described. To this framework will be added a consideration of
evidence relating to artificial sources of UV when it is available.
The most direct evidence of the carcinogenicity of UV in humans
should come, in principle, from observation of the effects of
personal exposure to the sun. In practice, it is very difficult to
make measurements of personal sun exposure accurately. Most often
they are made by questionnaire and require recall of rather
non-salient details of life over 60 or more years. This is a very
difficult task (Kricker et al., 1993). A difficulty is presented
by the fact that people who have sun sensitive skin and are at
higher risk of skin cancer will tend to expose themselves less to
the sun. To obtain an accurate measure of the effects of personal
sun exposure, this confounding with sun sensitivity should be
controlled - this has not always been done. It may not be
surprising, therefore, that measures of personal exposure to the sun
have not been consistently associated with risk of cancers thought
to be related to the sun and that more indirect evidence has proved
to be stronger.
8.5.1 Nonmelanocytic skin cancer
Introduction
There are two major histopathological types of nonmelanocytic
skin cancer: basal cell carcinoma (BCC) and squamous cell carcinoma
(SCC). BCC is the commoner type in white populations.
The epidemiology of nonmelanocytic skin cancer is difficult to
describe accurately. Its routine recording is often not attempted by
cancer registries because of the large numbers of cases involved
and, if attempted, is invariably incomplete because of the rarity
with which primary nonmelanocytic skin cancers require hospital
treatment and the frequency with which probable nonmelanocytic skin
cancers are not sent for histopathological verification of the
diagnosis (Muir et al., 1987).
These difficulties have influenced the quality of the
epidemiological evidence relating nonmelanocytic skin cancer to sun
exposure. In addition, most of the earlier cross-sectional and
case-control studies of nonmelanocytic skin cancer , and some of the
more recent ones, (see, for example, table 11 in IARC, 1992) are
deficient in that the control series were rarely population-based,
appropriate effect measures or p values were often not estimated,
and confounding by age and sex were not controlled in the analysis.
In the narrative that follows, little reliance will be placed on
these studies which include those reported by Lancaster & Nelson
(1957), Gellin et al. (1965), O'Beirn et al. (1970), Urbach et
al. (1974), Aubry & MacGibbon (1985), O'Loughlin et al. (1985),
Herity et al. (1989), Hogan et al. (1989) and Gafa et al.
(1991).
The results of most of the studies referred to below have been
described and their results tabulated in detail in other
publications (e.g., IARC, 1992) thus only the immediately salient
features will be described here.
Sun-sensitivity
Race
Nonmelanocytic skin cancer is much less frequent in populations
with dark skins than those with light skins (Fitzpatrick & Sober,
1985; Hoffman, 1987; Urbach, 1987). Data from cancer registries
represented in Cancer Incidence in Five Continents Volumes II-VI
(Doll et al., 1970; Waterhouse et al., 1976, 1982; Muir et al.,
1987; and Parkin et al., 1992) in which direct comparisons of
nonmelanocytic skin cancer incidence in different ethnic groups can
be made within a single geographical area show that the incidence
rates in the light skinned populations are consistently the highest.
Similarly, in the 1977-78 US survey of nonmelanocytic skin cancer;
rates in whites were 232.6 per 100 000 person years compared with
3.4 in blacks (Scotto et al., 1983).
Available evidence suggests that BCC occurs less frequently
than SCC among dark skinned populations. The opposite is the case in
light skinned populations. BCC was the most common nonmelanocytic
skin cancer reported in South African whites in 1949-1975 but was
rare among black Africans, occurring mainly in albinos (Oettlé,
1963; Oluwasanmi et al., 1969; Rippey and Schmaman, 1972; Isaacson
et al., 1978). SCC, principally on the lower limb and associated
with previous trauma, was the commoner of the two in blacks (Oettlé,
1963; Oluwasanmi et al., 1969; Rippey and Schmaman, 1972; Isaacson
et al., 1978; Rose, 1973). Melanesians (Foster & Webb, 1988) and
Polynesians (Paksoy et al., 1991) had less BCC than SCC, while no
BCCs were reported in the Melanesians of North Samoa, a particularly
heavily pigmented people (Foster & Webb, 1988).
Ethnic background is an important determinant of risk of
nonmelanocytic skin cancer in Caucasians. In people of southern
European ethnic origin born in Australia, relative to other people
born in Australia, risk of BCC was 0.56 (95% CI 0.14-1.65) for those
with one southern European grandparent, 0.17 (0.00-1.05) for two and
0.00 (0.00-0.86) for three or four (p for trend 0.002; Kricker et
al., 1991a). No subject with SCC in this study had any southern
European grandparents (RR 0.00, 0.00-1.23).
Individual sun sensitivity
Among recent cross-sectional, case-control and cohort studies
several have reported significantly elevated relative risk (RR)
estimates for red or light hair colour with BCC (RRs between 1.5 and
2.9; Hunter et al., 1990; Green & Battistutta, 1990; Kricker et
al., 1991a) and SCC (RR=2.4; Kricker et al., 1991a). However, a
light complexion was significantly associated only with SCC (RR=3.3;
Kricker et al., 1991a), and high RRs (of 3.4) for light hair
colour with BCC and SCC were reported in only one study (Vitasa et
al., 1990).
Sensitivity of the skin to the sun, as measured by ability to
tan and susceptibility to sunburn, was more consistently related to
risk of BCC and SCC than hair, skin and eye colour. RRs of 2.0 or
more for BCC were found with a skin that burns rather than tans
(Marks et al., 1989; Vitasa et al., 1990; Hunter et al., 1990;
Kricker et al., 1991a). For SCC, the evidence of an increased risk
with sun-sensitive skin was somewhat weaker, with RRs around 2.0 and
95% confidence intervals (CI) that were wide and included 1.0, in
the two studies that reported RRs adjusted for age, sex, and other
relevant confounders (Vitasa et al., 1990; Kricker et al.,
1991a).
Xeroderma pigmentosum and albinism
Xeroderma pigmentosum (XP) is a recessively inherited syndrome
characterised by clinical and cellular hypersensitivity to solar
radiation and a defect in the capacity to repair UV-induced damage
in DNA (Fitzpatrick et al., 1963, Cleaver, 1973). Evidence of
cutaneous sun damage may appear as early as 1-2 years of age in the
absence of specific protection from the sun, and skin cancers are
very frequent (Kraemer et al., 1987). In a survey of 830 cases of
XP located through published case reports, 45% were reported to have
had skin cancers (Kraemer et al., 1987). The median age of
diagnosis of the first skin cancer was 8 years. Ninety seven per
cent of BCC and SCC were on constantly exposed sites, that is, the
face, head, and neck, compared with an estimated 80% in the general
US population. In 220 XP patients in whom the number of skin cancers
was stated, half had more than two cancers and 5% had more than 30;
79 patients were described as having BCC and 112 SCC (Kraemer et
al., 1987).
Albinism is an inherited disorder of melanin metabolism with a
decrease in or complete absence of melanin; as a result, the skin of
albinos is highly sensitive to the sun. The most common type of
albinism occurs in 1 in 15,000 American blacks, 1 in 40,000 European
or American Caucasians, and has estimated frequencies as high as
1:3,900 in Soweto, South Africa (Kromberg and Jenkins, 1982), and 1
in 1,000 in Nigeria (Cervenka et al., 1979). African albinos have
been reported to have a high incidence of SCC and a somewhat lower
rate of BCC (Cervenka et al., 1979; Luande et al., 1985;
Kromberg et al., 1989).
Body-site distribution
It is commonly stated that the site distributions of
non-melanocytic skin cancers, particularly SCC, correspond well to
what would be expected from the exposure of different body sites to
the sun. They do generally conform to this pattern with more than
60% of lesions occurring on the head and neck (Kricker et al.,
1993). It is consistently observed, however, that the proportion of
SCC on the upper limbs is higher than that of BCC while BCC has the
higher proportion (some 10% or more) on the trunk (Scotto et al.,
1983; Osterlind et al., 1988a; Giles et al., 1988; Glass &
Hoover, 1989; Levi et al., 1988; Karjalainen et al., 1989;
Kricker et al., 1990; Gallagher et al., 1990a; Roberts, 1990;
Magnus, 1991; Serrano et al., 1991; Kricker et al., 1993). At
the sub-site level, BCCs are almost completely absent on the heavily
exposed backs of hands, and infrequent on the forearms compared with
the upper arms (Brodkin et al., 1969; Scotto et al., 1983;
Goodman et al., 1984; Kricker et al., 1990); in addition, this
subsite distribution on the face is not highly correlated with the
distribution of UV erythema on the face (Diffey et al., 1979).
Residence in areas of high ambient solar irradiance
Geographical variation
Annual incidence rates of nonmelanocytic skin cancer in 29
populations of mainly Western European origin in Cancer Incidence in
Five Continents, volume 6 (Parkin et al., 1992) show little
evidence of any consistent relationship between incidence and
latitude. The highest incidence rates were in the populations of
Tasmania, Australia (42°S) (213.2 per 100,000 in men and 113.1 per
100,000 in women) and British Columbia, Canada (49°N) (134.1 in men
and 91.2 in women), and were also comparatively high in Switzerland
(47°N) (78.7 in men and 50.2 in women) and Southern Ireland (53°N)
(71.5 in men and 48.0 in women).
In contrast to these international patterns, incidence rates of
nonmelanocytic skin cancer within countries do appear to increase
with proximity to the equator as indicated by broad place of
residence, latitude, or measures of solar irradiance. Geographical
variation in nonmelanocytic skin cancer incidence in the USA has
been described in three National Cancer Surveys (Mountin & Dorn,
1939; Dorn, 1944a, 1944b; Auerbach, 1961; Haenszel, 1963; Scotto et
al., 1974) and several related studies (Scotto et al., 1982,
1983; Serrano et al., 1991). Incidence of all types increased with
increasing proximity to the equator, with similar gradients for men,
women and all ages. The same pattern is seen in Australia (Marks et
al., 1993)
Migrants
The nonmelanocytic skin cancer experience of light-skinned
migrants from areas of low to areas of high ambient solar irradiance
has generally been consistent with an effect of sun exposure on skin
cancer incidence. In Australia, incidence and mortality were found
to be lower in migrants, most of whom had come from the UK, an area
of lower sun exposure, than in those born in Australia (Armstrong
et al., 1983; Giles et al., 1988). Kricker et al. (1991a)
examined in some detail the relationship between BCC and SCC and
migration to Australia. Migrants (excluding those from southern
Europe who may be at lower constitutional risk for skin cancer) had
a lower risk of BCC and SCC (RRs around 0.3) than did those born in
Australia. In addition, for BCC (which had sufficient cases to
analyze) the RR in those who migrated in the first 10 years of life
(1.05) was the same as that in those born in Australia (1.0) but
then fell to low levels in those who migrated later in life (RR
about 0.2). Hunter et al., (1990) observed an association between
risk of BCC and residence in southern parts relative to elsewhere in
the USA (RRs of 1.6 for residence in California and 2.1 for
Florida).
Personal sun exposure
Total sun exposure
Reported measurements of total current or accumulated sun
exposure in studies of nonmelanocytic skin cancer are likely to be
subject to substantial error not only because of the difficulties in
recalling sun exposure over periods of 60 years or more (Kricker et
al., 1993) but also because of the use of broad summary variables
such as "estimated average daily outdoor exposure" (Gellin et al.,
1965). The two studies which attempted a more quantitative measure
found no evidence of a positive association between estimates of
total sun exposure and risk of BCC (RRs of 1.0 or less) (Hunter et
al., 1990; Vitasa et al., 1990). Only one of these studies
included SCC (with only about 50% confirmed histopathologically);
for sun exposure above the 75th centile the estimated RR was 2.5
(95% CI 1.2-5.4) (Vitasa et al., 1990).
Occupational sun exposure
Early clinical reports associated skin cancer with outdoor
occupation (Molesworth, 1927; Blum, 1948; Emmett, 1973). However,
the relationship between occupation and skin cancer has been
examined adequately in relatively few studies and the evidence for
an association is weak. At the population level, the best conducted
studies which attempted to classify occupational sun exposure on the
basis of occupational title found only small differences in skin
cancer incidence between outdoor and indoor workers. Several
population studies reported associations with employment in
agriculture (Atkin et al., 1949; Whitaker et al., 1979; Teppo
et al., 1980) and outdoor employment generally (Beral & Robinson,
1981; Vågerö et al., 1986) but their interpretation is complicated
by incomplete and potentially biased case ascertainment (Vågerö et
al., 1986; Teppo et al., 1980), inclusion of other cancers
(Atkin et al., 1949), and confounding between occupation and
social class (Beral & Robinson, 1981).
There have been few well-conducted studies of occupational sun
exposure at the individual level. They show a generally consistent
but not strong association between BCC and SCC and various crude
measures of outdoor employment, e.g., "indoors", or "outdoors"
occupation. The RRs have generally been under 2.0 for BCC (Hogan et
al., 1989; Marks et al., 1989; Green and Battistutta, 1990; Gafa
et al., 1991). One study reported a high RR for SCC with an
"outdoors" occupation but with an extremely wide confidence interval
(Green and Battistutta, 1990). Other studies found RRs close to 1.0
(Aubry and MacGibbon, 1985, Hogan et al., 1989; Marks et al.,
1989; Gafa et al., 1991) and only one appeared to be statistically
significant (Hogan et al., 1989).
Recreational exposure
The relationship of exposure to sunlight in non-working hours
(largely recreational sun exposure) with nonmelanocytic skin cancer
has been described in only two studies. One study reported an RR
below 1.0 for BCC in those with "mainly outdoor leisure" and a
positive but not statistically significant association for SCC: RR
of 3.9 (95% CI 0.5-30.9) (Green & Battistutta, 1990). In the study
of Aubry & MacGibbon (1985), the highest category of a
"non-occupational exposure score" showed an elevated risk of SCC (RR
1.6; p-value=0.07).
Sunburn
Risk of BCC was significantly increased in subjects with a
history of sunburn in one of two recent studies. Hunter et al.
(1990) observed an increasing risk with increasing frequency of
painful sunburns (RR 2.91, 95% CI 2.37-3.58, for 6+ occasions).
Green and Battistutta (1990), on the other hand, found no
discernible association between BCC and number of painful sunburns.
The risk of SCC, however, was increased with any history of painful
sunburn in this study: RRs 3.3 (95% CI 0.9-12.3) for 1-5 sunburns
and 3.0 (0.7-12.2) for 6+.
Other sun-related skin conditions
Cutaneous microtopography (a measure of loss of the fine skin
markings on the backs of the hands presumed due to loss of dermal
collagen; Holman et al., 1984a), prevalence of solar elastosis of
the neck, solar telangiectasia and solar lentigines and a history of
solar keratoses have been taken as indicators of a high level of
total accumulated exposure to the sun in those who have BCC or SCC.
The evidence that sun exposure causes these conditions, however, is
no stronger, and may be weaker, than the evidence that it causes
BCC, SCC or cutaneous melanoma.
Increasingly severe sun-related skin damage as measured by
cutaneous microtopography was associated with increasing risks of
both BCC (RR 3.1, 95% CI 1.5-6.4 for the highest grade) and SCC (RR
1.8, 95% CI 0.8-4.2 for the highest grade) (Kricker et al.,
1991a). Other indicators of sun damage to the skin (e.g. freckles,
telangiectasia, and elastosis) were also strongly related to risk of
both BCC and SCC, whether considered separately (Green and
Battistutta, 1990; Kricker et al., 1991a) or together (Holman et
al., 1984a; Green et al., 1988a), and including when adjusted
for cutaneous sun sensitivity (Kricker et al., 1991a). The RRs
associated with the presence of 40 or more solar keratoses were 10.4
(95% CI 5.8-18.8) for BCC and 34.3 (95% CI, 14.0-84.0) for SCC
(Kricker et al., 1991a).
Exposure to artificial sources of UV
Any use of a sunlamp was associated with a statistically
significantly increased risk of SCC in the study of Aubry & McGibbon
(1985) but three other studies (O'Loughlin et al., 1985; Herity
et al., 1989; Hogan et al., 1989) found no association with
"artificial sunlight" or use of sunlamps or sunbeds.
8.5.2 Cutaneous Melanoma
Introduction
Cutaneous melanoma began to be studied epidemiologically in
1948 when it was separated from other primary malignancies of the
skin in the 6th Revision of the International Statistical
Classification of Diseases (WHO, 1948), and Eleanor MacDonald
published the first population-based study, covering 272 incident
cases of melanoma in Connecticut (MacDonald, 1948). Because of its
likely fatality if not treated, melanoma is much more likely to come
to medical attention and to be diagnosed histopathologically than
nonmelanocytic skin cancer. It has, therefore, been more readily
recorded on a population basis than nonmelanocytic skin cancer and
better studied epidemiologically. Thus, while generally considered
less clearly associated with sun exposure, a series of large and
well-conducted studies in different countries over the last 10 years
have served to clarify this issue.
Sun-sensitivity
Race
Melanoma is a disease primarily of light-skinned populations
and occurs much less frequently in people with darker skins. In the
United States, the incidence in whites is ten fold or more higher
than in blacks living in the same areas (Parkin et al., 1992).
Rates are lowest (less than 0.7 per 100 000 person years) in parts
of Asia (eg Japan, Singapore, India, China, Philippines) while in
Los Angeles, USA, incidence was less than 1 per 100 000 in Japanese
and Chinese people compared with slightly higher rates, around 2.0,
in blacks (Parkin et al., 1992).
Among Caucasians, ethnic background is an important determinant
of melanoma incidence. The incidence is substantially lower among
Hispanics than among other whites in the United States. For example,
the incidence among Hispanics in New Mexico is less than 2 per
100,000 person years, but in other whites it is about 11 per 100,000
(Muir et al., 1987). In several case-control studies, subjects
with a southern or eastern European background had substantially
lower risks than those of northern European or United Kingdom
origins (Elwood et al., 1984; Holman and Armstrong, 1984a; Graham
et al., 1985).
Individual sun sensitivity
Blue eyes, fair or red hair, and pale complexion in people of
European origin are well established risk factors for melanoma.
These pigmentary characteristics were documented in most melanoma
case-control studies (see table 16 in IARC, 1992 and, as examples,
Elwood et al., 1984, Holman & Armstrong, 1984a, and Green et al.,
1985). Relative risk for light skin colour ranged from a little more
than 1.0 to about 3. Compared with those with dark brown to black
hair, those with fair hair generally had a less than two-fold
increase in risk, but those with red hair usually had a 2 to 4-fold
increase in risk. Eye colour was generally a weak risk factor with
relative risks less than 2, and what increased risk there was
generally disappeared after adjustment for the other traits.
Increased risks of melanoma in those with a reduced ability to
tan and an increased tendency to sunburn were observed in all
case-control studies in which these sun-sensitivity measures were
examined (see, for example, Elwood et al., 1984, Holman &
Armstrong, 1984a, and Green et al., 1985).
Xeroderma pigmentosum and albinism
In a review of reports of 378 patients with XP in which cancer
was mentioned, 37 patients had a melanoma (Kraemer et al., 1987).
African albinos have been reported to have a low rate of
melanoma (Cervenka et al., 1979; Luande et al., 1985; Kromberg
et al., 1989). Levine et al. (1992) state that there have been
only 16 documented cases of melanoma in albino patients reported in
the English language literature. Since albinos generally have a
normal number of melanocytes (Dargent et al., 1992), the rarity of
melanoma in them may indicate that melanin plays an important role
in the genesis of this cancer.
Body site distribution
The distribution of melanoma appears to favour sites which are
less heavily sun exposed, that is the back and face in Caucasian men
and the lower limbs in women (Crombie, 1981). However, when
whole-population series of cancer cases have been examined and body
surface area taken into account, sites that were usually covered by
clothing had lower rates than those usually exposed, with the
exception of the forearms and hands for which the rates per unit
area were low (Elwood & Gallagher, 1983; Green et al., 1993). The
anatomic site distribution of melanoma in blacks is quite different
from that in Caucasians with the majority of melanomas on the soles
of the feet (Higginson and Oettlé, 1959; Lewis, 1967; Fleming et
al., 1975).
Residence in areas of high ambient solar irradiance
Geographical variation
Internationally, the incidence of melanoma varies over
100-fold. Among countries included in Cancer Incidence in Five
Continents, Volume VI, the lowest rates reported around 1983-87 were
0.1-0.2 per 100 000 person years in China (Qidong), Japan (Osaka),
India (Bombay), and in Kuwaitis in Kuwait, while the highest were
about 25 per 100 000 person years in parts of Australia (Parkin et
al., 1992). In the USA and Australia, which have reasonably
homogeneous populations of mainly European origin distributed across
a wide latitude range, melanoma incidence increases with increasing
proximity to the equator or with increasing measured, annual ambient
UV irradiance (Scotto & Fears, 1987; Jones et al., 1992).
A simple latitude gradient for melanoma is not evident in
Europe. Armstrong (1984) showed that the incidence of melanoma in
Europe decreased from about latitude 35°N to a minimum around 55o N
and then rose with increasing latitude because of high rates in
Scandinavian and Scottish populations. The gradient of risk for
melanoma from north to south in northern Europe matches the
increasing natural pigmentation of the skin and may also, in part,
be due to differing patterns of sun exposure, particularly
recreational and vacation sun exposure.
Migrants
If a person migrates in childhood to a country of high ambient
solar radiation such as Australia, Israel, and New Zealand, their
risk of melanoma is observed to be similar to that in those born in
the country to which they migrate, whereas if they migrate after
this age their risk is substantially less than that in the
locally-born population (Katz et al., 1982; Cooke and Fraser,
1985; McCredie & Coates 1989; Steinitz et al., 1989; Khlat et
al., 1992). Overall, British immigrants to Australia and New
Zealand, where the populations are of predominantly British origin,
had incidence and mortality rates of melanoma of about a half the
levels in those born in these countries (Cooke and Fraser, 1985;
Khlat et al., 1992).
Residence history
Studies of individual lifetime ambient solar radiance, based on
latitude, location (eg, tropical, near coast), or average sunshine
hours of all places of residence are consistent in showing an
increased risk of melanoma with increased average radiance (Green &
Siskind, 1983; Holman et al., 1984b; Graham et al., 1985;
Osterlind et al., 1988b; MacKie et al., 1989; Weinstock et al.,
1989; Beitner et al., 1990). RRs for the highest category of
ambient solar radiance were 2.8 for mean annual hours of bright
sunlight at places of residence in Australia (Holman et al.,
1984b), between 1.4 and 2.6 for a southerly latitude in the USA
(Graham et al., 1985; Weinstock et al., 1989), or a tropical or
Mediterranean residence (MacKie et al., 1989; Beitner et al.,
1990). RRs differed greatly for residence near the coast in Denmark
(RR=1.7; Osterlind et al., 1988b) and Australia (RR=5.0; Green &
Siskind, 1983).
Personal sun exposure
Lifetime total exposure
Inconsistent results have been obtained in studies in which
lifetime total sun exposure were assessed by questionnaires. RRs for
the highest exposure category were between 0.6 and 5.3 (see table 18
in IARC, 1992). Two of five studies showed a statistically
significant positive association with RRs of 3.4 and 5.4 (Grob et
al., 1990; Lê et al., 1992) while one study showed a significant
negative association (RR 0.6; Graham et al., 1985). The other two
RRs were 1.1 (Dubin et al., 1986) and 5.3 (95% CI 0.8-30.8; Green,
1984).
Usual or recent total exposure
In this context, total means exposure from all sources (i.e.
occupational and non-occupational) rather than the total over some
period of time. This concept has been measured in a variety of ways.
For example, it has been common to enquire about present "usual"
exposure to the sun or usual exposure in some time period in the
fairly recent past. The evidence of any effect of such exposures on
risk of melanoma is weak. Of five studies, four had relative risks
for the highest categories of exposure ranging from 0.7 to 1.2
(Elwood et al., 1985b; Dubin et al., 1986; Holman et al.,
1986; Cristofolini et al., 1987). The remaining study found a
relative risk of 2.5 (p < 0.001) for the highest category of
exposure of average daily sun exposure 10-20 years ago (Rigel et
al., 1983).
Occupational exposure
Four studies of melanoma have shown statistically significant
positive associations with estimated lifetime occupational exposure
to the sun (see table 20 in IARC, 1992) with RRs ranging up to 6.0
(95% CI, 2.1 to 17.4) for outdoor employment (Paffenbarger et al.,
1978; Dubin et al., 1986; Garbe et al., 1989; Grob et al.,
1990). On the other hand, four studies showed statistically
significant results in which the RR for the highest category of
exposure was < 1.0 (Elwood et al., 1985b; Holman et al., 1986;
Osterlind et al., 1988b; Beitner et al., 1990). The remaining
studies have shown RRs around 1.0.
Recreational exposure
Recreational exposure to the sun has generally been measured by
either the type of recreational activity or the frequency or
duration of outdoor recreation (see table 22: in IARC, 1992). Some
studies have also recorded sun exposure during vacations separately
from other recreational sun exposure. At least one statistically
significant positive association was found in 11 of 16 studies in
which recreational exposure was considered (Adam et al., 1981; Lew
et al., 1983; Rigel et al., 1983; Elwood et al., 1985b; Dubin
et al., 1986; Holman et al., 1986; Osterlind et al., 1988b;
Zanetti et al., 1988; Beitner et al., 1990; Grob et al., 1990;
Nelemans et al. 1993). The RRs in the highest categories of
exposure were generally between 1.5 and 2.5. Only one study has
shown a negative association with recreational exposure: MacKie &
Aitchison (1982), in Scotland, observed an RR of 0.4 (95% CI
0.2-0.9) for the highest category of hours a week in outdoor
recreation. Their statistical model, however, included socioeconomic
status and history of sunburn, both of which may measure sun
exposure; thus overadjustment of the estimate of RR is likely. All
measures of number or frequency of vacations in sunny places were
positively associated with risk of melanoma. The RRs for the highest
category of this variable ranged from 1.2 to 5 and the finding was
statistically significant in six of eight studies (Lew et al.,
1983; Elwood et al., 1985b, Osterlind et al., 1988b, Zanetti et
al., 1988; Beitner et al., 1990; Nelemans et al., 1993).
Sunburn
Fifteen of seventeen studies of melanoma in which sunburn was
recorded (see table 23 in IARC, 1992) showed a statistically
significant, moderately to strongly positive association with a
history of sunburn (MacKie & Aitchison, 1982; Lew et al., 1983;
Elwood et al., 1985a; Green et al., 1985; Sorahan & Grimley,
1985; Elwood et al., 1986; Holman et al., 1986; Holly et al.,
1987; Osterlind et al., 1988b; Zanetti et al., 1988; MacKie et
al., 1989; Weinstock et al., 1989; Beitner et al., 1990;
Elwood et al., 1990; Nelemans et al., 1993) (see table 23: IARC,
1992). Twelve of these studies showed RRs greater than 2.0 for the
highest category of sunburn. Positive associations were obtained for
both sunburn in childhood and sunburn at any age. The greater
consistency of the relationship of melanoma with sunburn compared to
that with other exposure variables may indicate a specific
association with sunburn per se or simply that sunburn is a more
accurately measured indicator of sun exposure.
Presence of other sun-related skin conditions
Cutaneous microtopography
Severity of sun damage to the skin as measured by cutaneous
microtopography was strongly associated with risk of melanoma in
Western Australia. The RR for the most severe grade of damage was
2.7 (95% CI 1.4-5.0; p-value for trend=0.003; Holman et al.,
1984b). There was no similar relationship, however, in a study in
Denmark, in which similar methods were used (Osterlind et al.,
1988b).
Freckling of the skin has been shown to be associated with an
increased risk of melanoma in several studies (see for example,
Elwood et al., 1986, 1990, Osterlind et al., 1988b; Dubin et
al., 1986).
Solar keratoses and other skin cancers
The risk of melanoma was significantly increased in association
with a past history of nonmelanocytic skin cancer with RRs of 3.7
(95% CI 2.1-6.6) in Australia (Holman et al., 1984b) and 3.8
(1.2-12.4) in the USA (Holly et al., 1987). Relative risks for a
history of solar keratoses or "actinic tumours on the face" were
similarly high (Green & O'Rourke, 1985; Dubin et al., 1986).
Exposure to artificial sources of UV
Several studies have reported increased risks of cutaneous
melanoma in users of sunlamps or sunbeds (IARC, 1992). In the most
recent of these studies (Walter et al., 1990), the relative risks
for any use of sunbeds or sunlamps were 1.9 (95% CI 1.2-3.0) in men
and 1.5 (95% CI 1.0-2.1) in women. Relative risk increased with
increasing duration of use: for more than 12 months use the relative
risks were 2.1 (0.9-5.3) in men and 3.0 (1.1-9.6) in women. Positive
associations were found in two of four other case-control studies in
which these exposures were studied (Swerdlow et al., 1988; MacKie
et al., 1989; see also IARC, 1992). Sun exposure is a potential
confounding variable in studies of sunlamps and sunbeds but has not
been taken into account. Specifically it was considered by Walter
et al. (1990). Exposure to a group of other artificial sources of
UV, including plan printers, laboratory equipment emitting UV,
insect tubes, black lights and photocopiers, was not associated with
melanoma in one case control study in Australia (Holman et al.,
1986) but for a similar group plus welding the RR was 2.2 (95% CI
1.0-4.9) in Canada (Elwood et al., 1986). Siemiatycki (1991) found
no association between arc welding or other occupational exposure to
UV in a study of occupation and cancer in Canada.
A number of case-control studies have examined the association
between melanoma and exposure to fluorescent lighting. Only the
first such study found a statistically significant positive result
(Beral et al., 1982). Six subsequent studies found little or no
evidence of a positive association (IARC, 1992).
8.5.3 Cancer of the lip
Cancer of the lip is defined as cancer of the vermilion border
and adjacent mucous membranes and thus excludes cancers of the skin
of the lip (WHO, 1977). Most are SCC and occur on the lower lip
(Keller, 1970; Lindqvist, 1979), a site more heavily exposed to the
sun than the upper lip (Urbach et al., 1966). The inclusion of
adjacent oral mucosa in the definition of cancer of the lip raises
the possibility of confounding of tobacco and alcohol use (well
established causes of cancer of the mouth; Tomatis, 1990).
The incidence of cancer of the lip is much more common in men
than women. Among men in the USA, it is some 20 times more common in
whites than blacks and it is rare in black and Asian populations
worldwide (Parkin et al., 1992). Incidence of and mortality from
cancer of the lip are substantially lower in migrants to Australia
and Israel, who come from places with potentially lower sun
exposure, than those born in these countries (Armstrong et al.,
1983; McCredie & Coates, 1989; Steinitz et al., 1989).
The incidence of cancer of the lip is higher in rural than
urban areas (Doll, 1991) and descriptive studies have consistently
reported higher incidence or mortality rates of this cancer in men
with outdoor occupations such as farmers, agricultural labourers and
fishermen. (Atkin et al., 1949; Clemmesen, 1965; Gallagher et
al., 1984; Olsen & Jensen, 1987; Lynge & Thygesen, 1990). Four
case-control studies of varying quality have examined the
association between outdoor work and cancer of the lip. Keller
(1970) compared 301 men with cancer of the lip with 301 oral cancer
controls. Crude RRs for employment as a farmer or in any outdoor
work were 4.0 and 2.6 respectively. The use of patients with oral
cancer as controls allowed a rough adjustment for confounding with
tobacco and alcohol use in this study. Spitzer et al. (1975)
compared 339 men with cancer of the lip with 199 matched population
controls. The relative risk for any outdoor work was 1.52 and, for
employment as a fisherman, 1.50 (p < 0.05 in both cases). Pipe
smoking but not alcohol drinking was controlled in the analysis. The
other two studies also showed a positive association between outdoor
work and cancer of the lip (Lindqvist, 1979; Dardanoni et al.,
1984) but did not control for smoking and alcohol use and showed
other methodological difficulties (IARC, 1992).
8.5.4 Ocular cancers
Ocular melanoma and other ocular cancers are dealt with in
Chapter 10.
8.5.5 Other cancers
Observations of increasing mortality with increasing latitude
have formed the basis of suggestions that cancers of the breast,
colon and prostate may be prevented by increasing exposure to the
sun (Garland & Garland, 1980; Gorham et al., 1989, 1990; Garland
et al., 1989, 1990; Hanchette & Schwartz, 1992), perhaps through
proposed anti-carcinogenic actions of vitamin D (Eisman et al.,
1980; Colston et al., 1989; Schwartz & Hulka, 1990; Ainsleigh,
1993). Variation with latitude in medical care, death certification
practices, diet and other lifestyle factors are alternative
explanations for the latitude gradients in these cancers. These
observations, therefore, can be taken only to raise hypotheses about
indirect anti-carcinogenic effects of UV which require testing by
other means.
8.5.6 Action spectrum
There are few data from which the action spectrum of UV
carcinogenesis in humans can be inferred. For practical purposes,
therefore, reliance is placed on action spectra which have been
determined in experimental animals (see chapter 7).
If, as is argued below, it is accepted that the formation of
cyclobutylthymidine dimers is a step in the production of mutations
that are associated at least with BCC and SCC, then it would be
reasonable to accept the action spectrum for production of thymidine
dimers in humans as possibly indicative of the action spectrum for
production of nonmelanocytic skin cancer (figure 8.1). This action
spectrum was determined by Freeman et al. (1989) who irradiated
untanned gluteal skin of 30 caucasian volunteers with 1 or 2 MED of
narrow band UV at 275 nm, 282 nm, 290 nm, 296 nm, 304 nm, 334 nm and
365 nm. At least five subjects were irradiated at each wavelength.
Following irradiation, shave biopsies were taken of the irradiated
skin, DNA extracted and incubated with Micrococcus luteus
endonuclease to determine the frequency of endonuclease sensitive
sites which indicate the presence of thymidine dimers. Figure 8.1
shows that the effectiveness of UV in producing dimers increased
from 275 nm to a peak at 296 to 304 nm and then fell by four orders
of magnitude to the lowest measured level at 365 nm. This fall is
quite consistent with that observed in the relative effectiveness of
UV for carcinogenesis in mouse skin over the same wavelength range.
The authors reported, also, that observations had been made at 385
nm and 405 nm and that these wavelengths were "largely ineffective"
in net production of dimers.
Some indication of the effectiveness of UVA in causing skin
cancer could be obtained from the experience of people exposed to
high doses of UVA from sunbeds or in tanning salons. There is some
evidence to suggest that these exposures are associated with an
increase in risk of nonmelanocytic skin cancer, cutaneous melanoma
and ocular melanoma (see above). In none of the studies carried out,
however, has exposure to mainly UVA sources been distinguished from
sources that emit both UVA and UVB. In addition, confounding with
sun exposure is a possibility in all these studies.
8.5.7 Dose-response
In theory, the best way to estimate the dose-response
relationship between UV and cancer in humans would be to measure the
lifetime exposure to the sun of each member of a population and
relate this measurement, by way of a cohort or a case-control study,
to their probability of developing skin cancer. In practice this is
very difficult. First, measurement of lifetime exposure to the sun
generally requires recall of amount and pattern of sun exposure and
use of protective measures against the sun for as long as 40 or more
years in the past.
Second, exposure in early life may be particularly important in
determining risk of skin cancer, at least, and this is to a large
extent outside the range of accurate recall. Third, while problems
of recall could be solved by conduct of a cohort study, such a study
would require measurements of sun exposure beginning in the first
few years of life and repeated periodically throughout life to the
age at which UV-related cancers become common. Such a study would be
logistically and economically impracticable. The whole process is
complicated by the fact that a simple measure of lifetime exposure
may be insufficient for adequate description of dose-response. The
evidence that period and pattern of exposure may be important
determinants of some UV-related cancers (see below) means that to
model incidence correctly as a function of exposure it would be
necessary to collect data on dose-rate of UV and its variation
throughout life.
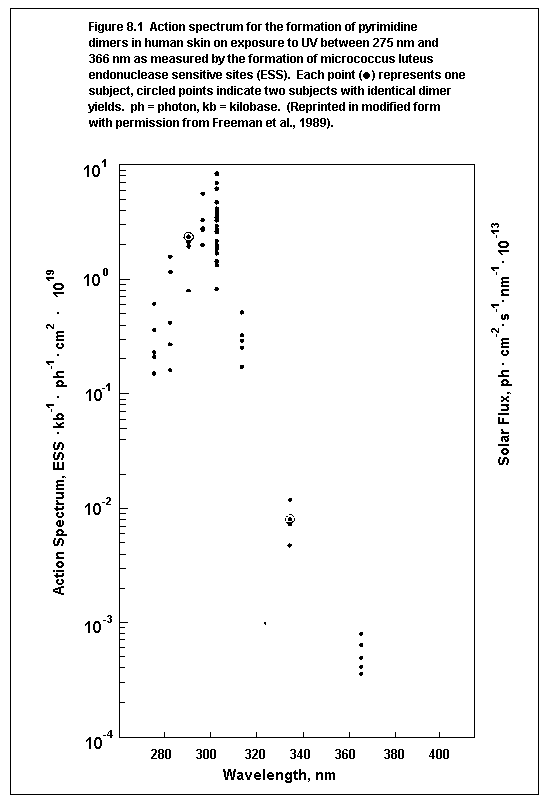 One attempt has been made to estimate the dose-response
relationship between UV and SCC and BCC in a retrospective cohort
study of Chesapeake Bay watermen (Strickland et al., 1989). The
watermen work mainly in traditional fishing tasks that are regulated
by law and have undergone few changes in the past 80 years. They
presented, therefore, better prospects than most populations for the
accurate recall of past sun exposure. Briefly, 808 caucasian
subjects among 1 250 licensed watermen aged 30 years or more
participated in an interview and skin examination. Present skin
cancers were identified at examination and a history of past skin
cancer was taken and; as far as possible, histological confirmation
of the diagnosis was obtained for both present and past skin cancer.
Totals of 47 SCC in 35 subjects, 60 BCC in 33 subjects and 344 solar
keratoses in 202 subjects were identified. About a half of the SCC
and BCC had been diagnosed before the survey examination while over
90% of the solar keratoses were diagnosed at the examination.
Histological confirmation was available for at least one lesion in
51% of subjects with SCC, 72% with BCC and 13% with a solar
keratosis (Vitasa et al., 1990). The average annual exposure of
facial skin to solar UVB was estimated for each subject by the
combination of data from a personal history of mainly occupational
outdoor exposure from 16 years of age to the interview, estimates of
ambient UV radiance in the area in which the watermen lived and
worked, based on the US network of Robertson Berger meters (Berger &
Urbach, 1982), and field measurements of individual exposure to UVB
under working conditions as recorded by polysulphone film
dosimeters. Annual average ambient UVB for the area was estimated at
about 1.14 x 106 J m-2 or 3260 MED assuming 350 J m-2 to be
equivalent to 1 MED. Personal annual average exposure of facial skin
ranged from about 1% to 8% of ambient UVB, i.e., 33 to 260 MED a
year.
The dose-response relationships obtained are shown in figure
8.2. They were derived from the following mathematical model:
prevalence = (exposure)a (age)b
or, equivalently,
log (prevalence) = a log (exposure) + b log (age).
One attempt has been made to estimate the dose-response
relationship between UV and SCC and BCC in a retrospective cohort
study of Chesapeake Bay watermen (Strickland et al., 1989). The
watermen work mainly in traditional fishing tasks that are regulated
by law and have undergone few changes in the past 80 years. They
presented, therefore, better prospects than most populations for the
accurate recall of past sun exposure. Briefly, 808 caucasian
subjects among 1 250 licensed watermen aged 30 years or more
participated in an interview and skin examination. Present skin
cancers were identified at examination and a history of past skin
cancer was taken and; as far as possible, histological confirmation
of the diagnosis was obtained for both present and past skin cancer.
Totals of 47 SCC in 35 subjects, 60 BCC in 33 subjects and 344 solar
keratoses in 202 subjects were identified. About a half of the SCC
and BCC had been diagnosed before the survey examination while over
90% of the solar keratoses were diagnosed at the examination.
Histological confirmation was available for at least one lesion in
51% of subjects with SCC, 72% with BCC and 13% with a solar
keratosis (Vitasa et al., 1990). The average annual exposure of
facial skin to solar UVB was estimated for each subject by the
combination of data from a personal history of mainly occupational
outdoor exposure from 16 years of age to the interview, estimates of
ambient UV radiance in the area in which the watermen lived and
worked, based on the US network of Robertson Berger meters (Berger &
Urbach, 1982), and field measurements of individual exposure to UVB
under working conditions as recorded by polysulphone film
dosimeters. Annual average ambient UVB for the area was estimated at
about 1.14 x 106 J m-2 or 3260 MED assuming 350 J m-2 to be
equivalent to 1 MED. Personal annual average exposure of facial skin
ranged from about 1% to 8% of ambient UVB, i.e., 33 to 260 MED a
year.
The dose-response relationships obtained are shown in figure
8.2. They were derived from the following mathematical model:
prevalence = (exposure)a (age)b
or, equivalently,
log (prevalence) = a log (exposure) + b log (age).
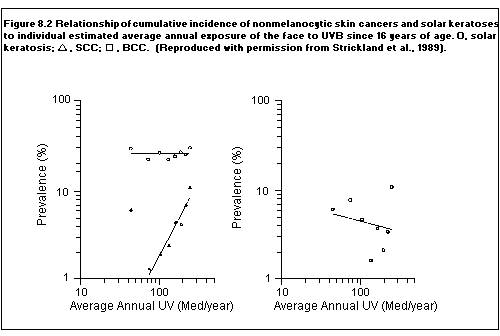 While prevalence is stated in the above expressions and the y
axis in figure 8.2 is labelled as prevalence, it is believed that
the figures are cumulative incidence proportions in which each
subject with more than one of a particular type of lesion was
counted only once. Only cumulative incidence of SCC appeared to
increase with increasing estimated exposure to solar UVB. The value
of a for SCC was 1.7. It should be noted however, that 4 subjects
with SCC among 64 men in the lowest sun exposure category were
excluded from the regression because of the anomalously high
incidence in this group and because of the possibility that these
subjects may have been hypersusceptible to UV. Apart from all having
solar keratoses, however, they did not consistently show phenotypic
features of high susceptibility to UV and it was not possible on any
basis, therefore, to remove hypersusceptible subjects from the
higher dose categories. The values of a for BCC and solar
keratoses were -0.2 and 0.005 respectively. It was suggested that
because of the high sun exposure of all men in the study the
cumulative incidence of BCC may have reached a saturation point at
the lowest level of sun exposure such that further increases in
exposure did not lead to further increases in incidence.
An alternative approach to estimating dose-response
relationships has been to examine the relationship, geographically,
between incidence or mortality of skin cancers in whole populations
and estimated or measured ambient UV (Armstrong, 1993). This work
has been done in the context of estimating the increase in skin
cancer that might be expected from some increment in ground-level UV
caused by depletion of stratospheric ozone. It has made the
fundamental assumption that the observed geographical variations in
incidence or morbidity rates of skin cancer is due largely to
geographical variations in UV. The results have commonly been
expressed in terms of the biological amplification factor (BAF)
defined as follows (de Gruijl and van der Leun, 1980).
There are substantial uncertainties regarding the dose-response
relationships shown in figure 8.2. First, the estimates of dose were
based on exposure of the face only, excluded dose in the first 16
years of life and did not take account of dose-rate or pattern of
exposure. Second, there is likely to have been appreciable
underascertainment of cancers diagnosed before the survey and high
proportions of all lesions were not confirmed histopathologically.
Third, the exclusion of four men with SCC must have had an
appreciable effect on the value of a for SCC. Fourth, no mention
is made of having controlled in the analysis for potential negative
confounding between cutaneous sun sensitivity and sun exposure. Such
confounding might be expected to reduce the slope of the
dose-response relationships. Finally, no measure of precision was
given for the values of a; given the small numbers of subjects with
SCC and BCC, those for these two cancers must have had quite wide
95% confidence intervals. This study gives only a hint, therefore,
as to what the dose-response relationship between solar UVB and
nonmelanocytic skin cancer might be at one end of the human exposure
range. No similar observations are available for cutaneous melanoma.
BAF = (dI/I)/(dD/D)
where dI equals a small increment in the existing incidence of skin
cancer, I, which results, in the steady state, from a small
increment dD in the existing biologically effective ambient level,
D, of solar radiation (i.e., spectral dose weighted by the action
spectrum for production of skin cancer).
The best known and most commonly used geographical
relationships between skin cancer incidence and ambient UV used for
dose-response estimation are those established from data on
nonmelanocytic skin cancer collected in a special survey in the USA
in 1977 and 1978 and UV measurements collected through the US
network of Robertson-Berger meters (Scotto et al., 1983).
Corresponding relationships were later established between melanoma
incidence measured by the US SEER cancer registries and the
Robertson-Berger meter data (Scotto and Fears, 1987). The
relationship for melanoma is shown in figure 8.3. In this figure,
the logarithm of age-adjusted annual incidence of melanoma is
plotted against the logarithm of the annual average Robertson-Berger
meter counts at each location. The lowest annual average meter count
was 95 x 104 units at Seattle and the highest, Albuquerque, New
Mexico, at 197 x 104 units. These values translate, approximately,
into 0.75 x 106 and 1.56 x 106 J m-2 respectively. The
estimated slopes of the regression lines were 0.7 in males and 0.8
in females which are also the estimated BAFs for melanoma from these
data.
While prevalence is stated in the above expressions and the y
axis in figure 8.2 is labelled as prevalence, it is believed that
the figures are cumulative incidence proportions in which each
subject with more than one of a particular type of lesion was
counted only once. Only cumulative incidence of SCC appeared to
increase with increasing estimated exposure to solar UVB. The value
of a for SCC was 1.7. It should be noted however, that 4 subjects
with SCC among 64 men in the lowest sun exposure category were
excluded from the regression because of the anomalously high
incidence in this group and because of the possibility that these
subjects may have been hypersusceptible to UV. Apart from all having
solar keratoses, however, they did not consistently show phenotypic
features of high susceptibility to UV and it was not possible on any
basis, therefore, to remove hypersusceptible subjects from the
higher dose categories. The values of a for BCC and solar
keratoses were -0.2 and 0.005 respectively. It was suggested that
because of the high sun exposure of all men in the study the
cumulative incidence of BCC may have reached a saturation point at
the lowest level of sun exposure such that further increases in
exposure did not lead to further increases in incidence.
An alternative approach to estimating dose-response
relationships has been to examine the relationship, geographically,
between incidence or mortality of skin cancers in whole populations
and estimated or measured ambient UV (Armstrong, 1993). This work
has been done in the context of estimating the increase in skin
cancer that might be expected from some increment in ground-level UV
caused by depletion of stratospheric ozone. It has made the
fundamental assumption that the observed geographical variations in
incidence or morbidity rates of skin cancer is due largely to
geographical variations in UV. The results have commonly been
expressed in terms of the biological amplification factor (BAF)
defined as follows (de Gruijl and van der Leun, 1980).
There are substantial uncertainties regarding the dose-response
relationships shown in figure 8.2. First, the estimates of dose were
based on exposure of the face only, excluded dose in the first 16
years of life and did not take account of dose-rate or pattern of
exposure. Second, there is likely to have been appreciable
underascertainment of cancers diagnosed before the survey and high
proportions of all lesions were not confirmed histopathologically.
Third, the exclusion of four men with SCC must have had an
appreciable effect on the value of a for SCC. Fourth, no mention
is made of having controlled in the analysis for potential negative
confounding between cutaneous sun sensitivity and sun exposure. Such
confounding might be expected to reduce the slope of the
dose-response relationships. Finally, no measure of precision was
given for the values of a; given the small numbers of subjects with
SCC and BCC, those for these two cancers must have had quite wide
95% confidence intervals. This study gives only a hint, therefore,
as to what the dose-response relationship between solar UVB and
nonmelanocytic skin cancer might be at one end of the human exposure
range. No similar observations are available for cutaneous melanoma.
BAF = (dI/I)/(dD/D)
where dI equals a small increment in the existing incidence of skin
cancer, I, which results, in the steady state, from a small
increment dD in the existing biologically effective ambient level,
D, of solar radiation (i.e., spectral dose weighted by the action
spectrum for production of skin cancer).
The best known and most commonly used geographical
relationships between skin cancer incidence and ambient UV used for
dose-response estimation are those established from data on
nonmelanocytic skin cancer collected in a special survey in the USA
in 1977 and 1978 and UV measurements collected through the US
network of Robertson-Berger meters (Scotto et al., 1983).
Corresponding relationships were later established between melanoma
incidence measured by the US SEER cancer registries and the
Robertson-Berger meter data (Scotto and Fears, 1987). The
relationship for melanoma is shown in figure 8.3. In this figure,
the logarithm of age-adjusted annual incidence of melanoma is
plotted against the logarithm of the annual average Robertson-Berger
meter counts at each location. The lowest annual average meter count
was 95 x 104 units at Seattle and the highest, Albuquerque, New
Mexico, at 197 x 104 units. These values translate, approximately,
into 0.75 x 106 and 1.56 x 106 J m-2 respectively. The
estimated slopes of the regression lines were 0.7 in males and 0.8
in females which are also the estimated BAFs for melanoma from these
data.
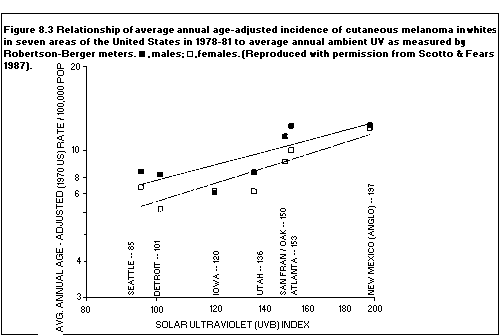 Table 8.2 summarises the most recent estimates of the BAFs for
nonmelanocytic skin cancer (BCC and SCC) and cutaneous melanoma from
data in the USA and Scandinavia. These estimates were based on
various action spectra: the response spectrum of the Robertson-
Berger meter (Scotto et al., 1983; Scotto & Fears, 1987; Pitcher &
Longtreth, 1991), the CIE reference spectrum of McKinlay and Diffey
(1987), Moan et al. (1989), Moan and Dahlbach, (1992), and the
Sterenberg-Slaper action spectrum for UV radiation of skin cancer in
albino hairless mice (de Gruijl and van der Leun, 1991). Generally,
the BAF for BCC lay between about 1.5 and 2.0. For SCC and melanoma,
there was much greater variation in estimates. Those for SCC were
between about 2.0 and 4.0 when based on the US data but between 1.0
and 2.0 when based on Norwegian data. Similarly, BAFs for melanoma
were between 0.3 and 0.5 when based on US data but between about 1.0
and 3.0 when based on Scandinavian data. While an attempt had been
made to adjust the US estimates for confounding with population
constitutional sensitivity to the sun and sun-related behaviour,
even the unadjusted estimates (ranging from 0.6 to 1.0; Scotto &
Fears, 1987; Pitcher & Longstreth, 1991) were considerably less than
those for Scandinavia. The most likely reasons for the differences
between the estimates is differences in their error.
Table 8.2 Recent estimates of the biological amplification factors (BAF) for
nonmelanocytic skin cancer and cutaneous melanoma based on geographical correlations
between average annual ambient UV and skin cancer incidence or mortality.
Authors Region Sex BAFs for BCC BAFs for SCC
Incidence Incidence
Nonmelanocytic skin cancer
Scotto et al, 1983 USA (8 centres) M 1.3-2.6a 2.1-4.1
F 1.1-2.1 2.2-4.3
de Gruijl & van der Leun 1991 USA (8 centres) MF 1.4 2.5
Moan et al 1989 Norway (6 areas) M 1.5-2.0a 1.2-1.5
Melanoma
Scotto & Fears, 1987 USA (7 areas) M 0.4c
F 0.5c
Pitcher & Longstreth, 1991 USA 215 SMAs M 0.4d
F 0.3d
Table 8.2 (contd).
Authors Region Sex BAFs for BCC BAFs for SCC
Incidence Incidence
Moan & Dahlback, 1992 Norway M 1.9
F 3.2
Finland M 1.3
F 2.2
Sweden M 1.9
F 2.3
a Exponential model used in which the value of the BAF varies with ambient UV, thus range
of values given.
b Same data as used by Scotto et al (1983) except that a power model was used instead of
an exponential model and the most recent action spectrum for UV carcinogenesis in mouse
skin was assumed.
c Adjusted for population estimates of ethnic origin, pigmentary characteristics, use of
sunscreens, and hours per week of outdoor exposure.
d Adjusted for population estimates of ethnic origin, household income, outdoor occupation
and education.
This approach to estimation of dose-response has assumed, among
other things, that: the correct action spectrum has been used to
weight spectral UV irradiance when producing a single figure for
ground level UV in each area; all members of the populations giving
rise to the incidence rates have lived their whole lives in their
present environment; the skin cancer incidence rates have been
measured accurately and, in particular, that their error does not
correlate with ambient UV radiance; and that possible confounding of
ambient UV with constitutional sensitivity to the sun and
sun-related behaviour is either unimportant or has been taken
adequately into account. None of these assumptions is likely to be
correct in any of the estimates of BAF made so far and the estimates
are all likely, therefore, to be inaccurate (Armstrong, 1993).
It should be noted that even if dose-response relationships
between UV and skin cancer incidence determined at the population
level are accurate, they may not reflect dose-response relationships
at the individual level. This is because of the averaging of
exposure and risk that occurs at the population level. The actual
population exposure of the skin at any level of ambient UV is an
average of many different exposures and the actual population
incidence of skin cancer is an average of many individual risks of
skin cancer. The association of individual risk to individual
exposure may be quite complex (as has been postulated for melanoma;
Armstrong, 1988) but at the population level this could still
translate into a simple exponential or power relationship between
ambient exposure and incidence.
No attempts have been made to estimate the dose-response
relationships for UV and cancers other than skin cancers that may be
caused by UV.
8.5.8 Effects of pattern of exposure
There are a number of aspects of the epidemiology of cutaneous
melanoma in European populations that appear inconsistent with a
simple relationship between it and sun exposure (English &
Armstrong, in press). First, in many populations, melanoma occurs as
commonly in women as in men, although men are more likely to work
outdoors; second, there is a relative peak in incidence in middle
life, which is not the pattern to be expected from life-long
exposure to an environmental agent; third, it most frequently occurs
on the back in men and lower limbs in women, sites which are not
maximally exposed to the sun; finally, melanoma is more common in
indoor than outdoor workers and in those of higher socioeconomic
status than those of lower status.
These observations led to postulation of the "intermittent
exposure hypothesis" for the relationship of sunlight to melanoma.
This hypothesis states that incidence of melanoma is determined as
much (or more) by the pattern of sun exposure as by the total
accumulated dose of sun exposure and, specifically, that infrequent
or intermittent exposure of untanned skin to intense sunlight is
particularly effective in increasing incidence of melanoma
(Armstrong, 1988). If, as seems plausible, this postulated effect of
intermittency of sun exposure is due to protective thickening and
pigmentation of the epidermis with more continuous sun exposure,
then intermittent exposure would be maximally effective if it
occurred at intervals longer than the time it takes for the skin to
return to its prior level of sensitivity after a single episode of
sun exposure (two to three weeks).
The epidemiological evidence, summarised above, that risk of
melanoma is apparently not increased with total or occupational
exposure to the sun but is increased with increasing
non-occupational exposure to the sun, in particular increasing time
spent in sun-related vacations in populations with otherwise
comparatively low sun exposure (Klepp & Magnus, 1979; Lew et al.,
1983; Elwood et al., 1985b; Osterlind et al., 1988b; Beitner
et al., 1990; Zanetti et al., 1992), and episodes of sunburn
(IARC, 1992) is consistent with this hypothesis.
There are less grounds for believing that pattern of UV is
important in causing nonmelanocytic skin cancer than there are for
melanoma. Specifically, nonmelanocytic skin cancer as a whole, and
BCC and SCC separately, are more common in men than women (Kricker
et al., 1990; Parkin et al., 1992; Marks et al., 1993) and, in
the largest, well-collected, population-based incidence series
available, they showed steadily increasing incidence rates with age,
except for a downturn in older age-groups possibly due to
underascertainment (Scotto et al., 1983). On the other hand,
recent series of BCC have shown unexpectedly high proportions on the
trunk (Kricker et al., 1990; Magnus, 1991) and, as reviewed above,
the relationship of nonmelanocytic skin cancer with occupational
exposure to the sun is not at all clear. Moreover, there is some
evidence of an association between sunburn and other indicators of
intermittent exposure and BCC (Hunter et al., 1990; Kricker et
al., 1991b).
8.5.9 Interactions between UV and other agents
5-methoxypsoralen and 8-methoxypsoralen (methoxsalen)
Methoxsalen, 5-methoxypsoralen and some other psoralens
(hereafter referred to collectively as psoralens) are used in
combination with UVA (referred to as PUVA) for the treatment of
psoriasis and to produce repigmentation in vitiligo. A working group
of International Agency for Research on Cancer concluded in 1987
that there was sufficient evidence that PUVA (defined as
8-methoxypsoralen plus long-wave UV radiation) causes cancer in
humans, based mainly on evidence of its association with SCC.
Recent studies have confirmed and extended this conclusion.
Stern & Lange (1988) reported on 5 to 10 year follow-up of patients
treated with PUVA in the USA. In 13 384 person years of follow-up of
1 380 people, 100 people had developed 391 SCC and 94 had developed
218 BCC. There were statistically significantly increasing risks of
both BCC and SCC occurring more than 58 months after entry into the
cohort with increasing number of treatments with PUVA. Risk of BCC
rose to 6.9 (95% CI 3.2-13.1) with 260 or more treatments and risk
of SCC rose to 50.1 (95% CI 24.9-89.5). Therapeutic exposure to
ionizing radiation, high doses of tar and UVB was unrelated to
number of treatments with PUVA and, therefore, did not confound the
observed dose-response relationship. Lindelöf et al. (1991)
reported on follow-up for an average of 6.9 years of 4 799 Swedish
patients treated with PUVA. Risk of SCC increased with increasing
number of treatments with PUVA. In those who received 200 or more
treatments, the relative risk, with reference to the general Swedish
population, was 30.7 (14.7-56.5) in men and 18.5 (0.5-103.2) in
women. BCC were not recorded in this study.
Cases of melanoma have been reported in patients treated with
PUVA but the rates of melanoma in US and Swedish PUVA cohorts were
little higher than those expected on the basis of population
incidence rates. In the US cohort the relative risk was 1.46 (95% CI
0.3-7.3; Gupta et al., 1988), while in the Swedish cohort it was
1.1 (0.2-3.2) in men and 0.8 (0.1-3.0) in women (Lindelöf et al.,
1991).
It is not possible in any of these studies to estimate what the
risk of skin cancer would have been if the patients had been treated
with UV alone. It would seem reasonable to assume, however, on the
basis of the evidence from experimental animals (see above) that
administration of psoralens potentiated the effect of UV. In
addition analysis of clinical data even suggests that this
carcinogenicity of PUVA exceeds that of UVB treatment by an order of
magnitude (Slaper, 1987).
Ionizing radiation
Ionizing radiation causes nonmelanocytic skin cancer, mainly
BCC, generally after a long latent period (Shore, 1990; Fragu et
al., 1991; Hogan et al., 1991; Sadamori et al., 1991; Moller
et al., 1993). Incidence in the atomic bomb survivors was related
strongly to exposure, although not for those closest to the blast
(Sadamori et al., 1991). There is evidence that ionizing radiation
and UV may act synergistically in causing skin cancer. In a study of
skin cancer following irradiation of the scalp for ringworm, Shore
et al. (1984) found a substantial excess of BCC in white subjects.
In blacks, no skin cancers were observed when 13 would have been
expected from the rate on the UV-shielded scalp in whites. This lack
of skin cancers in irradiated blacks suggest that UV radiation
contributes more than additively to the production of these cancers
in irradiated whites. This inference is supported by the observation
that the excess of BCC around the UV-exposed fringe of the scalp in
irradiated whites was some four to five times higher per unit dose
and area than that on the UV-shielded hairy scalp (Shore et al.,
1984). Further, Modan et al. (1993) observed, in a separate cohort
of people irradiated for ringworm of the scalp, that sunbathing
increased the risk of skin cancer 2.6 times (95% CI 1.1-6.1). All
except one of 39 cancers were BCCs and 60% were on the scalp, thus
suggesting that the ionizing radiation had also contributed to their
causation. However, no formal analysis of interaction was reported
from this study.
Other agents
One or more SCCs occur in 30% to 50% of patients with
epidermodysplasia verruciformis, a rare chronic skin disease in
which there are multiple benign skin lesions caused by a variety of
different types of human papilloma viruses (HPV; Quan & Moy, 1991).
HPV DNA sequences have been identified in the SCC occurring in these
patients and most of the cancers occur on sun-exposed sites. This
coincidence suggests that the UV interacts with the virus in causing
these cancers. Benign warts are also increased in frequency in renal
transplant patients and it is possible that they, with UV, are
involved in causing the excess of nonmelanocytic skin cancers that
occur in these patients (Bouwes-Bavinck, 1992). These cancers, too,
occur mainly on sun-exposed sites.
8.5.10 Mechanisms of UV carcinogenesis
Precursor lesions
Solar keratoses are very strongly related to risk of
nonmelanocytic skin cancer (Kricker et al., 1991a) and it is a
matter of clinical observation that SCC often occurs in lesions
which, previously, had the clinical appearances of solar keratoses.
There is evidence that solar keratoses are, themselves, caused by
exposure to the sun. Thus solar keratoses could be the visible
expression of an early, UV-caused, mutational step in the pathway to
skin carcinogenesis.
Analogously with solar keratoses, benign melanocytic naevi are
commonly observed in clinical or histopathological contiguity with
melanomas and have been found, epidemiologically to be strongly
related to risk of cutaneous melanoma (Holman & Armstrong, 1984a;
Armstrong & English, 1988). Naevi may, therefore, represent an
early, UV-caused, mutational step in the development of melanoma. In
this regard, it is interesting to note that the most active
development of naevi occurs during the first 15 years of life, thus
providing a possible link to early life sun exposure as suggested by
migrant data (see above). The appearance of naevi in children who
have received chemotherapy for cancer may indicate that they can be
caused by mutagens other then UV (Hughes et al., 1989; Baird et
al., 1992; Green et al., 1992).
DNA damage
The evidence of much increased incidence rates of BCC, SCC
cutaneous melanoma and tumours of the anterior eye in patients with
xeroderma pigmentosum (XP) (see above) strongly suggests that
unrepaired, UV-induced photoproducts in DNA form part of the genesis
of these cancers. Almost all forms of XP have an inherited
deficiency in excision repair of DNA photoproducts (Kraemer et al.,
1987). The position is made somewhat less clear, however, by the
existence of two other inherited syndromes involving deficiency in
DNA excision repair, trichothiodystrophy and Cockaynes syndrome,
that are not associated with increased incidence rates of skin
cancer (Bridges, 1990; Barrett et al., 1991).
There is evidence also that low levels of DNA repair may be
associated with skin cancer in the general population, although
there is some disagreement in the results so far obtained .
Munch-Petersen et al. (1985) studied 29 patients with multiple
skin neoplasms, 19 with multiple BCC only and 10 with both BCC and
squamous lesions (SCC, solar keratosis, or Bowen's disease), and 25
control subjects with roughly the same age and sex distributions.
Patients with both BCC and squamous lesions had much higher
UV-induced DNA synthesis in lymphocytes, as measured by
incorporation of [3H]thymidine following irradiation with a Philips,
TUV, 6 watt lamp (peak emission at 254 nm), than did controls.
Patients with multiple BCC were not different from controls. The
authors attributed this difference to possible defects in the DNA
ligation process. Roth et al. (1987) studied the rate of loss of
dithymidine dimers in DNA of cultured fibroblasts that had been
irradiated with a Philips 6-V germicidal lamp with peak emission at
254 nm. Dimers were measured with a monoclonal antibody highly
specific for the conformational change in DNA caused by the dimers.
No significant difference was seen between 16 BCC cases and 30
controls in the rate of loss of dimers. In ten melanoma patients,
however, the mean percentage of bound antibody at 60 minutes was
50.5 (SD 18.2) compared with 29.8 (SD 5.7) in controls (p=0.001)
thus suggesting that average dimer repair capacity was less in the
melanoma patients than the controls. Alcalay et al. (1990)
compared 22 patients with a history of one (13 patients) or more BCC
in the past with 19 healthy volunteers. Each subject was irradiated
at two locations on the lower back with 1 MED of UV from a 150 W
xenon arc solar UV simulator from which wavelengths < 295 nm had
been removed by a 1 mm WG 320 filter. Shave biopsies of skin were
taken from one of the sites immediately and the other 6 hours after
irradiation and the concentrations of pyrimidine dimers in DNA
measured by the micrococcus luteus endonuclease assay. Dimer yields
immediately after irradiation were similar in BCC patients and
controls but, after 6 hours, the proportion of dimers that had been
repaired was 22% in the BCC patients compared with 33% in the
controls (p=0.06). Wei et al. (1993) measured DNA repair capacity
in 88 patients with a history of one or more histologically
confirmed BCCs and 135 control subjects. Repair capacity was
measured by assaying the capacity of lymphocytes to repair
UV-induced damage in DNA of a nonreplicating recombinant plasmid.
Damage was induced by irradiating the plasmid with either 3500 J
m-2 or 7000 J m-2 of UV at 254 nm. The BCC cases had a
significantly lower mean DNA repair capacity (p=0.047) than controls
only when 29 controls with a family history of BCC or who themselves
had an actinic keratosis were removed from the comparison.
It now seems highly likely that UV can mutate the p53 tumour
suppressor gene in human skin and that UV-induced mutations in this
gene may be involved in the aetiology of some human cancers. First,
a recent study has shown that mutations that are strongly suggestive
of a UV effect, CC-TT changes at dipyrimidine sites, could be found
in 17 of 23 (74%) samples of sun-exposed normal skin in Australian
skin cancer patients compared with 1 of 20 (5%) samples of skin from
sites not exposed to the sun (Nakazawa et al., in press). Focal
overexpression of p53 protein has also been observed in normal, sun
exposed skin adjacent to BCC in 37 of 39 subjects but only in one
keratinocyte in 14 samples of buttock skin from the same subjects
(Shea et al., 1992). Second, several studies have found mutations
at dipyrimidine sites in the p53 gene in 36% to 56% of BCC (Rady et
al., 1992; Molès et al., 1993; Ziegler et al., 1993). In three
similar studies of SCC one found these mutations in 14 (57%) of 24
cases (Brash et al., 1991) one in two of 10 cases (Pierceall et
al., 1991) and, the other, none of 13 (Molès et al., 1993). The
more detailed findings of these five studies are summarised in table
8.3. Mutations were found in the p53 gene, following examinations of
varying extent, in 46% of BCC and SCC, the majority of which (88%)
were base substitutions or one or two base deletions at dipyrimidine
sites (the proportion expected by chance is 75%). The highest
proportions of these mutations resulted from C -> T, CC -> TT and
C -> A base changes which is consistent with what would be expected
if they had been caused by UV (Brash et al., 1991). Several
studies have found p53 protein by immunostaining (evidence of
overexpression of possibly abnormal p53 protein) in from 42% to 83%
of BCC and 56% to 65% of SCC (Barbareschi et al., 1992; Shea et
al., 1992; Stephenson et al., 1992, Ro et al., 1993) and are
thus reasonably consistent with the observations as to prevalence of
mutations.
The position with respect to p53 mutation is much less clear
for other cancers possibly caused by UV. A number of studies have
reported on the prevalence of detectable p53 protein in primary
melanomas: the proportions have varied from 3.6% to 97% (see, for
example, Stretch et al., 1991; Akslen & Morkve, 1992; Lassam et
al., 1993; Ro et al., 1993). There has been only one report of a
p53 mutation in melanoma, a C -> T transition in codon 248 of the
gene in the melanoma cell line SK-MEL-13 (Volkenandt et al.,
1991). The comparative lack of evidence of expression of p53 protein
in melanocytic naevi (Yu et al., 1992; Lassam et al., 1993)
would suggest that if p53 gene mutation is important in the genesis
of melanoma it is not the earliest mutation. The protein product of
p53 was detected in 12 of 18 (67%) choroidal melanomas (Tobal et
al., 1992) but none of 7 choroidal naevi. The DNA of exons 5, 7
and 8 of the p53 was sequenced from two of the strongly positive
choroidal melanomas: a mutation was found in each, one G -> T and
one C -> G.
Mutations in the ras oncogenes appear to be less frequent than
in the p53 gene in nonmelanocytic skin cancers. Mutations were
sought in one or more codons of Ha-ras, Ki-ras and N-ras in 77 BCC
and found in five (6%) (van der Schroeff et al., 1990; Lieu et
al., 1991; Campbell et al., 1993). There were two mutations, one
in a dipyrimidine site, in codon 61 of Ha-ras and three mutations at
dipyrimidine sites in codon 12 of Ki-ras; two mutations were C ->
A, one was C -> T and one was A -> T. These mutations were also
sought in 79 SCC and found in 7 (9%) (van der Schroeff et al.,
1990; Corominas et al., 1991; Lieu et al., 1991; Campbell et
al., 1993). There were three mutations in dipyrimidine sites in
codon 12 of Ha-ras, three, but none in dipyrimidine sites, in codon
61 of Ha-ras and one at a dipyrimidine site in codon 12 of Ki-ras;
three mutations were C -> A, one was C -> T and three were T ->
A. Overall, 8 of 12 (67%) mutations were at dipyrimidine sites.
There is nothing about the pattern of base changes that would favour
UV as the cause of the mutations. The comparative rarity of ras gene
mutations in nonmelanocytic skin cancers is supported by a study of
26 such tumours in Japanese patients with xeroderma pigmentosum
(Ishizaki et al., 1992). Only one mutation was found, an A -> T
change in codon 61 of Ki-ras.
Mutations in ras oncogenes have been demonstrated in some human
cutaneous melanomas. In three series totalling 59 primary melanomas
(van't Veer et al., 1989; Shukla et al., 1989; Albino et al.,
1991), ras gene mutations were found in 11 (19%). The mutations
occurred in codon 61 of N-ras (6), codon 13 of N-ras (4), codon 12
of Ki-ras (3) and codon 12 of Ha-ras (1) (two mutations were found
in each of 3 melanomas). Ten of the 13 mutations (77%) were at
dipyrimidine sites and nine were C -> A, three C -> T, one T -> C
and one T to A or G (IARC, 1992). The pattern of base changes is not
such as to suggest that UV was responsible for the mutations.
Mutations were sought but not found in codons 12, 13 and 61 of
Ha-ras, Ki-ras and N-ras in up to 68 uveal melanomas (Mooy et al.,
1991; Soparker et al., 1993).
Table 8.3 Summary of results of detection of mutations in the p53d
gene of BCC and SCC (Brash et al, 1991; Pierceall et al 1991; Rady
et al., 1992; Molès et al., 1993; Ziegler et al., 1993).
Mutations Detected BCC SCC
(60)a (46)
Any mutation 52% 39%
Base substitutions or 1 or 2 base deletions 45% 35%
at a dipyrimidine site
Types of base substitutions or 1 or 2 base 52%c 38%
deletions at dipyrimidine sites b
C -> T 13% 19%
CC -> TT 13% 31%
C -> A 10% 6%
C -> G 3% 0%
Table 8.3 (contd.)
Mutations Detected BCC SCC
T -> C 3% 0%
T -> A 3% 0%
G -> C 0% 6%
C deleted 3% 0%
CC deleted
a Total numbers of cancers examined.
b In consistency with the format adopted by Brash et al (1991) and
Ziegler et al (1993), four base substitutions reported as G -> A
(Pierceall et al, 1991; Rady et al, 1992) were represented as
C -> T and two reported as G -> T (Pierceall et al, 1991; Molès
et al, 1992) were represented as C -> A.
c Proportions of all point mutations at dipyrimidine sites.
Oxidative processes
Oxidative effects are an important consequence of exposure to
UV, especially UVA (see Chapter 6) and if they are involved in
mediating its carcinogenic effects in humans it might be expected
that anti-oxidant vitamins would reduce the risk of UV-related
cancers. There is some evidence of such effects, mainly for
cutaneous melanoma. Risk of melanoma was slightly but not
significantly reduced by high intakes of vitamin E and carotene and
high plasma concentrations of -carotene and -tocopherol in a
case-control study in the USA (Stryker et al., 1990). More
persuasively, a nested case-control study in a cohort study in
Finland showed significantly lower serum concentrations of -carotene
and -tocopherol in 10 cases of melanoma and 18 controls (Knekt et
al., 1991). Melanoma was the only cancer in this study to be
associated with low levels of anti-oxidant vitamins. A similar
result was obtained for -tocopherol in a case-control study of
melanoma carried out in Moscow (crude relative risk for highest
tertile of -tocopherol was 0.08, 95% CI 0.01-0.38; Zaridze et al.,
1992); risk was not reduced by high serum concentrations of
-carotene. As to nonmelanocytic skin cancer, in a hospital-based
case-control study covering 53 cases of BCC and 35 of SCC, high
intakes of vegetables were protective and cases had a significantly
lower mean level of -carotene than did controls (Kune et al.,
1992). It should be noted, however, that cases were on average, five
years older than controls. No evidence was found for a protective
effect of dietary carotenoids with vitamin A activity, vitamin C or
vitamin E in a study of diet and BCC in a cohort of US nurses
(Hunter et al., 1992).
Immune-suppression
UV has been shown to suppress immune functions in humans (see
Chapter 9). In addition there are a number of lines of evidence that
link immune suppression from other sources with skin cancer in
humans.
As early as 1980 it was clear that renal transplant patients,
who receive long-term immunosuppressive therapy, had an increased
incidence of nonmelanocytic skin cancer (Kinlen et al., 1979;
Hardie et al., 1980). Among 290 transplant patients in Queensland,
Australia, it was estimated that the incidence of nonmelanocytic
skin cancer was 21 times that in the general population Hardie et
al., 1980); the ratio of BCC to SCC was reversed from 4:1 in the
general population to 1:1.7 in the transplant patients suggesting
that the incidence of SCC was particularly increased in them.
Similar results were obtained in Canadian and Dutch series of
transplant patients (Gupta et al., 1986; Hartevelt et al., 1990)
In the Dutch study it was estimated that the incidence of SCC was
250 times higher than that in the general population and that of BCC
10 times higher.
There is evidence to suggest that the nonmelanocytic skin
cancers occurring in renal transplant patients are caused by sun
exposure. First their site distribution is similar to that of all
nonmelanocytic skin cancers and strongly favours sun exposed sites
(Hartevelt et al., 1990). Second, all 5 nonmelanocytic skin
cancers found by Boyle et al. (1984) in 94 transplant patients
occurred in the 17 patients judged to have high sun exposure. In a
more thorough study based on 137 Dutch transplant patients, 20 of
whom had SCC, 7 BCC and 9 both types of cancer, Bouwes-Bavinck
(1992) found a relative risk for 20 000 or more cumulated hours of
sun exposure compared with 10 000 or less of 97.5 (95% CI 6.6-1444)
for SCC and 49.3 (2.8-878) for BCC thus suggesting that sun exposure
had a strong effect on development of the transplant related skin
cancers.
The incidence of cutaneous melanoma has also been observed to
be increased in patients with renal transplants (relative risk 3.9,
95% CI 1.4-8.5; Hoover, 1977; Greene et al., 1981). All 14 renal
transplant patients with melanoma reported by Greene et al. (1981)
had fair complexions, light-coloured hair, light-coloured eyes and a
tendency to freckle thus suggesting that there may have been an
interaction between sun exposure and immune suppression in causing
their melanomas. The rapid appearance of new melanocytic naevi has
been reported in two renal transplant patients (Barker & MacDonald,
1988; McGregor et al., 1991). Melanoma incidence is also increased
in patients with lymphohaematopoetic neoplasms which are associated
with immune suppression (Greene & Wilson, 1985; Tucker et al.,
1985b; Travis et al., 1991, 1992) and has been reported to be
increased in patients with human immunodeficiency virus infection
(McGregor et al., 1992; Reynolds et al., 1993). Rapid appearance
of melanocytic naevi has also been reported in HIV infection (Duvic
et al., 1989).
8.6 Conclusions
UV exposure of the skin has both beneficial and harmful
effects. The beneficial effects include photochemical production of
vitamin D and widely believed but poorly documented effects on
general well-being. The harmful effects include sunburn,
phototoxicity, photoallergy, benign abnormalities of melanocytes
(freckles, melanocytic naevi and solar or senile lentigines) a range
of other chronic abnormalities resulting from UV injury to
keratinocytes, blood vessels and fibrous tissue, often described
together as "photoageing", skin cancer (melanoma and non-
melanocytic cancer) and possibly cancer of the lip.
The indirect epidemiological evidence that sun exposure causes
cutaneous melanoma and non-melanocytic cancer is strong. Their
incidence is less in darker-skinned ethnic groups than in those with
lighter skins residing in the same geographic area and, within
populations that are reasonably homogeneous ethnically, risk of skin
cancer increases with decreasing pigmentation of the skin and
reduced ability to produce a protective tan. Albinos, who lack
cutaneous pigmentation, appear to have an increased risk of
non-melanocytic skin cancer but not melanoma. The anatomic site
distibution of SCC favours the head and neck and upper limbs, sites
that are more or less continuously exposed to the sun when outdoors.
This concentration is less pronounced for BCC and melanoma, but both
are rare on sites that are rarely exposed to the sun.
Within countries covering an appreciable span of latitude, an
inverse relationship within latitude and incidence of both
non-melanocytic skin cancer and cutaneous melanoma has generally
been observed. The incidence of both cancers is substantially lower
in migrants from the United Kingdom (an area of low solar
irradiance) to Australia (an area of high solar irradiance) than it
is in persons of similar ethnic origin born in Australia. This
difference is not observed for BCC and melanoma in those who migrate
to Australia within the first ten years of life; this observation
may suggest that sun exposure in childhood is particularly important
in determining subsequent risk of skin cancer. Similar observations
have been made for melanoma in migrants to other countries with high
solar irradiance. Risk of melanoma has been shown to increase with
increasing lifetime average ambient solar irradiance at an
individual's places of residence.
The direct epidemiological evidence linking sun exposure and
skin cancer is weaker. Estimated total sun exposure of individuals
has not been consistently associated with risk of either
non-melanocytic skin cancer or melanoma. Indicators of benign sun
damage to the skin, however, are positively associated with risk of
both types of skin cancer. Occupational sun exposure has been found
to be weakly associated with non-melanocytic skin cancer in a few
studies but not consistently associated, either positively or
negatively, with melanoma. Non-occupational or recreational sun
exposure is consistently and quite strongly associated with risk of
melanoma whereas there are insufficient data to draw a conclusion
regarding non-occuptional exposure and non-melanocytic skin cancer.
The same is substantially true of history of sunburn although a few
observations suggest that it is associated with non-melanocytic skin
cancer.
Together, the indirect and direct evidence is sufficient to
conclude that sun exposure causes both melanoma and non- melanocytic
skin cancer.
Use of sunlamps or sunbeds has not been consistently associated
with risk of non-melanocytic skin cancer in several rather poorly
conducted studies. There is stronger evidence that it may be
associated with risk of melanoma in a number of better studies but
possible confounding of use of sunlamps and sunbeds with sun
exposure has not been consistently controlled.
Cancer of the lip is much more common in white than black
populations and is less frequent in migrants from areas of low
ambient solar irradiance to Australia and Israel than in those born
in these countries. It is associated with outdoor work but possible
confounding with tobacco and alcohol use has not been adequately
controlled in any study.
There are no data in humans from which the action spectrum for
production of either non-melanocytic skin cancer or cutaneous
melanoma can be inferred directly.
Problems in the quantitative measurement of the sun exposure of
individuals have prevented the determination, in humans, of the
relationship between individual dose of UV and risk of any form of
skin cancer. The observed geographical relationship between ambient
UV irradiance and incidence of skin cancers has been used to
estimate the quantitative relationship between ambient UV irradiance
and population risk of these cancers. Exponential or power
relationships have generally given an adequate fit to the data, but
provide rather variable estimates of the proportional increase in
incidence of skin cancer per unit proportional increase in
biologically effective UV irradiance. The accuracy of these
estimates is uncertain, particularly because of difficulties in the
accurate measurement of skin cancer incidence and difficulties in
control of likely confounding with cutaneous sensitivity to the sun
and sun-related behaviour.
A number of features of the epidemiology of melanoma suggest
that infrequent or intermittent exposure of skin that is unadapted
to sun exposure may be particularly important in its causation. This
hypothesis is supported by the relatively strong evidence relating
non-occupational (recreational) sun exposure and sunburn to
melanoma. There is little evidence that pattern of sun exposure is
important in the aetiology of non-melanocytic skin cancer but few
directly relevant observations exist.
The much increased rates of all skin cancers in patients with
xeroderma pigmentosum, who are deficient in the capacity to repair
UV-induced DNA damage, suggest that direct UV damage of DNA may be a
step in the causation of these cancers. This suggestion has been
supported by observation of UV specific mutations of the p53 tumour
suppressor gene in a proportion of patients with non-melanocytic
skin cancer. No such evidence is available for melanoma. Oxidative
and immune suppressant effects may also contribute to the capacity
of UV to cause skin cancers.
9. HUMAN STUDIES: IMMUNE FUNCTION
9.1 Immune Function Assays
Several investigators have examined the effect of UV on contact
hypersensitivity (CHS) responses to dinitrochlorobenzene (DNCB) or
other contact sensitizers in humans (Hersey et al., 1983a, 1983b;
O'Dell et al., 1980; Kalimo et al., 1983; Halprin et al.,
1981; Friedmann et al., 1989; Sjovall et al., 1985; Yoshikawa
et al., 1990; Vermeer et al., 1991; Cooper et al., 1992). Some
of these studies are inconclusive because they used patients with
skin diseases or recent UV-exposure, had insufficient subjects for
statistical analyses, or used subjective assessments of CHS. Hence
only 3 of these studies are discussed here.
Yoshikawa et al. (1990) exposed human buttock skin to 4 daily
UV doses of 1440 J m-2 using a high pressure mercury vapour lamp
(290-320 nm).Immediately after the last exposure, the irradiated
site was sensitized with 2000 µg DNCB. The inner surface of the
forearm was challenged 30 days later with 50 µg DNCB and CHS was
assessed. In this study 40-45% of healthy irradiated adults failed
to develop CHS. They designated these subjects UVB-S and suggested
that, as in mice, susceptibility to UV is genetically controlled.
Among biopsy proven skin cancer patients a much higher percentage
(92%) of UV-exposed individuals failed to developed CHS. The UVB-S
phenotype appeared to be a risk factor for the development of skin
cancer. Suppression of CHS was also demonstrated in 50% of
black-skinned individuals indicating that melanin does not protect
against the deleterious effects of UVB on the development of CHS
(Vermeer et al., 1991). Attempts to resensitize healthy UVB-S
individuals through normal skin following primary sensitization to
DNCB through irradiated skin were generally successful; however, 50%
of skin cancer patients failed to respond to resensitization
attempts suggesting that these patients developed immunological
tolerance similar to that demonstrated in experimental animal models
(Yoshikawa et al., 1990; Vermeer et al., 1991).
Cooper et al. (1992) exposed human buttock skin (using FS
lamps) to 0.75 or 2 MEDs of UVB (1 MED = 291 J m-2 to 325 J m-2
depending on the individual) for 4 days and sensitized with 30 µg
DNCB through irradiated skin immediately after the last exposure.
Subjects were challenged 3 weeks later with 4 serial 2-fold
dilutions of DNCB, the highest of which was 12.5 µg. In addition,
some subjects were exposed to 4 MED (moderate sunburn) and
sensitized 3 days later with DNCB. Analysis of overall individual
responses revealed decreased frequencies of fully successful
immunizations in all UVB exposed groups. Increasing doses of UVB
resulted in a linear decrease in immunological responsiveness to
DNCB. Only 5% of individuals exposed to 2 MED had strong positive
responses as opposed to 73% of unexposed individuals. The rate of
immunologic tolerance to DNCB (lasting up to 4 months) in the groups
that were initially sensitized on skin receiving erythemagenic doses
of UVB was 31% compared to 7% in controls. Similar results were
observed with UVA (320-340 nm) exposure (Cooper, 1993). The
differences in the Cooper and Yoshikawa studies may have to do with
different sensitization and challenge regimens, and differences in
methods used to quantitate the CHS response. Also, Cooper controlled
for diminished levels of CHS that occur in menstruating women except
during midcycle (Oberhelman et al., 1992) by sensitizing all
female subjects 14 days after the onset of menses.
Despite differences in the 2 studies it appears that
unresponsiveness following the application of a contact sensitizer
on UV-exposed skin can be induced in some parts of humans following
exposure to moderate levels of UV and that some individuals become
immunologically tolerant to the sensitizer in a manner reminiscent
of animal studies. Cooper et al. (1992) also demonstrated
modulation (although not clear-cut suppression) of contact
sensitivity to diphenylcyclopropenone (DPCP) in subjects exposed to
4 MED and sensitized 3 days later through unirradiated skin. A
similar effect was not observed with the 2 MED exposure regimen,
either because the dose was lower or the sensitization was
immediately following the last exposure. Hence the systemic effects
observed in mice may also occur in humans, although currently the
experimental data to support such effects are minimal.
As in the mouse, UV treatment of human skin resulted in altered
antigen presentation. UV caused depletion of Langerhans (CD1a+DR+)
cells followed by an influx of CD1a- DR+ macrophages that
preferentially activate CD4+ cells (suppressor-inducer) which, in
turn, induce maturation of CD-8+ suppressor T lymphocytes and
deregulate lymphocyte activation (Baadsgaard et al., 1990; Cooper
et al., 1986). Rasanen et al. (1989) reported a 70-80%
suppression in the ability of epidermal cells exposed to 2000 J
m-2 UVB in vivo to present PPD or HSV to lymphocytes in vitro.
In these studies recovery was observed 3 and 7 days post exposure.
The UV wavelengths responsible for induction of CD1a-DR+ cells were
found to lie predominantly within the UVB band and to a lesser
extent in the UVC band; UVA was a poor inducer of these
non-Langerhans cell antigen presenting cells (Baadsgaard et al.,
1987,1989). Hersey et al. (1983b) reported an increase in CD8+ T
cells and a decrease in CD4+ T cells in subjects exposed for 1
hour/day for 12 days to natural sunlight and a suppressor T cell
activity which lasted in many subjects for up to 2 weeks. Filtration
of solarium radiation through mylar prevented these changes
suggesting effects were due to UVB (Hersey et al., 1988). Hence in
humans as in mice, UVB appears to act by altering antigen
presentation in ways that favours suppressor cells.
Robinson & Rademaker (1992) studied 61 patients with two or
more BCC for occurrence of a further BCC. Patients were counselled
to avoid sun exposure, however there was a clear distinction between
those who reduced their sun exposure and those who did not. The
numbers of new BCC occurring after 36 months was determined by
regular examination of all patients. Plasma lymphocyte
subpopulations were measured at 0, 6, 12 and 18 months. At 36 months
an index of sun exposure was developed from questions on sun-related
behaviour before and after entry to the study. All 35 patients with
high sun exposure had low T helper to T suppressor cell (CD4/CD8)
ratios. The mean number of BCCs occurring during follow-up was 5.5
in those with high sun exposure and low CD4/CD8 ratios, 2.2 in those
with low sun exposure and low CD4/CD8 ratios and 1.1 in those with
low sun exposure and high CD4/CD8 ratios. The difference between the
first and the third groups was statistically significant. While
there is total confounding between high sun exposure and low CD4/CD8
ratios in this study it is consistent with the possibility that the
effect of sun exposure on risk of recurrent BCC was mediated by way
of its effect on cell-mediated immunity.
UV from solaria suppressed natural killer(NK) cell activity in
the blood of subjects exposed for 1 hr/day for 12 days and tested 1
and 7 days after exposure. NK activity returned to normal 21 days
post exposure. Filtration of UV through mylar did not affect this
response; hence effects on NK activity were attributed to UVA
(Hersey et al., 1983a; Hersey et al., 1988).
9.2 Susceptibility to Tumours, Infectious and Autoimmune
Diseases
As previously indicated Yoshikawa et al. (1990) found that
suppression of contact sensitivity following UV exposure was more
common among skin cancer patients than in healthy subjects. In their
study only skin cancer patients developed an immunological
tolerance. They suggested that individuals who were phenotypically
sensitive to the immunosuppressive effects of UV were at greater
risk of skin cancer.
Adverse effects of UV on 4 types of human infections have been
reported. Smallpox lesions were made larger by exposure to sunlight
(Finsen 1901). Lesions from Herpes Simplex Virus(HSV) types I and II
were reactivated by exposure to UV (Spruance, 1985; Klein, 1986).
Using the criteria established by Yoshikawa et al. (1990) for the
UVB-S phenotype, Taylor et al. (1993) reported that 66% of
individuals who had a strong history of HSV lip lesions provoked by
sun exposure were UVB-S as compared to 40-45% in the general
population and 92% in skin cancer patients (Yoshikawa et al.,
1990). Also, exposure of immunosuppressed patients to sunlight led
to an increased incidence of viral warts caused by papilloma virus,
presumably due to UVB exposure (Boyle et al., 1984; Dyall-Smith &
Varigos, 1985). Hence there is some indication that UV may
exacerbate certain infections in humans.
Recent attention has been paid to the effects of UV on immune
response in patients with human immune deficiency virus (HIV)
because UV has been shown to activate HIV in vitro (reviewed by
Zmudzka & Beer, 1990). It has been suggested that UV exposure could
progress these patients to full blown AIDS by interfering with
protective immunity, since Th1 responses appear to be protective
against AIDS whereas Th2 responses are not (Shearer & Clerici,
1992). However, Warfel et al. (1993) found no difference in CD4+
cell counts in HIV patients before and after treatment with 50-60%
of their MED for dermatological disease. More research in this area
is needed to resolve these issues.
Finally, it has been known for some time that UV exposure
adversely affects the clinical course of systemic lupus
erythematosus, an autoimmune disease (Epstein et al., 1965);
however, the relationship of UV in this immune response is unclear.
9.3 Conclusions
The above studies suggest that UV exposures at environmental
levels suppress immune responses in both rodents and man. In rodents
this immune suppression results in enhanced susceptibility to
certain infections with skin involvement as well as some systemic
infections. The mechanisms associated with UV-induced
immunosuppression in rodents and man are similar. Also, host defense
mechanisms which provide protection against infectious agents are
similar in rodents and man as shown in figure 9.1. It is therefore
reasonable to assume that exposure to UV may enhance the risk of
infection and decrease the effectiveness of vaccines in humans.
Additional research is needed to substantiate these
assumptions. In particular, experimental studies are needed in
rodents and man to access effects on immune function parameters in a
fashion that allows quantitative comparisons between the two
species. Results of experiments on rodents showing the effects of UV
exposure on the susceptibility to various infections could be used
to extrapolate the risk of infectious disease in humans. Ultimately
epidemiological studies on the effects of UV exposure on
susceptibility to infection and vaccine effectiveness are needed to
validate this hypothesis.
Table 8.2 summarises the most recent estimates of the BAFs for
nonmelanocytic skin cancer (BCC and SCC) and cutaneous melanoma from
data in the USA and Scandinavia. These estimates were based on
various action spectra: the response spectrum of the Robertson-
Berger meter (Scotto et al., 1983; Scotto & Fears, 1987; Pitcher &
Longtreth, 1991), the CIE reference spectrum of McKinlay and Diffey
(1987), Moan et al. (1989), Moan and Dahlbach, (1992), and the
Sterenberg-Slaper action spectrum for UV radiation of skin cancer in
albino hairless mice (de Gruijl and van der Leun, 1991). Generally,
the BAF for BCC lay between about 1.5 and 2.0. For SCC and melanoma,
there was much greater variation in estimates. Those for SCC were
between about 2.0 and 4.0 when based on the US data but between 1.0
and 2.0 when based on Norwegian data. Similarly, BAFs for melanoma
were between 0.3 and 0.5 when based on US data but between about 1.0
and 3.0 when based on Scandinavian data. While an attempt had been
made to adjust the US estimates for confounding with population
constitutional sensitivity to the sun and sun-related behaviour,
even the unadjusted estimates (ranging from 0.6 to 1.0; Scotto &
Fears, 1987; Pitcher & Longstreth, 1991) were considerably less than
those for Scandinavia. The most likely reasons for the differences
between the estimates is differences in their error.
Table 8.2 Recent estimates of the biological amplification factors (BAF) for
nonmelanocytic skin cancer and cutaneous melanoma based on geographical correlations
between average annual ambient UV and skin cancer incidence or mortality.
Authors Region Sex BAFs for BCC BAFs for SCC
Incidence Incidence
Nonmelanocytic skin cancer
Scotto et al, 1983 USA (8 centres) M 1.3-2.6a 2.1-4.1
F 1.1-2.1 2.2-4.3
de Gruijl & van der Leun 1991 USA (8 centres) MF 1.4 2.5
Moan et al 1989 Norway (6 areas) M 1.5-2.0a 1.2-1.5
Melanoma
Scotto & Fears, 1987 USA (7 areas) M 0.4c
F 0.5c
Pitcher & Longstreth, 1991 USA 215 SMAs M 0.4d
F 0.3d
Table 8.2 (contd).
Authors Region Sex BAFs for BCC BAFs for SCC
Incidence Incidence
Moan & Dahlback, 1992 Norway M 1.9
F 3.2
Finland M 1.3
F 2.2
Sweden M 1.9
F 2.3
a Exponential model used in which the value of the BAF varies with ambient UV, thus range
of values given.
b Same data as used by Scotto et al (1983) except that a power model was used instead of
an exponential model and the most recent action spectrum for UV carcinogenesis in mouse
skin was assumed.
c Adjusted for population estimates of ethnic origin, pigmentary characteristics, use of
sunscreens, and hours per week of outdoor exposure.
d Adjusted for population estimates of ethnic origin, household income, outdoor occupation
and education.
This approach to estimation of dose-response has assumed, among
other things, that: the correct action spectrum has been used to
weight spectral UV irradiance when producing a single figure for
ground level UV in each area; all members of the populations giving
rise to the incidence rates have lived their whole lives in their
present environment; the skin cancer incidence rates have been
measured accurately and, in particular, that their error does not
correlate with ambient UV radiance; and that possible confounding of
ambient UV with constitutional sensitivity to the sun and
sun-related behaviour is either unimportant or has been taken
adequately into account. None of these assumptions is likely to be
correct in any of the estimates of BAF made so far and the estimates
are all likely, therefore, to be inaccurate (Armstrong, 1993).
It should be noted that even if dose-response relationships
between UV and skin cancer incidence determined at the population
level are accurate, they may not reflect dose-response relationships
at the individual level. This is because of the averaging of
exposure and risk that occurs at the population level. The actual
population exposure of the skin at any level of ambient UV is an
average of many different exposures and the actual population
incidence of skin cancer is an average of many individual risks of
skin cancer. The association of individual risk to individual
exposure may be quite complex (as has been postulated for melanoma;
Armstrong, 1988) but at the population level this could still
translate into a simple exponential or power relationship between
ambient exposure and incidence.
No attempts have been made to estimate the dose-response
relationships for UV and cancers other than skin cancers that may be
caused by UV.
8.5.8 Effects of pattern of exposure
There are a number of aspects of the epidemiology of cutaneous
melanoma in European populations that appear inconsistent with a
simple relationship between it and sun exposure (English &
Armstrong, in press). First, in many populations, melanoma occurs as
commonly in women as in men, although men are more likely to work
outdoors; second, there is a relative peak in incidence in middle
life, which is not the pattern to be expected from life-long
exposure to an environmental agent; third, it most frequently occurs
on the back in men and lower limbs in women, sites which are not
maximally exposed to the sun; finally, melanoma is more common in
indoor than outdoor workers and in those of higher socioeconomic
status than those of lower status.
These observations led to postulation of the "intermittent
exposure hypothesis" for the relationship of sunlight to melanoma.
This hypothesis states that incidence of melanoma is determined as
much (or more) by the pattern of sun exposure as by the total
accumulated dose of sun exposure and, specifically, that infrequent
or intermittent exposure of untanned skin to intense sunlight is
particularly effective in increasing incidence of melanoma
(Armstrong, 1988). If, as seems plausible, this postulated effect of
intermittency of sun exposure is due to protective thickening and
pigmentation of the epidermis with more continuous sun exposure,
then intermittent exposure would be maximally effective if it
occurred at intervals longer than the time it takes for the skin to
return to its prior level of sensitivity after a single episode of
sun exposure (two to three weeks).
The epidemiological evidence, summarised above, that risk of
melanoma is apparently not increased with total or occupational
exposure to the sun but is increased with increasing
non-occupational exposure to the sun, in particular increasing time
spent in sun-related vacations in populations with otherwise
comparatively low sun exposure (Klepp & Magnus, 1979; Lew et al.,
1983; Elwood et al., 1985b; Osterlind et al., 1988b; Beitner
et al., 1990; Zanetti et al., 1992), and episodes of sunburn
(IARC, 1992) is consistent with this hypothesis.
There are less grounds for believing that pattern of UV is
important in causing nonmelanocytic skin cancer than there are for
melanoma. Specifically, nonmelanocytic skin cancer as a whole, and
BCC and SCC separately, are more common in men than women (Kricker
et al., 1990; Parkin et al., 1992; Marks et al., 1993) and, in
the largest, well-collected, population-based incidence series
available, they showed steadily increasing incidence rates with age,
except for a downturn in older age-groups possibly due to
underascertainment (Scotto et al., 1983). On the other hand,
recent series of BCC have shown unexpectedly high proportions on the
trunk (Kricker et al., 1990; Magnus, 1991) and, as reviewed above,
the relationship of nonmelanocytic skin cancer with occupational
exposure to the sun is not at all clear. Moreover, there is some
evidence of an association between sunburn and other indicators of
intermittent exposure and BCC (Hunter et al., 1990; Kricker et
al., 1991b).
8.5.9 Interactions between UV and other agents
5-methoxypsoralen and 8-methoxypsoralen (methoxsalen)
Methoxsalen, 5-methoxypsoralen and some other psoralens
(hereafter referred to collectively as psoralens) are used in
combination with UVA (referred to as PUVA) for the treatment of
psoriasis and to produce repigmentation in vitiligo. A working group
of International Agency for Research on Cancer concluded in 1987
that there was sufficient evidence that PUVA (defined as
8-methoxypsoralen plus long-wave UV radiation) causes cancer in
humans, based mainly on evidence of its association with SCC.
Recent studies have confirmed and extended this conclusion.
Stern & Lange (1988) reported on 5 to 10 year follow-up of patients
treated with PUVA in the USA. In 13 384 person years of follow-up of
1 380 people, 100 people had developed 391 SCC and 94 had developed
218 BCC. There were statistically significantly increasing risks of
both BCC and SCC occurring more than 58 months after entry into the
cohort with increasing number of treatments with PUVA. Risk of BCC
rose to 6.9 (95% CI 3.2-13.1) with 260 or more treatments and risk
of SCC rose to 50.1 (95% CI 24.9-89.5). Therapeutic exposure to
ionizing radiation, high doses of tar and UVB was unrelated to
number of treatments with PUVA and, therefore, did not confound the
observed dose-response relationship. Lindelöf et al. (1991)
reported on follow-up for an average of 6.9 years of 4 799 Swedish
patients treated with PUVA. Risk of SCC increased with increasing
number of treatments with PUVA. In those who received 200 or more
treatments, the relative risk, with reference to the general Swedish
population, was 30.7 (14.7-56.5) in men and 18.5 (0.5-103.2) in
women. BCC were not recorded in this study.
Cases of melanoma have been reported in patients treated with
PUVA but the rates of melanoma in US and Swedish PUVA cohorts were
little higher than those expected on the basis of population
incidence rates. In the US cohort the relative risk was 1.46 (95% CI
0.3-7.3; Gupta et al., 1988), while in the Swedish cohort it was
1.1 (0.2-3.2) in men and 0.8 (0.1-3.0) in women (Lindelöf et al.,
1991).
It is not possible in any of these studies to estimate what the
risk of skin cancer would have been if the patients had been treated
with UV alone. It would seem reasonable to assume, however, on the
basis of the evidence from experimental animals (see above) that
administration of psoralens potentiated the effect of UV. In
addition analysis of clinical data even suggests that this
carcinogenicity of PUVA exceeds that of UVB treatment by an order of
magnitude (Slaper, 1987).
Ionizing radiation
Ionizing radiation causes nonmelanocytic skin cancer, mainly
BCC, generally after a long latent period (Shore, 1990; Fragu et
al., 1991; Hogan et al., 1991; Sadamori et al., 1991; Moller
et al., 1993). Incidence in the atomic bomb survivors was related
strongly to exposure, although not for those closest to the blast
(Sadamori et al., 1991). There is evidence that ionizing radiation
and UV may act synergistically in causing skin cancer. In a study of
skin cancer following irradiation of the scalp for ringworm, Shore
et al. (1984) found a substantial excess of BCC in white subjects.
In blacks, no skin cancers were observed when 13 would have been
expected from the rate on the UV-shielded scalp in whites. This lack
of skin cancers in irradiated blacks suggest that UV radiation
contributes more than additively to the production of these cancers
in irradiated whites. This inference is supported by the observation
that the excess of BCC around the UV-exposed fringe of the scalp in
irradiated whites was some four to five times higher per unit dose
and area than that on the UV-shielded hairy scalp (Shore et al.,
1984). Further, Modan et al. (1993) observed, in a separate cohort
of people irradiated for ringworm of the scalp, that sunbathing
increased the risk of skin cancer 2.6 times (95% CI 1.1-6.1). All
except one of 39 cancers were BCCs and 60% were on the scalp, thus
suggesting that the ionizing radiation had also contributed to their
causation. However, no formal analysis of interaction was reported
from this study.
Other agents
One or more SCCs occur in 30% to 50% of patients with
epidermodysplasia verruciformis, a rare chronic skin disease in
which there are multiple benign skin lesions caused by a variety of
different types of human papilloma viruses (HPV; Quan & Moy, 1991).
HPV DNA sequences have been identified in the SCC occurring in these
patients and most of the cancers occur on sun-exposed sites. This
coincidence suggests that the UV interacts with the virus in causing
these cancers. Benign warts are also increased in frequency in renal
transplant patients and it is possible that they, with UV, are
involved in causing the excess of nonmelanocytic skin cancers that
occur in these patients (Bouwes-Bavinck, 1992). These cancers, too,
occur mainly on sun-exposed sites.
8.5.10 Mechanisms of UV carcinogenesis
Precursor lesions
Solar keratoses are very strongly related to risk of
nonmelanocytic skin cancer (Kricker et al., 1991a) and it is a
matter of clinical observation that SCC often occurs in lesions
which, previously, had the clinical appearances of solar keratoses.
There is evidence that solar keratoses are, themselves, caused by
exposure to the sun. Thus solar keratoses could be the visible
expression of an early, UV-caused, mutational step in the pathway to
skin carcinogenesis.
Analogously with solar keratoses, benign melanocytic naevi are
commonly observed in clinical or histopathological contiguity with
melanomas and have been found, epidemiologically to be strongly
related to risk of cutaneous melanoma (Holman & Armstrong, 1984a;
Armstrong & English, 1988). Naevi may, therefore, represent an
early, UV-caused, mutational step in the development of melanoma. In
this regard, it is interesting to note that the most active
development of naevi occurs during the first 15 years of life, thus
providing a possible link to early life sun exposure as suggested by
migrant data (see above). The appearance of naevi in children who
have received chemotherapy for cancer may indicate that they can be
caused by mutagens other then UV (Hughes et al., 1989; Baird et
al., 1992; Green et al., 1992).
DNA damage
The evidence of much increased incidence rates of BCC, SCC
cutaneous melanoma and tumours of the anterior eye in patients with
xeroderma pigmentosum (XP) (see above) strongly suggests that
unrepaired, UV-induced photoproducts in DNA form part of the genesis
of these cancers. Almost all forms of XP have an inherited
deficiency in excision repair of DNA photoproducts (Kraemer et al.,
1987). The position is made somewhat less clear, however, by the
existence of two other inherited syndromes involving deficiency in
DNA excision repair, trichothiodystrophy and Cockaynes syndrome,
that are not associated with increased incidence rates of skin
cancer (Bridges, 1990; Barrett et al., 1991).
There is evidence also that low levels of DNA repair may be
associated with skin cancer in the general population, although
there is some disagreement in the results so far obtained .
Munch-Petersen et al. (1985) studied 29 patients with multiple
skin neoplasms, 19 with multiple BCC only and 10 with both BCC and
squamous lesions (SCC, solar keratosis, or Bowen's disease), and 25
control subjects with roughly the same age and sex distributions.
Patients with both BCC and squamous lesions had much higher
UV-induced DNA synthesis in lymphocytes, as measured by
incorporation of [3H]thymidine following irradiation with a Philips,
TUV, 6 watt lamp (peak emission at 254 nm), than did controls.
Patients with multiple BCC were not different from controls. The
authors attributed this difference to possible defects in the DNA
ligation process. Roth et al. (1987) studied the rate of loss of
dithymidine dimers in DNA of cultured fibroblasts that had been
irradiated with a Philips 6-V germicidal lamp with peak emission at
254 nm. Dimers were measured with a monoclonal antibody highly
specific for the conformational change in DNA caused by the dimers.
No significant difference was seen between 16 BCC cases and 30
controls in the rate of loss of dimers. In ten melanoma patients,
however, the mean percentage of bound antibody at 60 minutes was
50.5 (SD 18.2) compared with 29.8 (SD 5.7) in controls (p=0.001)
thus suggesting that average dimer repair capacity was less in the
melanoma patients than the controls. Alcalay et al. (1990)
compared 22 patients with a history of one (13 patients) or more BCC
in the past with 19 healthy volunteers. Each subject was irradiated
at two locations on the lower back with 1 MED of UV from a 150 W
xenon arc solar UV simulator from which wavelengths < 295 nm had
been removed by a 1 mm WG 320 filter. Shave biopsies of skin were
taken from one of the sites immediately and the other 6 hours after
irradiation and the concentrations of pyrimidine dimers in DNA
measured by the micrococcus luteus endonuclease assay. Dimer yields
immediately after irradiation were similar in BCC patients and
controls but, after 6 hours, the proportion of dimers that had been
repaired was 22% in the BCC patients compared with 33% in the
controls (p=0.06). Wei et al. (1993) measured DNA repair capacity
in 88 patients with a history of one or more histologically
confirmed BCCs and 135 control subjects. Repair capacity was
measured by assaying the capacity of lymphocytes to repair
UV-induced damage in DNA of a nonreplicating recombinant plasmid.
Damage was induced by irradiating the plasmid with either 3500 J
m-2 or 7000 J m-2 of UV at 254 nm. The BCC cases had a
significantly lower mean DNA repair capacity (p=0.047) than controls
only when 29 controls with a family history of BCC or who themselves
had an actinic keratosis were removed from the comparison.
It now seems highly likely that UV can mutate the p53 tumour
suppressor gene in human skin and that UV-induced mutations in this
gene may be involved in the aetiology of some human cancers. First,
a recent study has shown that mutations that are strongly suggestive
of a UV effect, CC-TT changes at dipyrimidine sites, could be found
in 17 of 23 (74%) samples of sun-exposed normal skin in Australian
skin cancer patients compared with 1 of 20 (5%) samples of skin from
sites not exposed to the sun (Nakazawa et al., in press). Focal
overexpression of p53 protein has also been observed in normal, sun
exposed skin adjacent to BCC in 37 of 39 subjects but only in one
keratinocyte in 14 samples of buttock skin from the same subjects
(Shea et al., 1992). Second, several studies have found mutations
at dipyrimidine sites in the p53 gene in 36% to 56% of BCC (Rady et
al., 1992; Molès et al., 1993; Ziegler et al., 1993). In three
similar studies of SCC one found these mutations in 14 (57%) of 24
cases (Brash et al., 1991) one in two of 10 cases (Pierceall et
al., 1991) and, the other, none of 13 (Molès et al., 1993). The
more detailed findings of these five studies are summarised in table
8.3. Mutations were found in the p53 gene, following examinations of
varying extent, in 46% of BCC and SCC, the majority of which (88%)
were base substitutions or one or two base deletions at dipyrimidine
sites (the proportion expected by chance is 75%). The highest
proportions of these mutations resulted from C -> T, CC -> TT and
C -> A base changes which is consistent with what would be expected
if they had been caused by UV (Brash et al., 1991). Several
studies have found p53 protein by immunostaining (evidence of
overexpression of possibly abnormal p53 protein) in from 42% to 83%
of BCC and 56% to 65% of SCC (Barbareschi et al., 1992; Shea et
al., 1992; Stephenson et al., 1992, Ro et al., 1993) and are
thus reasonably consistent with the observations as to prevalence of
mutations.
The position with respect to p53 mutation is much less clear
for other cancers possibly caused by UV. A number of studies have
reported on the prevalence of detectable p53 protein in primary
melanomas: the proportions have varied from 3.6% to 97% (see, for
example, Stretch et al., 1991; Akslen & Morkve, 1992; Lassam et
al., 1993; Ro et al., 1993). There has been only one report of a
p53 mutation in melanoma, a C -> T transition in codon 248 of the
gene in the melanoma cell line SK-MEL-13 (Volkenandt et al.,
1991). The comparative lack of evidence of expression of p53 protein
in melanocytic naevi (Yu et al., 1992; Lassam et al., 1993)
would suggest that if p53 gene mutation is important in the genesis
of melanoma it is not the earliest mutation. The protein product of
p53 was detected in 12 of 18 (67%) choroidal melanomas (Tobal et
al., 1992) but none of 7 choroidal naevi. The DNA of exons 5, 7
and 8 of the p53 was sequenced from two of the strongly positive
choroidal melanomas: a mutation was found in each, one G -> T and
one C -> G.
Mutations in the ras oncogenes appear to be less frequent than
in the p53 gene in nonmelanocytic skin cancers. Mutations were
sought in one or more codons of Ha-ras, Ki-ras and N-ras in 77 BCC
and found in five (6%) (van der Schroeff et al., 1990; Lieu et
al., 1991; Campbell et al., 1993). There were two mutations, one
in a dipyrimidine site, in codon 61 of Ha-ras and three mutations at
dipyrimidine sites in codon 12 of Ki-ras; two mutations were C ->
A, one was C -> T and one was A -> T. These mutations were also
sought in 79 SCC and found in 7 (9%) (van der Schroeff et al.,
1990; Corominas et al., 1991; Lieu et al., 1991; Campbell et
al., 1993). There were three mutations in dipyrimidine sites in
codon 12 of Ha-ras, three, but none in dipyrimidine sites, in codon
61 of Ha-ras and one at a dipyrimidine site in codon 12 of Ki-ras;
three mutations were C -> A, one was C -> T and three were T ->
A. Overall, 8 of 12 (67%) mutations were at dipyrimidine sites.
There is nothing about the pattern of base changes that would favour
UV as the cause of the mutations. The comparative rarity of ras gene
mutations in nonmelanocytic skin cancers is supported by a study of
26 such tumours in Japanese patients with xeroderma pigmentosum
(Ishizaki et al., 1992). Only one mutation was found, an A -> T
change in codon 61 of Ki-ras.
Mutations in ras oncogenes have been demonstrated in some human
cutaneous melanomas. In three series totalling 59 primary melanomas
(van't Veer et al., 1989; Shukla et al., 1989; Albino et al.,
1991), ras gene mutations were found in 11 (19%). The mutations
occurred in codon 61 of N-ras (6), codon 13 of N-ras (4), codon 12
of Ki-ras (3) and codon 12 of Ha-ras (1) (two mutations were found
in each of 3 melanomas). Ten of the 13 mutations (77%) were at
dipyrimidine sites and nine were C -> A, three C -> T, one T -> C
and one T to A or G (IARC, 1992). The pattern of base changes is not
such as to suggest that UV was responsible for the mutations.
Mutations were sought but not found in codons 12, 13 and 61 of
Ha-ras, Ki-ras and N-ras in up to 68 uveal melanomas (Mooy et al.,
1991; Soparker et al., 1993).
Table 8.3 Summary of results of detection of mutations in the p53d
gene of BCC and SCC (Brash et al, 1991; Pierceall et al 1991; Rady
et al., 1992; Molès et al., 1993; Ziegler et al., 1993).
Mutations Detected BCC SCC
(60)a (46)
Any mutation 52% 39%
Base substitutions or 1 or 2 base deletions 45% 35%
at a dipyrimidine site
Types of base substitutions or 1 or 2 base 52%c 38%
deletions at dipyrimidine sites b
C -> T 13% 19%
CC -> TT 13% 31%
C -> A 10% 6%
C -> G 3% 0%
Table 8.3 (contd.)
Mutations Detected BCC SCC
T -> C 3% 0%
T -> A 3% 0%
G -> C 0% 6%
C deleted 3% 0%
CC deleted
a Total numbers of cancers examined.
b In consistency with the format adopted by Brash et al (1991) and
Ziegler et al (1993), four base substitutions reported as G -> A
(Pierceall et al, 1991; Rady et al, 1992) were represented as
C -> T and two reported as G -> T (Pierceall et al, 1991; Molès
et al, 1992) were represented as C -> A.
c Proportions of all point mutations at dipyrimidine sites.
Oxidative processes
Oxidative effects are an important consequence of exposure to
UV, especially UVA (see Chapter 6) and if they are involved in
mediating its carcinogenic effects in humans it might be expected
that anti-oxidant vitamins would reduce the risk of UV-related
cancers. There is some evidence of such effects, mainly for
cutaneous melanoma. Risk of melanoma was slightly but not
significantly reduced by high intakes of vitamin E and carotene and
high plasma concentrations of -carotene and -tocopherol in a
case-control study in the USA (Stryker et al., 1990). More
persuasively, a nested case-control study in a cohort study in
Finland showed significantly lower serum concentrations of -carotene
and -tocopherol in 10 cases of melanoma and 18 controls (Knekt et
al., 1991). Melanoma was the only cancer in this study to be
associated with low levels of anti-oxidant vitamins. A similar
result was obtained for -tocopherol in a case-control study of
melanoma carried out in Moscow (crude relative risk for highest
tertile of -tocopherol was 0.08, 95% CI 0.01-0.38; Zaridze et al.,
1992); risk was not reduced by high serum concentrations of
-carotene. As to nonmelanocytic skin cancer, in a hospital-based
case-control study covering 53 cases of BCC and 35 of SCC, high
intakes of vegetables were protective and cases had a significantly
lower mean level of -carotene than did controls (Kune et al.,
1992). It should be noted, however, that cases were on average, five
years older than controls. No evidence was found for a protective
effect of dietary carotenoids with vitamin A activity, vitamin C or
vitamin E in a study of diet and BCC in a cohort of US nurses
(Hunter et al., 1992).
Immune-suppression
UV has been shown to suppress immune functions in humans (see
Chapter 9). In addition there are a number of lines of evidence that
link immune suppression from other sources with skin cancer in
humans.
As early as 1980 it was clear that renal transplant patients,
who receive long-term immunosuppressive therapy, had an increased
incidence of nonmelanocytic skin cancer (Kinlen et al., 1979;
Hardie et al., 1980). Among 290 transplant patients in Queensland,
Australia, it was estimated that the incidence of nonmelanocytic
skin cancer was 21 times that in the general population Hardie et
al., 1980); the ratio of BCC to SCC was reversed from 4:1 in the
general population to 1:1.7 in the transplant patients suggesting
that the incidence of SCC was particularly increased in them.
Similar results were obtained in Canadian and Dutch series of
transplant patients (Gupta et al., 1986; Hartevelt et al., 1990)
In the Dutch study it was estimated that the incidence of SCC was
250 times higher than that in the general population and that of BCC
10 times higher.
There is evidence to suggest that the nonmelanocytic skin
cancers occurring in renal transplant patients are caused by sun
exposure. First their site distribution is similar to that of all
nonmelanocytic skin cancers and strongly favours sun exposed sites
(Hartevelt et al., 1990). Second, all 5 nonmelanocytic skin
cancers found by Boyle et al. (1984) in 94 transplant patients
occurred in the 17 patients judged to have high sun exposure. In a
more thorough study based on 137 Dutch transplant patients, 20 of
whom had SCC, 7 BCC and 9 both types of cancer, Bouwes-Bavinck
(1992) found a relative risk for 20 000 or more cumulated hours of
sun exposure compared with 10 000 or less of 97.5 (95% CI 6.6-1444)
for SCC and 49.3 (2.8-878) for BCC thus suggesting that sun exposure
had a strong effect on development of the transplant related skin
cancers.
The incidence of cutaneous melanoma has also been observed to
be increased in patients with renal transplants (relative risk 3.9,
95% CI 1.4-8.5; Hoover, 1977; Greene et al., 1981). All 14 renal
transplant patients with melanoma reported by Greene et al. (1981)
had fair complexions, light-coloured hair, light-coloured eyes and a
tendency to freckle thus suggesting that there may have been an
interaction between sun exposure and immune suppression in causing
their melanomas. The rapid appearance of new melanocytic naevi has
been reported in two renal transplant patients (Barker & MacDonald,
1988; McGregor et al., 1991). Melanoma incidence is also increased
in patients with lymphohaematopoetic neoplasms which are associated
with immune suppression (Greene & Wilson, 1985; Tucker et al.,
1985b; Travis et al., 1991, 1992) and has been reported to be
increased in patients with human immunodeficiency virus infection
(McGregor et al., 1992; Reynolds et al., 1993). Rapid appearance
of melanocytic naevi has also been reported in HIV infection (Duvic
et al., 1989).
8.6 Conclusions
UV exposure of the skin has both beneficial and harmful
effects. The beneficial effects include photochemical production of
vitamin D and widely believed but poorly documented effects on
general well-being. The harmful effects include sunburn,
phototoxicity, photoallergy, benign abnormalities of melanocytes
(freckles, melanocytic naevi and solar or senile lentigines) a range
of other chronic abnormalities resulting from UV injury to
keratinocytes, blood vessels and fibrous tissue, often described
together as "photoageing", skin cancer (melanoma and non-
melanocytic cancer) and possibly cancer of the lip.
The indirect epidemiological evidence that sun exposure causes
cutaneous melanoma and non-melanocytic cancer is strong. Their
incidence is less in darker-skinned ethnic groups than in those with
lighter skins residing in the same geographic area and, within
populations that are reasonably homogeneous ethnically, risk of skin
cancer increases with decreasing pigmentation of the skin and
reduced ability to produce a protective tan. Albinos, who lack
cutaneous pigmentation, appear to have an increased risk of
non-melanocytic skin cancer but not melanoma. The anatomic site
distibution of SCC favours the head and neck and upper limbs, sites
that are more or less continuously exposed to the sun when outdoors.
This concentration is less pronounced for BCC and melanoma, but both
are rare on sites that are rarely exposed to the sun.
Within countries covering an appreciable span of latitude, an
inverse relationship within latitude and incidence of both
non-melanocytic skin cancer and cutaneous melanoma has generally
been observed. The incidence of both cancers is substantially lower
in migrants from the United Kingdom (an area of low solar
irradiance) to Australia (an area of high solar irradiance) than it
is in persons of similar ethnic origin born in Australia. This
difference is not observed for BCC and melanoma in those who migrate
to Australia within the first ten years of life; this observation
may suggest that sun exposure in childhood is particularly important
in determining subsequent risk of skin cancer. Similar observations
have been made for melanoma in migrants to other countries with high
solar irradiance. Risk of melanoma has been shown to increase with
increasing lifetime average ambient solar irradiance at an
individual's places of residence.
The direct epidemiological evidence linking sun exposure and
skin cancer is weaker. Estimated total sun exposure of individuals
has not been consistently associated with risk of either
non-melanocytic skin cancer or melanoma. Indicators of benign sun
damage to the skin, however, are positively associated with risk of
both types of skin cancer. Occupational sun exposure has been found
to be weakly associated with non-melanocytic skin cancer in a few
studies but not consistently associated, either positively or
negatively, with melanoma. Non-occupational or recreational sun
exposure is consistently and quite strongly associated with risk of
melanoma whereas there are insufficient data to draw a conclusion
regarding non-occuptional exposure and non-melanocytic skin cancer.
The same is substantially true of history of sunburn although a few
observations suggest that it is associated with non-melanocytic skin
cancer.
Together, the indirect and direct evidence is sufficient to
conclude that sun exposure causes both melanoma and non- melanocytic
skin cancer.
Use of sunlamps or sunbeds has not been consistently associated
with risk of non-melanocytic skin cancer in several rather poorly
conducted studies. There is stronger evidence that it may be
associated with risk of melanoma in a number of better studies but
possible confounding of use of sunlamps and sunbeds with sun
exposure has not been consistently controlled.
Cancer of the lip is much more common in white than black
populations and is less frequent in migrants from areas of low
ambient solar irradiance to Australia and Israel than in those born
in these countries. It is associated with outdoor work but possible
confounding with tobacco and alcohol use has not been adequately
controlled in any study.
There are no data in humans from which the action spectrum for
production of either non-melanocytic skin cancer or cutaneous
melanoma can be inferred directly.
Problems in the quantitative measurement of the sun exposure of
individuals have prevented the determination, in humans, of the
relationship between individual dose of UV and risk of any form of
skin cancer. The observed geographical relationship between ambient
UV irradiance and incidence of skin cancers has been used to
estimate the quantitative relationship between ambient UV irradiance
and population risk of these cancers. Exponential or power
relationships have generally given an adequate fit to the data, but
provide rather variable estimates of the proportional increase in
incidence of skin cancer per unit proportional increase in
biologically effective UV irradiance. The accuracy of these
estimates is uncertain, particularly because of difficulties in the
accurate measurement of skin cancer incidence and difficulties in
control of likely confounding with cutaneous sensitivity to the sun
and sun-related behaviour.
A number of features of the epidemiology of melanoma suggest
that infrequent or intermittent exposure of skin that is unadapted
to sun exposure may be particularly important in its causation. This
hypothesis is supported by the relatively strong evidence relating
non-occupational (recreational) sun exposure and sunburn to
melanoma. There is little evidence that pattern of sun exposure is
important in the aetiology of non-melanocytic skin cancer but few
directly relevant observations exist.
The much increased rates of all skin cancers in patients with
xeroderma pigmentosum, who are deficient in the capacity to repair
UV-induced DNA damage, suggest that direct UV damage of DNA may be a
step in the causation of these cancers. This suggestion has been
supported by observation of UV specific mutations of the p53 tumour
suppressor gene in a proportion of patients with non-melanocytic
skin cancer. No such evidence is available for melanoma. Oxidative
and immune suppressant effects may also contribute to the capacity
of UV to cause skin cancers.
9. HUMAN STUDIES: IMMUNE FUNCTION
9.1 Immune Function Assays
Several investigators have examined the effect of UV on contact
hypersensitivity (CHS) responses to dinitrochlorobenzene (DNCB) or
other contact sensitizers in humans (Hersey et al., 1983a, 1983b;
O'Dell et al., 1980; Kalimo et al., 1983; Halprin et al.,
1981; Friedmann et al., 1989; Sjovall et al., 1985; Yoshikawa
et al., 1990; Vermeer et al., 1991; Cooper et al., 1992). Some
of these studies are inconclusive because they used patients with
skin diseases or recent UV-exposure, had insufficient subjects for
statistical analyses, or used subjective assessments of CHS. Hence
only 3 of these studies are discussed here.
Yoshikawa et al. (1990) exposed human buttock skin to 4 daily
UV doses of 1440 J m-2 using a high pressure mercury vapour lamp
(290-320 nm).Immediately after the last exposure, the irradiated
site was sensitized with 2000 µg DNCB. The inner surface of the
forearm was challenged 30 days later with 50 µg DNCB and CHS was
assessed. In this study 40-45% of healthy irradiated adults failed
to develop CHS. They designated these subjects UVB-S and suggested
that, as in mice, susceptibility to UV is genetically controlled.
Among biopsy proven skin cancer patients a much higher percentage
(92%) of UV-exposed individuals failed to developed CHS. The UVB-S
phenotype appeared to be a risk factor for the development of skin
cancer. Suppression of CHS was also demonstrated in 50% of
black-skinned individuals indicating that melanin does not protect
against the deleterious effects of UVB on the development of CHS
(Vermeer et al., 1991). Attempts to resensitize healthy UVB-S
individuals through normal skin following primary sensitization to
DNCB through irradiated skin were generally successful; however, 50%
of skin cancer patients failed to respond to resensitization
attempts suggesting that these patients developed immunological
tolerance similar to that demonstrated in experimental animal models
(Yoshikawa et al., 1990; Vermeer et al., 1991).
Cooper et al. (1992) exposed human buttock skin (using FS
lamps) to 0.75 or 2 MEDs of UVB (1 MED = 291 J m-2 to 325 J m-2
depending on the individual) for 4 days and sensitized with 30 µg
DNCB through irradiated skin immediately after the last exposure.
Subjects were challenged 3 weeks later with 4 serial 2-fold
dilutions of DNCB, the highest of which was 12.5 µg. In addition,
some subjects were exposed to 4 MED (moderate sunburn) and
sensitized 3 days later with DNCB. Analysis of overall individual
responses revealed decreased frequencies of fully successful
immunizations in all UVB exposed groups. Increasing doses of UVB
resulted in a linear decrease in immunological responsiveness to
DNCB. Only 5% of individuals exposed to 2 MED had strong positive
responses as opposed to 73% of unexposed individuals. The rate of
immunologic tolerance to DNCB (lasting up to 4 months) in the groups
that were initially sensitized on skin receiving erythemagenic doses
of UVB was 31% compared to 7% in controls. Similar results were
observed with UVA (320-340 nm) exposure (Cooper, 1993). The
differences in the Cooper and Yoshikawa studies may have to do with
different sensitization and challenge regimens, and differences in
methods used to quantitate the CHS response. Also, Cooper controlled
for diminished levels of CHS that occur in menstruating women except
during midcycle (Oberhelman et al., 1992) by sensitizing all
female subjects 14 days after the onset of menses.
Despite differences in the 2 studies it appears that
unresponsiveness following the application of a contact sensitizer
on UV-exposed skin can be induced in some parts of humans following
exposure to moderate levels of UV and that some individuals become
immunologically tolerant to the sensitizer in a manner reminiscent
of animal studies. Cooper et al. (1992) also demonstrated
modulation (although not clear-cut suppression) of contact
sensitivity to diphenylcyclopropenone (DPCP) in subjects exposed to
4 MED and sensitized 3 days later through unirradiated skin. A
similar effect was not observed with the 2 MED exposure regimen,
either because the dose was lower or the sensitization was
immediately following the last exposure. Hence the systemic effects
observed in mice may also occur in humans, although currently the
experimental data to support such effects are minimal.
As in the mouse, UV treatment of human skin resulted in altered
antigen presentation. UV caused depletion of Langerhans (CD1a+DR+)
cells followed by an influx of CD1a- DR+ macrophages that
preferentially activate CD4+ cells (suppressor-inducer) which, in
turn, induce maturation of CD-8+ suppressor T lymphocytes and
deregulate lymphocyte activation (Baadsgaard et al., 1990; Cooper
et al., 1986). Rasanen et al. (1989) reported a 70-80%
suppression in the ability of epidermal cells exposed to 2000 J
m-2 UVB in vivo to present PPD or HSV to lymphocytes in vitro.
In these studies recovery was observed 3 and 7 days post exposure.
The UV wavelengths responsible for induction of CD1a-DR+ cells were
found to lie predominantly within the UVB band and to a lesser
extent in the UVC band; UVA was a poor inducer of these
non-Langerhans cell antigen presenting cells (Baadsgaard et al.,
1987,1989). Hersey et al. (1983b) reported an increase in CD8+ T
cells and a decrease in CD4+ T cells in subjects exposed for 1
hour/day for 12 days to natural sunlight and a suppressor T cell
activity which lasted in many subjects for up to 2 weeks. Filtration
of solarium radiation through mylar prevented these changes
suggesting effects were due to UVB (Hersey et al., 1988). Hence in
humans as in mice, UVB appears to act by altering antigen
presentation in ways that favours suppressor cells.
Robinson & Rademaker (1992) studied 61 patients with two or
more BCC for occurrence of a further BCC. Patients were counselled
to avoid sun exposure, however there was a clear distinction between
those who reduced their sun exposure and those who did not. The
numbers of new BCC occurring after 36 months was determined by
regular examination of all patients. Plasma lymphocyte
subpopulations were measured at 0, 6, 12 and 18 months. At 36 months
an index of sun exposure was developed from questions on sun-related
behaviour before and after entry to the study. All 35 patients with
high sun exposure had low T helper to T suppressor cell (CD4/CD8)
ratios. The mean number of BCCs occurring during follow-up was 5.5
in those with high sun exposure and low CD4/CD8 ratios, 2.2 in those
with low sun exposure and low CD4/CD8 ratios and 1.1 in those with
low sun exposure and high CD4/CD8 ratios. The difference between the
first and the third groups was statistically significant. While
there is total confounding between high sun exposure and low CD4/CD8
ratios in this study it is consistent with the possibility that the
effect of sun exposure on risk of recurrent BCC was mediated by way
of its effect on cell-mediated immunity.
UV from solaria suppressed natural killer(NK) cell activity in
the blood of subjects exposed for 1 hr/day for 12 days and tested 1
and 7 days after exposure. NK activity returned to normal 21 days
post exposure. Filtration of UV through mylar did not affect this
response; hence effects on NK activity were attributed to UVA
(Hersey et al., 1983a; Hersey et al., 1988).
9.2 Susceptibility to Tumours, Infectious and Autoimmune
Diseases
As previously indicated Yoshikawa et al. (1990) found that
suppression of contact sensitivity following UV exposure was more
common among skin cancer patients than in healthy subjects. In their
study only skin cancer patients developed an immunological
tolerance. They suggested that individuals who were phenotypically
sensitive to the immunosuppressive effects of UV were at greater
risk of skin cancer.
Adverse effects of UV on 4 types of human infections have been
reported. Smallpox lesions were made larger by exposure to sunlight
(Finsen 1901). Lesions from Herpes Simplex Virus(HSV) types I and II
were reactivated by exposure to UV (Spruance, 1985; Klein, 1986).
Using the criteria established by Yoshikawa et al. (1990) for the
UVB-S phenotype, Taylor et al. (1993) reported that 66% of
individuals who had a strong history of HSV lip lesions provoked by
sun exposure were UVB-S as compared to 40-45% in the general
population and 92% in skin cancer patients (Yoshikawa et al.,
1990). Also, exposure of immunosuppressed patients to sunlight led
to an increased incidence of viral warts caused by papilloma virus,
presumably due to UVB exposure (Boyle et al., 1984; Dyall-Smith &
Varigos, 1985). Hence there is some indication that UV may
exacerbate certain infections in humans.
Recent attention has been paid to the effects of UV on immune
response in patients with human immune deficiency virus (HIV)
because UV has been shown to activate HIV in vitro (reviewed by
Zmudzka & Beer, 1990). It has been suggested that UV exposure could
progress these patients to full blown AIDS by interfering with
protective immunity, since Th1 responses appear to be protective
against AIDS whereas Th2 responses are not (Shearer & Clerici,
1992). However, Warfel et al. (1993) found no difference in CD4+
cell counts in HIV patients before and after treatment with 50-60%
of their MED for dermatological disease. More research in this area
is needed to resolve these issues.
Finally, it has been known for some time that UV exposure
adversely affects the clinical course of systemic lupus
erythematosus, an autoimmune disease (Epstein et al., 1965);
however, the relationship of UV in this immune response is unclear.
9.3 Conclusions
The above studies suggest that UV exposures at environmental
levels suppress immune responses in both rodents and man. In rodents
this immune suppression results in enhanced susceptibility to
certain infections with skin involvement as well as some systemic
infections. The mechanisms associated with UV-induced
immunosuppression in rodents and man are similar. Also, host defense
mechanisms which provide protection against infectious agents are
similar in rodents and man as shown in figure 9.1. It is therefore
reasonable to assume that exposure to UV may enhance the risk of
infection and decrease the effectiveness of vaccines in humans.
Additional research is needed to substantiate these
assumptions. In particular, experimental studies are needed in
rodents and man to access effects on immune function parameters in a
fashion that allows quantitative comparisons between the two
species. Results of experiments on rodents showing the effects of UV
exposure on the susceptibility to various infections could be used
to extrapolate the risk of infectious disease in humans. Ultimately
epidemiological studies on the effects of UV exposure on
susceptibility to infection and vaccine effectiveness are needed to
validate this hypothesis.
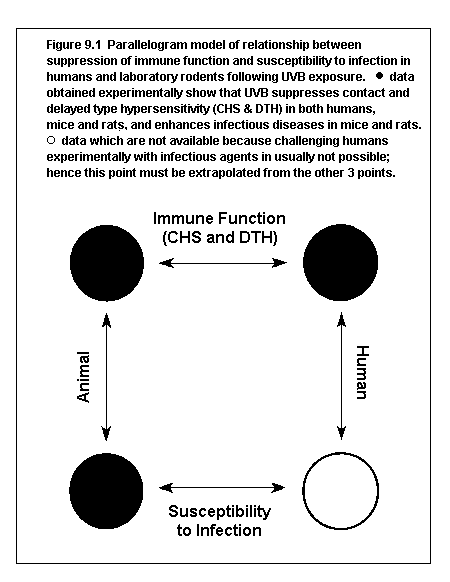 10. HUMAN STUDIES: THE EYE
10.1 Introduction
Since the early part of this century ophthalmologists have
suggested an association between sunlight exposure and UV and the
development of cataracts and other ocular effects (Widmark 1889,
1901; Hess 1907; Martin 1912; Birch-Hirschfeld, 1914; Verhoeff and
Bell, 1916; Duke-Elder, 1926b); however, only in the past twenty
years have epidemiological studies provided a scientific link. UV is
probably one of a number of factors associated with the development
of cataract. Despite the number of animal and human studies, many
questions remain as to the validity of interpretations of past data
and the biological and physical factors that influence the outcome
of UV exposure of the eye.
10.2 The Eye
The eyeball is deeply set in a bony orbital cavity, and the
upper bony ridge provides not only protection from mechanical injury
but also serves as a shield from overhead sky light. The upper and
lower eyelids (figure 10.1) also provide considerable protection by
serving as " shutters" against bright light. The eyeball consists
of three layers of tissue: (a) an outer protective layer, the sclera
and cornea; (b) a middle layer of blood vessels, pigment cells and
muscle fibres called the uvea; and (c) an inner, light sensitive
layer called the retina.
The sclera, the outer posterior layer, is a tough, thick,
opaque tissue formed of collagen fibres. The cornea, the anterior
transparent part of the eyeball, consists of multiple layers. The
surface epithelium continues with the surface epithelium of the
conjunctiva. The epithelial cells are known to be constantly
changing and the basal layer overlies the Bowman layer. The
outermost epithelial cells undergo rapid turnover, having a lifetime
at the surface of approximately 48 hours.
The main bulk of the cornea, the stroma, consists of highly
organized collagen fibres in a pattern that makes the cornea
transparent. The innermost layer, the endothelium, is a single layer
of an active ion pump (Na-K) that maintains the hydration state of
the stroma, an important factor for corneal transparency.
The middle layer of the eyeball consists of the iris
anteriorly, the ciliary body, and the choroid. The iris is a
diaphragm that adapts by changing the pupillary size according to
ambient light level. The iris is formed of mainly pigmented cells of
various densities, blood vessels and smooth muscle fibres that are
attached to the anterior part of the ciliary body. The smooth muscle
fibres form the sphincter and dilator of the pupil.
10. HUMAN STUDIES: THE EYE
10.1 Introduction
Since the early part of this century ophthalmologists have
suggested an association between sunlight exposure and UV and the
development of cataracts and other ocular effects (Widmark 1889,
1901; Hess 1907; Martin 1912; Birch-Hirschfeld, 1914; Verhoeff and
Bell, 1916; Duke-Elder, 1926b); however, only in the past twenty
years have epidemiological studies provided a scientific link. UV is
probably one of a number of factors associated with the development
of cataract. Despite the number of animal and human studies, many
questions remain as to the validity of interpretations of past data
and the biological and physical factors that influence the outcome
of UV exposure of the eye.
10.2 The Eye
The eyeball is deeply set in a bony orbital cavity, and the
upper bony ridge provides not only protection from mechanical injury
but also serves as a shield from overhead sky light. The upper and
lower eyelids (figure 10.1) also provide considerable protection by
serving as " shutters" against bright light. The eyeball consists
of three layers of tissue: (a) an outer protective layer, the sclera
and cornea; (b) a middle layer of blood vessels, pigment cells and
muscle fibres called the uvea; and (c) an inner, light sensitive
layer called the retina.
The sclera, the outer posterior layer, is a tough, thick,
opaque tissue formed of collagen fibres. The cornea, the anterior
transparent part of the eyeball, consists of multiple layers. The
surface epithelium continues with the surface epithelium of the
conjunctiva. The epithelial cells are known to be constantly
changing and the basal layer overlies the Bowman layer. The
outermost epithelial cells undergo rapid turnover, having a lifetime
at the surface of approximately 48 hours.
The main bulk of the cornea, the stroma, consists of highly
organized collagen fibres in a pattern that makes the cornea
transparent. The innermost layer, the endothelium, is a single layer
of an active ion pump (Na-K) that maintains the hydration state of
the stroma, an important factor for corneal transparency.
The middle layer of the eyeball consists of the iris
anteriorly, the ciliary body, and the choroid. The iris is a
diaphragm that adapts by changing the pupillary size according to
ambient light level. The iris is formed of mainly pigmented cells of
various densities, blood vessels and smooth muscle fibres that are
attached to the anterior part of the ciliary body. The smooth muscle
fibres form the sphincter and dilator of the pupil.
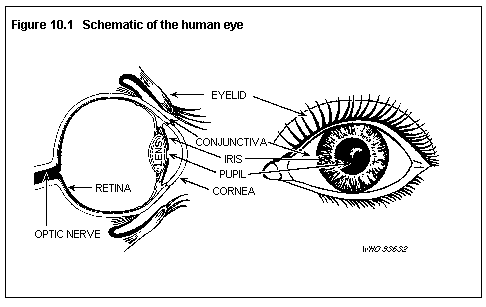 As part of the middle layer lining the sclera, the choroid
consists of a meshwork of blood vessels, nerves and pigment cells
that contribute to the nutrition of the retina and support the
function of the innermost layer, the retina, that contains the
photoreceptors and the neuronal network.
The lens, embryologically of ectodermal (skin) in origin,
consists of closely and orderly packed transparent elongated lens
cells that are enclosed in a capsule. New lens cells are constantly
formed at the so-called lens equator and old ones are displaced
towards the centre of the lens. The lens is suspended to the ciliary
body by fine ligaments. The lens shape can be changed by contraction
or relaxation of the ciliary muscles. This change provides a
focusing power to the eye that is called accommodation and is made
possible by the elasticity of the young lens. The lens is avascular
and obtains its nutrient from the aqueous fluid in front of it and
the vitreous body that fills the posterior cavity behind it.
Notably, the lens proteins cannot be renewed and therefore
accumulate lesions inflicted on them throughout life.
The photoreceptors, the rods and cones of the retina,
constitute the primary light receptor with rods functioning at low
light levels (scotopic vision) and cones operating at high light
levels (photopic vision). Thus the rod system subserves the function
of retinal sensitivity, whereas cones provide colour vision, high
resolution visual acuity and motion perception. The retinal pigment
epithelium is essential for the maintenance of photoreceptor
metabolism including, the transport, storage and regeneration of the
visual pigments.
Visible light (400-760 nm) incident upon the eye is strongly
refracted at the cornea, then transmitted through the aqueous humour
in the anterior chamber to the lens where it is refracted further.
After transmission through the vitreous gel-like structure, it
finally reaches the light-sensitive receptors in the retina. It is
these structures that are primarily damaged by UV and visible
radiations. The neuronal layers perform the complex task of
information processing. The primary visual signal is transformed and
ultimately transmitted to the visual cortex in the brain, thus
providing the image seen by the eye.
Most of the UV incident on the eye is absorbed in the tear
film, the cornea and the lens. The lens and the tissues in the
anterior part of the eye may however, be exposed to UV at
wavelengths above 295 nm and the retina is exposed to a fraction of
the incident UVA. Absorption of UV in the ocular media is given in
Fig 10.2 (Sliney & Wolbarsht, 1980). Boettner & Wolter (1962)
measured the transmission of direct forward scattering UV in the
cornea, aqueous humour, lens and vitreous humour from freshly
enucleated normal human eyes. The cornea absorbed all UV with
wavelengths <300 nm, while above 300 nm some UV was transmitted
through the cornea. About 60% of UV at 320 nm and 80% at 380 nm was
transmitted through the cornea. The aqueous humour transmitted most
incident UV (90% transmission at 400 nm) with no evidence of
scattering.
Recently Barker and Brainard (1993) quantified the change in UV
transmittance of the human lens with age. All of these studies
clearly show the steady decrease in UV transmittance of the lens
age. At birth there is a small window of transmission to the retina
at 320 nm. This window almost disappears by the second decade due to
an age-related yellowing of the human lens. As shown in Fig 10.3 the
lens absorption is strongest in the 340-380 nm band with somewhat
less absorption in the 310-320 nm range (Rosen, 1986; Barker and
Brainard 1993). The human lens is unique in that it contains a UVA
absorbing filter (O-beta of 3-hydroxykynurenine) which protects the
retina.
10.3 Study Design
This chapter reviews the epidemiological evidence for a causal
association between exposure to UV and development of specific eye
diseases which have at some point been linked with exposure to UV.
Two distinctive types of UV exposure assessments have been used in
the epidemiological studies. Some studies have related the
occurrence of eye disease to non-personal factors associated with
place of residence, such as meteorological data on average annual UV
dose or average annual hours of sunlight. Other studies have
obtained estimates of exposure at the individual level (e.g. hours
of sunshine exposure, lifetime exposure to UV) and related these to
disease occurrence.
Three types of epidemiological studies have been used to
investigate an association with UV exposure: geographical
correlation studies, cross-sectional studies, and case-control
studies. In the geographical correlation studies the prevalence of
eye disease in different areas has been related to non-personal
factors associated with place of residence. These studies are useful
for generating hypotheses but of limited value in testing a
particular hypothesis because observed correlations may result from
confounding by other factors which also vary geographically, and
because the level of exposure for persons with the disease is not
known.
As part of the middle layer lining the sclera, the choroid
consists of a meshwork of blood vessels, nerves and pigment cells
that contribute to the nutrition of the retina and support the
function of the innermost layer, the retina, that contains the
photoreceptors and the neuronal network.
The lens, embryologically of ectodermal (skin) in origin,
consists of closely and orderly packed transparent elongated lens
cells that are enclosed in a capsule. New lens cells are constantly
formed at the so-called lens equator and old ones are displaced
towards the centre of the lens. The lens is suspended to the ciliary
body by fine ligaments. The lens shape can be changed by contraction
or relaxation of the ciliary muscles. This change provides a
focusing power to the eye that is called accommodation and is made
possible by the elasticity of the young lens. The lens is avascular
and obtains its nutrient from the aqueous fluid in front of it and
the vitreous body that fills the posterior cavity behind it.
Notably, the lens proteins cannot be renewed and therefore
accumulate lesions inflicted on them throughout life.
The photoreceptors, the rods and cones of the retina,
constitute the primary light receptor with rods functioning at low
light levels (scotopic vision) and cones operating at high light
levels (photopic vision). Thus the rod system subserves the function
of retinal sensitivity, whereas cones provide colour vision, high
resolution visual acuity and motion perception. The retinal pigment
epithelium is essential for the maintenance of photoreceptor
metabolism including, the transport, storage and regeneration of the
visual pigments.
Visible light (400-760 nm) incident upon the eye is strongly
refracted at the cornea, then transmitted through the aqueous humour
in the anterior chamber to the lens where it is refracted further.
After transmission through the vitreous gel-like structure, it
finally reaches the light-sensitive receptors in the retina. It is
these structures that are primarily damaged by UV and visible
radiations. The neuronal layers perform the complex task of
information processing. The primary visual signal is transformed and
ultimately transmitted to the visual cortex in the brain, thus
providing the image seen by the eye.
Most of the UV incident on the eye is absorbed in the tear
film, the cornea and the lens. The lens and the tissues in the
anterior part of the eye may however, be exposed to UV at
wavelengths above 295 nm and the retina is exposed to a fraction of
the incident UVA. Absorption of UV in the ocular media is given in
Fig 10.2 (Sliney & Wolbarsht, 1980). Boettner & Wolter (1962)
measured the transmission of direct forward scattering UV in the
cornea, aqueous humour, lens and vitreous humour from freshly
enucleated normal human eyes. The cornea absorbed all UV with
wavelengths <300 nm, while above 300 nm some UV was transmitted
through the cornea. About 60% of UV at 320 nm and 80% at 380 nm was
transmitted through the cornea. The aqueous humour transmitted most
incident UV (90% transmission at 400 nm) with no evidence of
scattering.
Recently Barker and Brainard (1993) quantified the change in UV
transmittance of the human lens with age. All of these studies
clearly show the steady decrease in UV transmittance of the lens
age. At birth there is a small window of transmission to the retina
at 320 nm. This window almost disappears by the second decade due to
an age-related yellowing of the human lens. As shown in Fig 10.3 the
lens absorption is strongest in the 340-380 nm band with somewhat
less absorption in the 310-320 nm range (Rosen, 1986; Barker and
Brainard 1993). The human lens is unique in that it contains a UVA
absorbing filter (O-beta of 3-hydroxykynurenine) which protects the
retina.
10.3 Study Design
This chapter reviews the epidemiological evidence for a causal
association between exposure to UV and development of specific eye
diseases which have at some point been linked with exposure to UV.
Two distinctive types of UV exposure assessments have been used in
the epidemiological studies. Some studies have related the
occurrence of eye disease to non-personal factors associated with
place of residence, such as meteorological data on average annual UV
dose or average annual hours of sunlight. Other studies have
obtained estimates of exposure at the individual level (e.g. hours
of sunshine exposure, lifetime exposure to UV) and related these to
disease occurrence.
Three types of epidemiological studies have been used to
investigate an association with UV exposure: geographical
correlation studies, cross-sectional studies, and case-control
studies. In the geographical correlation studies the prevalence of
eye disease in different areas has been related to non-personal
factors associated with place of residence. These studies are useful
for generating hypotheses but of limited value in testing a
particular hypothesis because observed correlations may result from
confounding by other factors which also vary geographically, and
because the level of exposure for persons with the disease is not
known.
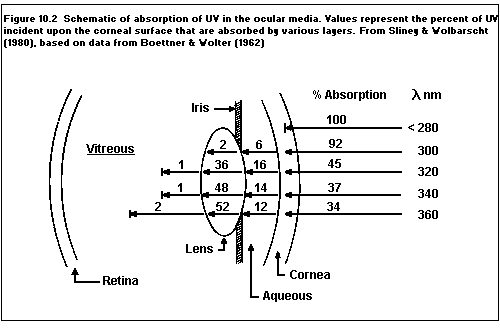
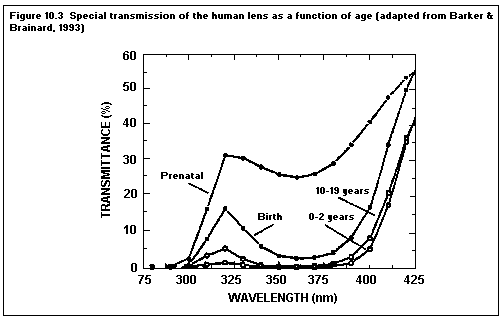 The second, and most common type of study design, has been the
cross-sectional study in which a population or occupational group is
surveyed and disease prevalence measured. Cross-sectional surveys
identify all persons with the disease, some of which are new cases
while other persons may have had the disease for a period of time.
Some cross-sectional studies have related non-personal factors
associated with place of residence to disease status. Other studies
have collected detailed information from each study participant on
personal exposure to UV or indices of exposure (e.g. hours of
exposure to sunshine, occupation) and related these exposures to
disease status.
The third type of study design has been the case-control study,
where differences in UV exposure between persons with the disease
and those without have been compared. Cases have usually been drawn
from a hospital or clinic. Controls have been drawn from other
hospital or clinic patients or from the general population and have
usually only included persons with good visual acuity, without the
disease of interest, and without a disease that is associated with
UV exposure. Some case-control studies have related disease status
to non-personal factors associated with place of residence while
others have used personal exposure information.
Data from both cross-sectional and case-control studies can be
useful in confirming a hypothesis, but have a number of limitations.
If there is an excess risk of death associated with the disease, as
has been suggested for cataract (Minassian et al., 1992; Vitale
et al., 1992), both types of study will be biased towards the
survivors. In addition, disease status and prior exposure indices
are measured at the same time and it may not be possible to
differentiate between cause and effect, especially if the disease
has a long latency period.
10.4 Diseases of the External Eye
10.4.1 Photokeratitis and photoconjunctivitis
Cases of photokeratitis and photoconjunctivitis have occurred
between 0.5 and 24 hours after prolonged exposure to intense solar
radiation, often in highly reflective environments (Wittenberg,
1986). The most severe cases are usually manifested as snow
blindness, suggesting that UV is the cause of this condition.
The action spectrum for UV photokeratitis produced in the
rabbit was first measured by Cogan and Kinsey (1946). Pitts (1974,
1978) in a series of laboratory studies on humans estimated the mean
threshold of UVB (290-315 nm) for photokeratitis at 3500 J m-2.
These laboratory data are supported by Blumthaler et al. (1987),
who estimated that the radiant exposures in clinically observed
cases of photokeratitis ranged from 1200 to 5600 J m-2. It is
estimated that 100 to 200 seconds of direct, unattenuated exposure
to 295-315 nm solar radiation will result in photokeratitis (Sliney,
1987; Wittenberg, 1986). Blotting out the solar disc would remove
around 40% of the UV, still leaving a threshold of around 5.5
minutes. Sliney (1986) has estimated that the reflected levels of UV
from light sand should be sufficient to cause a threshold
photokeratitis within exposure periods of 6-8 hours centred around
midday, and within 1 hour for UV reflected from snow.
Experimental data shows photokeratitis can be induced in
animals by UVB exposure and that use of UVB absorbing contact lens
or chromophores can prevent UVB induced photokeratitis in laboratory
animals. Collectively, there is sufficient experimental and
epidemiological evidence that exposure to intense UVB radiation
causes photokeratitis and photoconjunctivitis.
10.4.2 Climatic droplet keratopathy
Climatic droplet keratopathy, among a variety of other names
(Gray et al., 1992), is also known as spheroidal degeneration from
its histological appearance. It is a degenerative condition usually
affecting both eyes symmetrically, and restricted to the exposed
interpalpebral band of the cornea. This condition is of major
significance for vision in some parts of the world, reducing vision
to blindness levels in older people. For example, in Mongolia it has
been found in an initial survey to be the third cause of blindness
(Baasanhu et al., in press).
Climatic droplet keratopathy occurs throughout the world, but
is more common in areas with snowfall persisting late into the
summer in the northern hemisphere, such as parts of northern Canada,
Siberia and Mongolia, and in areas of sand and desert in other
latitudes, including Somalia, the Arabian peninsula, Iran, and
Australia. It is also particularly common on sea coasts where there
is coral sand or the sand is impregnated with salt, such as the
islands of the Red Sea (Gray el al., 1992).
In a cross-sectional study of Australian aborigines Taylor
(1980a) found no correlation between the prevalence of climatic
droplet keratopathy and ambient UVB levels, although the condition
was more common among those working as stockmen. Johnson (1981)
reported a geographical correlation with the calculated flux of
reflected UV from snow and ice throughout the year in the eastern
coast of Newfoundland and Labrador and the eastern Arctic of Canada.
In a cross-sectional study of Chesapeake Bay waterman study
Taylor et al. (1989) examined the risk of climatic droplet
keratopathy with chronic UVB exposure. Although a positive
association was found (RR= 6.4, 95%CI=2.5-11.7) for those in the
highest quarter of exposure compared to those with the bottom
quarter, further analyses of this data (Taylor et al., 1992)
showed the risk of climatic droplet keratopathy was also related to
UVA.
There is strong evidence that the corneal degeneration is due
to environmental factors. Circumstantial evidence exists that it is
caused by solar UV, mainly reflected from ground surfaces such as
snow and sand which are particularly reflective of UV.
Histologically, the material deposited in the superficial corneal
stroma as spheroidal droplets is most likely to be derived from a
mixture of altered plasma proteins, including fibrinogen, albumin,
and immunoglobulins (Johnson & Overall, 1978).
Other proposed aetiological agents such as low atmospheric
humidity, low temperature or high temperature have been excluded. It
is possible that particulate injury by wind-blown ice or snow or
sand particles may contribute to the development of the condition by
causing inflammation and therefore outpouring of additional plasma
proteins from the blood vessels of the limbs.
10.4.3 Pinguecula
Pinguecula is a fibro-fatty degeneration of the interpalpebral
conjunctiva. The pathological changes that occur in pinguecula are
similar to actinic elastosis of the skin, a condition thought to be
linked to sunlight exposure. This indirect evidence suggests that
exposure to sunlight may be a risk factor for pinguecula.
Geographical variation in the occurrence of pinguecula has been
reported, with higher prevalence in Arabs living near the Red Sea
than in Eskimos from Greenland or Caucasians in Copenhagen (Norn,
1982). Johnson et al. (1981) in a study of pinguecula in Labrador
found the size of pinguecula was correlated with the severity of
climatic droplet keratopathy. Taylor et al. (1989) in the study of
Chesapeake Bay watermen found a weak association for the presence or
absence of pinguecula with exposure to UVA and UVB. The relative
risk for the top quartile of exposure was 1.4 (95%CI=0.9-2.2), less
than for climatic droplet keratopathy or pterygium. Karai &
Horiguchi (1984) in a study of 191 Japanese welders found no
difference in the occurrence of pinguecula between welders and
controls.
It is concluded that there is currently insufficient
epidemiological or experimental data for an assessment of the risk
of pinguecula with exposure to UV.
10.4.4 Pterygium
Pterygium is a triangular shaped degeneration and hyperplastic
process in which the bulbar conjunctiva encroaches on the cornea.
A geographical association between variation in the occurrence
of pterygium and variation in sunlight exposure was first suggested
by Talbot (1948). Based on observation of pterygium in New Zealand
and South Pacific Islands Elliott (1961) suggested pterygium in
these locations resulted from UV exposure.
Studies of non-personal factors associated with place of
residence
In a study of pterygium patients in US Veterans Administration
hospitals during 1957-59 Darrell & Bachrach (1963) related mean
daily UV (319nm) levels to the ratio of pterygium to all hospital
discharges. A trend was found between UV level and pterygium ratio
for persons born in rural counties. A similar association with UV
level was seen for persons residing in a rural county at the time of
the hospital discharge. Cameron (1965) examined the global pattern
of pterygium and reported an inverse gradient with latitude. In a
study of Australian aborigines Taylor (1980a) found pterygium was
correlated with ambient UVB level and hours of sunshine at place of
residence. Data from Canada indicates that pterygium is also common
in arctic and sub-arctic environments (Johnson et al., 1981).
Moran & Hollows (1984) found a nonsignificant increase in the
prevalence of pterygium among Australian aborigines residing in
areas with higher ambient UV levels.
Studies with personal exposure measurements
Four studies have related occurrence of pterygium to personal
measurements. Karai & Horiguchi (1984) examined 191 Japanese welders
for the presence of pterygium. A trend of increasing risk of
pterygium was found with years of employment as a welder, an
indirect measure of cumulative occupational exposure to UV.
Booth (1985) undertook a hospital-base case-control study of
pterygium in Sydney. No difference between cases and controls was
found in subjective assessment of exposure to sunlight in work or
sport. However, a family history of pterygium was found to be a
strong risk factor.
Among Chesapeake Bay watermen, Taylor et al. (1989) found a
dose-response relationship between risk of pterygium and exposure to
UVA and UVB. The relative risk for the top quarter of UVB exposure
was 3.1 (95%CI=1.8-5.3) compared to the lowest quarter. However, it
is noted that pterygium was equally associated with ocular exposure
to UVA and visible light.
Mackenzie et al. (1992) undertook a hospital-based
case-control study of pterygium in Queensland. A strong
dose-response relationship was found with closeness of place of
residence to the equator, type of outdoor work environment (e.g.
sandy) and amount of time spend outdoors. The most striking finding
was the magnitude of risk associated with spending most of the time
outdoors was stronger when related to childhood exposure (RR=17.2,
95%CI=6.2-47.6) than to adult exposure (RR=5.7, 95%CI=3.1-10.6). The
risk associated with working at ages 20-29 in an outdoors
environment of mainly sand or concrete was associated with a
relative risk of 11.3 compared with indoor workers. Corneo (1993)
has suggested that the cornea is acting as a side on lens focusing
light and also UV across the anterior chamber to the nasal limbus.
This hypothesis may explain why pterygia usually commence on the
nasal side of the eye.
Evaluation of epidemiological evidence
While several geographical studies have reported an inverse
trend with latitude, the common occurrence of pterygium in arctic
and subarctic locations suggest that closeness to the equator does
not fully explain the distribution of this disease.
The strength of the association with time spent outdoors
reported by Mackenzie et al. (1992) suggests that the association
may be causal. However, there is insufficient evidence to show that
the observed association with UV exposure is not, in part, due to
confounding. The findings from three of the studies lend support to
a hypothesis that irritation by particulate matter is associated
with pterygium. The Australian aborigines live in a dry, dusty
environment and welders are occupationally exposed to a range of
particles. Similarly, the Queensland study found highest risk among
those who worked in sandy locations. The particulate matter
hypothesis is also supported by a report (Dhir et al., 1967) of
higher prevalence of pterygium among Punjabi Indians working in
sawmills (an indoor occupation) in New Delhi and British Columbia
than Punjabi farmers (an outdoor occupation). Similar findings of
increased risk of pterygium among sawmill workers in Thailand and
Taiwan have been reported (Detels & Dhir, 1967). The evidence of
possible confounding by particulate matter is inconsistent, with the
Chesapeake Bay watermen study finding an association with sunlight
exposure in a location that was neither hot, dry or dusty.
It is not possible, based on available epidemiological data, to
assess the risk of pterygium with exposure to UV because of possible
confounding of observed associations by exposure to particulate
matter or other factors.
10.4.5 Hyperkeratosis, carcinoma-in-situ, and squamous cell
carcinoma of the conjunctiva
These conditions probably form a gradation of development and
cannot necessarily be distinguished clinically. Invasive squamous
cell carcinoma is often said to arise from a pre-cancerous lesion.
Epithelial dysplasia and carcinoma-in-situ look the same, and are
sometimes keratinized and present as leucoplakia in which case the
term actinic keratosis may be applied (Naumann & Apple, 1986;
Garner, 1989). The main argument for an actinic causation is that
these tumours usually present in the exposed area of the eye between
the lids (the interpalpebral fissure) and under conditions where
they may be expected to be exposed to solar radiation.
Xeroderma pigmentosum is a recessively inherited syndrome
characterized by clinical and cellular hypersensitivity to solar
radiation and a defect in the capacity to repair UV-induced damage
in DNA (Fitzpatrick, et al., 1963). Among reports of 337 patients
with xeroderma pigmentosum for whom ocular findings had been
described, Kraemer et al. (1987) identified 88 ocular tumours of
which 73 were specific to the corneal-scleral limbus (34), the
cornea (24) or the conjunctiva (15). Of the non- melanomas for which
histopathological type was specified, 28 were squamous cell
carcinomas and 12 were basal cell carcinomas. Among 64 patients with
ocular neoplasms whose age was stated, half the neoplasms had
occurred before 11 years of age. While the eyelids are a site of
preference for basal cell carcinoma, this tumour rarely, if ever,
arises in the conjunctiva in otherwise normal individuals.
Squamous cell carcinoma of the conjunctiva is an uncommon
tumour. Garner (1989) reviewed all cases of tumours at the limbus
sent over a 40 year period for examination to the Institute of
Ophthalmology, London. The total was only 636 tumours, of which 73
were squamous carcinomas. This amounts to less than 2 cases per year
coming to the Pathology Laboratory, even though Moorfields Eye
Hospital which the Laboratory serves, attracts patients from all
over the country and from overseas.
Lee & Hirst (1992) attempted to provide population-based
figures and estimate the incidence of these tumours in metropolitan
Brisbane (latitude 30° south). They surveyed the histological
records of all ocular surface tumours examined in the pathological
laboratories over the previous 10 years, serving a population of
more than 745,000 in 1989. There were 139 cases of which 79 were
corneal epithelial dysplasia, 28 carcinomas-in-situ and 32 were
squamous cell carcinomas. There was a strong male preponderance. The
incidence ranged from 1 per 100,000 in 1980 to 2.8 per 100,000 in
1982. This is well below the rate for squamous cell carcinoma of the
skin and melanoma of the skin in Queensland as a whole. On the other
hand, it is a substantially higher rate than that recorded in London
where the pathological laboratory referred to also covers a much
larger population.
Squamous cell carcinoma of the conjunctiva has been reported to
form a greater proportion of eye tumours in Africans living in areas
close to the equator (Templeton, 1967) than in the south of Africa
(Higginson & Oettlé, 1959), and much higher than in Baltimore
(39°N). The incidence (0.3 per 100,000) in Uganda (0°) has been
reported to be twice that in Denmark (55°N) despite the potential
underascertainment in Africa (Templeton, 1967).
It is extremely rare for a neoplasm to arise de novo in the
corneal epithelium, where it may be called a corneal intra-
epithelial neoplasm. Most such intra-epithelial sheets are connected
at the corneo-scleral limbus to a conjunctival lesion, such as a
papilloma or a leucoplakia over a pterygium or pinguecula (Waring
et al., 1984). The only available evidence for an UV aetiology is
the location of the lesion within the interpalpebral fissure, and
the fact that it may arise from a lesion which is itself associated
with UV. Three cases of corneal intra-epithelial neoplasia have been
recently reported in people aged 31 to 38 who wore contact lenses
and were considered to have had substantial exposure to artificial
and solar UV (Guex-Crosier & Herbort, 1993).
10.5 Diseases of the Lens
10.5.1 Cataract
For the purpose of this review, a cataract is defined as an
opacity of the lens of the eye. The three major types of cataract
are cortical, nuclear and posterior subcapsular (PSC). When a lens
opacity interferes with vision, a clinically significant cataract is
present. If left untreated, cataract will often progress to
blindness. Cataract causes half of the world's blindness.
Definitions of cataract and methods used to assess the presence
and severity of cataract have not been uniform in epidemiological
studies of cataract. Many studies include lens opacities that are
not necessarily accompanied by a decrease in visual acuity. Some
have combined all three major types of lens opacities into a single
"cataract" category, while others have investigated associations for
specific types of opacity. Methods of assessing the presence and
sometimes the severity of opacities range from reviews of existing
charts to clinical examinations using written definitions of
cataract, and the use of standardized grading systems that have been
found to be highly reliable.
Occupational case series
When cataracts result from occupational exposure to UV, it may
be difficult to differentiate between the contribution of
occupational and non-occupational factors to the development of the
disease. Lerman (1980) described the onset of lens opacities in
three persons who worked in a dental clinic and were exposed to UV
(300-400 nm) from a dental curing unit. The lens damage varied from
posterior subcapsular cataract in the dentist, who was reported to
have received the highest dose, to zonular type opacities in one of
the dental assistants. However, any retrospective reconstruction of
the actual ocular exposure has a large degree of uncertainty, and
the results from such an exercise must be interpreted with caution.
Studies in which UV exposure was inferred from place of residence
Selected studies of humans exposed to solar UV are presented in
tables 10.1 and 10.2. Studies were selected for inclusion in the
tables on the basis of the scientific quality of the published
report and the overall contribution of the paper to the evaluation
of the UV-cataract hypothesis. The tables do not include the
relative risks for other factors, which in some instances are higher
than for sunlight or UV exposure.
Hiller et al. (1977) investigated sunlight and cataracts
using data from the large sources (blindness registries in 14 states
and the cross-sectional Health and Nutrition Examination Survey
(HANES) of 35 geographic areas of the US) and US Weather Bureau
geographical data on annual hours of sunlight in each geographical
area. Above the age of 65 the prevalence of cataract increased with
annual hours of sunlight, with the highest prevalence found in
locations with 3000+ annual hours of sunshine. At ages 45-64 there
was some evidence of an association with hours of sunshine but the
gradient was weaker. Below age 45 there was little association
between annual hours of sunshine and prevalence of cataract. In a
further analysis of HANES data Hiller et al. (1983) reported a
correlation between average daily UVB levels and prevalence of
cataract. Analysis of HANES data by type of cataract (Hiller et
al., 1986) revealed UVB levels at location of residence were
associated with pure cortical cataract but not with pure nuclear or
posterior subcapsular cataract.
The prevalence of cataract among Australian aborigines was
found to be correlated with annual ambient UVB level at place of
residence (Taylor et al., 1980b). Hollows & Moran (1981) found the
prevalence of cataract was highest among aborigines living in the
north of Australia, an area with high average daily UVB radiation.
Mao & Hu (1982) studied age related cataract in seven rural areas of
China and found the prevalence of cataract was correlated with
annual direct solar radiation.
Residents of rural villages in Nepal had a prevalence of
cataract related in different zones of the country to average hours
of sunshine (Brilliant et al., 1983). The prevalence was higher in
the plains where there were 12 hours of direct sunshine compared to
the mountains with 7-9 hours per day. Factors such as use of glasses
and hats modify personal ocular exposure to UV and should be
assessed.
TABLE 10.1 SELECTED HUMAN CATARACT STUDIES WITH SUNLIGHT OR UV EXPOSURE BASED ON RESIDENCE
AUTHOR POPULATION MEASURE OF MEASURE OF ASSOCIATIONS OBSERVED COMMENTS
OUTCOME SUNLIGHT EXPOSURE
Hiller et al. MRA: 9110 persons Blind from cataract, Average hours of RR= 3.3 (age 65-74) Adjusted for age and
(1977, 1983, registered as blind visual acuity (VA) sunlight < 2400 vs sex; blind registry data
1986) in 14 US States; 6/60 3000+
whites;
NHANES: 3580 persons; Lens opacity and VA Average hours of RR= 2.7 (age 65.74) Adjusted for age and sex
probability sample of < 6/7.5 sunlight < 2400 vs
US population; 3000+
NHANES data Lens opacity and VA Average daily UVB RR= 1.58 (age 45-74, Adjusted analysis
< 6/9 count in area of p<.05)
residency 6000 vs 2600
NHANES data Nuclear and cortical Average daily UVB RR= 3.6 for cortical Adjusted analyses; pure
opacity count in area of opacity; no association opacity types only
residency 6000 vs 2600 with nuclear opacity
Taylor (1980b) Survey of 350 Lens opacity with Average daily sunlight RR= 4.2 (95% CI=0.9-18.91 Unadjusted for age or
Australian Aborigines good vision, poor hours in area of potential confounding
vision or blindness residence: 9.5+ vs < 8 factors
Annual mean UVB RR= 1.8 (95%ci=0.9-3.4)1
radiation level for
area of residence: 1 95%CI estimated from
3000 vs 2000 published data
Hollows & Moran Survey of 64,307 Lens opacity and VA Average daily UVB Significant positive Wide age bands;
(1981) Aborigines and 41,254 < 6/6 count in 5 zones of correlation between unadjusted analyses
non-Aborigines, Australia: 3000 vs prevalence of lens
Australia 1000 opacity and UVB counts
in Aborigines; no
association in
non-Aborigines
TABLE 10.1 (contd).
AUTHOR POPULATION MEASURE OF MEASURE OF ASSOCIATIONS OBSERVED COMMENTS
OUTCOME SUNLIGHT EXPOSURE
Brilliant et Survey of 27,785 Lens opacities or Average daily sunlight RR= 3.8 (p<.001) Adjusted for age and
al. (1983) Nepalese; national aphakia hours: 12 vs 712 vs sex; RR decreased with
probability sample; 7-9 increasing altitude; sun
lifelong residents; blocked by mountains at
high elevations
12 vs 712 vs 7-9 RR= 2.6 (p<.005)
Cruickshanks et Cross-sectional Nuclear, cortical Average annual ambient UVB exposure associated Adjusted for other risk
al. (1993) survey of 4926 and PSC opacities UVB exposure with cortical opacities factors; measure of
persons, Wisconsin, in men (RR=1.36, exposure represents
USA 95%CI=1.02-1.79) but not average potential
women, not associated exposure at residency;
with nuclear or PSC also Table 9.2
opacities
TABLE 10.2 SELECTED HUMAN CATARACT STUDIES WITH INDIVIDUAL UV EXPOSURE ASSESSMENT
AUTHOR POPULATION MEASURE OF MEASURE OF SUNLIGHT ASSOCIATIONS OBSERVED COMMENTS
OUTCOME EXPOSURE
Collmann et Clinic-based case Nuclear, cortical Average annual sunlight No significant association with Low power to
al. (1988) control study of 113 and PSC opacities exposure, based on any type of opacity detect
cases and 168 residential history and association;
controls, North amount of time spent in sun matched on age
Carolina, USA; whites and sex
Taylor et Cross-sectional Nuclear and Cumulative ocular exposure Dose-response relationship in High exposure
al. (1988) survey of 838 cortical to UV since age 16, based which a doubling of cumulative UVR study population;
watermen, Maryland, opacities on life history and ocular exposure increased risk of detailed ocular
USA exposure model cortical opacity by 1.60 exposure model
(95%CI=1.01-2.64). RR= 3.30
(95%CI=0.90-9.97) for highest vs
lowest quartile. No association
between ocular exposure to UVR and
nuclear opacity
Bochow et Clinic-based PSC cataract Cumulative ocular exposure Increased levels of UVB exposure Adjusted analyses;
al. (1989) case-control study of (surgical since age 16, based on life associated with increased risk of association
168 cases and 168 patients) history and exposure model PSC cataract present when
controls, Maryland, adjusted for
USA cortical cataract
Dolezal et Clinic-based Cataract Lifetime sunlight exposure, No association between lifetime Only partially
al. (1989) case-control study of (scheduled for based on life history, sunlight exposure and risk of adjusted for
160 cases and 160 surgery) amount of time in sun and cataract; use of head covering potential
controls, Iowa, USA use of glasses and hat reduced risk of cataract in males confounding
(RR=0.48, 95%CI=0.25-0.94) factors; crude
index of exposure,
low power
TABLE 10.2 (contd).
AUTHOR POPULATION MEASURE OF MEASURE OF SUNLIGHT ASSOCIATIONS OBSERVED COMMENTS
OUTCOME EXPOSURE
Italian-Am. Clinical-based Nuclear, Work location in the Cortical and mixed opacities Adjusted for other
study (1991) case-control study of cortical, PSC and sunlight; leisure time in associated with work location in risk factors;
1008 cases and 469 mixed opacities the sunlight; use of sunlight (RR=1.75, crude indices of
controls, Italy glasses and hat 95%CI=1.15-2.65), leisure time in exposure; also
sunlight (RR=1.45, Table 9.1
95%CI=1.09-1.93). Cortical, PSC
and mixed opacities associated
with use of a hat in summer
(RR=1.80, 95%CI=1.17-2.47). No
association between sun exposure
indices and nuclear opacities
Leske et al. Clinical-based Nuclear, cortical Work in sunlight; leisure Work in sunlight significantly Analyses adjusted
(1991) case-control study of and PSC mixed time in sun; residence and reduced risk of nuclear opacity for other risk
945 cases and 435 opacities travel to areas of high sun (RR=0.61, 95%CI=0.37-0.99); no factor
controls, exposure, use of hat and significant associations between
Massachusetts, USA sunglasses exposure and cortical or PSC
opacities
Cruikshanks Cross-sectional Nuclear, cortical Leisure and work time No associations with cortical Adjusted for other
et al. survey of 4926 and PSC opacities outside; use of glasses and opacities; reduced risk of nuclear factors; crude
(1993) persons, Wisconsin, hat and PSC opacities amount men for indices of
USA outdoor leisure time in winter; exposure; also
use of hats and sunglasses Table 9.1
significantly increased risk of
PSC opacity in women
Age and sex adjusted prevalence for all types of cataract in
persons aged 40 years and older was found to be 60% greater in Tibet
than in Beijing (14.6% versus 9.1%, p>0.001) (Hu et al., 1989).
The authors suggested a relationship with higher UV at the higher
altitudes of Tibet, but confounding factors could not be excluded
and prevalences were higher in women than men.
Studies with personal exposure measurements
A number of studies have collected information from each study
participant and estimated personal exposure to either sunlight or
UV. Factors such as use of glasses and hats should be assessed. The
characteristics of selected studies are outlined in Table 10.2.
In a cross-sectional study of cataract in the Punjab related
prevalence of cataract to work environment Chatterjee et al.
(1982) found a suggestion of lower cataract incidence among men
whose main work location was outdoors (RR=0.7, 95%CI=0.5-1.1).
Collman et al. (1988) examined lifetime exposure to sunlight
in a clinic-based case-control study of cortical, nuclear or PSC.
Lifetime exposure to sunlight was estimated from intensity of solar
radiation in area of residence, years of residence and average
amount of time spent outdoors during daylight hours. A
non-significant risk (RR=1.1) of cataract was found for the highest
category of lifetime exposure to sunlight.
Personal exposure history and ambient UVB data were combined to
estimate an individual's lifetime annual ocular exposure to UVB
after age 15 in a cross-sectional study of Chesapeake Bay watermen
(Taylor et al., 1988). This included information on occupational
and leisure exposures, type of work surfaces, seasons, and use of
head wear and eyewear. A moderate association with a trend of
increasing risk with exposure to UVB was seen for cortical cataract,
with a RR of 3.3 (95%CI=0.9-10.0) for the top quarter of exposure
relative to the bottom quarter. A nonsignificant association was
also found between exposure to UVA (320 -340 nm) and prevalence of
cortical cataract. Little evidence was found for an association
between UVA or UVB exposure and nuclear cataract. It is noted that
UVA and UVB exposures were highly correlated and that the study
would not have been able to differentiate between the effects of UVA
and UVB. Further analyses suggested a significant difference between
the cumulative lifetime ocular exposure among cases of cortical
cataract compared to non- cataract controls. No threshold or latency
period was observed.
In a clinic-based study of PSC cataracts in Maryland, cases
were persons who underwent PSC extraction in an ophthalmic practice
(Bochow et al., 1989). Controls, matched on age, sex, and type of
referral were chosen from other patients on the appointment book of
the same ophthalmic practice who did not have a PSC cataract or a
previous cataract extraction. Annual and cumulative ocular UVB
exposures were estimated for each individual using the same method
as the studies of Chesapeake Bay watermen. Thirty-nine percent of
cases had a pure PSC cataract, the remaining 61% had mixed PSC and
other cataracts. Almost half the controls had a non PSC cataract
(nuclear, cortical or other lens opacity). UVB exposure was
significantly associated with PSC cataracts. Both the average
cumulative exposure and average annual exposure were higher in cases
than controls, after adjusting for steroid use, eye colour,
education, diabetes and presence of cortical cataracts.
In a hospital-based study of cataract patients in Iowa Dolezal
et al. (1989) found little evidence of an association between
individual lifetime sunlight exposure and cataract. Mohan et al.
(1989) in a similar study of cataract in New Delhi examined a range
of environmental factors, including occupation. An increase in cloud
cover was significantly associated (RR=0.8, 95%CI=0.7-0.9) with
cataract when adjusted for each of the other environmental
variables. The study did not quantify individual lifetime exposure
to sunlight or UV.
In a study of cataract patients in a Massachussetts hospital,
Leske et al. (1991) investigated occupational exposure to
sunshine. No association was found for PSC cataract (RR=1.3,
95%CI=0.7-2.3), cortical cataract (RR=0.9, 95%CI=0.6-1.3), or mixed
cataract (RR=0.8, 95%CI=0.6-1.1) among those with at least 2 hours
of exposure to bright sunshine per day for at least 2 months. The
risk of nuclear cataract was reduced (RR=0.5, 95%CI=0.3-0.9).
A hospital-based study from Italy (Italian-American Cataract
Study Group, 1991) found an excess of pure cortical and mixed
cataract (RR=1.8, 95%CI=1.2-2.6) and a nonsignificant deficit of
nuclear (RR=0.6) and PSC cataract (RR=0.8) among those with a work
location in the sunlight. Leisure time spent in the sunlight was
associated with an excess of cortical and mixed cataract (RR=1.4,
95%CI=1.1-1.9) and a nonsignificant deficit of posterior subcapsular
cataract (RR=0.6).
In an Indian clinic-based study of cataract and history of
severe diarrhoeal diseases Bhatnagar et al. (1991) found an
elevated risk of cataract (RR=2.1, 95%CI=1.2-3.6) for outdoor
occupations compare with indoor occupations. However Zaunuddin &
Saski (1991) found no relationship between hours of exposure to
sunshine and prevalence of nuclear or cortical cataract in Sumatra
(0° latitude). In Beaver Dam, Wisconsin, Cruickshanks et al.
(1993) found no association between average annual exposure to UVB
and cortical, PSC or nuclear cataract. Wong et al. (1993) surveyed
fishermen in Hong Kong. A sun exposure score was calculated based on
daily sunlight exposure, and protection from use of a canopy, hat,
and glasses. The highest grades of cataract of all types considered
together were more common in subjects with the highest sun exposure
scores, but none of these associations was significant at the 5%
level. A population-based case-control study (Shibati et al.,
1993) reported an increased risk of cortical cataract among men aged
40-50 years who spent 5 or more hours per day outdoors compared with
those who spent less time outdoors (RR = 6.89; 95%CI = 1.22-39).
Evaluation of epidemiological evidence
An association has been demonstrated between prevalence of
cataract and residence in areas at low latitudes, with long hours of
sunlight or high ambient UV radiance in several studies undertaken
in different parts of the world. However, in each study, the
observed association may be confounded by other possibly causal
factors. Certain of the earlier studies did not classify the lens
opacities into types of cataract.
Cortical cataract was examined separately in four studies. Only
one study assessed individual exposure. Taylor et al. (1988) found
a dose-response relationship with exposure to UVB radiation. The
relative risk for the highest exposure category was three times that
for the lowest exposure category. It is unlikely that the exposure
assessment was able to distinguish between UVA and UVB exposure. The
other two used simple measures of sun-related behaviour. Leske et
al. (1991) found no association between exposure to bright
sunshine and cortical cataract, while in the Italian-American
Cataract Study (1991), a work location in the sunlight was related
to cortical and mixed cataract. The Italian study also found an
association between leisure time outdoors and cortical and mixed
cataract. The other two studies showed non-significant trends in
opposite directions. More recently, Cruickshanks et al. (1993)
found annual UVB exposure was associated with cortical opacities
among men, but no association was found for women.
Four studies report risk estimates for posterior subcapsular
(PSC) cataracts. Bochow et al. (1989) measured individual exposure
and found PSC cataract patients had higher annual and cumulative
exposures to UVB than controls, even after allowing for the effects
of several other factors. The other two studies used simple measures
of sun-related behaviour and showed non-significant trends in
opposite directions. Leske et al. (1991) found elevated risk for
pure PSC cataract patients compared to controls. However, the
Italian-American Cataract Study (1991) reported reduced risk for
pure PSC cataract patients with a work location in the sunlight or
who spent leisure time in the sunlight. Cruickshanks et al. (1993)
found no association between annual UVB exposure and risk of PSC
cataract.
Five studies provide risk estimates separately for nuclear
cataracts. These studies are consistent in showing no association
between UV exposure and nuclear cataract (Taylor et al., 1988;
Dolezal et al., 1989; Leske et al., 1991; The Italian-American
Cataract Study, 1991; and Cruickshanks et al., 1993). Collectively
these studies are consistent in showing no association between UV
exposure and nuclear cataract.
All of the published epidemiological studies of UV and cataract
have been challenged by the enormous difficulty of determining
ocular exposure in different climates. As noted previously, the
cornea and lens are seldom directly exposed to light rays from much
of the sky; hence the sunlight scattered from the ground and the
horizon determine the actual accumulated UV dose.
These studies clearly demonstrate that UV is at least one
aetiologic factor in cataractogenesis. However, extrapolation of
strong associations found in a mid-latitude population where no
serious nutritional problems are present (e.g. Taylor, et al.,
1988) to a tropical population in less developed regions, may not be
valid, since the contribution of UV relative to other factors such
as malnutrition and dehydration may be far more important.
10.5.2 Exfoliation syndrome
The exfoliation syndrome (pseudoexfoliation of lens capsule)
consists of abnormal material deposited on or arising from various
parts of the anterior eye. This condition was originally described
from Finland by Lindberg (1917). In the Nordic countries it
contributes to a high proportion of glaucoma in the older
population. This appears as a round area in the centre of the
anterior lens capsule, corresponding to normal pupil size, on which
bluish-grey flakes are deposited. This is surrounded by a clear
zone, which in turn is surrounded by a peripheral band of
involvement as well. On the border of the pupil it looks like
"dandruff". Similar material is trapped in the pores of the
trabecula meshwork and may be seen on the ciliary processes, on the
zonulas, surrounding the conjunctival vessels and in retro-orbital
tissues. It is a basement membrane material, akin to amyloid in some
respects, although many histochemical studies do not support this
identification.
The prevalence varies enormously from country to country, and
even within countries. The highest prevalence was found in the
Navajo Indians of New Mexico, in which 38% were over 60 years of
age. At the other extreme, only 2 cases have ever been recorded in
Eskimos, and these were two Greenlanders, possibly of mixed
ancestry, aged over 70 years (Ostenfeld-Åkerblom, 1988). The average
prevalence in central Europe is around 2% on the basis of figures
from several authors (Forsius, 1988).
The possibility of environmental factors was proposed by Taylor
(1979) based on observations of exfoliation in Australian
aborigines. The distribution of exfoliation was linked to annual
global radiation and to climatic droplet keratopathy.
Exfoliation syndrome sometimes occurs in other areas of high
UV, and high prevalence of climatic keratopathy. Examples include
Somalia, Djibouti and Saudi Arabia. There is, however, considerable
evidence to suggest that UV is not the main factor associated with
the development of exfoliation. The geographic distribution does not
consistently correspond with that of climatic keratopathy. There may
be wide differences in prevalence of exfoliation syndrome at similar
latitudes. For example, it is frequent in parts of East Africa, but
rare in West Africa. Similarly it may be seen at high prevalence in
the Lapps of Finland and Sweden, but not in Eskimos at the same
latitude. The prevalence may vary within the same country. A total
of 4,042 patients aged over 50 were examined in clinics in 6 areas
in different parts of France over a 2 week period. The prevalence
was high in Brittany (20.6% in those over 60 years) and extremely
rare in Picardy at a similar latitude (Colin et al., 1985).
Exfoliation is usually more frequent in females than males. It is
found in parts of the eye, such as the ciliary body and in the
orbit, remote from the influence of light. Forsius (1988) has
reviewed the evidence for genetic aetiology for the condition.
The present conclusion is that environment, at least in the
form of UV, is not the primary cause. There is not a consistent
direct relationship with solar radiation. There is at least a major
racial or genetic predisposition, but it is possible that light or
some other environmental factor activates or induces the development
of the exfoliation syndrome in those who are genetically
predisposed.
10.5.3 Anterior lens capsule
In 1989 a previously unrecorded condition was reported from
Somalia (Johnson et al., 1989). This consisted of alterations of
the pupillary area of the anterior capsule of the lens. The first
stage appeared to be an opalescence of the capsule, which then
became a plateau-shaped elevation above the surrounding contour of
the anterior lens. In its most developed form it was a bagging of
the anterior lens capsule and contents through the pupil, appearing
like a hernia. This condition was invariably associated with
climatic droplet keratopathy, but not necessarily with cataract. In
fact, there appeared to be an inverse relation with cataract.
The absolute association with climatic keratopathy suggests
that it also may be due to excessive UV exposure. Attempts to secure
histology on extracted lenses with this condition were difficult
because the lens capsule so frequently tore from the rest of the
lens as it was extracted by the cryoprobe. The capsules examined
showed thinning and splitting of the layers, and death of many of
the nuclei of the epithelial cells. However, there were no controls
from the same geographical and ethnic area of the same ages for
comparison.
10.6 Diseases of the Choroid and Retina
Among adults, only extremely small amounts of UVA and UVB at
wavelengths below 380 nm reach the retina, because of the very
strong absorption by the cornea and lens. Less than 1% of radiation
below 340 nm and 2% of radiation between 340 and 360 nm reaches the
retina (Barker and Brainard, 1993). Even in early childhood the
highest spectral transmittance reaches about 4% in the UVB and is
generally of the order of 1%. However, because of the biological
activity of the shorter wavelengths of UVB, the biological
importance of the small amount of this radiation that does reach the
retina cannot be completely neglected. As children age, UV is
increasingly absorbed by the cornea and lens, and the proportion
reaching the retina decreases. This suggests firstly, that exposure
to UV during childhood may be of more importance than exposure to UV
during adult life, and secondly, that exposure to longer wavelength
radiation (e.g. visible light) may be of more importance in
adulthood.
10.6.1 Uveal melanoma
Exposure to solar radiation is considered to be causally
associated with the development of cutaneous malignant melanoma
(IARC, 1993). There is a possibility that exposure to UV may also
cause melanoma of the uveal tract. There is no separate ICD code for
intra-ocular melanoma, so descriptive studies have generally been
based on cancer of the eye (ICD-9 190), of which it has been
estimated that 80% are intra-ocular melanomas (Osterlind, 1987). In
the case-control studies cases of uveal-tract melanomas were
confirmed histologically, but also included tumours of iris and
ciliary body with those of the choroid.
The incidence of cancer of the eye is higher among white than
black or Asian populations residing at the same latitude. For
example, in US whites the incidence rates are 0.7 per 100 000 person
years in males and 0.6 in females compared with 0.2 in both sexes in
blacks (Parkin et al., 1992). Among people of European ancestry,
risk of ocular melanoma was observed to be least in those of
southern European ethnic origin; for example, in comparison with an
RR of 1.0 in those of southern European origin, the RR in people of
northern European origin was 6.5 (95% CI 1.9-22.4; Seddon et al.,
1990). Risk of ocular melanoma was observed to be increased in those
with light eye colour, with RRs of 1.7 to 2.1 (Gallagher et al.,
1985; Tucker et al., 1985c; Holly et al., 1990), but not when
ethnicity was taken into account (Seddon et al., 1990). Kraemer
(1987) found five cases of ocular melanoma among reports of 337
patients with xeroderma pigmentosum for whom ocular findings had
been described. The defect in this condition is failure to repair
DNA after damage by UV.
There is no evident latitude gradient in incidence of ocular
melanoma in white populations of the northern hemisphere or
Australia (IARC, 1992) and, within the USA, its risks in those born
in southern parts of the country, where ambient solar radiation is
highest, has variously been reported to be more (Tucker et al.,
1985c), less (Seddon et al., 1990) or the same (Schwartz & Weiss,
1988; Mack & Floderus, 1991) as that in those born elsewhere in the
country. Similarly Gallagher et al. (1985) in Canada found no
association with latitude of residence.
Two studies have examined place of birth and risk of uveal
melanoma, but the findings are inconsistent. Tucker et al. (1985)
found an excess of cases were born south of latitude 40°N, whereas
Seddon et al. (1990) found a deficit.
Indicators of personal sun exposure have been inconsistently
associated with risk of cancer of the eye or ocular melanoma. A
small rural excess in incidence of cancer of the eye has been
reported (Doll, 1991). Two descriptive studies reported an
association with farming (Saftlas et al., 1987; Gallagher, 1988)
but this was not found in several other such studies (Milham, 1983;
Office of Population, Censuses and Surveys, 1986; Vågerö et al.,
1990) or two case-control studies of ocular melanoma (Gallagher et
al., 1985; Seddon et al., 1990). Some high exposure activities
such as gardening (RR 1.6, 95% CI 0.7-1.6) and taking sunny
vacations (RR 1.5, 95% CI 1.0-2.3, for highest category) were
significantly associated with increased risks of ocular melanoma in
one case-control study (Tucker et al., 1985c) but no similar
associations with personal sun exposure at work in leisure time, or
in vacation were found in three other studies (Gallagher et al.,
1985; Holly et al., 1990; Seddon et al., 1990). Indeed,
Gallagher et al., (1985) found an elevated risk for government
workers, a predominantly indoor managerial group. The lack of use of
protective eyewear (sunglasses, visors, headgear) was associated
with an increased risk of ocular melanoma in one study (Tucker et
al., 1985c) with an RR for infrequent or rare use of 1.6 (95% CI
1.2-2.2). Weak evidence of a similar effect was found by Seddon et
al. (1990).
No statistically significant association has been observed
between ocular melanoma and a personal history of skin cancer in
several studies of cancer registry or other data (Osterlind et al.,
1985; Tucker et al., 1985a; Holly et al., 1991; Lischko et al.,
1989; Turner et al., 1989).
There is evidence of associations between exposure to sunlamps
and some other artificial sources of UV and risk of ocular melanoma
in the three case-control studies in which they have been examined.
Tucker et al. (1985c) found a relative risk of 2.1 (95% CI
0.3-17.9) for frequent use of sunlamps compared with no use (p=0.10
for trend over four categories of use); Holly et al. (1990) found
a relative risk of 3.7 (95% CI 1.6-8.7) for ever having an exposure
to "artificial UV or black light" and with welding burn, sunburn to
eyelids, or snow-blindness, RR 7.2 (95% CI 2.5-20.6); and Seddon et
al. (1990) found a relative risk of 3.4 (95% CI 1.1-10.3) for
frequent or occasional use of sunlamps compared with never used. In
one of these studies, there was also a strong association with
employment as a welder (RR 10.9, 95% CI 2.1-56.5; Tucker et al.,
1985c). No similar association was found by Seddon et al. (1990)
but an increased risk in welders (RR 8.3, 95% CI 2.5-27.1) was found
in an occupational study of French Canadians (Siemiatycki, 1991).
Overall, the epidemiological studies do not provide convincing
evidence of an association between exposure to solar UV and uveal
melanoma. None of the studies has developed a practical assessment
of individual cumulative ocular exposure to UVB. They have all used
various simple estimates of sun-related behaviour.
On the other hand, the use of a sunlamp, an artificial source
of UV, was significantly associated with uveal melanoma in the two
case-control studies that examined their use. Another study found
elevated risk of uveal melanoma with exposure to UV or black lights.
Collectively, these studies suggest frequent use of a sunlamp may be
associated with a 2-4 fold increase in risk of developing uveal
melanoma. Sunlamp use can produce over five-fold more DNA damage per
unit of erythema than the sun (Nachtwey & Rundel, 1981).
The large number of ocular melanomas in xeroderma pigmentosum
patients also means that exposure to UV cannot be ruled out as a
causative factor.
10.6.2 Age-related macular degeneration
Age-related macular degeneration (AMD) is one of the leading
causes of blindness in the industrialized world. Visual loss can
occur because of the development of geographic atrophy (loss of the
outer retinal segments and retinal pigment epithelium), retinal
pigment epithelial detachment or sub-retinal neovascularization
(exudative AMD). Prior to visual loss AMD is characterized by the
presence of drusen (lipofuscin and other material deposited between
the retinal pigment epithelial cells and Bruch's membrane and
appearing as yellow-white nodules with distinct and indistinct edges
on retinal examination).
There is evidence for association of AMD with UV exposure.
Photochemical retinal damage can occur from prolonged exposure to
high intensity light. Whether such damage is directly related to AMD
is unknown. Although aged Rhesus monkeys have drusenoid deposits, no
good experimental animal models for AMD currently exist.
In a case control study, Hyman et al. (1983) found no
association of AMD and light exposure based on residential history.
They also found no association of AMD to occupational light
exposure.
In studies based on individual exposure data; the results are
equivocal. The initial evaluation of the association of AMD and UV
exposure in the cross-sectional study of Chesapeake Bay watermen
revealed no statistically significant association (West et al.,
1989). However, a reanalysis based on the small number of cases of
AMD with exudative disease or geographic atrophy suggested an
association with 20 year exposure to blue light but not UVA or UVB
(Taylor, 1992).
The Beaver Dam Eye Study found an association of late stage AMD
(exudative AMD or geographic atrophy) and summer leisure time
outdoors (RR=2.2 CI=1.1-4.2). It was also suggested that an
association existed in men only between early stage AMD and summer
leisure time outdoors. The magnitude of the risk estimates were
unchanged after adjusting for numerous possible confounding factors
(Cruickshanks et.al. 1993).
It can be concluded that there are very limited data
demonstrating an association of AMD with UV exposure. The finding of
an association with blue light exposure is consistent with the
wavelengths of visible light reaching the retina and needs further
investigation.
10.7 Conclusion
The causal links between UVB exposure and various ocular
conditions were evaluated on the basis of the following definitions:
Sufficient evidence for a causal association indicates that
positive associations have been observed between human exposure to
UV and the effect in which chance, bias and confounding could be
ruled out with reasonable confidence.
Limited evidence for a causal association indicates that positive
associations have been observed between exposure to UV and the
effect for which a causal interpretation is considered to be
credible, but chance, bias or confounding could not be ruled out
with reasonable confidence.
Inadequate evidence for a causal association indicates that the
available studies are of insufficient quality, consistency, or
statistical power to permit a conclusion regarding the presence or
absence of a causal association between UV and the effect, or no
data were available.
Evidence for lack of causal association indicates that there are
several adequate studies covering the range of exposure that humans
are known to encounter which are consistent in not showing a
positive association between UV and the effect.
There is sufficient evidence to link photokeratitis to acute
ocular exposure to UVB.
Sufficient evidence exists to link the production of cortical
and PSC cataracts to UVB exposure in animals. There is limited
evidence to link cortical and PSC cataract in humans to chronic
ocular exposure to UVB. Inadequate evidence is available to link PSC
cataract in humans to chronic UVB exposure. Insufficient data have
been collected upon which to evaluate the risk of cataract
associated with childhood exposure to UVB. Half the world's
35-million blind people are blind because of cataract. The
proportion of cataract that results from UVB exposure is unknown,
but may be as high as 20%.
There is limited evidence to link sunlight exposure of the eye
to the development of pterygium. It is unclear whether the observed
association is specific for UV. The contribution from other
environmental factors remains unclear.
There is limited evidence to associate climatic droplet
keratopathy with UV exposure and insufficient to link pinguecular
and cancers of the anterior ocular structures. Insufficient evidence
exists to link uveal melanoma to ocular exposure to solar UV
radiation. However, several epidemiological studies have suggested
that the use of sunbeds (an artificial source of UV) is associated
with uveal melanoma.
There is inadequate evidence of an association between ocular
UV exposure and acute solar retinitis, age-related macular
degeneration, acceleration of pigmentary retinopathies and
exfoliation syndrome.
11. EFFECTS ON PLANT AND AQUATIC ECOSYSTEMS
11.1 Introduction
Human populations may be affected by direct and indirect
consequences of increased solar UVB on aquatic food webs. Because
more than 30% of the world's animal protein for human consumption
comes from the sea (in many developing countries this percentage is
even larger), a substantial decrease in biomass production would
diminish fishery resources in the face of growing world populations.
Reductions of leaf area, fresh and dry weight, lipid content and
photosynthetic activity were typically found in UVB sensitive plant
species. Additionally, alterations of leaf surface, epicuticular
waxes, diffusion of water vapour through the stomata have been
reported. For previous comprehensive publications see Caldwell et
al. (1989), Wellmann (1991) and SCOPE/UNEP (1993).
11.2 Effects on Terrestrial Plants
11.2.1 UV penetration into the leaf
UVB has a direct effect on photosynthesis. Reductions in
photosynthesis often accompany changes in leaf pigmentation,
anatomy, and leaf thickness. After exposure to enhanced UVB, the
internal light regime of leaves was altered (Bornman & Vogelmann,
1991). In a recent study, Brassica campestris (origin: northern
latitudes) was subjected to 6.3 kJ m-2 day-1 of UVB and
responded by increasing leaf thickness by 45% and UVB screening
pigments by 21% relative to controls (Bornman & Vogelmann, 1991).
Chlorophyll content (per leaf area) and photosynthetic activity
decreased while scattered light within the leaves of UV-treated
plants increased. Since the distribution of photosynthetically
active radiation was altered at different depths within leaves after
UV, these changes can also be expected to have an indirect effect on
photosynthetic capacity.
In a study on a group of 22 diverse plant species (including
herbaceous and woody dicotyledons, grasses and conifers), widely
varying UVB penetration was found. For instance, epidermal
transmittance of the herbaceous dicotyledons ranged from 18% to 41%
with penetration up to 140 µm, while conifer needles excluded a
large percentage of the incident UVB. Penetration of UVB into leaves
of the woody dicotyledons and grasses was in between that of the
herbaceous dicotyledons and conifers (UNEP 1989).
11.2.2 Changes in growth
The growth of many plant species is reduced by enhanced levels
of UVB. The main components of plants affected by UVB are shown in
figure 11.1 (UNEP 1989). The ozone filter technique was used to
simulate a relative solar UVB enhancement of 20% by providing 54.4
kJ m-2 day-1 (unweighted) or 5.1 kJ m-2 day-1 of
biologically effective radiation (UVBBE) through one cuvette and
45.3 kJ m-2 day-1 (unweighted) or 3.6 kJ m-2 day-1 UVBBE
through the other cuvette (Tevini et al., 1991b). These were average
values measured from May 1990 to August 1990 and are equivalent to
an ozone depletion of approximately 10%. Plant height, leaf area,
and the dry weight of sunflower, corn, and rye seedlings were
significantly reduced, while oat seedling remained almost unaffected
(Tevini et al., 1991b). The reduction of hypocotyl growth of
sunflower seedlings under artificial UVB irradiation is associated
with a UV dependent destruction of the growth regulator
indole-3-acetic acid (IAA) and the formation of growth inhibiting
IAA photoproducts. The inhibition of elongation in UV-irradiated
sunflower seedlings might also be due to the action of peroxidases
working as IAA-oxidase, causing a decrease in cell wall
extensibility of the hypocotyl epidermis (Ros, 1990). Shading of
shoot apex was shown to reduce UVB induced reduction in growth of
Vigna seedlings (Kulandaivelu et al., 1993).
11.2.3 Effects on plant function
When high UVB irradiances were used in combination with low
levels of white light, such as commonly found in growth chambers,
effects on photosynthesis were generally deleterious. However, even
in the presence of higher levels of white light in green houses and
in the field, reductions in photosynthesis of up to 17% were
reported in the UVB sensitive soybean cultivar Essex when supplied
with UVB equivalent to an 18% ozone depletion (Murali & Teramura,
1987). Solar UVB also reduced net photosynthesis in sunflower
seedlings by about 15% when a 12% ozone depletion was simulated by
using the ozone filter technique (Tevini et al., 1991c). One
reason for the reduction in overall photosynthesis might be due to
stomatal closure by enhanced UVB.
The second, and most common type of study design, has been the
cross-sectional study in which a population or occupational group is
surveyed and disease prevalence measured. Cross-sectional surveys
identify all persons with the disease, some of which are new cases
while other persons may have had the disease for a period of time.
Some cross-sectional studies have related non-personal factors
associated with place of residence to disease status. Other studies
have collected detailed information from each study participant on
personal exposure to UV or indices of exposure (e.g. hours of
exposure to sunshine, occupation) and related these exposures to
disease status.
The third type of study design has been the case-control study,
where differences in UV exposure between persons with the disease
and those without have been compared. Cases have usually been drawn
from a hospital or clinic. Controls have been drawn from other
hospital or clinic patients or from the general population and have
usually only included persons with good visual acuity, without the
disease of interest, and without a disease that is associated with
UV exposure. Some case-control studies have related disease status
to non-personal factors associated with place of residence while
others have used personal exposure information.
Data from both cross-sectional and case-control studies can be
useful in confirming a hypothesis, but have a number of limitations.
If there is an excess risk of death associated with the disease, as
has been suggested for cataract (Minassian et al., 1992; Vitale
et al., 1992), both types of study will be biased towards the
survivors. In addition, disease status and prior exposure indices
are measured at the same time and it may not be possible to
differentiate between cause and effect, especially if the disease
has a long latency period.
10.4 Diseases of the External Eye
10.4.1 Photokeratitis and photoconjunctivitis
Cases of photokeratitis and photoconjunctivitis have occurred
between 0.5 and 24 hours after prolonged exposure to intense solar
radiation, often in highly reflective environments (Wittenberg,
1986). The most severe cases are usually manifested as snow
blindness, suggesting that UV is the cause of this condition.
The action spectrum for UV photokeratitis produced in the
rabbit was first measured by Cogan and Kinsey (1946). Pitts (1974,
1978) in a series of laboratory studies on humans estimated the mean
threshold of UVB (290-315 nm) for photokeratitis at 3500 J m-2.
These laboratory data are supported by Blumthaler et al. (1987),
who estimated that the radiant exposures in clinically observed
cases of photokeratitis ranged from 1200 to 5600 J m-2. It is
estimated that 100 to 200 seconds of direct, unattenuated exposure
to 295-315 nm solar radiation will result in photokeratitis (Sliney,
1987; Wittenberg, 1986). Blotting out the solar disc would remove
around 40% of the UV, still leaving a threshold of around 5.5
minutes. Sliney (1986) has estimated that the reflected levels of UV
from light sand should be sufficient to cause a threshold
photokeratitis within exposure periods of 6-8 hours centred around
midday, and within 1 hour for UV reflected from snow.
Experimental data shows photokeratitis can be induced in
animals by UVB exposure and that use of UVB absorbing contact lens
or chromophores can prevent UVB induced photokeratitis in laboratory
animals. Collectively, there is sufficient experimental and
epidemiological evidence that exposure to intense UVB radiation
causes photokeratitis and photoconjunctivitis.
10.4.2 Climatic droplet keratopathy
Climatic droplet keratopathy, among a variety of other names
(Gray et al., 1992), is also known as spheroidal degeneration from
its histological appearance. It is a degenerative condition usually
affecting both eyes symmetrically, and restricted to the exposed
interpalpebral band of the cornea. This condition is of major
significance for vision in some parts of the world, reducing vision
to blindness levels in older people. For example, in Mongolia it has
been found in an initial survey to be the third cause of blindness
(Baasanhu et al., in press).
Climatic droplet keratopathy occurs throughout the world, but
is more common in areas with snowfall persisting late into the
summer in the northern hemisphere, such as parts of northern Canada,
Siberia and Mongolia, and in areas of sand and desert in other
latitudes, including Somalia, the Arabian peninsula, Iran, and
Australia. It is also particularly common on sea coasts where there
is coral sand or the sand is impregnated with salt, such as the
islands of the Red Sea (Gray el al., 1992).
In a cross-sectional study of Australian aborigines Taylor
(1980a) found no correlation between the prevalence of climatic
droplet keratopathy and ambient UVB levels, although the condition
was more common among those working as stockmen. Johnson (1981)
reported a geographical correlation with the calculated flux of
reflected UV from snow and ice throughout the year in the eastern
coast of Newfoundland and Labrador and the eastern Arctic of Canada.
In a cross-sectional study of Chesapeake Bay waterman study
Taylor et al. (1989) examined the risk of climatic droplet
keratopathy with chronic UVB exposure. Although a positive
association was found (RR= 6.4, 95%CI=2.5-11.7) for those in the
highest quarter of exposure compared to those with the bottom
quarter, further analyses of this data (Taylor et al., 1992)
showed the risk of climatic droplet keratopathy was also related to
UVA.
There is strong evidence that the corneal degeneration is due
to environmental factors. Circumstantial evidence exists that it is
caused by solar UV, mainly reflected from ground surfaces such as
snow and sand which are particularly reflective of UV.
Histologically, the material deposited in the superficial corneal
stroma as spheroidal droplets is most likely to be derived from a
mixture of altered plasma proteins, including fibrinogen, albumin,
and immunoglobulins (Johnson & Overall, 1978).
Other proposed aetiological agents such as low atmospheric
humidity, low temperature or high temperature have been excluded. It
is possible that particulate injury by wind-blown ice or snow or
sand particles may contribute to the development of the condition by
causing inflammation and therefore outpouring of additional plasma
proteins from the blood vessels of the limbs.
10.4.3 Pinguecula
Pinguecula is a fibro-fatty degeneration of the interpalpebral
conjunctiva. The pathological changes that occur in pinguecula are
similar to actinic elastosis of the skin, a condition thought to be
linked to sunlight exposure. This indirect evidence suggests that
exposure to sunlight may be a risk factor for pinguecula.
Geographical variation in the occurrence of pinguecula has been
reported, with higher prevalence in Arabs living near the Red Sea
than in Eskimos from Greenland or Caucasians in Copenhagen (Norn,
1982). Johnson et al. (1981) in a study of pinguecula in Labrador
found the size of pinguecula was correlated with the severity of
climatic droplet keratopathy. Taylor et al. (1989) in the study of
Chesapeake Bay watermen found a weak association for the presence or
absence of pinguecula with exposure to UVA and UVB. The relative
risk for the top quartile of exposure was 1.4 (95%CI=0.9-2.2), less
than for climatic droplet keratopathy or pterygium. Karai &
Horiguchi (1984) in a study of 191 Japanese welders found no
difference in the occurrence of pinguecula between welders and
controls.
It is concluded that there is currently insufficient
epidemiological or experimental data for an assessment of the risk
of pinguecula with exposure to UV.
10.4.4 Pterygium
Pterygium is a triangular shaped degeneration and hyperplastic
process in which the bulbar conjunctiva encroaches on the cornea.
A geographical association between variation in the occurrence
of pterygium and variation in sunlight exposure was first suggested
by Talbot (1948). Based on observation of pterygium in New Zealand
and South Pacific Islands Elliott (1961) suggested pterygium in
these locations resulted from UV exposure.
Studies of non-personal factors associated with place of
residence
In a study of pterygium patients in US Veterans Administration
hospitals during 1957-59 Darrell & Bachrach (1963) related mean
daily UV (319nm) levels to the ratio of pterygium to all hospital
discharges. A trend was found between UV level and pterygium ratio
for persons born in rural counties. A similar association with UV
level was seen for persons residing in a rural county at the time of
the hospital discharge. Cameron (1965) examined the global pattern
of pterygium and reported an inverse gradient with latitude. In a
study of Australian aborigines Taylor (1980a) found pterygium was
correlated with ambient UVB level and hours of sunshine at place of
residence. Data from Canada indicates that pterygium is also common
in arctic and sub-arctic environments (Johnson et al., 1981).
Moran & Hollows (1984) found a nonsignificant increase in the
prevalence of pterygium among Australian aborigines residing in
areas with higher ambient UV levels.
Studies with personal exposure measurements
Four studies have related occurrence of pterygium to personal
measurements. Karai & Horiguchi (1984) examined 191 Japanese welders
for the presence of pterygium. A trend of increasing risk of
pterygium was found with years of employment as a welder, an
indirect measure of cumulative occupational exposure to UV.
Booth (1985) undertook a hospital-base case-control study of
pterygium in Sydney. No difference between cases and controls was
found in subjective assessment of exposure to sunlight in work or
sport. However, a family history of pterygium was found to be a
strong risk factor.
Among Chesapeake Bay watermen, Taylor et al. (1989) found a
dose-response relationship between risk of pterygium and exposure to
UVA and UVB. The relative risk for the top quarter of UVB exposure
was 3.1 (95%CI=1.8-5.3) compared to the lowest quarter. However, it
is noted that pterygium was equally associated with ocular exposure
to UVA and visible light.
Mackenzie et al. (1992) undertook a hospital-based
case-control study of pterygium in Queensland. A strong
dose-response relationship was found with closeness of place of
residence to the equator, type of outdoor work environment (e.g.
sandy) and amount of time spend outdoors. The most striking finding
was the magnitude of risk associated with spending most of the time
outdoors was stronger when related to childhood exposure (RR=17.2,
95%CI=6.2-47.6) than to adult exposure (RR=5.7, 95%CI=3.1-10.6). The
risk associated with working at ages 20-29 in an outdoors
environment of mainly sand or concrete was associated with a
relative risk of 11.3 compared with indoor workers. Corneo (1993)
has suggested that the cornea is acting as a side on lens focusing
light and also UV across the anterior chamber to the nasal limbus.
This hypothesis may explain why pterygia usually commence on the
nasal side of the eye.
Evaluation of epidemiological evidence
While several geographical studies have reported an inverse
trend with latitude, the common occurrence of pterygium in arctic
and subarctic locations suggest that closeness to the equator does
not fully explain the distribution of this disease.
The strength of the association with time spent outdoors
reported by Mackenzie et al. (1992) suggests that the association
may be causal. However, there is insufficient evidence to show that
the observed association with UV exposure is not, in part, due to
confounding. The findings from three of the studies lend support to
a hypothesis that irritation by particulate matter is associated
with pterygium. The Australian aborigines live in a dry, dusty
environment and welders are occupationally exposed to a range of
particles. Similarly, the Queensland study found highest risk among
those who worked in sandy locations. The particulate matter
hypothesis is also supported by a report (Dhir et al., 1967) of
higher prevalence of pterygium among Punjabi Indians working in
sawmills (an indoor occupation) in New Delhi and British Columbia
than Punjabi farmers (an outdoor occupation). Similar findings of
increased risk of pterygium among sawmill workers in Thailand and
Taiwan have been reported (Detels & Dhir, 1967). The evidence of
possible confounding by particulate matter is inconsistent, with the
Chesapeake Bay watermen study finding an association with sunlight
exposure in a location that was neither hot, dry or dusty.
It is not possible, based on available epidemiological data, to
assess the risk of pterygium with exposure to UV because of possible
confounding of observed associations by exposure to particulate
matter or other factors.
10.4.5 Hyperkeratosis, carcinoma-in-situ, and squamous cell
carcinoma of the conjunctiva
These conditions probably form a gradation of development and
cannot necessarily be distinguished clinically. Invasive squamous
cell carcinoma is often said to arise from a pre-cancerous lesion.
Epithelial dysplasia and carcinoma-in-situ look the same, and are
sometimes keratinized and present as leucoplakia in which case the
term actinic keratosis may be applied (Naumann & Apple, 1986;
Garner, 1989). The main argument for an actinic causation is that
these tumours usually present in the exposed area of the eye between
the lids (the interpalpebral fissure) and under conditions where
they may be expected to be exposed to solar radiation.
Xeroderma pigmentosum is a recessively inherited syndrome
characterized by clinical and cellular hypersensitivity to solar
radiation and a defect in the capacity to repair UV-induced damage
in DNA (Fitzpatrick, et al., 1963). Among reports of 337 patients
with xeroderma pigmentosum for whom ocular findings had been
described, Kraemer et al. (1987) identified 88 ocular tumours of
which 73 were specific to the corneal-scleral limbus (34), the
cornea (24) or the conjunctiva (15). Of the non- melanomas for which
histopathological type was specified, 28 were squamous cell
carcinomas and 12 were basal cell carcinomas. Among 64 patients with
ocular neoplasms whose age was stated, half the neoplasms had
occurred before 11 years of age. While the eyelids are a site of
preference for basal cell carcinoma, this tumour rarely, if ever,
arises in the conjunctiva in otherwise normal individuals.
Squamous cell carcinoma of the conjunctiva is an uncommon
tumour. Garner (1989) reviewed all cases of tumours at the limbus
sent over a 40 year period for examination to the Institute of
Ophthalmology, London. The total was only 636 tumours, of which 73
were squamous carcinomas. This amounts to less than 2 cases per year
coming to the Pathology Laboratory, even though Moorfields Eye
Hospital which the Laboratory serves, attracts patients from all
over the country and from overseas.
Lee & Hirst (1992) attempted to provide population-based
figures and estimate the incidence of these tumours in metropolitan
Brisbane (latitude 30° south). They surveyed the histological
records of all ocular surface tumours examined in the pathological
laboratories over the previous 10 years, serving a population of
more than 745,000 in 1989. There were 139 cases of which 79 were
corneal epithelial dysplasia, 28 carcinomas-in-situ and 32 were
squamous cell carcinomas. There was a strong male preponderance. The
incidence ranged from 1 per 100,000 in 1980 to 2.8 per 100,000 in
1982. This is well below the rate for squamous cell carcinoma of the
skin and melanoma of the skin in Queensland as a whole. On the other
hand, it is a substantially higher rate than that recorded in London
where the pathological laboratory referred to also covers a much
larger population.
Squamous cell carcinoma of the conjunctiva has been reported to
form a greater proportion of eye tumours in Africans living in areas
close to the equator (Templeton, 1967) than in the south of Africa
(Higginson & Oettlé, 1959), and much higher than in Baltimore
(39°N). The incidence (0.3 per 100,000) in Uganda (0°) has been
reported to be twice that in Denmark (55°N) despite the potential
underascertainment in Africa (Templeton, 1967).
It is extremely rare for a neoplasm to arise de novo in the
corneal epithelium, where it may be called a corneal intra-
epithelial neoplasm. Most such intra-epithelial sheets are connected
at the corneo-scleral limbus to a conjunctival lesion, such as a
papilloma or a leucoplakia over a pterygium or pinguecula (Waring
et al., 1984). The only available evidence for an UV aetiology is
the location of the lesion within the interpalpebral fissure, and
the fact that it may arise from a lesion which is itself associated
with UV. Three cases of corneal intra-epithelial neoplasia have been
recently reported in people aged 31 to 38 who wore contact lenses
and were considered to have had substantial exposure to artificial
and solar UV (Guex-Crosier & Herbort, 1993).
10.5 Diseases of the Lens
10.5.1 Cataract
For the purpose of this review, a cataract is defined as an
opacity of the lens of the eye. The three major types of cataract
are cortical, nuclear and posterior subcapsular (PSC). When a lens
opacity interferes with vision, a clinically significant cataract is
present. If left untreated, cataract will often progress to
blindness. Cataract causes half of the world's blindness.
Definitions of cataract and methods used to assess the presence
and severity of cataract have not been uniform in epidemiological
studies of cataract. Many studies include lens opacities that are
not necessarily accompanied by a decrease in visual acuity. Some
have combined all three major types of lens opacities into a single
"cataract" category, while others have investigated associations for
specific types of opacity. Methods of assessing the presence and
sometimes the severity of opacities range from reviews of existing
charts to clinical examinations using written definitions of
cataract, and the use of standardized grading systems that have been
found to be highly reliable.
Occupational case series
When cataracts result from occupational exposure to UV, it may
be difficult to differentiate between the contribution of
occupational and non-occupational factors to the development of the
disease. Lerman (1980) described the onset of lens opacities in
three persons who worked in a dental clinic and were exposed to UV
(300-400 nm) from a dental curing unit. The lens damage varied from
posterior subcapsular cataract in the dentist, who was reported to
have received the highest dose, to zonular type opacities in one of
the dental assistants. However, any retrospective reconstruction of
the actual ocular exposure has a large degree of uncertainty, and
the results from such an exercise must be interpreted with caution.
Studies in which UV exposure was inferred from place of residence
Selected studies of humans exposed to solar UV are presented in
tables 10.1 and 10.2. Studies were selected for inclusion in the
tables on the basis of the scientific quality of the published
report and the overall contribution of the paper to the evaluation
of the UV-cataract hypothesis. The tables do not include the
relative risks for other factors, which in some instances are higher
than for sunlight or UV exposure.
Hiller et al. (1977) investigated sunlight and cataracts
using data from the large sources (blindness registries in 14 states
and the cross-sectional Health and Nutrition Examination Survey
(HANES) of 35 geographic areas of the US) and US Weather Bureau
geographical data on annual hours of sunlight in each geographical
area. Above the age of 65 the prevalence of cataract increased with
annual hours of sunlight, with the highest prevalence found in
locations with 3000+ annual hours of sunshine. At ages 45-64 there
was some evidence of an association with hours of sunshine but the
gradient was weaker. Below age 45 there was little association
between annual hours of sunshine and prevalence of cataract. In a
further analysis of HANES data Hiller et al. (1983) reported a
correlation between average daily UVB levels and prevalence of
cataract. Analysis of HANES data by type of cataract (Hiller et
al., 1986) revealed UVB levels at location of residence were
associated with pure cortical cataract but not with pure nuclear or
posterior subcapsular cataract.
The prevalence of cataract among Australian aborigines was
found to be correlated with annual ambient UVB level at place of
residence (Taylor et al., 1980b). Hollows & Moran (1981) found the
prevalence of cataract was highest among aborigines living in the
north of Australia, an area with high average daily UVB radiation.
Mao & Hu (1982) studied age related cataract in seven rural areas of
China and found the prevalence of cataract was correlated with
annual direct solar radiation.
Residents of rural villages in Nepal had a prevalence of
cataract related in different zones of the country to average hours
of sunshine (Brilliant et al., 1983). The prevalence was higher in
the plains where there were 12 hours of direct sunshine compared to
the mountains with 7-9 hours per day. Factors such as use of glasses
and hats modify personal ocular exposure to UV and should be
assessed.
TABLE 10.1 SELECTED HUMAN CATARACT STUDIES WITH SUNLIGHT OR UV EXPOSURE BASED ON RESIDENCE
AUTHOR POPULATION MEASURE OF MEASURE OF ASSOCIATIONS OBSERVED COMMENTS
OUTCOME SUNLIGHT EXPOSURE
Hiller et al. MRA: 9110 persons Blind from cataract, Average hours of RR= 3.3 (age 65-74) Adjusted for age and
(1977, 1983, registered as blind visual acuity (VA) sunlight < 2400 vs sex; blind registry data
1986) in 14 US States; 6/60 3000+
whites;
NHANES: 3580 persons; Lens opacity and VA Average hours of RR= 2.7 (age 65.74) Adjusted for age and sex
probability sample of < 6/7.5 sunlight < 2400 vs
US population; 3000+
NHANES data Lens opacity and VA Average daily UVB RR= 1.58 (age 45-74, Adjusted analysis
< 6/9 count in area of p<.05)
residency 6000 vs 2600
NHANES data Nuclear and cortical Average daily UVB RR= 3.6 for cortical Adjusted analyses; pure
opacity count in area of opacity; no association opacity types only
residency 6000 vs 2600 with nuclear opacity
Taylor (1980b) Survey of 350 Lens opacity with Average daily sunlight RR= 4.2 (95% CI=0.9-18.91 Unadjusted for age or
Australian Aborigines good vision, poor hours in area of potential confounding
vision or blindness residence: 9.5+ vs < 8 factors
Annual mean UVB RR= 1.8 (95%ci=0.9-3.4)1
radiation level for
area of residence: 1 95%CI estimated from
3000 vs 2000 published data
Hollows & Moran Survey of 64,307 Lens opacity and VA Average daily UVB Significant positive Wide age bands;
(1981) Aborigines and 41,254 < 6/6 count in 5 zones of correlation between unadjusted analyses
non-Aborigines, Australia: 3000 vs prevalence of lens
Australia 1000 opacity and UVB counts
in Aborigines; no
association in
non-Aborigines
TABLE 10.1 (contd).
AUTHOR POPULATION MEASURE OF MEASURE OF ASSOCIATIONS OBSERVED COMMENTS
OUTCOME SUNLIGHT EXPOSURE
Brilliant et Survey of 27,785 Lens opacities or Average daily sunlight RR= 3.8 (p<.001) Adjusted for age and
al. (1983) Nepalese; national aphakia hours: 12 vs 712 vs sex; RR decreased with
probability sample; 7-9 increasing altitude; sun
lifelong residents; blocked by mountains at
high elevations
12 vs 712 vs 7-9 RR= 2.6 (p<.005)
Cruickshanks et Cross-sectional Nuclear, cortical Average annual ambient UVB exposure associated Adjusted for other risk
al. (1993) survey of 4926 and PSC opacities UVB exposure with cortical opacities factors; measure of
persons, Wisconsin, in men (RR=1.36, exposure represents
USA 95%CI=1.02-1.79) but not average potential
women, not associated exposure at residency;
with nuclear or PSC also Table 9.2
opacities
TABLE 10.2 SELECTED HUMAN CATARACT STUDIES WITH INDIVIDUAL UV EXPOSURE ASSESSMENT
AUTHOR POPULATION MEASURE OF MEASURE OF SUNLIGHT ASSOCIATIONS OBSERVED COMMENTS
OUTCOME EXPOSURE
Collmann et Clinic-based case Nuclear, cortical Average annual sunlight No significant association with Low power to
al. (1988) control study of 113 and PSC opacities exposure, based on any type of opacity detect
cases and 168 residential history and association;
controls, North amount of time spent in sun matched on age
Carolina, USA; whites and sex
Taylor et Cross-sectional Nuclear and Cumulative ocular exposure Dose-response relationship in High exposure
al. (1988) survey of 838 cortical to UV since age 16, based which a doubling of cumulative UVR study population;
watermen, Maryland, opacities on life history and ocular exposure increased risk of detailed ocular
USA exposure model cortical opacity by 1.60 exposure model
(95%CI=1.01-2.64). RR= 3.30
(95%CI=0.90-9.97) for highest vs
lowest quartile. No association
between ocular exposure to UVR and
nuclear opacity
Bochow et Clinic-based PSC cataract Cumulative ocular exposure Increased levels of UVB exposure Adjusted analyses;
al. (1989) case-control study of (surgical since age 16, based on life associated with increased risk of association
168 cases and 168 patients) history and exposure model PSC cataract present when
controls, Maryland, adjusted for
USA cortical cataract
Dolezal et Clinic-based Cataract Lifetime sunlight exposure, No association between lifetime Only partially
al. (1989) case-control study of (scheduled for based on life history, sunlight exposure and risk of adjusted for
160 cases and 160 surgery) amount of time in sun and cataract; use of head covering potential
controls, Iowa, USA use of glasses and hat reduced risk of cataract in males confounding
(RR=0.48, 95%CI=0.25-0.94) factors; crude
index of exposure,
low power
TABLE 10.2 (contd).
AUTHOR POPULATION MEASURE OF MEASURE OF SUNLIGHT ASSOCIATIONS OBSERVED COMMENTS
OUTCOME EXPOSURE
Italian-Am. Clinical-based Nuclear, Work location in the Cortical and mixed opacities Adjusted for other
study (1991) case-control study of cortical, PSC and sunlight; leisure time in associated with work location in risk factors;
1008 cases and 469 mixed opacities the sunlight; use of sunlight (RR=1.75, crude indices of
controls, Italy glasses and hat 95%CI=1.15-2.65), leisure time in exposure; also
sunlight (RR=1.45, Table 9.1
95%CI=1.09-1.93). Cortical, PSC
and mixed opacities associated
with use of a hat in summer
(RR=1.80, 95%CI=1.17-2.47). No
association between sun exposure
indices and nuclear opacities
Leske et al. Clinical-based Nuclear, cortical Work in sunlight; leisure Work in sunlight significantly Analyses adjusted
(1991) case-control study of and PSC mixed time in sun; residence and reduced risk of nuclear opacity for other risk
945 cases and 435 opacities travel to areas of high sun (RR=0.61, 95%CI=0.37-0.99); no factor
controls, exposure, use of hat and significant associations between
Massachusetts, USA sunglasses exposure and cortical or PSC
opacities
Cruikshanks Cross-sectional Nuclear, cortical Leisure and work time No associations with cortical Adjusted for other
et al. survey of 4926 and PSC opacities outside; use of glasses and opacities; reduced risk of nuclear factors; crude
(1993) persons, Wisconsin, hat and PSC opacities amount men for indices of
USA outdoor leisure time in winter; exposure; also
use of hats and sunglasses Table 9.1
significantly increased risk of
PSC opacity in women
Age and sex adjusted prevalence for all types of cataract in
persons aged 40 years and older was found to be 60% greater in Tibet
than in Beijing (14.6% versus 9.1%, p>0.001) (Hu et al., 1989).
The authors suggested a relationship with higher UV at the higher
altitudes of Tibet, but confounding factors could not be excluded
and prevalences were higher in women than men.
Studies with personal exposure measurements
A number of studies have collected information from each study
participant and estimated personal exposure to either sunlight or
UV. Factors such as use of glasses and hats should be assessed. The
characteristics of selected studies are outlined in Table 10.2.
In a cross-sectional study of cataract in the Punjab related
prevalence of cataract to work environment Chatterjee et al.
(1982) found a suggestion of lower cataract incidence among men
whose main work location was outdoors (RR=0.7, 95%CI=0.5-1.1).
Collman et al. (1988) examined lifetime exposure to sunlight
in a clinic-based case-control study of cortical, nuclear or PSC.
Lifetime exposure to sunlight was estimated from intensity of solar
radiation in area of residence, years of residence and average
amount of time spent outdoors during daylight hours. A
non-significant risk (RR=1.1) of cataract was found for the highest
category of lifetime exposure to sunlight.
Personal exposure history and ambient UVB data were combined to
estimate an individual's lifetime annual ocular exposure to UVB
after age 15 in a cross-sectional study of Chesapeake Bay watermen
(Taylor et al., 1988). This included information on occupational
and leisure exposures, type of work surfaces, seasons, and use of
head wear and eyewear. A moderate association with a trend of
increasing risk with exposure to UVB was seen for cortical cataract,
with a RR of 3.3 (95%CI=0.9-10.0) for the top quarter of exposure
relative to the bottom quarter. A nonsignificant association was
also found between exposure to UVA (320 -340 nm) and prevalence of
cortical cataract. Little evidence was found for an association
between UVA or UVB exposure and nuclear cataract. It is noted that
UVA and UVB exposures were highly correlated and that the study
would not have been able to differentiate between the effects of UVA
and UVB. Further analyses suggested a significant difference between
the cumulative lifetime ocular exposure among cases of cortical
cataract compared to non- cataract controls. No threshold or latency
period was observed.
In a clinic-based study of PSC cataracts in Maryland, cases
were persons who underwent PSC extraction in an ophthalmic practice
(Bochow et al., 1989). Controls, matched on age, sex, and type of
referral were chosen from other patients on the appointment book of
the same ophthalmic practice who did not have a PSC cataract or a
previous cataract extraction. Annual and cumulative ocular UVB
exposures were estimated for each individual using the same method
as the studies of Chesapeake Bay watermen. Thirty-nine percent of
cases had a pure PSC cataract, the remaining 61% had mixed PSC and
other cataracts. Almost half the controls had a non PSC cataract
(nuclear, cortical or other lens opacity). UVB exposure was
significantly associated with PSC cataracts. Both the average
cumulative exposure and average annual exposure were higher in cases
than controls, after adjusting for steroid use, eye colour,
education, diabetes and presence of cortical cataracts.
In a hospital-based study of cataract patients in Iowa Dolezal
et al. (1989) found little evidence of an association between
individual lifetime sunlight exposure and cataract. Mohan et al.
(1989) in a similar study of cataract in New Delhi examined a range
of environmental factors, including occupation. An increase in cloud
cover was significantly associated (RR=0.8, 95%CI=0.7-0.9) with
cataract when adjusted for each of the other environmental
variables. The study did not quantify individual lifetime exposure
to sunlight or UV.
In a study of cataract patients in a Massachussetts hospital,
Leske et al. (1991) investigated occupational exposure to
sunshine. No association was found for PSC cataract (RR=1.3,
95%CI=0.7-2.3), cortical cataract (RR=0.9, 95%CI=0.6-1.3), or mixed
cataract (RR=0.8, 95%CI=0.6-1.1) among those with at least 2 hours
of exposure to bright sunshine per day for at least 2 months. The
risk of nuclear cataract was reduced (RR=0.5, 95%CI=0.3-0.9).
A hospital-based study from Italy (Italian-American Cataract
Study Group, 1991) found an excess of pure cortical and mixed
cataract (RR=1.8, 95%CI=1.2-2.6) and a nonsignificant deficit of
nuclear (RR=0.6) and PSC cataract (RR=0.8) among those with a work
location in the sunlight. Leisure time spent in the sunlight was
associated with an excess of cortical and mixed cataract (RR=1.4,
95%CI=1.1-1.9) and a nonsignificant deficit of posterior subcapsular
cataract (RR=0.6).
In an Indian clinic-based study of cataract and history of
severe diarrhoeal diseases Bhatnagar et al. (1991) found an
elevated risk of cataract (RR=2.1, 95%CI=1.2-3.6) for outdoor
occupations compare with indoor occupations. However Zaunuddin &
Saski (1991) found no relationship between hours of exposure to
sunshine and prevalence of nuclear or cortical cataract in Sumatra
(0° latitude). In Beaver Dam, Wisconsin, Cruickshanks et al.
(1993) found no association between average annual exposure to UVB
and cortical, PSC or nuclear cataract. Wong et al. (1993) surveyed
fishermen in Hong Kong. A sun exposure score was calculated based on
daily sunlight exposure, and protection from use of a canopy, hat,
and glasses. The highest grades of cataract of all types considered
together were more common in subjects with the highest sun exposure
scores, but none of these associations was significant at the 5%
level. A population-based case-control study (Shibati et al.,
1993) reported an increased risk of cortical cataract among men aged
40-50 years who spent 5 or more hours per day outdoors compared with
those who spent less time outdoors (RR = 6.89; 95%CI = 1.22-39).
Evaluation of epidemiological evidence
An association has been demonstrated between prevalence of
cataract and residence in areas at low latitudes, with long hours of
sunlight or high ambient UV radiance in several studies undertaken
in different parts of the world. However, in each study, the
observed association may be confounded by other possibly causal
factors. Certain of the earlier studies did not classify the lens
opacities into types of cataract.
Cortical cataract was examined separately in four studies. Only
one study assessed individual exposure. Taylor et al. (1988) found
a dose-response relationship with exposure to UVB radiation. The
relative risk for the highest exposure category was three times that
for the lowest exposure category. It is unlikely that the exposure
assessment was able to distinguish between UVA and UVB exposure. The
other two used simple measures of sun-related behaviour. Leske et
al. (1991) found no association between exposure to bright
sunshine and cortical cataract, while in the Italian-American
Cataract Study (1991), a work location in the sunlight was related
to cortical and mixed cataract. The Italian study also found an
association between leisure time outdoors and cortical and mixed
cataract. The other two studies showed non-significant trends in
opposite directions. More recently, Cruickshanks et al. (1993)
found annual UVB exposure was associated with cortical opacities
among men, but no association was found for women.
Four studies report risk estimates for posterior subcapsular
(PSC) cataracts. Bochow et al. (1989) measured individual exposure
and found PSC cataract patients had higher annual and cumulative
exposures to UVB than controls, even after allowing for the effects
of several other factors. The other two studies used simple measures
of sun-related behaviour and showed non-significant trends in
opposite directions. Leske et al. (1991) found elevated risk for
pure PSC cataract patients compared to controls. However, the
Italian-American Cataract Study (1991) reported reduced risk for
pure PSC cataract patients with a work location in the sunlight or
who spent leisure time in the sunlight. Cruickshanks et al. (1993)
found no association between annual UVB exposure and risk of PSC
cataract.
Five studies provide risk estimates separately for nuclear
cataracts. These studies are consistent in showing no association
between UV exposure and nuclear cataract (Taylor et al., 1988;
Dolezal et al., 1989; Leske et al., 1991; The Italian-American
Cataract Study, 1991; and Cruickshanks et al., 1993). Collectively
these studies are consistent in showing no association between UV
exposure and nuclear cataract.
All of the published epidemiological studies of UV and cataract
have been challenged by the enormous difficulty of determining
ocular exposure in different climates. As noted previously, the
cornea and lens are seldom directly exposed to light rays from much
of the sky; hence the sunlight scattered from the ground and the
horizon determine the actual accumulated UV dose.
These studies clearly demonstrate that UV is at least one
aetiologic factor in cataractogenesis. However, extrapolation of
strong associations found in a mid-latitude population where no
serious nutritional problems are present (e.g. Taylor, et al.,
1988) to a tropical population in less developed regions, may not be
valid, since the contribution of UV relative to other factors such
as malnutrition and dehydration may be far more important.
10.5.2 Exfoliation syndrome
The exfoliation syndrome (pseudoexfoliation of lens capsule)
consists of abnormal material deposited on or arising from various
parts of the anterior eye. This condition was originally described
from Finland by Lindberg (1917). In the Nordic countries it
contributes to a high proportion of glaucoma in the older
population. This appears as a round area in the centre of the
anterior lens capsule, corresponding to normal pupil size, on which
bluish-grey flakes are deposited. This is surrounded by a clear
zone, which in turn is surrounded by a peripheral band of
involvement as well. On the border of the pupil it looks like
"dandruff". Similar material is trapped in the pores of the
trabecula meshwork and may be seen on the ciliary processes, on the
zonulas, surrounding the conjunctival vessels and in retro-orbital
tissues. It is a basement membrane material, akin to amyloid in some
respects, although many histochemical studies do not support this
identification.
The prevalence varies enormously from country to country, and
even within countries. The highest prevalence was found in the
Navajo Indians of New Mexico, in which 38% were over 60 years of
age. At the other extreme, only 2 cases have ever been recorded in
Eskimos, and these were two Greenlanders, possibly of mixed
ancestry, aged over 70 years (Ostenfeld-Åkerblom, 1988). The average
prevalence in central Europe is around 2% on the basis of figures
from several authors (Forsius, 1988).
The possibility of environmental factors was proposed by Taylor
(1979) based on observations of exfoliation in Australian
aborigines. The distribution of exfoliation was linked to annual
global radiation and to climatic droplet keratopathy.
Exfoliation syndrome sometimes occurs in other areas of high
UV, and high prevalence of climatic keratopathy. Examples include
Somalia, Djibouti and Saudi Arabia. There is, however, considerable
evidence to suggest that UV is not the main factor associated with
the development of exfoliation. The geographic distribution does not
consistently correspond with that of climatic keratopathy. There may
be wide differences in prevalence of exfoliation syndrome at similar
latitudes. For example, it is frequent in parts of East Africa, but
rare in West Africa. Similarly it may be seen at high prevalence in
the Lapps of Finland and Sweden, but not in Eskimos at the same
latitude. The prevalence may vary within the same country. A total
of 4,042 patients aged over 50 were examined in clinics in 6 areas
in different parts of France over a 2 week period. The prevalence
was high in Brittany (20.6% in those over 60 years) and extremely
rare in Picardy at a similar latitude (Colin et al., 1985).
Exfoliation is usually more frequent in females than males. It is
found in parts of the eye, such as the ciliary body and in the
orbit, remote from the influence of light. Forsius (1988) has
reviewed the evidence for genetic aetiology for the condition.
The present conclusion is that environment, at least in the
form of UV, is not the primary cause. There is not a consistent
direct relationship with solar radiation. There is at least a major
racial or genetic predisposition, but it is possible that light or
some other environmental factor activates or induces the development
of the exfoliation syndrome in those who are genetically
predisposed.
10.5.3 Anterior lens capsule
In 1989 a previously unrecorded condition was reported from
Somalia (Johnson et al., 1989). This consisted of alterations of
the pupillary area of the anterior capsule of the lens. The first
stage appeared to be an opalescence of the capsule, which then
became a plateau-shaped elevation above the surrounding contour of
the anterior lens. In its most developed form it was a bagging of
the anterior lens capsule and contents through the pupil, appearing
like a hernia. This condition was invariably associated with
climatic droplet keratopathy, but not necessarily with cataract. In
fact, there appeared to be an inverse relation with cataract.
The absolute association with climatic keratopathy suggests
that it also may be due to excessive UV exposure. Attempts to secure
histology on extracted lenses with this condition were difficult
because the lens capsule so frequently tore from the rest of the
lens as it was extracted by the cryoprobe. The capsules examined
showed thinning and splitting of the layers, and death of many of
the nuclei of the epithelial cells. However, there were no controls
from the same geographical and ethnic area of the same ages for
comparison.
10.6 Diseases of the Choroid and Retina
Among adults, only extremely small amounts of UVA and UVB at
wavelengths below 380 nm reach the retina, because of the very
strong absorption by the cornea and lens. Less than 1% of radiation
below 340 nm and 2% of radiation between 340 and 360 nm reaches the
retina (Barker and Brainard, 1993). Even in early childhood the
highest spectral transmittance reaches about 4% in the UVB and is
generally of the order of 1%. However, because of the biological
activity of the shorter wavelengths of UVB, the biological
importance of the small amount of this radiation that does reach the
retina cannot be completely neglected. As children age, UV is
increasingly absorbed by the cornea and lens, and the proportion
reaching the retina decreases. This suggests firstly, that exposure
to UV during childhood may be of more importance than exposure to UV
during adult life, and secondly, that exposure to longer wavelength
radiation (e.g. visible light) may be of more importance in
adulthood.
10.6.1 Uveal melanoma
Exposure to solar radiation is considered to be causally
associated with the development of cutaneous malignant melanoma
(IARC, 1993). There is a possibility that exposure to UV may also
cause melanoma of the uveal tract. There is no separate ICD code for
intra-ocular melanoma, so descriptive studies have generally been
based on cancer of the eye (ICD-9 190), of which it has been
estimated that 80% are intra-ocular melanomas (Osterlind, 1987). In
the case-control studies cases of uveal-tract melanomas were
confirmed histologically, but also included tumours of iris and
ciliary body with those of the choroid.
The incidence of cancer of the eye is higher among white than
black or Asian populations residing at the same latitude. For
example, in US whites the incidence rates are 0.7 per 100 000 person
years in males and 0.6 in females compared with 0.2 in both sexes in
blacks (Parkin et al., 1992). Among people of European ancestry,
risk of ocular melanoma was observed to be least in those of
southern European ethnic origin; for example, in comparison with an
RR of 1.0 in those of southern European origin, the RR in people of
northern European origin was 6.5 (95% CI 1.9-22.4; Seddon et al.,
1990). Risk of ocular melanoma was observed to be increased in those
with light eye colour, with RRs of 1.7 to 2.1 (Gallagher et al.,
1985; Tucker et al., 1985c; Holly et al., 1990), but not when
ethnicity was taken into account (Seddon et al., 1990). Kraemer
(1987) found five cases of ocular melanoma among reports of 337
patients with xeroderma pigmentosum for whom ocular findings had
been described. The defect in this condition is failure to repair
DNA after damage by UV.
There is no evident latitude gradient in incidence of ocular
melanoma in white populations of the northern hemisphere or
Australia (IARC, 1992) and, within the USA, its risks in those born
in southern parts of the country, where ambient solar radiation is
highest, has variously been reported to be more (Tucker et al.,
1985c), less (Seddon et al., 1990) or the same (Schwartz & Weiss,
1988; Mack & Floderus, 1991) as that in those born elsewhere in the
country. Similarly Gallagher et al. (1985) in Canada found no
association with latitude of residence.
Two studies have examined place of birth and risk of uveal
melanoma, but the findings are inconsistent. Tucker et al. (1985)
found an excess of cases were born south of latitude 40°N, whereas
Seddon et al. (1990) found a deficit.
Indicators of personal sun exposure have been inconsistently
associated with risk of cancer of the eye or ocular melanoma. A
small rural excess in incidence of cancer of the eye has been
reported (Doll, 1991). Two descriptive studies reported an
association with farming (Saftlas et al., 1987; Gallagher, 1988)
but this was not found in several other such studies (Milham, 1983;
Office of Population, Censuses and Surveys, 1986; Vågerö et al.,
1990) or two case-control studies of ocular melanoma (Gallagher et
al., 1985; Seddon et al., 1990). Some high exposure activities
such as gardening (RR 1.6, 95% CI 0.7-1.6) and taking sunny
vacations (RR 1.5, 95% CI 1.0-2.3, for highest category) were
significantly associated with increased risks of ocular melanoma in
one case-control study (Tucker et al., 1985c) but no similar
associations with personal sun exposure at work in leisure time, or
in vacation were found in three other studies (Gallagher et al.,
1985; Holly et al., 1990; Seddon et al., 1990). Indeed,
Gallagher et al., (1985) found an elevated risk for government
workers, a predominantly indoor managerial group. The lack of use of
protective eyewear (sunglasses, visors, headgear) was associated
with an increased risk of ocular melanoma in one study (Tucker et
al., 1985c) with an RR for infrequent or rare use of 1.6 (95% CI
1.2-2.2). Weak evidence of a similar effect was found by Seddon et
al. (1990).
No statistically significant association has been observed
between ocular melanoma and a personal history of skin cancer in
several studies of cancer registry or other data (Osterlind et al.,
1985; Tucker et al., 1985a; Holly et al., 1991; Lischko et al.,
1989; Turner et al., 1989).
There is evidence of associations between exposure to sunlamps
and some other artificial sources of UV and risk of ocular melanoma
in the three case-control studies in which they have been examined.
Tucker et al. (1985c) found a relative risk of 2.1 (95% CI
0.3-17.9) for frequent use of sunlamps compared with no use (p=0.10
for trend over four categories of use); Holly et al. (1990) found
a relative risk of 3.7 (95% CI 1.6-8.7) for ever having an exposure
to "artificial UV or black light" and with welding burn, sunburn to
eyelids, or snow-blindness, RR 7.2 (95% CI 2.5-20.6); and Seddon et
al. (1990) found a relative risk of 3.4 (95% CI 1.1-10.3) for
frequent or occasional use of sunlamps compared with never used. In
one of these studies, there was also a strong association with
employment as a welder (RR 10.9, 95% CI 2.1-56.5; Tucker et al.,
1985c). No similar association was found by Seddon et al. (1990)
but an increased risk in welders (RR 8.3, 95% CI 2.5-27.1) was found
in an occupational study of French Canadians (Siemiatycki, 1991).
Overall, the epidemiological studies do not provide convincing
evidence of an association between exposure to solar UV and uveal
melanoma. None of the studies has developed a practical assessment
of individual cumulative ocular exposure to UVB. They have all used
various simple estimates of sun-related behaviour.
On the other hand, the use of a sunlamp, an artificial source
of UV, was significantly associated with uveal melanoma in the two
case-control studies that examined their use. Another study found
elevated risk of uveal melanoma with exposure to UV or black lights.
Collectively, these studies suggest frequent use of a sunlamp may be
associated with a 2-4 fold increase in risk of developing uveal
melanoma. Sunlamp use can produce over five-fold more DNA damage per
unit of erythema than the sun (Nachtwey & Rundel, 1981).
The large number of ocular melanomas in xeroderma pigmentosum
patients also means that exposure to UV cannot be ruled out as a
causative factor.
10.6.2 Age-related macular degeneration
Age-related macular degeneration (AMD) is one of the leading
causes of blindness in the industrialized world. Visual loss can
occur because of the development of geographic atrophy (loss of the
outer retinal segments and retinal pigment epithelium), retinal
pigment epithelial detachment or sub-retinal neovascularization
(exudative AMD). Prior to visual loss AMD is characterized by the
presence of drusen (lipofuscin and other material deposited between
the retinal pigment epithelial cells and Bruch's membrane and
appearing as yellow-white nodules with distinct and indistinct edges
on retinal examination).
There is evidence for association of AMD with UV exposure.
Photochemical retinal damage can occur from prolonged exposure to
high intensity light. Whether such damage is directly related to AMD
is unknown. Although aged Rhesus monkeys have drusenoid deposits, no
good experimental animal models for AMD currently exist.
In a case control study, Hyman et al. (1983) found no
association of AMD and light exposure based on residential history.
They also found no association of AMD to occupational light
exposure.
In studies based on individual exposure data; the results are
equivocal. The initial evaluation of the association of AMD and UV
exposure in the cross-sectional study of Chesapeake Bay watermen
revealed no statistically significant association (West et al.,
1989). However, a reanalysis based on the small number of cases of
AMD with exudative disease or geographic atrophy suggested an
association with 20 year exposure to blue light but not UVA or UVB
(Taylor, 1992).
The Beaver Dam Eye Study found an association of late stage AMD
(exudative AMD or geographic atrophy) and summer leisure time
outdoors (RR=2.2 CI=1.1-4.2). It was also suggested that an
association existed in men only between early stage AMD and summer
leisure time outdoors. The magnitude of the risk estimates were
unchanged after adjusting for numerous possible confounding factors
(Cruickshanks et.al. 1993).
It can be concluded that there are very limited data
demonstrating an association of AMD with UV exposure. The finding of
an association with blue light exposure is consistent with the
wavelengths of visible light reaching the retina and needs further
investigation.
10.7 Conclusion
The causal links between UVB exposure and various ocular
conditions were evaluated on the basis of the following definitions:
Sufficient evidence for a causal association indicates that
positive associations have been observed between human exposure to
UV and the effect in which chance, bias and confounding could be
ruled out with reasonable confidence.
Limited evidence for a causal association indicates that positive
associations have been observed between exposure to UV and the
effect for which a causal interpretation is considered to be
credible, but chance, bias or confounding could not be ruled out
with reasonable confidence.
Inadequate evidence for a causal association indicates that the
available studies are of insufficient quality, consistency, or
statistical power to permit a conclusion regarding the presence or
absence of a causal association between UV and the effect, or no
data were available.
Evidence for lack of causal association indicates that there are
several adequate studies covering the range of exposure that humans
are known to encounter which are consistent in not showing a
positive association between UV and the effect.
There is sufficient evidence to link photokeratitis to acute
ocular exposure to UVB.
Sufficient evidence exists to link the production of cortical
and PSC cataracts to UVB exposure in animals. There is limited
evidence to link cortical and PSC cataract in humans to chronic
ocular exposure to UVB. Inadequate evidence is available to link PSC
cataract in humans to chronic UVB exposure. Insufficient data have
been collected upon which to evaluate the risk of cataract
associated with childhood exposure to UVB. Half the world's
35-million blind people are blind because of cataract. The
proportion of cataract that results from UVB exposure is unknown,
but may be as high as 20%.
There is limited evidence to link sunlight exposure of the eye
to the development of pterygium. It is unclear whether the observed
association is specific for UV. The contribution from other
environmental factors remains unclear.
There is limited evidence to associate climatic droplet
keratopathy with UV exposure and insufficient to link pinguecular
and cancers of the anterior ocular structures. Insufficient evidence
exists to link uveal melanoma to ocular exposure to solar UV
radiation. However, several epidemiological studies have suggested
that the use of sunbeds (an artificial source of UV) is associated
with uveal melanoma.
There is inadequate evidence of an association between ocular
UV exposure and acute solar retinitis, age-related macular
degeneration, acceleration of pigmentary retinopathies and
exfoliation syndrome.
11. EFFECTS ON PLANT AND AQUATIC ECOSYSTEMS
11.1 Introduction
Human populations may be affected by direct and indirect
consequences of increased solar UVB on aquatic food webs. Because
more than 30% of the world's animal protein for human consumption
comes from the sea (in many developing countries this percentage is
even larger), a substantial decrease in biomass production would
diminish fishery resources in the face of growing world populations.
Reductions of leaf area, fresh and dry weight, lipid content and
photosynthetic activity were typically found in UVB sensitive plant
species. Additionally, alterations of leaf surface, epicuticular
waxes, diffusion of water vapour through the stomata have been
reported. For previous comprehensive publications see Caldwell et
al. (1989), Wellmann (1991) and SCOPE/UNEP (1993).
11.2 Effects on Terrestrial Plants
11.2.1 UV penetration into the leaf
UVB has a direct effect on photosynthesis. Reductions in
photosynthesis often accompany changes in leaf pigmentation,
anatomy, and leaf thickness. After exposure to enhanced UVB, the
internal light regime of leaves was altered (Bornman & Vogelmann,
1991). In a recent study, Brassica campestris (origin: northern
latitudes) was subjected to 6.3 kJ m-2 day-1 of UVB and
responded by increasing leaf thickness by 45% and UVB screening
pigments by 21% relative to controls (Bornman & Vogelmann, 1991).
Chlorophyll content (per leaf area) and photosynthetic activity
decreased while scattered light within the leaves of UV-treated
plants increased. Since the distribution of photosynthetically
active radiation was altered at different depths within leaves after
UV, these changes can also be expected to have an indirect effect on
photosynthetic capacity.
In a study on a group of 22 diverse plant species (including
herbaceous and woody dicotyledons, grasses and conifers), widely
varying UVB penetration was found. For instance, epidermal
transmittance of the herbaceous dicotyledons ranged from 18% to 41%
with penetration up to 140 µm, while conifer needles excluded a
large percentage of the incident UVB. Penetration of UVB into leaves
of the woody dicotyledons and grasses was in between that of the
herbaceous dicotyledons and conifers (UNEP 1989).
11.2.2 Changes in growth
The growth of many plant species is reduced by enhanced levels
of UVB. The main components of plants affected by UVB are shown in
figure 11.1 (UNEP 1989). The ozone filter technique was used to
simulate a relative solar UVB enhancement of 20% by providing 54.4
kJ m-2 day-1 (unweighted) or 5.1 kJ m-2 day-1 of
biologically effective radiation (UVBBE) through one cuvette and
45.3 kJ m-2 day-1 (unweighted) or 3.6 kJ m-2 day-1 UVBBE
through the other cuvette (Tevini et al., 1991b). These were average
values measured from May 1990 to August 1990 and are equivalent to
an ozone depletion of approximately 10%. Plant height, leaf area,
and the dry weight of sunflower, corn, and rye seedlings were
significantly reduced, while oat seedling remained almost unaffected
(Tevini et al., 1991b). The reduction of hypocotyl growth of
sunflower seedlings under artificial UVB irradiation is associated
with a UV dependent destruction of the growth regulator
indole-3-acetic acid (IAA) and the formation of growth inhibiting
IAA photoproducts. The inhibition of elongation in UV-irradiated
sunflower seedlings might also be due to the action of peroxidases
working as IAA-oxidase, causing a decrease in cell wall
extensibility of the hypocotyl epidermis (Ros, 1990). Shading of
shoot apex was shown to reduce UVB induced reduction in growth of
Vigna seedlings (Kulandaivelu et al., 1993).
11.2.3 Effects on plant function
When high UVB irradiances were used in combination with low
levels of white light, such as commonly found in growth chambers,
effects on photosynthesis were generally deleterious. However, even
in the presence of higher levels of white light in green houses and
in the field, reductions in photosynthesis of up to 17% were
reported in the UVB sensitive soybean cultivar Essex when supplied
with UVB equivalent to an 18% ozone depletion (Murali & Teramura,
1987). Solar UVB also reduced net photosynthesis in sunflower
seedlings by about 15% when a 12% ozone depletion was simulated by
using the ozone filter technique (Tevini et al., 1991c). One
reason for the reduction in overall photosynthesis might be due to
stomatal closure by enhanced UVB.
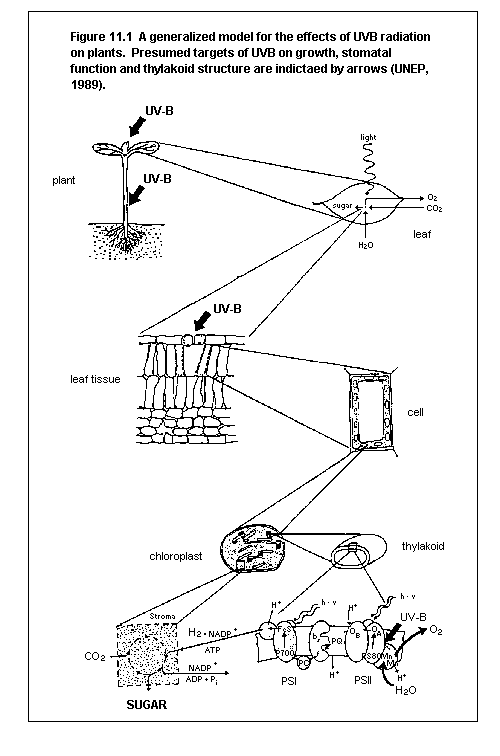 Recent studies reveal the effects of UVB radiation on tropical
plants. Rice is among the most important tropical crops in the
world. Sixteen rice ( Oryza sativa L.) cultivars from several
different geographical regions when grown for 12 weeks in
greenhouses with supplemental levels of UVB exposure equivalent to
20% ozone depletion over the equator (15.7 kJ m-2 day-1 UVBBE)
showed alterations in biomass, morphology, and photosynthesis.
Approximately one-third of all cultivars tested showed a
statistically significant decrease in total biomass with increased
UVB exposure. Photosynthetic capacity declined for some cultivars,
but only a weak relationship existed between changes in
photosynthesis and biomass with increasing UVB exposure. In one of
the rice cultivars tested, total biomass significantly increased by
20% when grown under enhanced levels of UVB exposure. Therefore,
despite the fact that the effects of UVB are generally damaging, in
some cases, it has been reported to have a stimulating effect. Such
positive growth effects are presently not easily explainable.
Results from this experiment indicate that 1) a number of rice
cultivars are sensitive to increases in UVB exposure; 2) the
diversity exhibited by rice in response to increased levels of UVB
suggests that selective breeding might be successfully used to
develop UVB tolerant rice cultivars. Other preliminary screening
studies on rice seedlings also corroborate these observations
(Coronel et al., 1990).
In a three year field study (Sullivan & Teramura, 1991),
photosynthetic capacity was generally reduced in loblolly pine trees
exposed to supplemental levels of UVB simulating a 16% and 25% ozone
depletion (11.5 and 13.6 kJ m-2 UVBBE). Absolute reductions
varied from 0 to 40% between the seed sources and with needle age.
For example, photosynthesis was significantly reduced by up to 40%
in needles which had been exposed to UVB for an entire season, but
only 19% on recently expanded needles. These reductions, however,
were only transient in some plants because they could not be
detected following the dormant winter period. This suggests that UVB
repair mechanisms may exist. Measurements of chlorophyll
fluorescence and the photosynthetic response to light indicated that
the quantum yield was significantly reduced in some cases by direct
effects on photosystem II. No significant effects were observed on
stomatal conductance of transpiration, and chlorophyll
concentrations were not generally altered by UVB exposure.
In vitro studies, using isolated chloroplasts, indicate that
UVB-induced damage to photochemical reactions is greater in C3
plants ( Dolichos lab lab, Phaseolus mungo, and Triticum vulgare)
than in C4 ( Amaranthus gangeticus, Zea mays, and Pennisetum
typhoides). Such differences are associated with the polypeptide
composition of the thylakoids (Kulandaivelu et al., 1993).
Studies in growth chambers with the ozone filter for
attenuating solar UVB report significant reductions in net
photosynthesis (measured under saturating light conditions) on a
leaf area and whole plant basis in sunflower seedlings, when grown
for three weeks at a daily maximum temperature of 28°C or 32°C under
a 20% higher UVB level compared to controls (5.1 kJ m-2 day-1
UVBBE vs. 3.6 kJ m-2 day-1). These represent average values
from May 1990 to August 1990 and are equivalent to approximately a
10% ozone depletion. In contrast, net photosynthesis was lower in
maize seedlings only during the earliest stages of development at
both temperatures (Tevini et al., 1991c).
11.2.4 Species competition
Enhanced UVB exposure can cause changes in the growth of plants
without necessarily decreasing plant production. Such changes
include reduced leaf length, increased branching, and increased
number of leaves (Barnes et al., 1990). These changes seem to be
general among different crop and weed species. Both graminoid and
broad-leaf species respond in this fashion, with graminoids
generally more responsive. Although these growth form changes do not
lead to changes in the production of monocultures, in mixed species
these alterations can lead to a change in the balance of competition
for light.
Multi-year field studies had shown that the balance of
competition between wheat and wild oat (a common weed) began to
favour wheat when mixtures of these species were subjected to
increased UVB irradiation (Barnes et al., 1988). In a recent study
involving canopy light microclimate assessments and a detailed
canopy radiation interception model, it was shown that the shift in
growth-form of the two species was sufficient to quantitatively
explain the change in the competitive balance (Ryel et al., 1990).
Thus, in many cases where plants are not necessarily depressed in
overall growth by increased UVB exposure, changes in growth-form can
have ecologically meaningful consequences. The direction of
competitive balance changes are not easily predicted at present.
However, altered competitive balance also has important implications
for mixed-crop agriculture and species composition of
nonagricultural ecosystems.
11.2.5 Plant diseases
Certain diseases may become more severe in plants exposed to
enhanced UVB levels. Sugar beet ( Beta vulgaris) plants infected
with Cercospora beticola, and receiving 6.9 kJ m-2 day-1
UVBBE, showed large reductions in leaf chlorophyll content, and
fresh and dry weight of total biomass. In another study, three
cucumber ( cucumis sativus) cultivars were exposed to a daily UVB
dose of 11.6 kJ m-2 UVBBE in a greenhouse before and/or after
infection with Colletotrichum lagenarium or Cladosporium
cucumerinum, and analyzed for disease development (Orth et al.,
1990). Two of the three cultivars were disease resistant and the
other was disease susceptible. Pre-infection treatment with UVB led
to greater disease development in the susceptible cultivar and in
one of the disease resistant cultivars. Post-infection treatment did
not alter disease development. The increased disease development in
UVB irradiated plants was found only on the cotyledons and not on
true leaves, suggesting that the effects of UVB on disease
development in cucumber vary according to the cultivar, timing of
UVB exposure, and tissue age.
11.2.6 UV-protection systems
Epidermal pigments
UVB induces flavonoid production (Wellmann, 1971), and may
regulate the synthesis of UV protective pigments (Braun & Tevini,
1991). In a study using two important crops (rye and oat),
UV-fluence and wavelength dependent accumulation of isovitexin
derivatives in the epidermal layer of rye seedlings prevented damage
to chloroplast functions. In contrast, photosynthetic function was
low without the accumulation of screening pigments (Tevini et al.,
1991a). Because the epidermal layer of oat seedlings already
accumulates large amounts of UV-absorbing pigments during early
development, the photosynthetic apparatus is better protected than
rye seedlings against UVB (Braun, 1991). This inherently higher
flavonoid production occurs even in absence of UVB irradiation, and
appears to be constitutive in nature. UVB induction of flavonoids
was demonstrated in two species of columbines, Aquilegia caerulea,
growing in alpine environments, and Aquilegia canadensis, which
grows at lower elevations (Larson et al., 1990). In both species,
flavonoid content increased upon UVB irradiation, even though the
alpine species accumulated higher amounts in the UVB-free controls
when compared to A canadensis after UVB irradiation. This
demonstrates that plants which are already genetically adapted to
higher UVB environments can further increase their adaptation
capacity.
Photorepair
A second protective mechanism in plants is photoreactivation.
The UV-induced production of DNA pyrimidine dimers can be repaired
by DNA photolysase. This enzyme was shown to increase with UVB
irradiation in Arabidopsis (Pang & Hays, 1991) but also by visible
light via phytochrome in bean seedlings (Langer & Wellmann, 1990).
This inducibility means that de novo synthesis of DNA photolysase
itself is a target for UV damage. Thus, the repair capacity of the
cell may be reduced in the presence of increasing UVB (Wellmann,
1991).
11.3 Effects on Aquatic Ecosystems
Aquatic ecosystems contribute more biomass (104 Gt a-1) than
all terrestrial ecosystems (100 Gt a-1) combined. Recent work on
UVB effects has concentrated on inhibition mechanisms and field
studies in the subpolar waters of Antarctica, because of its high
biomass production and the occurrence of the ozone hole over this
region.
Phytoplankton organisms orient within the water column using
external factors. Mobility and orientation mechanisms are impaired
by UVB exposure. Most organisms do not possess UVB receptors, and so
cannot avoid deleterious effects of enhanced UVB that produces
higher intensities deeper into the water column. New action spectra
indicate that, in addition to DNA, other targets absorb UVB
including proteins of the photoreceptor and photosynthetic
apparatus. The inability of phytoplankton to adjust their position
within the water column causes massive inhibition of photosynthesis.
In only a few cases have UVB inducible screening pigments been
identified.
11.3.1 Effects on phytoplankton
Recent UVB aquatic research has concentrated on phytoplankton
and the Antarctic ecosystem. As shown in figure 11.2, phytoplankton
is at the base of the aquatic food chain/trophic structure and
serves as food for primary consumers (e.g., larvae of fish and
shrimp), which in turn are consumed by secondary and tertiary
consumers (e.g. fish). The production of phytoplankton has been
estimated at about 6 x 1014 kg (UNEP, 1989). A loss of 10% would
far exceed the gross national product of all countries in the world,
assuming any reasonable price for biomass on the market. table 11.1
gives the estimated annual biomass production for plankton and fish.
Table 11.1 Estimated annual biomass production at different levels
in marine food web and possible loss after 10% decrease at the
phytoplankton level (adapted from UNEP 1989 report)
Type Annual Production 10% loss
(in million tonnes)
Phytoplankton 600,000 60,000
Zooplankton 60,000 6,000
Small fish 6,000 600
Large fish 600 60
Recent studies reveal the effects of UVB radiation on tropical
plants. Rice is among the most important tropical crops in the
world. Sixteen rice ( Oryza sativa L.) cultivars from several
different geographical regions when grown for 12 weeks in
greenhouses with supplemental levels of UVB exposure equivalent to
20% ozone depletion over the equator (15.7 kJ m-2 day-1 UVBBE)
showed alterations in biomass, morphology, and photosynthesis.
Approximately one-third of all cultivars tested showed a
statistically significant decrease in total biomass with increased
UVB exposure. Photosynthetic capacity declined for some cultivars,
but only a weak relationship existed between changes in
photosynthesis and biomass with increasing UVB exposure. In one of
the rice cultivars tested, total biomass significantly increased by
20% when grown under enhanced levels of UVB exposure. Therefore,
despite the fact that the effects of UVB are generally damaging, in
some cases, it has been reported to have a stimulating effect. Such
positive growth effects are presently not easily explainable.
Results from this experiment indicate that 1) a number of rice
cultivars are sensitive to increases in UVB exposure; 2) the
diversity exhibited by rice in response to increased levels of UVB
suggests that selective breeding might be successfully used to
develop UVB tolerant rice cultivars. Other preliminary screening
studies on rice seedlings also corroborate these observations
(Coronel et al., 1990).
In a three year field study (Sullivan & Teramura, 1991),
photosynthetic capacity was generally reduced in loblolly pine trees
exposed to supplemental levels of UVB simulating a 16% and 25% ozone
depletion (11.5 and 13.6 kJ m-2 UVBBE). Absolute reductions
varied from 0 to 40% between the seed sources and with needle age.
For example, photosynthesis was significantly reduced by up to 40%
in needles which had been exposed to UVB for an entire season, but
only 19% on recently expanded needles. These reductions, however,
were only transient in some plants because they could not be
detected following the dormant winter period. This suggests that UVB
repair mechanisms may exist. Measurements of chlorophyll
fluorescence and the photosynthetic response to light indicated that
the quantum yield was significantly reduced in some cases by direct
effects on photosystem II. No significant effects were observed on
stomatal conductance of transpiration, and chlorophyll
concentrations were not generally altered by UVB exposure.
In vitro studies, using isolated chloroplasts, indicate that
UVB-induced damage to photochemical reactions is greater in C3
plants ( Dolichos lab lab, Phaseolus mungo, and Triticum vulgare)
than in C4 ( Amaranthus gangeticus, Zea mays, and Pennisetum
typhoides). Such differences are associated with the polypeptide
composition of the thylakoids (Kulandaivelu et al., 1993).
Studies in growth chambers with the ozone filter for
attenuating solar UVB report significant reductions in net
photosynthesis (measured under saturating light conditions) on a
leaf area and whole plant basis in sunflower seedlings, when grown
for three weeks at a daily maximum temperature of 28°C or 32°C under
a 20% higher UVB level compared to controls (5.1 kJ m-2 day-1
UVBBE vs. 3.6 kJ m-2 day-1). These represent average values
from May 1990 to August 1990 and are equivalent to approximately a
10% ozone depletion. In contrast, net photosynthesis was lower in
maize seedlings only during the earliest stages of development at
both temperatures (Tevini et al., 1991c).
11.2.4 Species competition
Enhanced UVB exposure can cause changes in the growth of plants
without necessarily decreasing plant production. Such changes
include reduced leaf length, increased branching, and increased
number of leaves (Barnes et al., 1990). These changes seem to be
general among different crop and weed species. Both graminoid and
broad-leaf species respond in this fashion, with graminoids
generally more responsive. Although these growth form changes do not
lead to changes in the production of monocultures, in mixed species
these alterations can lead to a change in the balance of competition
for light.
Multi-year field studies had shown that the balance of
competition between wheat and wild oat (a common weed) began to
favour wheat when mixtures of these species were subjected to
increased UVB irradiation (Barnes et al., 1988). In a recent study
involving canopy light microclimate assessments and a detailed
canopy radiation interception model, it was shown that the shift in
growth-form of the two species was sufficient to quantitatively
explain the change in the competitive balance (Ryel et al., 1990).
Thus, in many cases where plants are not necessarily depressed in
overall growth by increased UVB exposure, changes in growth-form can
have ecologically meaningful consequences. The direction of
competitive balance changes are not easily predicted at present.
However, altered competitive balance also has important implications
for mixed-crop agriculture and species composition of
nonagricultural ecosystems.
11.2.5 Plant diseases
Certain diseases may become more severe in plants exposed to
enhanced UVB levels. Sugar beet ( Beta vulgaris) plants infected
with Cercospora beticola, and receiving 6.9 kJ m-2 day-1
UVBBE, showed large reductions in leaf chlorophyll content, and
fresh and dry weight of total biomass. In another study, three
cucumber ( cucumis sativus) cultivars were exposed to a daily UVB
dose of 11.6 kJ m-2 UVBBE in a greenhouse before and/or after
infection with Colletotrichum lagenarium or Cladosporium
cucumerinum, and analyzed for disease development (Orth et al.,
1990). Two of the three cultivars were disease resistant and the
other was disease susceptible. Pre-infection treatment with UVB led
to greater disease development in the susceptible cultivar and in
one of the disease resistant cultivars. Post-infection treatment did
not alter disease development. The increased disease development in
UVB irradiated plants was found only on the cotyledons and not on
true leaves, suggesting that the effects of UVB on disease
development in cucumber vary according to the cultivar, timing of
UVB exposure, and tissue age.
11.2.6 UV-protection systems
Epidermal pigments
UVB induces flavonoid production (Wellmann, 1971), and may
regulate the synthesis of UV protective pigments (Braun & Tevini,
1991). In a study using two important crops (rye and oat),
UV-fluence and wavelength dependent accumulation of isovitexin
derivatives in the epidermal layer of rye seedlings prevented damage
to chloroplast functions. In contrast, photosynthetic function was
low without the accumulation of screening pigments (Tevini et al.,
1991a). Because the epidermal layer of oat seedlings already
accumulates large amounts of UV-absorbing pigments during early
development, the photosynthetic apparatus is better protected than
rye seedlings against UVB (Braun, 1991). This inherently higher
flavonoid production occurs even in absence of UVB irradiation, and
appears to be constitutive in nature. UVB induction of flavonoids
was demonstrated in two species of columbines, Aquilegia caerulea,
growing in alpine environments, and Aquilegia canadensis, which
grows at lower elevations (Larson et al., 1990). In both species,
flavonoid content increased upon UVB irradiation, even though the
alpine species accumulated higher amounts in the UVB-free controls
when compared to A canadensis after UVB irradiation. This
demonstrates that plants which are already genetically adapted to
higher UVB environments can further increase their adaptation
capacity.
Photorepair
A second protective mechanism in plants is photoreactivation.
The UV-induced production of DNA pyrimidine dimers can be repaired
by DNA photolysase. This enzyme was shown to increase with UVB
irradiation in Arabidopsis (Pang & Hays, 1991) but also by visible
light via phytochrome in bean seedlings (Langer & Wellmann, 1990).
This inducibility means that de novo synthesis of DNA photolysase
itself is a target for UV damage. Thus, the repair capacity of the
cell may be reduced in the presence of increasing UVB (Wellmann,
1991).
11.3 Effects on Aquatic Ecosystems
Aquatic ecosystems contribute more biomass (104 Gt a-1) than
all terrestrial ecosystems (100 Gt a-1) combined. Recent work on
UVB effects has concentrated on inhibition mechanisms and field
studies in the subpolar waters of Antarctica, because of its high
biomass production and the occurrence of the ozone hole over this
region.
Phytoplankton organisms orient within the water column using
external factors. Mobility and orientation mechanisms are impaired
by UVB exposure. Most organisms do not possess UVB receptors, and so
cannot avoid deleterious effects of enhanced UVB that produces
higher intensities deeper into the water column. New action spectra
indicate that, in addition to DNA, other targets absorb UVB
including proteins of the photoreceptor and photosynthetic
apparatus. The inability of phytoplankton to adjust their position
within the water column causes massive inhibition of photosynthesis.
In only a few cases have UVB inducible screening pigments been
identified.
11.3.1 Effects on phytoplankton
Recent UVB aquatic research has concentrated on phytoplankton
and the Antarctic ecosystem. As shown in figure 11.2, phytoplankton
is at the base of the aquatic food chain/trophic structure and
serves as food for primary consumers (e.g., larvae of fish and
shrimp), which in turn are consumed by secondary and tertiary
consumers (e.g. fish). The production of phytoplankton has been
estimated at about 6 x 1014 kg (UNEP, 1989). A loss of 10% would
far exceed the gross national product of all countries in the world,
assuming any reasonable price for biomass on the market. table 11.1
gives the estimated annual biomass production for plankton and fish.
Table 11.1 Estimated annual biomass production at different levels
in marine food web and possible loss after 10% decrease at the
phytoplankton level (adapted from UNEP 1989 report)
Type Annual Production 10% loss
(in million tonnes)
Phytoplankton 600,000 60,000
Zooplankton 60,000 6,000
Small fish 6,000 600
Large fish 600 60
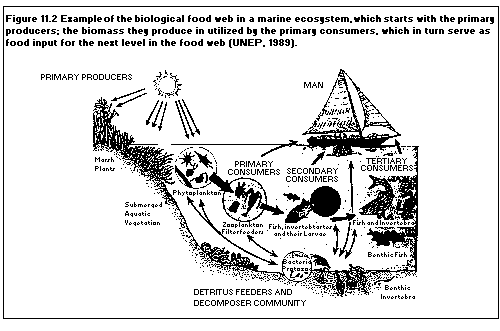 Concentrations of phytoplankton in subpolar waters may be 103
to 104 times greater than concentrations of phytoplankton found in
tropical and subtropical seas (Jeffery & Humphrey, 1975). Any
significant increase in UVB could well diminish growth and
productivity of phytoplankton, subsequently affecting all higher
trophic levels in the aquatic food web. Therefore, it is not
surprising that a majority of recent research has looked at the
effects of increased UVB exposure in Antarctic waters. Ongoing
research activities include investigations of both direct
(physiological and behavioural) and indirect (trophic implications)
effects.
Phytoplankton dwell in the top layers of the water column (the
photic zone) because of their requirement for solar energy
(Ignatiades, 1990). Their position within the column is maintained
by precise orientation strategies using light, gravity and other
external factors as guides. Phytoplankton in the photic zone would
be exposed to any increase in solar UV. Most phytoplankton organisms
do not possess UVB photoreceptors to guide them away from harmful
UV, a situation similar to humans. Previous work demonstrated that
mobility/orientation mechanisms in response to light are impaired by
solar UV (Häder & Worrest, 1991; Baker & Smith, 1982). The ability
of phytoplankton to adjust their position within the water column,
in response to constantly changing conditions, may even be affected
at ambient UVB levels. Although ambient UVB fluxes may cause damage
to some species of phytoplankton, it should be emphasized that there
are uncertainties regarding the magnitude of these effects. These
included problems of extrapolating laboratory findings to the open
sea and the nearly complete absence of data on long-term effects and
ecosystem responses. Likewise, there is a need to investigate
adaptation mechanisms. Before effects of exposure to solar UVB can
be predicted, information is required on seasonal abundances and
vertical distributions of marine organisms, vertical mixing, and the
penetration of UVB into appropriate water columns.
In their natural habitats, organisms are exposed to a wide
range of UVB intensities. This radiation has been shown to affect
growth, photosynthesis, nitrogen incorporation, and enzyme activity
(Döhler & Alt, 1989; Döhler, 1990).
11.3.2 UV increase and primary biomass production
Recent results indicate that orientation mechanisms responsive
to both light and gravity are affected by solar UV in a number of
ecologically significant phytoplankton groups (Häder & Lui, 1991).
Action spectra inhibitory effects are different from the DNA
absorption spectrum and the action spectra calculated for higher
plants, suggesting that UV exposure affects these organisms by a
different mechanism. Proteins essential for specific functions such
as orientation and photosynthesis are the primary targets of UVB.
Biochemical analyses conducted to reveal the molecular targets
of UVB inhibition show that specific photoreceptor proteins are
degraded. Simultaneously photosynthetic pigments (responsible for
converting solar energy) are bleached and destroyed by radiation
(UNEP, 1989). The results of these biochemical studies are further
supported by spectroscopic investigations showing losses in
pigmentation.
In order to evaluate the effects of enhanced UV, the vertical
movement of natural phytoplankton was analyzed in 3m Plexiglas
columns (Eggersdorfer & Häder, 1991). Most organisms moved to the
surface during daytime hours, although some species avoided periods
of intense UV during the midday hours by moving slightly down in the
water column. However, this avoidance response is not sufficient to
protect organisms under conditions of increased UVB irradiation
(UNEP, 1989).
The UVB irradiance in Antarctic waters significantly increased
during the occurrence of the ozone hole (Bidigare, 1989; Lubin et
al., 1989; Karentz & Lutze, 1990). However changes in marine
productivity accompanying UV flux changes have not been determined.
Recent measurements show that UVB penetrates 65 meters deep into
clear Antarctic waters (UNEP 1991). Consequently, measurements of
photosynthetic biomass production in Antarctic waters under the
ozone hole show a pronounced decrease in productivity by up to 25%
(Holm-Hansen, 1990).
Field studies indicate that photosynthesis is impaired first,
followed by decreases in protein concentration and changes in
pigment composition. As a result, a dramatic decrease in
photosynthetic oxygen production can be measured after exposure to
solar radiation (Smith et al., 1980). Other spectral bands, such
as UVA and visible radiation, may contribute to photosynthetic
inhibition (Smith et al., 1980). Likewise, photosynthetic
inhibition has been detected in macroalgae at their natural depth
(Bittersmann et al., 1988; Nultsch et al., 1990).
In contrast to higher plants only a few photoplanktons produce
UV absorbing substances (Carretto et al., 1990; Karentz et al.,
1991b). However, all of these mycosporine-like amino acids have
maximal absorption in the UVA range and only secondary peaks in the
UVB. It is not clear whether the production of these potentially
screening substances can be induced by exposing organisms to UV
(Raven, 1991). One exception is cyanobacteria where a UVB inducible
pigment has been found within the slime sheath surrounding the
organisms, which absorbs up to 88% of the incident UVB
(Garcia-Pichel and Castenholz, 1991).
11.4 Conclusion
Field and laboratory experiments on plant responses to
increased UVB radiation underscore the concern for agriculture,
forestry and natural ecosystems as the stratospheric ozone level is
depleted.
Growth and photosynthesis of certain crop plants can be
inhibited even under ambient levels of UVB radiation. Certain
environmental factors, both biotic (e.g. plant diseases and
competition with other plants) and abiotic (e.g. carbon dioxide,
temperature, heavy metals, and water availability) can alter UVB
effects in plants. This increases the difficulty in making any
quantitative predictions. Plants in temperate regions and certain
tropical species were found to be adversely affected by enhanced UVB
radiation.
Marine ecosystem which provides the primary food for human
consumption (in some countries) has been shown to be more sensitive
to UVB than terrestrial plants. One consequence of loss in
phytoplankton is reduced biomass production which would be
propagated throughout the whole food web. The marine phytoplankton
is a major absorber of atmospheric carbon dioxide. Any reduction in
this population would decrease the uptake of carbon dioxide and so
augment the greenhouse effect.
12. HEALTH HAZARD ASSESSMENT
12.1 Introduction
For the vast majority of people, the sun is the single largest
source of exposure to UV. In some cases solar exposure will be
elective such as from sunbathing, in others it will be adventitious
as a result of outdoor recreational and/or occupational activity.
Exposure may also occur from artificial sources of UV, either
deliberately for example during medical treatment or the use of a
sunbed for cosmetic purposes.
The health risks associated with exposure to UV include those
of both acute and chronic effects and will vary according to the
nature of the exposure. Factors important in assessing such risks
include: the biologically-effective irradiance of the UV incident on
the person exposed; the duration and frequency of occurrence of
exposures; and the individual sensitivity of the person to UV as
determined by genetic and other factors.
International guidelines on protection against UV given in
chapter 13 are based on available scientific data (IRPA/INIRC,
1991). The guidelines define occupational exposure limits (ELs)
below which it is expected that nearly all people may be repeatedly
exposed without adverse effects. The Els are intended to be used to
evaluate potentially hazardous exposures from, for example, solar
radiation, arcs, gas and vapour discharges, fluorescent lamps and
incandescent sources. The Els are generally below levels which are
often used for the UV exposure of patients required as part of
medical treatment and below levels associated with sunbed exposure.
IRPA/INIRC recommend that, where they are to be incorporated in
regulations, the ELs should be considered as absolute limits for the
eye, but only as advisory for the skin. This is because of the wide
range of susceptibility to skin injury depending on skin type. The
values were developed by considering lightly pigmented populations,
with greatest sensitivity to sunburn and non- melanocytic skin
cancer. In recommending similar limits, Threshold Limit Values
(TLVs), the American Conference of Governmental Industrial
Hygienists (ACGIH 1993) indicate that conditioned, tanned,
individuals can tolerate skin exposure in excess of the exposure
limits without sunburn effects, but that conditioning may not
protect individuals against skin cancer. ELs are not intended to
apply to exposure of pathologically photosensitive individuals, to
people concomitantly exposed to photosensitising agents or to
neonates.
The threshold for adverse acute effects may be exceeded in
certain people without exceeding the exposure limit. The risk of
skin cancer increases with cumulative exposure over time. Therefore,
it is recommended that all exposures should be reduced as far as is
reasonably practicable.
An estimate of the risk of cumulative exposure to UV can be
expressed in terms of the cumulative incidence (I) of non-melanoma
skin cancer in the form of the equation (Schothorst et al., 1985;
Diffey 1988).
I = gamma A Hß aalpha 12.1
where I is the total number of cases per 100,000 of the population
in the age group up to age 'a' years:
H is the annual carcinogenic-effective radiant exposure at the skin
surface:
A is the fraction of the body surface area exposed:
gamma, ß and alpha are numerical constants reflecting the genetic
susceptibility of the exposed population, the biological
amplification factor and the age dependence of the cumulative
incidence respectively.
The above equation applies only to situations where the annual
effective exposure remains constant year by year. This will not be
the case, for example, for workers whose job may result in
additional occupational exposure during their working lives (Diffey
1988). In these circumstances the cumulative incidence I can be
represented by the equation (Slaper et al., 1986).
I = gamma A Heß aalpha 12.2
where He is given by
He = H + Ho(a-as)/a
H is the annual effective radiant exposure of solar UV:
as is the age at which the occupational exposure began:
Ho is the annual effective radiant exposure from occupational
exposure.
In the following sections estimates of personal exposures
likely to result from different situations are considered and, where
the availability of data permits, examples of estimates of
quantitative risk are provided. The exposure situations include
elective exposure to solar UV, from medical treatment and from the
use of sunbeds, as well as adventitious exposure resulting from
solar radiation and from artificial sources of UV.
12.2 Elective Exposures
12.2.1 Medical exposure
UV is used in medicine for both the diagnosis and treatment of
disease. The highest and most extensive medical exposures to UV
result from phototherapy and photochemotherapy.
Medical treatment using UV carries the risk of acute side
effects and long term risks of chronic effects, in particular skin
cancer. The acute side effects of UVB phototherapy exposure include;
reddening, swelling, blistering and desquamation of the skin. The
acute effects following PUVA treatment include itching, skin pain
and nausea (Green et al., 1992).
Chronic effects include those on both the eyes and the skin.
However, the eyes of patients are generally well protected during
PUVA treatment and only one case of PUVA induced cataract in humans
appears to have been reported (Cyrlin 1980). Structural changes in
the skin, akin to those characteristic of long term damage resulting
from solar radiation exposure, have been observed as a result of
both UVB and PUVA phototherapy.
The risk of skin cancer from ultraviolet phototherapy has been
reviewed by Green et al. (1992). They conclude that data on the
association between UVB phototherapy and skin cancer are very poor.
Only one case control study has been reported and no correlation was
found between UVB phototherapy and skin cancer in 85 patients who
had received UVB phototherapy for up to 25 years. The association
between PUVA treatment and skin cancer in psoriasis patients has
been reported in ten studies. However, only one study (Stern et
al., 1979) indicated that PUVA acted as an independent carcinogen,
the risk of developing skin cancer being 6-12 times that of a
population survey. The results of other studies point to PUVA acting
in the role of a co-carcinogen, other factors involved being a
family history of skin cancer and treatment with antipsoriatic
medication. Comparisons and interpretations of data from US and
European studies have been complicated by differences in the
exposure regimens used.
12.2.2 Phototherapy of seasonal affective disorder (SAD)
Typical treatment consists of sitting in front of a panel of
fluorescent lamps and exposing the face, with the eyes open, to an
illuminance of about 2500 lux for 2 to 6 hours per day during the
winter months. This is equivalent to a daily erythemally-effective
radiant exposure of up to 40 J m-2 (Diffey 1993). An annual
cumulative exposure of up to 24 MEDs results, which, if continued
for several decades is estimated to result in a risk of non-melanoma
skin cancer of about 1.5, compared with someone (typical indoor
worker) minimally exposed.
12.2.3 Sunbeds
Among lightly pigmented people, a suntanned skin is
unfortunately still socially desirable. The establishment of the
'suntanning industry' has enabled the acquisition of a 'suntan'
irrespective of the availability of solar radiation. Suntanning is
caused by UV and as with exposure to the sun, there are attendant
risks in using sunbeds.
Adverse health effects of sunbed use include acute effects on
the skin, longer term structural damage of the skin and increased
risk of skin cancer (Roza et al., 1989). Provided that appropriate
protective eyewear is worn, adverse health effects on the eyes will
be avoided. The extent to which adverse, but relatively transient
skin effects occur is demonstrated by the results of a survey of the
use of sunbeds in the United Kingdom (Diffey 1986). Questionnaire
replies from over 1000 sunbed users indicated that the incidence of
acute adverse effects was substantial; 28% of users complained of
itching and about 8% developed a skin rash or had felt nauseous at
some time during or immediately after exposure. The incidence of
such effects was higher among women taking oral contraceptives than
in women who were not. No conclusion could be drawn about other
medications as few individuals were taking them. The survey also
revealed that 50% of sunbed users in the United Kingdom are female
and aged between 16 and 30 years. 43% of users had skin types I and
II, the most sensitive to the adverse effects of UV and the ones
least likely to tan well.
Structural damage to human skin from exposure to UVA, such as
demonstrated experimentally in mice (Kligman et al., 1987, Bissett
et al., 1989) might be expected in people as the result of
excessive use of sunbeds. Increased skin fragility and blistering
(Farr et al., 1988; Murphy 1989) and atypical melanocytic lesions
(Jones et al., 1987) have been observed in people who have used
UVA sunbeds excessively. In some people, photodermatosis and
polymorphic light eruption, is readily caused by exposure to UVA
radiation from a sunbed (Rivers et al., 1989). Certain
photo-aggravated dermatoses, such as lupus erythematosus, are
exacerbated by the use of UVA sunbeds (Stern and Docken 1986). The
use of certain medications such as antihypertensives and antibiotics
(Hawk 1984) and topical application of certain products, including
perfumes, body lotions, etc., may produce a photosensitising effect
on exposure to a sunbed.
Some localised skin and systemic changes in immunological
reactions result from exposure to UV and, of particular relevance to
sunbeds, from exposure to UVA. There is also evidence that exposure
to UV can accelerate the growth of human viruses (e.g. Otani and
Mori 1987, Perna et al., 1987), including human immunodeficiency
virus (HIV) (Zmudzka and Beer 1990). At present, the significance of
these observations with respect to the health of people exposed to
UV from sunbeds is unclear.
The risk of non-melanocytic skin cancer in northern Europeans
who sunbathe and who use sunbeds has been estimated by Diffey (1987)
using a mathematical model of skin cancer incidence that makes
allowance for childhood, occupational and recreational solar
radiation exposure. The model takes into account the fractions of
total skin surface area that might normally be exposed during
everyday activities and the larger fraction, normally unexposed, but
exposed during sunbathing and sunbed use. The annual effective
radiant exposures resulting from different activities and exposure
scenarios used as the basis for risk calculations are expressed as
representative minimal erythemal doses (MEDs) in table 12.1.
Table 12.1 Representative annual minimal erythemal doses (MEDs) for
various exposure scenarios in northern Europeans (Diffey, 1987,
1993a).
Scenario Annual
MEDs
Outdoor worker 270
Indoor worker 90
(including weekend exposure)
Sunbathing holiday in Mediterranean 50-100
area for 2 week period in summer
UVA sunbed (low pressure fluorescent 20
lamps, 30 x 30 minute sessions *
* Recommended maximum number of sessions per year (IRPA/INIRC,
1991a) with estimated exposure of 0.7 MED per session
(Diffey, 1987).
Diffey (1987) concluded that the increased risk of
non-melanocytic skin cancer associated with the use of a UVA sunbed
for 10 sessions per year is negligible. However, frequent use of a
sunbed, for example once per week from the age of 20 years will
result in an estimated doubling of the risk of non-melanocytic skin
cancer by age 45 years.
Several studies have reported increased risks of cutaneous
melanoma in users of sunlamps and sunbeds (IARC 1992).
12.2.4 Sunbathing
A significant contribution to the risk of non-melanocytic skin
cancer that results from sunbathing (Diffey, 1987). In these
calculations it was assumed that the subjects were indoor workers
who began sunbathing at age 20 years, and that their annual exposure
up to age 16 years is one half that of an outdoor worker. An indoor
worker who does not sunbathe is estimated to have a 2-3% risk of
non-melanoma skin cancer by age 70. Annual two week vacations spent
sunbathing at Mediterranean latitudes (approx. 40°N) will increase
this risk by a factor of about 5. Sunbathing for four weeks annually
is estimated to result in a 10-20 fold increase in cumulative risk
compared with non-sunbathers. It should be stressed that sunbathing
during vacation periods was assumed to take place for nearly the
whole day on every day of vacation. This is unlikely to be the case
for most holiday makers and so the estimated risk factors for most
people taking holidays in a sunny climate are likely to be one-third
to one-half of those given.
12.3 Adventitious Exposures
12.3.1 Outdoor exposures
Studies using personal dosimeters to monitor exposure of
individuals to UV have shown that, on average the
carcinogenic-effective UV exposure (excluding vacation exposure) of
outdoor workers is about 3 times that of indoor workers (Holman et
al., 1983; Larko and Diffey 1983; Schothorst et al., 1985).
Assuming that up to the age of 16, indoor and outdoor workers
receive the same carcinogenic-effective radiant exposure and that
both receive equivalent vacation exposures, the relative risk for
non-melanoma skin cancer for outdoor workers is estimated to be 3.7
times that for indoor workers (Diffey 1987). However, recent
population studies have found only small differences in skin cancer
incidence between outdoor and indoor workers (see chapter 8).
Armstrong & Kricker (1994) have estimated the proportion of
cutaneous malignant melanomas that is caused by sun exposure. The
estimated proportions varied from 0.97 in males and 0.96 in females
in Queensland, Australia, when the incidence on the whole body was
compared with the incidence on unexposed sites, to 0.68 when the
incidence in people born in Australia was compared with that in
migrants to Australia from areas of lower sun exposure. A comparison
of whites and blacks in the US in which the incidence in blacks was
taken as the incidence in unexposed whites, gave estimates of 0.96
in males and 0.92 in females. It was estimated that some 59,000
(65%) of about 92,000 malignant melanomas worldwide in 1985 were
caused by sun exposure.
12.3.2 Artificial sources
General public exposure
Exposure of the general public to potentially hazardous levels
of UV from artificial sources is unlikely. However, an area that
merits note is the increased use of tungsten halogen lamps for
lighting. Very high filament temperatures combined with quartz
envelopes results in the emission of significantly higher levels of
UV compared with conventional incandescent lamps (Césarini & Muel,
1989; McKinlay et al., 1989). For many applications of such lamps
the presence of a glass filter effectively attenuates the
potentially harmful UV. However, for some others, and particularly
when such lamps are incorporated in desk-top luminaires, no
protective filter is present, and significantly high levels of
erythemally effective UV have been measured. However, for lamps of
similar design and having nominally the same power, there is a wide
range of emitted levels of UV, (for example, between 2 and 56 mW
m-2 effective, in the beam of the lamp at 30 cm distance)
Occupational exposure
Sources that emit UV are used for a wide range of applications
in the workplace. High intensity discharge sources should be, and
often are, contained in interlocked enclosures thus obviating
hazardous exposure of people. Hazard evaluation surveys generally
consist of measurements of the effective exposure levels of UV and
comparison with recommended guidelines on limiting exposure. This
approach addresses the adverse acute effects of exposure, but very
few risk assessments have been carried out in relation to chronic
effects. The risk of non-melanocytic skin cancer for any
occupational situation can be calculated using multivariant
analysis, provided the effective exposures related to the occupation
are known. However, although data on levels of exposure exist for
some occupational situations there is generally a paucity of such
data. Where estimates of skin cancer risk have been made, they have
related to specific occupational exposure situations (Diffey 1988,
1989) or broadly with respect to general (fluorescent) lighting
(Lytle et al., 1993).
Examples where occupational exposure to UV from artificial
sources may occur include; electric arc plasma welding, the drying
and curing of inks, resins, plastics and paints; printing, graphics
arts, copying (using photographic processes) and photography,
photoetching, projector lamps operation, medicine, scientific
laboratory work, tanning and salon work and UV associated with
general lighting fluorescent lamps and desk-top and other luminaires
incorporating tungsten halogen lamps.
Welders are likely to be the largest occupational group with
potential exposure to UV (IARC 1992). The UV effective irradiance
levels around an operating welding arc can exceed recommended
exposure guideline levels by several orders of magnitude, and the
occurrence of acute effects of exposure, photokeratitis and
erythema, is common (Eriksen 1987).
Many industrial photoprocesses use high intensity discharge
lamps that emit copious quantities of UV. However, most of these
sources are effectively shielded and interlocked to prevent human
exposure. Some sources emit leakage radiation and, under these
circumstances, a detailed measurement survey is required to assess
the degree of hazard.
Medical physiotherapists involved in the phototherapy of
patients are occupationally exposed to UV and an analysis of
probable risk has been published (Diffey 1988). During a working
life of 40 years, the additional risk of non-melanoma skin cancer
for a member of this group is estimated to be around 25% compared
with that for non-exposed workers.
General lighting
Low pressure mercury vapour fluorescent lamps are ubiquitous in
the workplace and the home and concern has been expressed regarding
the potential role of the UV they emit in the aetiology of malignant
melanoma. Epidemiological data on exposure to general lighting
fluorescent lamps and malignant melanoma are few and inconsistent.
IRPA/INIRC (1991) concluded that UV exposure from indoor fluorescent
lighting should not be considered as a malignant melanoma risk.
The exposure levels associated with the use of general lighting
fluorescent lamps have been measured in a number of studies (Cole
et al., 1986; McKinlay & Whillock, 1987; Muel et al., 1988 and
Lytle et al., 1993). Exposure data vary considerably depending on
the types of lamps examined and the exposure conditions considered.
A recent study (Lytle et al., 1993) serves to illustrate estimates
of the risk associated with long term exposure to unfiltered general
lighting fluorescent lamps used in the United States. It was
estimated that 50 years exposure to typical unfiltered levels of UV
from fluorescent lamps added approximately 4% (1.6 - 12%) to any
risks associated with exposure to solar UV.
13. INTERNATIONAL GUIDELINES ON EXPOSURE TO ULTRAVIOLET RADIATION
A number of national and international organizations have
promulgated guidelines or standards on exposure to UV. Most are
based upon the same basic criteria of ACGIH (1993) and IRPA/INIRC
(1991).
The basic exposure limit (EL) for both general public and
occupational exposure to UV incident on the skin or eye is 30 J
m-2 effective), when the spectral irradiance Elambda at the eye
or skin surface is mathematically weighted with the hazard relative
spectral effectiveness factor Slambda from 180 nm to 400 nm. This
is given as follows:
Eeff = Sigma Elambda Slambda Deltalambda
where:
Eeff = effective irradiance W m-2
Elambda = spectral irradiance from measurements in W m-2
nm-1
Slambda = relative spectral effectiveness factor (unit-less)
Deltalambda = bandwidth of the calculation or measurement in
nm
At 270 nm in the UVC range, Slambda is 1.0, but at 360 nm in
the centre of the UVA range, its value falls to 0.00013, and
continues to fall for longer wavelengths.
For the UVA, the total radiant exposure incident on the
unprotected eye should not exceed 104 J m-2 (1 J cm-2) within
an 8 h period. The total 8 h radiant exposure incident on the
unprotected skin should not exceed the values in table 13.1.
The radiant UV exposure incident upon the unprotected skin or
eye within an 8-hour period should not exceed the values given in
table 13.1. The limits apply to sources whose emissions are measured
with an instrument having a cosine response detector oriented
perpendicular to the most directly exposed surfaces of the body when
assessing skin exposure and along (or parallel to) the line(s) of
sight when assessing ocular exposure. Although no measurement
averaging aperture is recommended, 1 mm is commonly used.
Table 13.1 International UV exposure limits and spectral weighting
factor (IRPA/INIRC, 1991)
Wavelengtha (nm) EL EL Relative Spectral
(J m-2) (mJ cm-2) Effectiveness Slamda
180 2,500 250 0.012
190 1,600 160 0.019
200 1,000 100 0.030
205 590 59 0.051
210 400 40 0.075
215 320 32 0.095
220 250 25 0.120
225 200 20 0.150
230 160 16 0.190
235 130 13 0.240
240 100 20 0.300
245 83 8.3 0.360
250 70 7.0 0.430
254b 60 6.0 0.500
255 58 5.8 0.520
260 46 4.6 0.650
265 37 3.7 0.810
270 30 3.0 1.000
275 31 3.1 0.960
280b 34 3.4 0.880
285 39 3.9 0.770
290 47 4.7 0.640
295 56 5.6 0.540
297b 65 6.5 0.460
300 100 10 0.300
303b 250 25 0.190
305 500 50 0.060
308 1,200 120 0.026
310 2,000 200 0.015
313b 5,000 500 0.006
315 1.0 x 104 1.0 x 103 0.003
316 1.3 x 104 1.3 x 103 0.0024
317 1.5 x 104 1.5 x 103 0.0020
318 1.9 x 104 1.9 x 103 0.0016
319 2.5 x 104 2.5 x 103 0.0012
320 2.9 x 104 2.9 x 103 0.0010
322 4.5 x 104 4.5 x 103 0.00067
323 5.6 x 104 5.6 x 103 0.00054
325 6.0 x 104 6.0 x 103 0.00050
328 6.8 x 104 6.8 x 103 0.00044
330 7.3 x 104 7.3 x 103 0.00041
333 8.1 x 104 8.1 x 103 0.00037
335 8.8 x 104 8.8 x 103 0.00034
340 1.1 x 105 1.1 x 104 0.00028
Table 13.1 (contd).
Wavelengtha (nm) EL EL Relative Spectral
(J m-2) (mJ cm-2) Effectiveness Slamda
345 1.3 x 105 1.3 x 104 0.00024
350 1.5 x 105 1.5 x 104 0.00020
355 1.9 x 105 1.9 x 104 0.00016
360 2.3 x 105 2.3 x 104 0.00013
365b 2.7 x 105 2.7 x 104 0.00011
370 3.2 x 105 3.2 x 104 0.000093
375 3.9 x 105 3.9 x 104 0.000077
380 4.7 x 105 4.7 x 104 0.000064
385 5.7 x 105 5.7 x 104 0.000053
390 6.8 x 105 6.8 x 104 0.000044
395 8.3 x 105 8.3 x 104 0.000036
400 1.0 x 106 1.0 x 105 0.000030
a Wavelengths chosen are representative; other values should be
interpolated at intermediate wavelengths.
b Emission lines of a mercury discharge spectrum.
The permissible exposure duration, tmax, for exposure (in
seconds) to UV is calculated by:
tmax = 30/ Eeff (W m-2)
Examples are provided in table 13.2.
Table 13.2 Limiting UV exposure durations based on exposure limits
(IRPA/INIRC, 1991)
Duration of exposure Effective irradiance
per day Eeff (W m-2) Eeff (µW cm-2)
8 hours 0.001 0.1
4 hours 0.002 0.2
2 hours 0.004 0.4
1 hour 0.008 0.8
30 minutes 0.017 1.7
15 minutes 0.033 3.3
10 minutes 0.05 5
The EL's were developed considering lightly pigmented
populations with greatest sensitivity and predisposition to adverse
health effects from exposure to UV. The limits apply to UV exposure
of the working population, but with some precaution also apply to
the general public. However, some rare, highly photosensitive
individuals exist who may react adversely to exposure at these
levels. These individuals are normally aware of their heightened
sensitivity. Likewise, if individuals are concomitantly exposed to
photosensitizing agents, an enhanced reaction can take place. Many
individuals who are exposed to photosensitizing agents (ingested or
externally applied chemicals, e.g., in cosmetics, foods, drugs,
industrial chemicals, etc.) may not be aware of their heightened
sensitivity. Lightly pigmented individuals conditioned by previous
UV exposure (leading to tanning and hyperplasia) and heavily
pigmented individuals can tolerate skin exposure in excess of the
EL's without erythemal effects. However, repeated tanning may
increase the risk of accelerated skin aging and even skin cancer.
Such risks should be understood prior to the use of UV for medical
phototherapy or cosmetic exposures.
14. PROTECTIVE MEASURES
14.1 Introduction
It is widely accepted by scientific and medical authorities
throughout the world that UV is potentially carcinogenic and capable
of producing other undesirable health effects. It is sensible
therefore to take steps to minimise UV exposure. Many risks in life
are completely beyond our control such as contracting a rare
disease. However, risks to health associated with exposure to UV
from both natural and artificial sources can be substantially
reduced by taking appropriate control measures.
Since UV exposure occurs externally, simple measures can be
taken to reduce the exposures received. A high degree of protection
can be afforded by protective clothing (including hats), UV
protective eyewear (welding helmets, face shields, goggles,
sunglasses, spectacles etc.) and by sunscreens for exposed skin.
However, the degree of protection afforded can be reduced by
ingestion of photosensitizing drugs or photoallergic/phototoxic
reactions produced by chemicals or cosmetics in contact with the
skin. Thus, education is also an important control measure.
14.2 Education
Concern about high incidences of skin cancer and eye damage
have led to national educational campaigns in some countries to
encourage people to protect themselves against excessive UV exposure
from the sun and in the workplace. Educational programmes directed
at both the workforce and the public are intended to create an
awareness of the adverse health effects that can result from
overexposure to UV.
Presently, in several different countries around the world,
daily environmental UV levels are supplied to the general public in
the form of UV indices. Their provision is intended to educate the
public on the basic climatology of UV, increase awareness of the
hazards of UV and provide information necessary to plan protection.
In a report commissioned to investigate Canadian attitudes to
Environment Canada's UV index (Environment Canada, 1993), it was
found that 73% of Canadians were aware of the UV index, 91% believed
UV affected human health, and most important, 59% of respondents who
were aware of the UV index had changed their sun exposure habits.
Different countries have adopted their own national UV indices.
Unfortunately, they are not all compatible. Clearly, international
uniformity would help to prevent confusion.
Educational programmes aim to produce a change in knowledge and
attitudes, then a change in behaviour and eventually a reduction in
the incidence and mortality rates of skin cancer. A survey of
sunscreen use on beaches in Brisbane, Australia found about 70% of
females and males applied sunscreen. Half of the sunscreens provided
the maximum protection (SPF 15+) and almost 90% used a waterproof
formulation. However, the sunscreen was not applied over the entire
body with over half neglecting ears and lower limbs (Pincus et al.,
1991). Survey results show that sunburn is still occurring; in a
randomly selected group of adults in Melbourne, Australia 16%
reported sunburn over a summer weekend (Hill et al., 1992). With
regard to skin cancers it is still too early to evaluate the
effectiveness of current educational/publicity campaigns.
With respect to changes in behaviour, there is good reason to
avoid exposure to the midday sun since it has been estimated that up
to one third of the day's erythemally effective UV is received
within the period one hour before noon to one hour after noon. Shade
is a useful method of protection but its value should not be
over-estimated, as one may be exposed to a quarter or more of the
total solar UV while shaded from direct sunlight, depending on the
prevailing exposure conditions.
14.3 Protection Factors
The concept of a protection factor is useful when attempting to
quantify the UV protection that items such as sunscreens, clothing
and eyewear can provide (Gies et al., 1992). To determine the
protection factor, the following procedure is conducted. An
effective dose (ED) of UV to the unprotected skin or eye is
calculated by summing the incident solar spectral power over the
wavelength range 280 to 400 nm. In order to determine the effective
dose (EDm) for the skin or eye when it is protected, the
calculation is repeated with the spectral transmission of the
protection item as an additional weighting. The protection factor
(PF) is then defined as the ratio of ED to EDm and is given by the
following equation:
ED SigmaElambda.Slambda.Delta lambda
PF =
EDm SigmaElambda.Slambda.Tlambda.Delta lambda
where:
Elambda = spectral irradiance (W m-2 nm-1) at wavelength lambda
Slambda = relative spectral effectiveness
Tlambda = spectral transmission of protective item at
wavelength lambda
Delta lambda = wavelength interval or bandwidth (nm)
lambda = wavelength (nm)
The inclusion of the spectral effectiveness function in the
calculation ensures that sufficient weighting is given to the
biologically effective wavelengths below 315 nm. A description of
how the protection factors for fabrics, eyewear and sunscreens are
determined is given in Roy and Gies (1993) and CIE (1991).
14.4 Clothing
Use of protective clothing provides one of the simplest means
of reducing UV exposure. Hats have been shown to afford protection,
to various degrees, to the forehead, scalp, ears and most of the
neck. Their protective properties have been studied by Diffey and
Cheeseman (1992). In this study hats were classified into four
categories, small brim, medium brim, large brim and peaked cap. The
protection afforded to different anatomical sites on the head and
neck is shown in table 14.1.
Table 14.1 Sun protection for various anatomical sites on the head
and neck provided by different types of hat (Diffey & Cheeseman,
1992).
Typical sun protection factor *
Style of hat Forehead Nose Cheek Chin Back of
neck
Small brim 15 1.5 1 1 1
< 2.5 cm
Medium brim >20 3 2 1 1
2.5-7.5 cm
Large brim >20 7 3 1.2 5
>7.5 cm
Peaked cap >20 5 1.5 1 1
* In this table 'sun protection factor' is defined as the reciprocal
of the fraction of UV exposure recorded relative to that of the
unprotected head.
The degree of protection provided by clothing depends on the
penetration of UV through materials and this can vary considerably.
Fabrics which are visibly opaque tend to be more highly absorbent of
UV, but the structure or weave of a material is the most important
factor in determining its protective value. Colour and thickness are
a poor guide to UV protection. The transmission properties of some
fabrics commonly used in the manufacture of clothing for everyday
wear are given in table 14.2 (Welsh and Diffey 1981). Here the
protection factor is an estimate of the protection afforded against
biologically effective solar radiation. A high protection factor is
associated with a tightly woven material. UV is transmitted and
scattered through the interstices of the material itself rather than
penetrating the fabric.
Gies et al. (1992) have recently extended the concept of
protection factor (PF) to fabrics and proposed the Ultraviolet
Protection Factor (UPF). The UPF scheme, as shown in table 14.3, is
designed to give the general public information on the amount of UV
protection available from fabrics and clothing and is now in use in
Australia. UPF is analogous to SPF and a fabric of UPF 10 would, in
principle, provide a similar level of protection as a sunscreen of
SPF 10.
Table 14.2 UVB transmission properties of common fabrics
(from Welsh & Diffey 1981)
Fabric Structure Colour Thickness % of Protection
(mm) incident factor
UV
transmitted
Nylon-tricel Woven Black 0.1 0.15 750
Nylon-viscose jacquard Woven Black 0.2 0.20 500
Nylon Woven White 0.1 1.7 55
Nylon-terylene mixture Knitted Blue 0.2 11 9
Nylon-acetate jersey Knitted Pink 0.2 24 4
Polyester-slub viscose Woven Pink 0.5 9 14
Polyester-printed lawn Woven Red 0.2 7 11
Polyester-jersey Knitted Fawn 0.3 7 14
Polyester-jersey Knitted Cream 0.4 5 19
Polyester-jersey Knitted Black 0.3 8 12
Polyester-jersey Knitted Orange 0.3 5 23
Polyester-jersey Knitted Turquoise 0.5 16 6
Polyester-jersey Knitted Brown 0.7 1.6 68
Polyester-jersey Knitted Black 0.5 4.4 23
Polyester-brushed jersey Knitted Blue 0.4 5.2 19
Polyester-brushed jersey Knitted Green 0.4 6.0 16
Polyester-bouclette Knitted Orange 0.4 3.0 33
Polyester-bouclette Knitted Purple 0.4 2.0 51
Cotton-needlecord Woven Brown 0.5 < 0.1 > 1000
Cotton-denim Woven Blue 0.5 < 0.1 > 1000
Cotton-printed Woven Brown 0.3 < 0.1 >1000
Cotton-printed Woven Cream 0.3 2.7 36
Wool-jersey Knitted Fawn 0.7 0.7 150
Table 14.3 Summary of the ultraviolet protection
factor scheme for fabrics
Ultraviolet Mean % UV Protection
protection transmission category
factor UPF
UPF 40+ less than 2.5 Maximum protection 40+
UPF 30 to 39 3.3 to 2.5 Very high protection
UPF 20 to 29 5.0 to 3.3 High protection
Whether a material is wet or dry is important in relation to
its UV transmission properties. The spectral transmittances of some
cotton and polyester-cotton samples are shown in figure 14.1.
Measurements show that the variation of UPF for wet and dry fabrics
is consistent for cotton, all examples showing a decrease in UPF
when wet (Gies et al., 1992). The variation in UPFs between wet
and dry was less consistent for polyester- cotton than for the
cotton, some UPFs increasing for wet fabric, while others decreased.
14.5 Sunscreens
Sunscreens are physical and chemical topical preparations which
attenuate the transmission of solar UV into the skin by absorption,
reflection or scattering. Physical sunscreens (sunblocks), for
example zinc oxide, titanium dioxide or red ferric oxide, function
by reflecting and scattering and provide protection against a broad
spectrum of UV and visible wavelengths. They are normally nontoxic
and have few known adverse effects. Sunscreens based on chemical
absorbers contain one or more colourless UV-absorbing ingredients
which generally absorb UVB radiation more strongly that UVA.
Para-aminobenzoic acid (PABA) and its derivatives, salicylates,
cinnamates and camphor derivatives primarily absorb UVB and transmit
UVA; benzophenones essentially absorb UV of wavelengths of less than
360 nm. The use of solely UVA absorbers (di-benzoyl-methane) is
allowed in only certain countries. These chemicals are all based on
benzene (Moseley 1988).
The application of any sunscreen normally changes the spectrum
of UV that reaches the target cells. Although most sunscreens are
designed to attenuate UV, some contain additives such as bergamot
oil (containing 5-methoxypsoralen) to enhance pigmentation and
photoprotection (Young et al., 1991). The role of such
preparations remains controversial.
Concentrations of phytoplankton in subpolar waters may be 103
to 104 times greater than concentrations of phytoplankton found in
tropical and subtropical seas (Jeffery & Humphrey, 1975). Any
significant increase in UVB could well diminish growth and
productivity of phytoplankton, subsequently affecting all higher
trophic levels in the aquatic food web. Therefore, it is not
surprising that a majority of recent research has looked at the
effects of increased UVB exposure in Antarctic waters. Ongoing
research activities include investigations of both direct
(physiological and behavioural) and indirect (trophic implications)
effects.
Phytoplankton dwell in the top layers of the water column (the
photic zone) because of their requirement for solar energy
(Ignatiades, 1990). Their position within the column is maintained
by precise orientation strategies using light, gravity and other
external factors as guides. Phytoplankton in the photic zone would
be exposed to any increase in solar UV. Most phytoplankton organisms
do not possess UVB photoreceptors to guide them away from harmful
UV, a situation similar to humans. Previous work demonstrated that
mobility/orientation mechanisms in response to light are impaired by
solar UV (Häder & Worrest, 1991; Baker & Smith, 1982). The ability
of phytoplankton to adjust their position within the water column,
in response to constantly changing conditions, may even be affected
at ambient UVB levels. Although ambient UVB fluxes may cause damage
to some species of phytoplankton, it should be emphasized that there
are uncertainties regarding the magnitude of these effects. These
included problems of extrapolating laboratory findings to the open
sea and the nearly complete absence of data on long-term effects and
ecosystem responses. Likewise, there is a need to investigate
adaptation mechanisms. Before effects of exposure to solar UVB can
be predicted, information is required on seasonal abundances and
vertical distributions of marine organisms, vertical mixing, and the
penetration of UVB into appropriate water columns.
In their natural habitats, organisms are exposed to a wide
range of UVB intensities. This radiation has been shown to affect
growth, photosynthesis, nitrogen incorporation, and enzyme activity
(Döhler & Alt, 1989; Döhler, 1990).
11.3.2 UV increase and primary biomass production
Recent results indicate that orientation mechanisms responsive
to both light and gravity are affected by solar UV in a number of
ecologically significant phytoplankton groups (Häder & Lui, 1991).
Action spectra inhibitory effects are different from the DNA
absorption spectrum and the action spectra calculated for higher
plants, suggesting that UV exposure affects these organisms by a
different mechanism. Proteins essential for specific functions such
as orientation and photosynthesis are the primary targets of UVB.
Biochemical analyses conducted to reveal the molecular targets
of UVB inhibition show that specific photoreceptor proteins are
degraded. Simultaneously photosynthetic pigments (responsible for
converting solar energy) are bleached and destroyed by radiation
(UNEP, 1989). The results of these biochemical studies are further
supported by spectroscopic investigations showing losses in
pigmentation.
In order to evaluate the effects of enhanced UV, the vertical
movement of natural phytoplankton was analyzed in 3m Plexiglas
columns (Eggersdorfer & Häder, 1991). Most organisms moved to the
surface during daytime hours, although some species avoided periods
of intense UV during the midday hours by moving slightly down in the
water column. However, this avoidance response is not sufficient to
protect organisms under conditions of increased UVB irradiation
(UNEP, 1989).
The UVB irradiance in Antarctic waters significantly increased
during the occurrence of the ozone hole (Bidigare, 1989; Lubin et
al., 1989; Karentz & Lutze, 1990). However changes in marine
productivity accompanying UV flux changes have not been determined.
Recent measurements show that UVB penetrates 65 meters deep into
clear Antarctic waters (UNEP 1991). Consequently, measurements of
photosynthetic biomass production in Antarctic waters under the
ozone hole show a pronounced decrease in productivity by up to 25%
(Holm-Hansen, 1990).
Field studies indicate that photosynthesis is impaired first,
followed by decreases in protein concentration and changes in
pigment composition. As a result, a dramatic decrease in
photosynthetic oxygen production can be measured after exposure to
solar radiation (Smith et al., 1980). Other spectral bands, such
as UVA and visible radiation, may contribute to photosynthetic
inhibition (Smith et al., 1980). Likewise, photosynthetic
inhibition has been detected in macroalgae at their natural depth
(Bittersmann et al., 1988; Nultsch et al., 1990).
In contrast to higher plants only a few photoplanktons produce
UV absorbing substances (Carretto et al., 1990; Karentz et al.,
1991b). However, all of these mycosporine-like amino acids have
maximal absorption in the UVA range and only secondary peaks in the
UVB. It is not clear whether the production of these potentially
screening substances can be induced by exposing organisms to UV
(Raven, 1991). One exception is cyanobacteria where a UVB inducible
pigment has been found within the slime sheath surrounding the
organisms, which absorbs up to 88% of the incident UVB
(Garcia-Pichel and Castenholz, 1991).
11.4 Conclusion
Field and laboratory experiments on plant responses to
increased UVB radiation underscore the concern for agriculture,
forestry and natural ecosystems as the stratospheric ozone level is
depleted.
Growth and photosynthesis of certain crop plants can be
inhibited even under ambient levels of UVB radiation. Certain
environmental factors, both biotic (e.g. plant diseases and
competition with other plants) and abiotic (e.g. carbon dioxide,
temperature, heavy metals, and water availability) can alter UVB
effects in plants. This increases the difficulty in making any
quantitative predictions. Plants in temperate regions and certain
tropical species were found to be adversely affected by enhanced UVB
radiation.
Marine ecosystem which provides the primary food for human
consumption (in some countries) has been shown to be more sensitive
to UVB than terrestrial plants. One consequence of loss in
phytoplankton is reduced biomass production which would be
propagated throughout the whole food web. The marine phytoplankton
is a major absorber of atmospheric carbon dioxide. Any reduction in
this population would decrease the uptake of carbon dioxide and so
augment the greenhouse effect.
12. HEALTH HAZARD ASSESSMENT
12.1 Introduction
For the vast majority of people, the sun is the single largest
source of exposure to UV. In some cases solar exposure will be
elective such as from sunbathing, in others it will be adventitious
as a result of outdoor recreational and/or occupational activity.
Exposure may also occur from artificial sources of UV, either
deliberately for example during medical treatment or the use of a
sunbed for cosmetic purposes.
The health risks associated with exposure to UV include those
of both acute and chronic effects and will vary according to the
nature of the exposure. Factors important in assessing such risks
include: the biologically-effective irradiance of the UV incident on
the person exposed; the duration and frequency of occurrence of
exposures; and the individual sensitivity of the person to UV as
determined by genetic and other factors.
International guidelines on protection against UV given in
chapter 13 are based on available scientific data (IRPA/INIRC,
1991). The guidelines define occupational exposure limits (ELs)
below which it is expected that nearly all people may be repeatedly
exposed without adverse effects. The Els are intended to be used to
evaluate potentially hazardous exposures from, for example, solar
radiation, arcs, gas and vapour discharges, fluorescent lamps and
incandescent sources. The Els are generally below levels which are
often used for the UV exposure of patients required as part of
medical treatment and below levels associated with sunbed exposure.
IRPA/INIRC recommend that, where they are to be incorporated in
regulations, the ELs should be considered as absolute limits for the
eye, but only as advisory for the skin. This is because of the wide
range of susceptibility to skin injury depending on skin type. The
values were developed by considering lightly pigmented populations,
with greatest sensitivity to sunburn and non- melanocytic skin
cancer. In recommending similar limits, Threshold Limit Values
(TLVs), the American Conference of Governmental Industrial
Hygienists (ACGIH 1993) indicate that conditioned, tanned,
individuals can tolerate skin exposure in excess of the exposure
limits without sunburn effects, but that conditioning may not
protect individuals against skin cancer. ELs are not intended to
apply to exposure of pathologically photosensitive individuals, to
people concomitantly exposed to photosensitising agents or to
neonates.
The threshold for adverse acute effects may be exceeded in
certain people without exceeding the exposure limit. The risk of
skin cancer increases with cumulative exposure over time. Therefore,
it is recommended that all exposures should be reduced as far as is
reasonably practicable.
An estimate of the risk of cumulative exposure to UV can be
expressed in terms of the cumulative incidence (I) of non-melanoma
skin cancer in the form of the equation (Schothorst et al., 1985;
Diffey 1988).
I = gamma A Hß aalpha 12.1
where I is the total number of cases per 100,000 of the population
in the age group up to age 'a' years:
H is the annual carcinogenic-effective radiant exposure at the skin
surface:
A is the fraction of the body surface area exposed:
gamma, ß and alpha are numerical constants reflecting the genetic
susceptibility of the exposed population, the biological
amplification factor and the age dependence of the cumulative
incidence respectively.
The above equation applies only to situations where the annual
effective exposure remains constant year by year. This will not be
the case, for example, for workers whose job may result in
additional occupational exposure during their working lives (Diffey
1988). In these circumstances the cumulative incidence I can be
represented by the equation (Slaper et al., 1986).
I = gamma A Heß aalpha 12.2
where He is given by
He = H + Ho(a-as)/a
H is the annual effective radiant exposure of solar UV:
as is the age at which the occupational exposure began:
Ho is the annual effective radiant exposure from occupational
exposure.
In the following sections estimates of personal exposures
likely to result from different situations are considered and, where
the availability of data permits, examples of estimates of
quantitative risk are provided. The exposure situations include
elective exposure to solar UV, from medical treatment and from the
use of sunbeds, as well as adventitious exposure resulting from
solar radiation and from artificial sources of UV.
12.2 Elective Exposures
12.2.1 Medical exposure
UV is used in medicine for both the diagnosis and treatment of
disease. The highest and most extensive medical exposures to UV
result from phototherapy and photochemotherapy.
Medical treatment using UV carries the risk of acute side
effects and long term risks of chronic effects, in particular skin
cancer. The acute side effects of UVB phototherapy exposure include;
reddening, swelling, blistering and desquamation of the skin. The
acute effects following PUVA treatment include itching, skin pain
and nausea (Green et al., 1992).
Chronic effects include those on both the eyes and the skin.
However, the eyes of patients are generally well protected during
PUVA treatment and only one case of PUVA induced cataract in humans
appears to have been reported (Cyrlin 1980). Structural changes in
the skin, akin to those characteristic of long term damage resulting
from solar radiation exposure, have been observed as a result of
both UVB and PUVA phototherapy.
The risk of skin cancer from ultraviolet phototherapy has been
reviewed by Green et al. (1992). They conclude that data on the
association between UVB phototherapy and skin cancer are very poor.
Only one case control study has been reported and no correlation was
found between UVB phototherapy and skin cancer in 85 patients who
had received UVB phototherapy for up to 25 years. The association
between PUVA treatment and skin cancer in psoriasis patients has
been reported in ten studies. However, only one study (Stern et
al., 1979) indicated that PUVA acted as an independent carcinogen,
the risk of developing skin cancer being 6-12 times that of a
population survey. The results of other studies point to PUVA acting
in the role of a co-carcinogen, other factors involved being a
family history of skin cancer and treatment with antipsoriatic
medication. Comparisons and interpretations of data from US and
European studies have been complicated by differences in the
exposure regimens used.
12.2.2 Phototherapy of seasonal affective disorder (SAD)
Typical treatment consists of sitting in front of a panel of
fluorescent lamps and exposing the face, with the eyes open, to an
illuminance of about 2500 lux for 2 to 6 hours per day during the
winter months. This is equivalent to a daily erythemally-effective
radiant exposure of up to 40 J m-2 (Diffey 1993). An annual
cumulative exposure of up to 24 MEDs results, which, if continued
for several decades is estimated to result in a risk of non-melanoma
skin cancer of about 1.5, compared with someone (typical indoor
worker) minimally exposed.
12.2.3 Sunbeds
Among lightly pigmented people, a suntanned skin is
unfortunately still socially desirable. The establishment of the
'suntanning industry' has enabled the acquisition of a 'suntan'
irrespective of the availability of solar radiation. Suntanning is
caused by UV and as with exposure to the sun, there are attendant
risks in using sunbeds.
Adverse health effects of sunbed use include acute effects on
the skin, longer term structural damage of the skin and increased
risk of skin cancer (Roza et al., 1989). Provided that appropriate
protective eyewear is worn, adverse health effects on the eyes will
be avoided. The extent to which adverse, but relatively transient
skin effects occur is demonstrated by the results of a survey of the
use of sunbeds in the United Kingdom (Diffey 1986). Questionnaire
replies from over 1000 sunbed users indicated that the incidence of
acute adverse effects was substantial; 28% of users complained of
itching and about 8% developed a skin rash or had felt nauseous at
some time during or immediately after exposure. The incidence of
such effects was higher among women taking oral contraceptives than
in women who were not. No conclusion could be drawn about other
medications as few individuals were taking them. The survey also
revealed that 50% of sunbed users in the United Kingdom are female
and aged between 16 and 30 years. 43% of users had skin types I and
II, the most sensitive to the adverse effects of UV and the ones
least likely to tan well.
Structural damage to human skin from exposure to UVA, such as
demonstrated experimentally in mice (Kligman et al., 1987, Bissett
et al., 1989) might be expected in people as the result of
excessive use of sunbeds. Increased skin fragility and blistering
(Farr et al., 1988; Murphy 1989) and atypical melanocytic lesions
(Jones et al., 1987) have been observed in people who have used
UVA sunbeds excessively. In some people, photodermatosis and
polymorphic light eruption, is readily caused by exposure to UVA
radiation from a sunbed (Rivers et al., 1989). Certain
photo-aggravated dermatoses, such as lupus erythematosus, are
exacerbated by the use of UVA sunbeds (Stern and Docken 1986). The
use of certain medications such as antihypertensives and antibiotics
(Hawk 1984) and topical application of certain products, including
perfumes, body lotions, etc., may produce a photosensitising effect
on exposure to a sunbed.
Some localised skin and systemic changes in immunological
reactions result from exposure to UV and, of particular relevance to
sunbeds, from exposure to UVA. There is also evidence that exposure
to UV can accelerate the growth of human viruses (e.g. Otani and
Mori 1987, Perna et al., 1987), including human immunodeficiency
virus (HIV) (Zmudzka and Beer 1990). At present, the significance of
these observations with respect to the health of people exposed to
UV from sunbeds is unclear.
The risk of non-melanocytic skin cancer in northern Europeans
who sunbathe and who use sunbeds has been estimated by Diffey (1987)
using a mathematical model of skin cancer incidence that makes
allowance for childhood, occupational and recreational solar
radiation exposure. The model takes into account the fractions of
total skin surface area that might normally be exposed during
everyday activities and the larger fraction, normally unexposed, but
exposed during sunbathing and sunbed use. The annual effective
radiant exposures resulting from different activities and exposure
scenarios used as the basis for risk calculations are expressed as
representative minimal erythemal doses (MEDs) in table 12.1.
Table 12.1 Representative annual minimal erythemal doses (MEDs) for
various exposure scenarios in northern Europeans (Diffey, 1987,
1993a).
Scenario Annual
MEDs
Outdoor worker 270
Indoor worker 90
(including weekend exposure)
Sunbathing holiday in Mediterranean 50-100
area for 2 week period in summer
UVA sunbed (low pressure fluorescent 20
lamps, 30 x 30 minute sessions *
* Recommended maximum number of sessions per year (IRPA/INIRC,
1991a) with estimated exposure of 0.7 MED per session
(Diffey, 1987).
Diffey (1987) concluded that the increased risk of
non-melanocytic skin cancer associated with the use of a UVA sunbed
for 10 sessions per year is negligible. However, frequent use of a
sunbed, for example once per week from the age of 20 years will
result in an estimated doubling of the risk of non-melanocytic skin
cancer by age 45 years.
Several studies have reported increased risks of cutaneous
melanoma in users of sunlamps and sunbeds (IARC 1992).
12.2.4 Sunbathing
A significant contribution to the risk of non-melanocytic skin
cancer that results from sunbathing (Diffey, 1987). In these
calculations it was assumed that the subjects were indoor workers
who began sunbathing at age 20 years, and that their annual exposure
up to age 16 years is one half that of an outdoor worker. An indoor
worker who does not sunbathe is estimated to have a 2-3% risk of
non-melanoma skin cancer by age 70. Annual two week vacations spent
sunbathing at Mediterranean latitudes (approx. 40°N) will increase
this risk by a factor of about 5. Sunbathing for four weeks annually
is estimated to result in a 10-20 fold increase in cumulative risk
compared with non-sunbathers. It should be stressed that sunbathing
during vacation periods was assumed to take place for nearly the
whole day on every day of vacation. This is unlikely to be the case
for most holiday makers and so the estimated risk factors for most
people taking holidays in a sunny climate are likely to be one-third
to one-half of those given.
12.3 Adventitious Exposures
12.3.1 Outdoor exposures
Studies using personal dosimeters to monitor exposure of
individuals to UV have shown that, on average the
carcinogenic-effective UV exposure (excluding vacation exposure) of
outdoor workers is about 3 times that of indoor workers (Holman et
al., 1983; Larko and Diffey 1983; Schothorst et al., 1985).
Assuming that up to the age of 16, indoor and outdoor workers
receive the same carcinogenic-effective radiant exposure and that
both receive equivalent vacation exposures, the relative risk for
non-melanoma skin cancer for outdoor workers is estimated to be 3.7
times that for indoor workers (Diffey 1987). However, recent
population studies have found only small differences in skin cancer
incidence between outdoor and indoor workers (see chapter 8).
Armstrong & Kricker (1994) have estimated the proportion of
cutaneous malignant melanomas that is caused by sun exposure. The
estimated proportions varied from 0.97 in males and 0.96 in females
in Queensland, Australia, when the incidence on the whole body was
compared with the incidence on unexposed sites, to 0.68 when the
incidence in people born in Australia was compared with that in
migrants to Australia from areas of lower sun exposure. A comparison
of whites and blacks in the US in which the incidence in blacks was
taken as the incidence in unexposed whites, gave estimates of 0.96
in males and 0.92 in females. It was estimated that some 59,000
(65%) of about 92,000 malignant melanomas worldwide in 1985 were
caused by sun exposure.
12.3.2 Artificial sources
General public exposure
Exposure of the general public to potentially hazardous levels
of UV from artificial sources is unlikely. However, an area that
merits note is the increased use of tungsten halogen lamps for
lighting. Very high filament temperatures combined with quartz
envelopes results in the emission of significantly higher levels of
UV compared with conventional incandescent lamps (Césarini & Muel,
1989; McKinlay et al., 1989). For many applications of such lamps
the presence of a glass filter effectively attenuates the
potentially harmful UV. However, for some others, and particularly
when such lamps are incorporated in desk-top luminaires, no
protective filter is present, and significantly high levels of
erythemally effective UV have been measured. However, for lamps of
similar design and having nominally the same power, there is a wide
range of emitted levels of UV, (for example, between 2 and 56 mW
m-2 effective, in the beam of the lamp at 30 cm distance)
Occupational exposure
Sources that emit UV are used for a wide range of applications
in the workplace. High intensity discharge sources should be, and
often are, contained in interlocked enclosures thus obviating
hazardous exposure of people. Hazard evaluation surveys generally
consist of measurements of the effective exposure levels of UV and
comparison with recommended guidelines on limiting exposure. This
approach addresses the adverse acute effects of exposure, but very
few risk assessments have been carried out in relation to chronic
effects. The risk of non-melanocytic skin cancer for any
occupational situation can be calculated using multivariant
analysis, provided the effective exposures related to the occupation
are known. However, although data on levels of exposure exist for
some occupational situations there is generally a paucity of such
data. Where estimates of skin cancer risk have been made, they have
related to specific occupational exposure situations (Diffey 1988,
1989) or broadly with respect to general (fluorescent) lighting
(Lytle et al., 1993).
Examples where occupational exposure to UV from artificial
sources may occur include; electric arc plasma welding, the drying
and curing of inks, resins, plastics and paints; printing, graphics
arts, copying (using photographic processes) and photography,
photoetching, projector lamps operation, medicine, scientific
laboratory work, tanning and salon work and UV associated with
general lighting fluorescent lamps and desk-top and other luminaires
incorporating tungsten halogen lamps.
Welders are likely to be the largest occupational group with
potential exposure to UV (IARC 1992). The UV effective irradiance
levels around an operating welding arc can exceed recommended
exposure guideline levels by several orders of magnitude, and the
occurrence of acute effects of exposure, photokeratitis and
erythema, is common (Eriksen 1987).
Many industrial photoprocesses use high intensity discharge
lamps that emit copious quantities of UV. However, most of these
sources are effectively shielded and interlocked to prevent human
exposure. Some sources emit leakage radiation and, under these
circumstances, a detailed measurement survey is required to assess
the degree of hazard.
Medical physiotherapists involved in the phototherapy of
patients are occupationally exposed to UV and an analysis of
probable risk has been published (Diffey 1988). During a working
life of 40 years, the additional risk of non-melanoma skin cancer
for a member of this group is estimated to be around 25% compared
with that for non-exposed workers.
General lighting
Low pressure mercury vapour fluorescent lamps are ubiquitous in
the workplace and the home and concern has been expressed regarding
the potential role of the UV they emit in the aetiology of malignant
melanoma. Epidemiological data on exposure to general lighting
fluorescent lamps and malignant melanoma are few and inconsistent.
IRPA/INIRC (1991) concluded that UV exposure from indoor fluorescent
lighting should not be considered as a malignant melanoma risk.
The exposure levels associated with the use of general lighting
fluorescent lamps have been measured in a number of studies (Cole
et al., 1986; McKinlay & Whillock, 1987; Muel et al., 1988 and
Lytle et al., 1993). Exposure data vary considerably depending on
the types of lamps examined and the exposure conditions considered.
A recent study (Lytle et al., 1993) serves to illustrate estimates
of the risk associated with long term exposure to unfiltered general
lighting fluorescent lamps used in the United States. It was
estimated that 50 years exposure to typical unfiltered levels of UV
from fluorescent lamps added approximately 4% (1.6 - 12%) to any
risks associated with exposure to solar UV.
13. INTERNATIONAL GUIDELINES ON EXPOSURE TO ULTRAVIOLET RADIATION
A number of national and international organizations have
promulgated guidelines or standards on exposure to UV. Most are
based upon the same basic criteria of ACGIH (1993) and IRPA/INIRC
(1991).
The basic exposure limit (EL) for both general public and
occupational exposure to UV incident on the skin or eye is 30 J
m-2 effective), when the spectral irradiance Elambda at the eye
or skin surface is mathematically weighted with the hazard relative
spectral effectiveness factor Slambda from 180 nm to 400 nm. This
is given as follows:
Eeff = Sigma Elambda Slambda Deltalambda
where:
Eeff = effective irradiance W m-2
Elambda = spectral irradiance from measurements in W m-2
nm-1
Slambda = relative spectral effectiveness factor (unit-less)
Deltalambda = bandwidth of the calculation or measurement in
nm
At 270 nm in the UVC range, Slambda is 1.0, but at 360 nm in
the centre of the UVA range, its value falls to 0.00013, and
continues to fall for longer wavelengths.
For the UVA, the total radiant exposure incident on the
unprotected eye should not exceed 104 J m-2 (1 J cm-2) within
an 8 h period. The total 8 h radiant exposure incident on the
unprotected skin should not exceed the values in table 13.1.
The radiant UV exposure incident upon the unprotected skin or
eye within an 8-hour period should not exceed the values given in
table 13.1. The limits apply to sources whose emissions are measured
with an instrument having a cosine response detector oriented
perpendicular to the most directly exposed surfaces of the body when
assessing skin exposure and along (or parallel to) the line(s) of
sight when assessing ocular exposure. Although no measurement
averaging aperture is recommended, 1 mm is commonly used.
Table 13.1 International UV exposure limits and spectral weighting
factor (IRPA/INIRC, 1991)
Wavelengtha (nm) EL EL Relative Spectral
(J m-2) (mJ cm-2) Effectiveness Slamda
180 2,500 250 0.012
190 1,600 160 0.019
200 1,000 100 0.030
205 590 59 0.051
210 400 40 0.075
215 320 32 0.095
220 250 25 0.120
225 200 20 0.150
230 160 16 0.190
235 130 13 0.240
240 100 20 0.300
245 83 8.3 0.360
250 70 7.0 0.430
254b 60 6.0 0.500
255 58 5.8 0.520
260 46 4.6 0.650
265 37 3.7 0.810
270 30 3.0 1.000
275 31 3.1 0.960
280b 34 3.4 0.880
285 39 3.9 0.770
290 47 4.7 0.640
295 56 5.6 0.540
297b 65 6.5 0.460
300 100 10 0.300
303b 250 25 0.190
305 500 50 0.060
308 1,200 120 0.026
310 2,000 200 0.015
313b 5,000 500 0.006
315 1.0 x 104 1.0 x 103 0.003
316 1.3 x 104 1.3 x 103 0.0024
317 1.5 x 104 1.5 x 103 0.0020
318 1.9 x 104 1.9 x 103 0.0016
319 2.5 x 104 2.5 x 103 0.0012
320 2.9 x 104 2.9 x 103 0.0010
322 4.5 x 104 4.5 x 103 0.00067
323 5.6 x 104 5.6 x 103 0.00054
325 6.0 x 104 6.0 x 103 0.00050
328 6.8 x 104 6.8 x 103 0.00044
330 7.3 x 104 7.3 x 103 0.00041
333 8.1 x 104 8.1 x 103 0.00037
335 8.8 x 104 8.8 x 103 0.00034
340 1.1 x 105 1.1 x 104 0.00028
Table 13.1 (contd).
Wavelengtha (nm) EL EL Relative Spectral
(J m-2) (mJ cm-2) Effectiveness Slamda
345 1.3 x 105 1.3 x 104 0.00024
350 1.5 x 105 1.5 x 104 0.00020
355 1.9 x 105 1.9 x 104 0.00016
360 2.3 x 105 2.3 x 104 0.00013
365b 2.7 x 105 2.7 x 104 0.00011
370 3.2 x 105 3.2 x 104 0.000093
375 3.9 x 105 3.9 x 104 0.000077
380 4.7 x 105 4.7 x 104 0.000064
385 5.7 x 105 5.7 x 104 0.000053
390 6.8 x 105 6.8 x 104 0.000044
395 8.3 x 105 8.3 x 104 0.000036
400 1.0 x 106 1.0 x 105 0.000030
a Wavelengths chosen are representative; other values should be
interpolated at intermediate wavelengths.
b Emission lines of a mercury discharge spectrum.
The permissible exposure duration, tmax, for exposure (in
seconds) to UV is calculated by:
tmax = 30/ Eeff (W m-2)
Examples are provided in table 13.2.
Table 13.2 Limiting UV exposure durations based on exposure limits
(IRPA/INIRC, 1991)
Duration of exposure Effective irradiance
per day Eeff (W m-2) Eeff (µW cm-2)
8 hours 0.001 0.1
4 hours 0.002 0.2
2 hours 0.004 0.4
1 hour 0.008 0.8
30 minutes 0.017 1.7
15 minutes 0.033 3.3
10 minutes 0.05 5
The EL's were developed considering lightly pigmented
populations with greatest sensitivity and predisposition to adverse
health effects from exposure to UV. The limits apply to UV exposure
of the working population, but with some precaution also apply to
the general public. However, some rare, highly photosensitive
individuals exist who may react adversely to exposure at these
levels. These individuals are normally aware of their heightened
sensitivity. Likewise, if individuals are concomitantly exposed to
photosensitizing agents, an enhanced reaction can take place. Many
individuals who are exposed to photosensitizing agents (ingested or
externally applied chemicals, e.g., in cosmetics, foods, drugs,
industrial chemicals, etc.) may not be aware of their heightened
sensitivity. Lightly pigmented individuals conditioned by previous
UV exposure (leading to tanning and hyperplasia) and heavily
pigmented individuals can tolerate skin exposure in excess of the
EL's without erythemal effects. However, repeated tanning may
increase the risk of accelerated skin aging and even skin cancer.
Such risks should be understood prior to the use of UV for medical
phototherapy or cosmetic exposures.
14. PROTECTIVE MEASURES
14.1 Introduction
It is widely accepted by scientific and medical authorities
throughout the world that UV is potentially carcinogenic and capable
of producing other undesirable health effects. It is sensible
therefore to take steps to minimise UV exposure. Many risks in life
are completely beyond our control such as contracting a rare
disease. However, risks to health associated with exposure to UV
from both natural and artificial sources can be substantially
reduced by taking appropriate control measures.
Since UV exposure occurs externally, simple measures can be
taken to reduce the exposures received. A high degree of protection
can be afforded by protective clothing (including hats), UV
protective eyewear (welding helmets, face shields, goggles,
sunglasses, spectacles etc.) and by sunscreens for exposed skin.
However, the degree of protection afforded can be reduced by
ingestion of photosensitizing drugs or photoallergic/phototoxic
reactions produced by chemicals or cosmetics in contact with the
skin. Thus, education is also an important control measure.
14.2 Education
Concern about high incidences of skin cancer and eye damage
have led to national educational campaigns in some countries to
encourage people to protect themselves against excessive UV exposure
from the sun and in the workplace. Educational programmes directed
at both the workforce and the public are intended to create an
awareness of the adverse health effects that can result from
overexposure to UV.
Presently, in several different countries around the world,
daily environmental UV levels are supplied to the general public in
the form of UV indices. Their provision is intended to educate the
public on the basic climatology of UV, increase awareness of the
hazards of UV and provide information necessary to plan protection.
In a report commissioned to investigate Canadian attitudes to
Environment Canada's UV index (Environment Canada, 1993), it was
found that 73% of Canadians were aware of the UV index, 91% believed
UV affected human health, and most important, 59% of respondents who
were aware of the UV index had changed their sun exposure habits.
Different countries have adopted their own national UV indices.
Unfortunately, they are not all compatible. Clearly, international
uniformity would help to prevent confusion.
Educational programmes aim to produce a change in knowledge and
attitudes, then a change in behaviour and eventually a reduction in
the incidence and mortality rates of skin cancer. A survey of
sunscreen use on beaches in Brisbane, Australia found about 70% of
females and males applied sunscreen. Half of the sunscreens provided
the maximum protection (SPF 15+) and almost 90% used a waterproof
formulation. However, the sunscreen was not applied over the entire
body with over half neglecting ears and lower limbs (Pincus et al.,
1991). Survey results show that sunburn is still occurring; in a
randomly selected group of adults in Melbourne, Australia 16%
reported sunburn over a summer weekend (Hill et al., 1992). With
regard to skin cancers it is still too early to evaluate the
effectiveness of current educational/publicity campaigns.
With respect to changes in behaviour, there is good reason to
avoid exposure to the midday sun since it has been estimated that up
to one third of the day's erythemally effective UV is received
within the period one hour before noon to one hour after noon. Shade
is a useful method of protection but its value should not be
over-estimated, as one may be exposed to a quarter or more of the
total solar UV while shaded from direct sunlight, depending on the
prevailing exposure conditions.
14.3 Protection Factors
The concept of a protection factor is useful when attempting to
quantify the UV protection that items such as sunscreens, clothing
and eyewear can provide (Gies et al., 1992). To determine the
protection factor, the following procedure is conducted. An
effective dose (ED) of UV to the unprotected skin or eye is
calculated by summing the incident solar spectral power over the
wavelength range 280 to 400 nm. In order to determine the effective
dose (EDm) for the skin or eye when it is protected, the
calculation is repeated with the spectral transmission of the
protection item as an additional weighting. The protection factor
(PF) is then defined as the ratio of ED to EDm and is given by the
following equation:
ED SigmaElambda.Slambda.Delta lambda
PF =
EDm SigmaElambda.Slambda.Tlambda.Delta lambda
where:
Elambda = spectral irradiance (W m-2 nm-1) at wavelength lambda
Slambda = relative spectral effectiveness
Tlambda = spectral transmission of protective item at
wavelength lambda
Delta lambda = wavelength interval or bandwidth (nm)
lambda = wavelength (nm)
The inclusion of the spectral effectiveness function in the
calculation ensures that sufficient weighting is given to the
biologically effective wavelengths below 315 nm. A description of
how the protection factors for fabrics, eyewear and sunscreens are
determined is given in Roy and Gies (1993) and CIE (1991).
14.4 Clothing
Use of protective clothing provides one of the simplest means
of reducing UV exposure. Hats have been shown to afford protection,
to various degrees, to the forehead, scalp, ears and most of the
neck. Their protective properties have been studied by Diffey and
Cheeseman (1992). In this study hats were classified into four
categories, small brim, medium brim, large brim and peaked cap. The
protection afforded to different anatomical sites on the head and
neck is shown in table 14.1.
Table 14.1 Sun protection for various anatomical sites on the head
and neck provided by different types of hat (Diffey & Cheeseman,
1992).
Typical sun protection factor *
Style of hat Forehead Nose Cheek Chin Back of
neck
Small brim 15 1.5 1 1 1
< 2.5 cm
Medium brim >20 3 2 1 1
2.5-7.5 cm
Large brim >20 7 3 1.2 5
>7.5 cm
Peaked cap >20 5 1.5 1 1
* In this table 'sun protection factor' is defined as the reciprocal
of the fraction of UV exposure recorded relative to that of the
unprotected head.
The degree of protection provided by clothing depends on the
penetration of UV through materials and this can vary considerably.
Fabrics which are visibly opaque tend to be more highly absorbent of
UV, but the structure or weave of a material is the most important
factor in determining its protective value. Colour and thickness are
a poor guide to UV protection. The transmission properties of some
fabrics commonly used in the manufacture of clothing for everyday
wear are given in table 14.2 (Welsh and Diffey 1981). Here the
protection factor is an estimate of the protection afforded against
biologically effective solar radiation. A high protection factor is
associated with a tightly woven material. UV is transmitted and
scattered through the interstices of the material itself rather than
penetrating the fabric.
Gies et al. (1992) have recently extended the concept of
protection factor (PF) to fabrics and proposed the Ultraviolet
Protection Factor (UPF). The UPF scheme, as shown in table 14.3, is
designed to give the general public information on the amount of UV
protection available from fabrics and clothing and is now in use in
Australia. UPF is analogous to SPF and a fabric of UPF 10 would, in
principle, provide a similar level of protection as a sunscreen of
SPF 10.
Table 14.2 UVB transmission properties of common fabrics
(from Welsh & Diffey 1981)
Fabric Structure Colour Thickness % of Protection
(mm) incident factor
UV
transmitted
Nylon-tricel Woven Black 0.1 0.15 750
Nylon-viscose jacquard Woven Black 0.2 0.20 500
Nylon Woven White 0.1 1.7 55
Nylon-terylene mixture Knitted Blue 0.2 11 9
Nylon-acetate jersey Knitted Pink 0.2 24 4
Polyester-slub viscose Woven Pink 0.5 9 14
Polyester-printed lawn Woven Red 0.2 7 11
Polyester-jersey Knitted Fawn 0.3 7 14
Polyester-jersey Knitted Cream 0.4 5 19
Polyester-jersey Knitted Black 0.3 8 12
Polyester-jersey Knitted Orange 0.3 5 23
Polyester-jersey Knitted Turquoise 0.5 16 6
Polyester-jersey Knitted Brown 0.7 1.6 68
Polyester-jersey Knitted Black 0.5 4.4 23
Polyester-brushed jersey Knitted Blue 0.4 5.2 19
Polyester-brushed jersey Knitted Green 0.4 6.0 16
Polyester-bouclette Knitted Orange 0.4 3.0 33
Polyester-bouclette Knitted Purple 0.4 2.0 51
Cotton-needlecord Woven Brown 0.5 < 0.1 > 1000
Cotton-denim Woven Blue 0.5 < 0.1 > 1000
Cotton-printed Woven Brown 0.3 < 0.1 >1000
Cotton-printed Woven Cream 0.3 2.7 36
Wool-jersey Knitted Fawn 0.7 0.7 150
Table 14.3 Summary of the ultraviolet protection
factor scheme for fabrics
Ultraviolet Mean % UV Protection
protection transmission category
factor UPF
UPF 40+ less than 2.5 Maximum protection 40+
UPF 30 to 39 3.3 to 2.5 Very high protection
UPF 20 to 29 5.0 to 3.3 High protection
Whether a material is wet or dry is important in relation to
its UV transmission properties. The spectral transmittances of some
cotton and polyester-cotton samples are shown in figure 14.1.
Measurements show that the variation of UPF for wet and dry fabrics
is consistent for cotton, all examples showing a decrease in UPF
when wet (Gies et al., 1992). The variation in UPFs between wet
and dry was less consistent for polyester- cotton than for the
cotton, some UPFs increasing for wet fabric, while others decreased.
14.5 Sunscreens
Sunscreens are physical and chemical topical preparations which
attenuate the transmission of solar UV into the skin by absorption,
reflection or scattering. Physical sunscreens (sunblocks), for
example zinc oxide, titanium dioxide or red ferric oxide, function
by reflecting and scattering and provide protection against a broad
spectrum of UV and visible wavelengths. They are normally nontoxic
and have few known adverse effects. Sunscreens based on chemical
absorbers contain one or more colourless UV-absorbing ingredients
which generally absorb UVB radiation more strongly that UVA.
Para-aminobenzoic acid (PABA) and its derivatives, salicylates,
cinnamates and camphor derivatives primarily absorb UVB and transmit
UVA; benzophenones essentially absorb UV of wavelengths of less than
360 nm. The use of solely UVA absorbers (di-benzoyl-methane) is
allowed in only certain countries. These chemicals are all based on
benzene (Moseley 1988).
The application of any sunscreen normally changes the spectrum
of UV that reaches the target cells. Although most sunscreens are
designed to attenuate UV, some contain additives such as bergamot
oil (containing 5-methoxypsoralen) to enhance pigmentation and
photoprotection (Young et al., 1991). The role of such
preparations remains controversial.
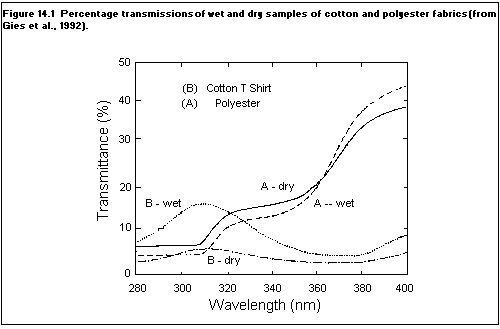 The generally accepted parameter for evaluating the efficacy of
sunscreen preparations is the sun protection factor (SPF), which is
defined as the ratio of the least amount of UV required to produce
minimal erythema after application of a standard quantity of the
sunscreen to the skin to that required to produce the same erythema
without sunscreen application. Several countries have published
recommendations for the efficacy testing of propriety sunscreens,
e.g. the US Food and Drug Administration (1978) and CIE (1991).
Many factors influence SPF values; particularly important are
the spectral power distribution of the source used for SPF testing
and a clear definition of the end-point used for assessment (Urbach,
1989). Variations in these factors can lead to considerable
differences in measured SPF values for the same product.
SPF values generally reflect the degree of protection against
solar UVB radiation, but their protective capacity against UVA must
also be defined. Several in-vivo and in-vitro methods have been
proposed for defining protection against UVA but there is currently
no international consensus on which is the most appropriate.
However, this issue is currently being considered by a technical
committee of the CIE.
Correctly used, sunscreens are effective in preventing sunburn.
Actual SPF values depend critically on the thickness of the
application and on other factors such as absorption into the skin,
sweating and contact with water (for example while swimming).
If sun exposure causes skin cancer, it would be expected that
the use of efficient sunscreens would prevent skin cancer. In animal
experiments, sunscreens with a SPF of as low as 4 have been shown to
be effective in preventing or delaying the onset of UV-induced skin
tumours and cancers (Forbes et al., 1989). There is, at present,
neither direct epidemiological nor laboratory evidence to suggest
that they prevent melanoma or basal cell carcinoma. The few studies
conducted to date suggest either no effect or a causal rather than a
protective effect. For example, Holman et al. (1986) found a
relative risk of melanoma of 1.06 (95% confidence interval
0.71-1.57) with 1-9 years use of sunscreens and 1.15 (95% confidence
interval 0.78-1.68) with 10+ years of use relative to "never use". A
statistically significant positive association of "often or very
often" use of sun protection agents with melanoma (relative risk
1.8, 95% confidence interval 1.5-3.8) was found by Beitner et al.
(1990). In their cohort study of basal cell carcinoma in US nurses,
Hunter et al. (1990) found a higher risk in those who spent 8+
hours per week outside and used sunscreen than in those who spent
the same time outside but did not use sunscreen.
These results cannot be taken at face value. First, there is
likely to be negative confounding of the use of sunscreens with
cutaneous sun sensitivity, people with highly sun sensitive skins
are likely to use sunscreens more than people with less sensitive
skin. Second, there is good evidence that the epidemiological
studies have identified an enthusiastic suntanning population using
sunscreens with minimal SPF to promote their suntan. Third, it is
likely that people adopt or increase their use of sunscreens after
their first skin cancer has been diagnosed, or their attention is
drawn to their personal risk in some other way (for example,
diagnosis of a solar keratosis). Thus present users of sunscreens
are likely to be enriched with people at higher than average risk of
skin cancer. While adjustment for confounding with skin sensitivity
has been carried out in some studies (Holman et al. 1986, Hunter
et al. 1990), this third issue has not been addressed in any
published analysis. Fourth, relatively little information is
available on the mutagenic and carcinogenic potential of various
sunscreens. The US National Cancer Institute (1989) recommended the
following six compounds to be evaluated for chronic testing in
rodents by the US National Toxicology Program: cinoxate;
2-ethylhexyl 2-cyno-3,3-diphenyl-acrylate; 2-ethylhexyl
para-methoxycinnamate; homosalate; methyl anthranilate; and
oxybenzone. Neither epidemiological nor long term mammalian
carcinogenicity data are available on these compounds. The results
of in-vitro studies were assessed as either negative or inconsistent
among systems or among batches of a compound (because of
impurities). 2-ethylhexyl para-methoxycinnamate was implicated as a
potential tumour initiator in one study in which hairless mice were
painted with the compound over a nine-week period and subsequently
treated with the tumour promoter, croton oil (Gallagher et al.,
1984a). Subsequent work by Reeve et al. (1985), however, failed to
confirm these results, and Forbes et al. (1989) found no evidence
of tumour initiation by the compound in an initiation-promotion
experiment in mice.
Sunscreens have been strongly promoted by the medical community
such as in the United States, Canada, Australia, and Scandinavian
countries. However, incidence rates of melanoma have risen steeply
in recent decades, even after the introduction of sunscreens (Lee
1989, Jensen and Bolander 1980, Magnus 1977, 1986; Gallagher et
al., 1986). The use of high SPF sunscreens is recommended by WHO
and by the International Union Against Cancer (UICC) as a supportive
part of a sun avoidance programme rather than its main thrust (Marks
& Hill, 1992).
Both UVA and UVB have been shown to mutate DNA and cause skin
cancers in animals (Staberg et al., 1983). UVA penetrates deeper
into the skin than UVB and because of the energy distribution of
sunlight and filtering by the outermost layers of the skin,
melanocytes receive up to 70 photons of UVA for every photon of UVB.
Sunscreens effectively block solar UVB. UVB is the normal stimulus
for accommodation of the skin such as thickening and increased
pigmentation.
Sunscreens suppress normal warnings of overexposure such as
erythema and sunburn and allow excessive exposure to wavelengths of
sunlight they do not block. Due to lack of these natural signs
sunscreens create a false sense of security and individuals tend to
stay in sun longer. In view of these behavioural changes which
increases individuals UVA exposure it has been suggested that,
because of the rising incidence of melanoma, UVA may be associated
with its occurrence (Garland et al., 1992). While a recent study
(Setlow et al., 1993) in fish reported melanoma induction by UVA,
the role of UVA in the causation of human malignant melanoma has yet
to be established.
Trans-urocanic acid, a natural compound of the stratum corneum
which absorbs UVB and is used as an additive in some commercial
sunscreen products, increased the yield of solar-simulated UV
induced tumours in hairless mice (Reeve et al., 1989). The
significance of this finding for human exposure has not been
evaluated.
Reports on sunscreen protection against UV induced
immunosuppression have been equivocal. Fisher et al. (1989)
reported no protection of hairless mice against UV-induced systemic
immunosuppression following application of Padimate 0 (a PABA ester
and UVB absorber) or oxybenzone (a UVB/UVA absorber) with sun
protection factors (SPF) of 6 or 15. Similarly SPF 15
octy-N-dimethyl-p-aminobenzoate (o-PABA) had no effect on UV-induced
systemic immunosuppression; nor was susceptibility to UV-induced
tumours altered in hairless mice (Reeve et al., 1991). In this
study however, 2-ethylhexyl-p-methoxycinnamate (2-EHMC, SPF-15) was
fully protective. Morison (1984) also reported protection of mice
with PABA against both immunosuppression and susceptibility to UV-
induced tumours. In recent studies (Wolf et al., 1993a, b),
sunscreens with SPFs of 4-6, including o-PABA, 2-EHMC, and
benzophenone all provided protection of mice against UV- induced
local and systemic immunosuppression, although they were less
effective in preventing systemic effects. Finally, in a human study,
a sunscreen containing 8% octyl dimethyl PABA, 2% 2-hydroxy-
4methoxybenzophenone, and 2% methoxydibenzoyl methane (SPF 15) did
not protect against effects of solarium exposure on NK activity,
recall antigen skin tests and immunoglobulin production in vitro
in mitogen stimulated cultures (Hersey et al., 1987).
While further studies are still needed to clarify concerns
raised about the ingredients and protectiveness of sunscreens, broad
spectrum sunscreens which absorb both UVA and UVB with an SPF of at
least 15 are still recommended as an effective means of personal
protection against UV exposure.
14.6 Tanning Devices
Our modern lifestyle has suggested that having a tan is
synonymous with good health. The increasing popularity of this
symbol of health status and the inability to suntan during
non-summer months, has led to the growth of an artificial tanning
industry using sunlamps or sunbeds (combination of fluorescent
lamp-shaped sunlamps into a bed). The dangers of excessive exposure
to UV have been described earlier in this document. They range from
mild erythema to severe burns of the skin from acute exposure, to
skin cancer and skin ageing from long-term exposure. When eyes are
exposed, damage can occur to the cornea, lens and retina, depending
on the UV wavelength.
The first generation of sunlamps emitted primarily UVB and, if
used correctly, were efficient tanning agents. Unfortunately, they
also tended to cause a painful 'sunburn' and other undesirable
side-effects. UVA lamps are now used and these, it is claimed, tan
safely without burning. However, as the carcinogenesis action
spectrum extends into the longer UVA spectral region, exposure to
these lamps is not without risk.
Recommendations regarding the use of sunbeds
Following a thorough review of this topic, the IRPA/INIRC
(1991a) issued recommendations on the use of sunlamps or sunbeds for
cosmetic purposes as follows:
General
The use of sunbeds for cosmetic purposes is not recommended.
Specific
(1) People with skin types I and II should not use sunbeds. They
are likely to be disappointed with the results of the
exposures, they have higher susceptibility to sunburn and have
a higher risk of developing skin cancer.
(2) Any person with a large number of nevi (moles), a tendency to
freckle, a history of severe sunburn especially in childhood,
or a family history of malignant melanoma should not use a
sunbed.
(3) Any person taking a medicine that is known to be photoactive
should not use a sunbed. If in doubt, they should seek the
advice of a physician.
(4) Any person who already has extensive skin "sunlight" damage, or
who has had premalignant or malignant skin lesions, should not
use sunbeds.
(5) Any person who has a skin disease should seek the advice of a
physician before using a sunbed.
(6) Children should not use sunbeds.
(7) Sunbeds should not be used if perfumes, body lotions or sprays
have been applied that day.
(8) Because the sensitivities of individuals vary greatly, it is
advisable to limit the duration of the first session to about
one-half of a regular session in order to establish the user's
skin response. If following the first session any adverse
reaction occurs, further use of the sunbed is not recommended.
(9) Regular exposure should not exceed two sessions per week with a
maximum of 30 sessions per year or 30 minimum erythemal doses
(MEDs) per year, whichever is the smaller erythemally effective
exposure. An occasional break from the regularity of exposure
is advisable.
(10) With respect to recommendation (9), the manufacturer of the
sunbed should supply a schedule of exposure and recommended
maximum exposure durations based on the emission
characteristics of the sunbed.
(11) Appropriate protective eyewear should be provided by the
manufacturer and should always be worn when using a sunbed.
(12) When the sunbed is being provided for use by a commercial
operator, it is the responsibility of the operator to provide
the person intending the use the sunbed with the appropriate
information as summarized in recommendations (1) to (11) above.
14.7 Occupational Protection
Occupational exposure to UV should be kept to a minimum. Some
UV sources emit a considerable amount of visible radiation, and in
this case, the natural aversion response is evoked, so there is
little chance of accidental over-exposure of the eyes. On the other
hand, artificial sources emitting short wavelength UV radiation
exist where accidental exposure is quite likely. While working
outdoors, skin and eye protection should be used. Exposure outdoors
during the periods of 2 hours either side of noon should be avoided.
Where UV levels, compared with the ELs and erythemal dose, are
such as to constitute a hazard, protection against hazardous
exposure may be achieved by a combination of engineering control
measures; administrative control measures and personal protection.
For artificial sources, wherever possible, priority should be given
to engineering and administrative controls to reduce the requirement
for personal protection.
Engineering controls
The principal and most effective engineering control measures
are those intended to contain the radiation. Wherever possible UV
from artificial sources should be contained within a sealed housing.
If observation windows are required they should be made of suitably
absorbent materials such as certain grades of acrylic and window
glass. Where the exposure process is required to take place external
to the source housing a screened area should be set aside where it
may be carried out; an example of this is arc welding where screens
must be provided to prevent exposure of people not involved with the
welding process. Any such screened area should be subject to
administrative control measures and persons working in the area
should be adequately protected from UV as described below. Where a
source of UV is normally enclosed during use but to which access is
required, for example for maintenance, the housing should be fitted
with safety interlocks. If anyone needs to gain access to the source
while it is energized, the interlock should immediately switch off
the power to the lamps and should not be able to be re-energized
until the interlocks are re-engaged Many such sources are used for a
variety of drying and curing purposes, particularly in the printing
industry and all should be subject to strict engineering controls.
Administrative control measures
The principal administrative control measures are those that
limit access to the source and provide information directed at
making people aware of potential hazards associated with it. Access
to an area where equipment emits UV should be limited to those
persons directly concerned with its use. All persons concerned with
the use of such equipment should be made aware of this and should be
informed of the potential hazards. Appropriate hazard warning signs
should be used to indicate the presence of UV and whenever possible
warning lights may be used to show that equipment is energized. The
user of a UV emitting source should keep as far from the source as
possible. At large distances (greater than twice the greatest
dimensions of the source) the irradiance (W m-2) falls off as the
square of the distance from the source. Closer to the source
irradiance falls off approximately linearly with distance. Exposures
should be kept to a minimum and the EL's recommended by IRPA/INIRC
(1991) given in chapter 13 should not be exceeded. Particular care
should be taken to prevent exposure of persons taking
photosensitising medications or concomitantly exposed to
photosensitisers in the environment.
Personal protection
The most effective way to protect the skin from UV is to cover
it. In indoor occupational situations, the areas of the body most at
risk are the face (and eyes) and neck, the forearms and the backs of
the hands. The face can be protected by a shield and this should
also provide eye protection. The arms should be covered by clothing
with a low UVB transmission; in general materials that are visibly
opaque are suitable (see section 14.4). Hands can be protected by
wearing gloves. Face shields, goggles or safety spectacles which
absorb UV should be worn where there is a potential eye hazard.
A range of suitable protective eyewear is commercially
available (see figure 14.2). Welders should be protected by a helmet
fitted with appropriate absorption filters. Some high pressure lamps
are potential explosion hazards and the eyes and face should be
protected against flying fragments of glass. Particular care should
be taken to protect the eyes, face and hands when such lamps are
being removed or replaced. The risk to outdoor workers such as
agricultural workers, labourers, construction workers, fishermen etc
from solar radiation exposure can be minimized by wearing
appropriate tightly woven clothing, and most importantly a brimmed
hat to reduce face and neck exposure. Sunscreens can be applied to
exposed skin to reduce exposure further (see section 14.5).
Hazards from ozone
The absorption of short wavelength UV by oxygen in the air
forms ozone, a powerful oxidising agent. The American Conference of
Governmental Hygienists has published a threshold limit value for
ozone exposure, 0.1 ppm, (ACGIH, 1993), and concentrations above
this value should be avoided.
Levels of ozone may be reduced by providing adequate
ventilation in the area in which the source is located. Very intense
sources emitting short wavelength UVB, for example, high pressure
linear and compact mercury and xenon lamps will normally require an
extraction system to remove ozone.
14.8 Protection in Medicine and Dentistry
Protection of both patients and staff must be considered. When
a patient is being exposed to UV for clinical purposes, sites not
intended to be treated should be covered and the eyes protected.
In medical care, chlorpromazine and thioridazine are
phenothiazine derivatives which are widely used as psychosedatives.
As well as producing photosensitivity as a side-effect in some
patients, these drugs may induce a similar type of reaction in
hospital staff. Contact dermatitis may occur in staff handling
phenothiazines or fouled laundry since metabolites of chlorpromazine
are phototoxic (Moseley 1988). To avoid contamination, staff should
wear protective clothing.
Problems may also occur in dentistry, both for patients and
staff. The mouth is lined with a relatively thin squamous epithelium
and so there may be considerable penetration of UV to underlying
cells. Patients and staff with systemic lupus erythematous (SLE)
have been reported to be at risk from lights used in dentistry
(Moseley 1988). Both operating lamps and sources used to polymerise
resins have been found to cause damage. In one case, a dentist, who
was a known SLE sufferer, experienced a facial eruption after using
a visible-light resin-curing source on a patient. Evidently, special
care is required when lights are used on patients and a known
history of SLE would be contraindicated for UV or visible light
polymerisation.
14.9 Nutrition
Nutrition can provide the body with essential antioxidants and
these molecules are distributed throughout the body. At the cellular
level, they enter a number of endogenous photoprotective systems to
control photochemical processes (Roberts et al., 1991). Quenchers
which can negate specific reactive intermediates may be important as
a defence mechanism against UV insult to the eye. Glutathione, due
to the low energy of the SH (thiol) bond (65 kcal) is an efficient
free-radical scavenger and singlet oxygen quencher. Ascorbic acid
quenches free radical and superoxide reactions. alpha-Tocopherol
quenches both singlet oxygen and free radicals. There are also
various antioxidant enzymes present in the eye. Exogenous scavengers
and quenchers may be able to prevent UV damage by interrupting
transient intermediates which cause ocular damage. An approach is to
increase the known endogenous quenchers, (antioxidants) ascorbic
acid, alpha-tocopherol and ß-carotene in the diet (Roberts et al.,
1991).
14.10 Additional Protective Agents
Although it has been known that glutathione is a particularly
effective quencher of excited state transients in the eye, until now
there has been no success in increasing this endogenous sulphydryl
compound in ocular tissues. There have recently been found (Roberts
et al., 1991) a promising group of compounds, phosphorylated
sulphydryls, which pass the blood retinal and blood ocular barrier,
and appear to mimic the protective effect of the endogenous thiol
glutathione. This offers a possible way of protecting the eye and
other tissues from UV induced damage.
Protection from UV damage was reported in mice maintained on a
diet supplemented with either carnosine (ß alanylhistidine), an
antioxidant known to have immunopotentiating properties (Reeve et
al., 1993) or retinal palmitate in combination with canthaxanthin,
a carotenoid (Gensler, 1989). Some protection was also afforded by
retinal palmitate alone. Protection has been shown in healthy humans
receiving a daily supplement of ß-carotene (Fuller et al., 1992).
14.11 Eye Protection
In industry there are many sources capable of causing acute eye
injury within a short exposure time, while in the natural
environment acute injury is likely to occur mostly in situations
where solar UV is reflected onto the eye, such as from snow while
skiing. A variety of eye protection is available with various
degrees of protection appropriate to their intended use (see figure
14.2). Those intended for industrial use include, welding helmets
(additionally providing protection from intense visible and infrared
radiation and face protection), face shields, goggles and UV
absorbing spectacles. For use in the outdoor environment, they
include (ski) goggles for extreme exposure conditions and
sunglasses.
The appropriateness and selection of protective eyewear is
dependent on the:
(1) intensity and spectral emission characteristics of the UV
source,
(2) behavioural pattern of people near UV sources (distance and
time are important),
(3) transmission properties of the protective eyewear material, and
(4) design of the frame of the eyewear to prevent exposure of the
eye from direct unabsorbed UV.
In industrial exposure situations the degree of ocular hazard
can be assessed by measurement and comparison with recommended
limits for exposure (IRPA/INIRC, 1991) (see Chapter 13). Welders and
nearby workers should routinely wear appropriate eye and face
protection. For protection against other less intense sources of UV
in the workplace tightly fitting goggles or spectacles with side
shields may be appropriate, but consideration should also be given
to the need for additional face protection. In general, protective
eyewear provided for industrial use should fit snugly to the face,
thus ensuring that there are no gaps through which UV can directly
reach the eye and should have adequate mechanical construction to
prevent physical injury.
For outdoor workers and the general public, the most hazardous
source of UV exposure is the sun. Adequately designed (ski) goggles
afford protection against exposure to solar UV at high altitudes and
on snow, but for most other exposure conditions, good UV absorbing
sunglasses are an adequate means of eye protection.
Transmission of UV through sunglasses varies considerably
(Wester 1987, Gies et al., 1992), yet consumers are provided with
little information about the protection afforded by them. Some
countries have drafted standards limiting UV transmission through
sunglasses and Gies et al. (1990a) have proposed a UV eye
protection factor (EPF) for sunglasses similar to the one developed
for fabrics and sunscreens.
The generally accepted parameter for evaluating the efficacy of
sunscreen preparations is the sun protection factor (SPF), which is
defined as the ratio of the least amount of UV required to produce
minimal erythema after application of a standard quantity of the
sunscreen to the skin to that required to produce the same erythema
without sunscreen application. Several countries have published
recommendations for the efficacy testing of propriety sunscreens,
e.g. the US Food and Drug Administration (1978) and CIE (1991).
Many factors influence SPF values; particularly important are
the spectral power distribution of the source used for SPF testing
and a clear definition of the end-point used for assessment (Urbach,
1989). Variations in these factors can lead to considerable
differences in measured SPF values for the same product.
SPF values generally reflect the degree of protection against
solar UVB radiation, but their protective capacity against UVA must
also be defined. Several in-vivo and in-vitro methods have been
proposed for defining protection against UVA but there is currently
no international consensus on which is the most appropriate.
However, this issue is currently being considered by a technical
committee of the CIE.
Correctly used, sunscreens are effective in preventing sunburn.
Actual SPF values depend critically on the thickness of the
application and on other factors such as absorption into the skin,
sweating and contact with water (for example while swimming).
If sun exposure causes skin cancer, it would be expected that
the use of efficient sunscreens would prevent skin cancer. In animal
experiments, sunscreens with a SPF of as low as 4 have been shown to
be effective in preventing or delaying the onset of UV-induced skin
tumours and cancers (Forbes et al., 1989). There is, at present,
neither direct epidemiological nor laboratory evidence to suggest
that they prevent melanoma or basal cell carcinoma. The few studies
conducted to date suggest either no effect or a causal rather than a
protective effect. For example, Holman et al. (1986) found a
relative risk of melanoma of 1.06 (95% confidence interval
0.71-1.57) with 1-9 years use of sunscreens and 1.15 (95% confidence
interval 0.78-1.68) with 10+ years of use relative to "never use". A
statistically significant positive association of "often or very
often" use of sun protection agents with melanoma (relative risk
1.8, 95% confidence interval 1.5-3.8) was found by Beitner et al.
(1990). In their cohort study of basal cell carcinoma in US nurses,
Hunter et al. (1990) found a higher risk in those who spent 8+
hours per week outside and used sunscreen than in those who spent
the same time outside but did not use sunscreen.
These results cannot be taken at face value. First, there is
likely to be negative confounding of the use of sunscreens with
cutaneous sun sensitivity, people with highly sun sensitive skins
are likely to use sunscreens more than people with less sensitive
skin. Second, there is good evidence that the epidemiological
studies have identified an enthusiastic suntanning population using
sunscreens with minimal SPF to promote their suntan. Third, it is
likely that people adopt or increase their use of sunscreens after
their first skin cancer has been diagnosed, or their attention is
drawn to their personal risk in some other way (for example,
diagnosis of a solar keratosis). Thus present users of sunscreens
are likely to be enriched with people at higher than average risk of
skin cancer. While adjustment for confounding with skin sensitivity
has been carried out in some studies (Holman et al. 1986, Hunter
et al. 1990), this third issue has not been addressed in any
published analysis. Fourth, relatively little information is
available on the mutagenic and carcinogenic potential of various
sunscreens. The US National Cancer Institute (1989) recommended the
following six compounds to be evaluated for chronic testing in
rodents by the US National Toxicology Program: cinoxate;
2-ethylhexyl 2-cyno-3,3-diphenyl-acrylate; 2-ethylhexyl
para-methoxycinnamate; homosalate; methyl anthranilate; and
oxybenzone. Neither epidemiological nor long term mammalian
carcinogenicity data are available on these compounds. The results
of in-vitro studies were assessed as either negative or inconsistent
among systems or among batches of a compound (because of
impurities). 2-ethylhexyl para-methoxycinnamate was implicated as a
potential tumour initiator in one study in which hairless mice were
painted with the compound over a nine-week period and subsequently
treated with the tumour promoter, croton oil (Gallagher et al.,
1984a). Subsequent work by Reeve et al. (1985), however, failed to
confirm these results, and Forbes et al. (1989) found no evidence
of tumour initiation by the compound in an initiation-promotion
experiment in mice.
Sunscreens have been strongly promoted by the medical community
such as in the United States, Canada, Australia, and Scandinavian
countries. However, incidence rates of melanoma have risen steeply
in recent decades, even after the introduction of sunscreens (Lee
1989, Jensen and Bolander 1980, Magnus 1977, 1986; Gallagher et
al., 1986). The use of high SPF sunscreens is recommended by WHO
and by the International Union Against Cancer (UICC) as a supportive
part of a sun avoidance programme rather than its main thrust (Marks
& Hill, 1992).
Both UVA and UVB have been shown to mutate DNA and cause skin
cancers in animals (Staberg et al., 1983). UVA penetrates deeper
into the skin than UVB and because of the energy distribution of
sunlight and filtering by the outermost layers of the skin,
melanocytes receive up to 70 photons of UVA for every photon of UVB.
Sunscreens effectively block solar UVB. UVB is the normal stimulus
for accommodation of the skin such as thickening and increased
pigmentation.
Sunscreens suppress normal warnings of overexposure such as
erythema and sunburn and allow excessive exposure to wavelengths of
sunlight they do not block. Due to lack of these natural signs
sunscreens create a false sense of security and individuals tend to
stay in sun longer. In view of these behavioural changes which
increases individuals UVA exposure it has been suggested that,
because of the rising incidence of melanoma, UVA may be associated
with its occurrence (Garland et al., 1992). While a recent study
(Setlow et al., 1993) in fish reported melanoma induction by UVA,
the role of UVA in the causation of human malignant melanoma has yet
to be established.
Trans-urocanic acid, a natural compound of the stratum corneum
which absorbs UVB and is used as an additive in some commercial
sunscreen products, increased the yield of solar-simulated UV
induced tumours in hairless mice (Reeve et al., 1989). The
significance of this finding for human exposure has not been
evaluated.
Reports on sunscreen protection against UV induced
immunosuppression have been equivocal. Fisher et al. (1989)
reported no protection of hairless mice against UV-induced systemic
immunosuppression following application of Padimate 0 (a PABA ester
and UVB absorber) or oxybenzone (a UVB/UVA absorber) with sun
protection factors (SPF) of 6 or 15. Similarly SPF 15
octy-N-dimethyl-p-aminobenzoate (o-PABA) had no effect on UV-induced
systemic immunosuppression; nor was susceptibility to UV-induced
tumours altered in hairless mice (Reeve et al., 1991). In this
study however, 2-ethylhexyl-p-methoxycinnamate (2-EHMC, SPF-15) was
fully protective. Morison (1984) also reported protection of mice
with PABA against both immunosuppression and susceptibility to UV-
induced tumours. In recent studies (Wolf et al., 1993a, b),
sunscreens with SPFs of 4-6, including o-PABA, 2-EHMC, and
benzophenone all provided protection of mice against UV- induced
local and systemic immunosuppression, although they were less
effective in preventing systemic effects. Finally, in a human study,
a sunscreen containing 8% octyl dimethyl PABA, 2% 2-hydroxy-
4methoxybenzophenone, and 2% methoxydibenzoyl methane (SPF 15) did
not protect against effects of solarium exposure on NK activity,
recall antigen skin tests and immunoglobulin production in vitro
in mitogen stimulated cultures (Hersey et al., 1987).
While further studies are still needed to clarify concerns
raised about the ingredients and protectiveness of sunscreens, broad
spectrum sunscreens which absorb both UVA and UVB with an SPF of at
least 15 are still recommended as an effective means of personal
protection against UV exposure.
14.6 Tanning Devices
Our modern lifestyle has suggested that having a tan is
synonymous with good health. The increasing popularity of this
symbol of health status and the inability to suntan during
non-summer months, has led to the growth of an artificial tanning
industry using sunlamps or sunbeds (combination of fluorescent
lamp-shaped sunlamps into a bed). The dangers of excessive exposure
to UV have been described earlier in this document. They range from
mild erythema to severe burns of the skin from acute exposure, to
skin cancer and skin ageing from long-term exposure. When eyes are
exposed, damage can occur to the cornea, lens and retina, depending
on the UV wavelength.
The first generation of sunlamps emitted primarily UVB and, if
used correctly, were efficient tanning agents. Unfortunately, they
also tended to cause a painful 'sunburn' and other undesirable
side-effects. UVA lamps are now used and these, it is claimed, tan
safely without burning. However, as the carcinogenesis action
spectrum extends into the longer UVA spectral region, exposure to
these lamps is not without risk.
Recommendations regarding the use of sunbeds
Following a thorough review of this topic, the IRPA/INIRC
(1991a) issued recommendations on the use of sunlamps or sunbeds for
cosmetic purposes as follows:
General
The use of sunbeds for cosmetic purposes is not recommended.
Specific
(1) People with skin types I and II should not use sunbeds. They
are likely to be disappointed with the results of the
exposures, they have higher susceptibility to sunburn and have
a higher risk of developing skin cancer.
(2) Any person with a large number of nevi (moles), a tendency to
freckle, a history of severe sunburn especially in childhood,
or a family history of malignant melanoma should not use a
sunbed.
(3) Any person taking a medicine that is known to be photoactive
should not use a sunbed. If in doubt, they should seek the
advice of a physician.
(4) Any person who already has extensive skin "sunlight" damage, or
who has had premalignant or malignant skin lesions, should not
use sunbeds.
(5) Any person who has a skin disease should seek the advice of a
physician before using a sunbed.
(6) Children should not use sunbeds.
(7) Sunbeds should not be used if perfumes, body lotions or sprays
have been applied that day.
(8) Because the sensitivities of individuals vary greatly, it is
advisable to limit the duration of the first session to about
one-half of a regular session in order to establish the user's
skin response. If following the first session any adverse
reaction occurs, further use of the sunbed is not recommended.
(9) Regular exposure should not exceed two sessions per week with a
maximum of 30 sessions per year or 30 minimum erythemal doses
(MEDs) per year, whichever is the smaller erythemally effective
exposure. An occasional break from the regularity of exposure
is advisable.
(10) With respect to recommendation (9), the manufacturer of the
sunbed should supply a schedule of exposure and recommended
maximum exposure durations based on the emission
characteristics of the sunbed.
(11) Appropriate protective eyewear should be provided by the
manufacturer and should always be worn when using a sunbed.
(12) When the sunbed is being provided for use by a commercial
operator, it is the responsibility of the operator to provide
the person intending the use the sunbed with the appropriate
information as summarized in recommendations (1) to (11) above.
14.7 Occupational Protection
Occupational exposure to UV should be kept to a minimum. Some
UV sources emit a considerable amount of visible radiation, and in
this case, the natural aversion response is evoked, so there is
little chance of accidental over-exposure of the eyes. On the other
hand, artificial sources emitting short wavelength UV radiation
exist where accidental exposure is quite likely. While working
outdoors, skin and eye protection should be used. Exposure outdoors
during the periods of 2 hours either side of noon should be avoided.
Where UV levels, compared with the ELs and erythemal dose, are
such as to constitute a hazard, protection against hazardous
exposure may be achieved by a combination of engineering control
measures; administrative control measures and personal protection.
For artificial sources, wherever possible, priority should be given
to engineering and administrative controls to reduce the requirement
for personal protection.
Engineering controls
The principal and most effective engineering control measures
are those intended to contain the radiation. Wherever possible UV
from artificial sources should be contained within a sealed housing.
If observation windows are required they should be made of suitably
absorbent materials such as certain grades of acrylic and window
glass. Where the exposure process is required to take place external
to the source housing a screened area should be set aside where it
may be carried out; an example of this is arc welding where screens
must be provided to prevent exposure of people not involved with the
welding process. Any such screened area should be subject to
administrative control measures and persons working in the area
should be adequately protected from UV as described below. Where a
source of UV is normally enclosed during use but to which access is
required, for example for maintenance, the housing should be fitted
with safety interlocks. If anyone needs to gain access to the source
while it is energized, the interlock should immediately switch off
the power to the lamps and should not be able to be re-energized
until the interlocks are re-engaged Many such sources are used for a
variety of drying and curing purposes, particularly in the printing
industry and all should be subject to strict engineering controls.
Administrative control measures
The principal administrative control measures are those that
limit access to the source and provide information directed at
making people aware of potential hazards associated with it. Access
to an area where equipment emits UV should be limited to those
persons directly concerned with its use. All persons concerned with
the use of such equipment should be made aware of this and should be
informed of the potential hazards. Appropriate hazard warning signs
should be used to indicate the presence of UV and whenever possible
warning lights may be used to show that equipment is energized. The
user of a UV emitting source should keep as far from the source as
possible. At large distances (greater than twice the greatest
dimensions of the source) the irradiance (W m-2) falls off as the
square of the distance from the source. Closer to the source
irradiance falls off approximately linearly with distance. Exposures
should be kept to a minimum and the EL's recommended by IRPA/INIRC
(1991) given in chapter 13 should not be exceeded. Particular care
should be taken to prevent exposure of persons taking
photosensitising medications or concomitantly exposed to
photosensitisers in the environment.
Personal protection
The most effective way to protect the skin from UV is to cover
it. In indoor occupational situations, the areas of the body most at
risk are the face (and eyes) and neck, the forearms and the backs of
the hands. The face can be protected by a shield and this should
also provide eye protection. The arms should be covered by clothing
with a low UVB transmission; in general materials that are visibly
opaque are suitable (see section 14.4). Hands can be protected by
wearing gloves. Face shields, goggles or safety spectacles which
absorb UV should be worn where there is a potential eye hazard.
A range of suitable protective eyewear is commercially
available (see figure 14.2). Welders should be protected by a helmet
fitted with appropriate absorption filters. Some high pressure lamps
are potential explosion hazards and the eyes and face should be
protected against flying fragments of glass. Particular care should
be taken to protect the eyes, face and hands when such lamps are
being removed or replaced. The risk to outdoor workers such as
agricultural workers, labourers, construction workers, fishermen etc
from solar radiation exposure can be minimized by wearing
appropriate tightly woven clothing, and most importantly a brimmed
hat to reduce face and neck exposure. Sunscreens can be applied to
exposed skin to reduce exposure further (see section 14.5).
Hazards from ozone
The absorption of short wavelength UV by oxygen in the air
forms ozone, a powerful oxidising agent. The American Conference of
Governmental Hygienists has published a threshold limit value for
ozone exposure, 0.1 ppm, (ACGIH, 1993), and concentrations above
this value should be avoided.
Levels of ozone may be reduced by providing adequate
ventilation in the area in which the source is located. Very intense
sources emitting short wavelength UVB, for example, high pressure
linear and compact mercury and xenon lamps will normally require an
extraction system to remove ozone.
14.8 Protection in Medicine and Dentistry
Protection of both patients and staff must be considered. When
a patient is being exposed to UV for clinical purposes, sites not
intended to be treated should be covered and the eyes protected.
In medical care, chlorpromazine and thioridazine are
phenothiazine derivatives which are widely used as psychosedatives.
As well as producing photosensitivity as a side-effect in some
patients, these drugs may induce a similar type of reaction in
hospital staff. Contact dermatitis may occur in staff handling
phenothiazines or fouled laundry since metabolites of chlorpromazine
are phototoxic (Moseley 1988). To avoid contamination, staff should
wear protective clothing.
Problems may also occur in dentistry, both for patients and
staff. The mouth is lined with a relatively thin squamous epithelium
and so there may be considerable penetration of UV to underlying
cells. Patients and staff with systemic lupus erythematous (SLE)
have been reported to be at risk from lights used in dentistry
(Moseley 1988). Both operating lamps and sources used to polymerise
resins have been found to cause damage. In one case, a dentist, who
was a known SLE sufferer, experienced a facial eruption after using
a visible-light resin-curing source on a patient. Evidently, special
care is required when lights are used on patients and a known
history of SLE would be contraindicated for UV or visible light
polymerisation.
14.9 Nutrition
Nutrition can provide the body with essential antioxidants and
these molecules are distributed throughout the body. At the cellular
level, they enter a number of endogenous photoprotective systems to
control photochemical processes (Roberts et al., 1991). Quenchers
which can negate specific reactive intermediates may be important as
a defence mechanism against UV insult to the eye. Glutathione, due
to the low energy of the SH (thiol) bond (65 kcal) is an efficient
free-radical scavenger and singlet oxygen quencher. Ascorbic acid
quenches free radical and superoxide reactions. alpha-Tocopherol
quenches both singlet oxygen and free radicals. There are also
various antioxidant enzymes present in the eye. Exogenous scavengers
and quenchers may be able to prevent UV damage by interrupting
transient intermediates which cause ocular damage. An approach is to
increase the known endogenous quenchers, (antioxidants) ascorbic
acid, alpha-tocopherol and ß-carotene in the diet (Roberts et al.,
1991).
14.10 Additional Protective Agents
Although it has been known that glutathione is a particularly
effective quencher of excited state transients in the eye, until now
there has been no success in increasing this endogenous sulphydryl
compound in ocular tissues. There have recently been found (Roberts
et al., 1991) a promising group of compounds, phosphorylated
sulphydryls, which pass the blood retinal and blood ocular barrier,
and appear to mimic the protective effect of the endogenous thiol
glutathione. This offers a possible way of protecting the eye and
other tissues from UV induced damage.
Protection from UV damage was reported in mice maintained on a
diet supplemented with either carnosine (ß alanylhistidine), an
antioxidant known to have immunopotentiating properties (Reeve et
al., 1993) or retinal palmitate in combination with canthaxanthin,
a carotenoid (Gensler, 1989). Some protection was also afforded by
retinal palmitate alone. Protection has been shown in healthy humans
receiving a daily supplement of ß-carotene (Fuller et al., 1992).
14.11 Eye Protection
In industry there are many sources capable of causing acute eye
injury within a short exposure time, while in the natural
environment acute injury is likely to occur mostly in situations
where solar UV is reflected onto the eye, such as from snow while
skiing. A variety of eye protection is available with various
degrees of protection appropriate to their intended use (see figure
14.2). Those intended for industrial use include, welding helmets
(additionally providing protection from intense visible and infrared
radiation and face protection), face shields, goggles and UV
absorbing spectacles. For use in the outdoor environment, they
include (ski) goggles for extreme exposure conditions and
sunglasses.
The appropriateness and selection of protective eyewear is
dependent on the:
(1) intensity and spectral emission characteristics of the UV
source,
(2) behavioural pattern of people near UV sources (distance and
time are important),
(3) transmission properties of the protective eyewear material, and
(4) design of the frame of the eyewear to prevent exposure of the
eye from direct unabsorbed UV.
In industrial exposure situations the degree of ocular hazard
can be assessed by measurement and comparison with recommended
limits for exposure (IRPA/INIRC, 1991) (see Chapter 13). Welders and
nearby workers should routinely wear appropriate eye and face
protection. For protection against other less intense sources of UV
in the workplace tightly fitting goggles or spectacles with side
shields may be appropriate, but consideration should also be given
to the need for additional face protection. In general, protective
eyewear provided for industrial use should fit snugly to the face,
thus ensuring that there are no gaps through which UV can directly
reach the eye and should have adequate mechanical construction to
prevent physical injury.
For outdoor workers and the general public, the most hazardous
source of UV exposure is the sun. Adequately designed (ski) goggles
afford protection against exposure to solar UV at high altitudes and
on snow, but for most other exposure conditions, good UV absorbing
sunglasses are an adequate means of eye protection.
Transmission of UV through sunglasses varies considerably
(Wester 1987, Gies et al., 1992), yet consumers are provided with
little information about the protection afforded by them. Some
countries have drafted standards limiting UV transmission through
sunglasses and Gies et al. (1990a) have proposed a UV eye
protection factor (EPF) for sunglasses similar to the one developed
for fabrics and sunscreens.
 15. FUTURE RESEARCH
15.1 Introduction
At the United Nations Conference on the Environment and
Development (UNCED) in 1992 it was declared under Agenda 21 that
there should be activities on the effects of UV. Specifically:
(i) Undertake as a matter of urgency, research on the effects on
human health of increasing UV reaching the earth's surface as a
consequence of depletion of the stratospheric ozone layer;
(ii) On the basis of the outcome of this research, consider taking
appropriate remedial measures to mitigate the above mentioned
effects on human beings.
This report gives a thorough review of the health hazards of UV
exposure. However, although it is known that the burden of
UV-related diseases on human populations is high, the exact nature
and extent of these diseases is still largely unknown.
There is great uncertainty about future trends in atmospheric
ozone. For example the Antarctic holes and large depletions of ozone
that have occurred recently were not predicted in any of the ozone
depletion models. While agreements have been reached to reduce
releases of CFCs into the environment, and this will have future
benefit on the ozone layer, there is uncertainty about the extent of
ozone depletion caused by chemical pollutants. What is apparent is
that decreased ozone levels will persist for many years to come and
the corresponding increases in UV intensities will result in more
significant adverse health effects on all populations of the world
for many decades to come (WMO 1993).
The health effects of UV are not restricted to fair skinned
populations. UV exposure is thought to cause diseases of the eye and
suppression of the immune system in all populations of the world. UV
induced immune suppression may have adverse consequences on
infectious disease immunization programmes, particularly in areas
where the UV intensities are high. The possibility that UV will
cause progression of various diseases such as for HIV positive
patients still has to be elucidated. Many such important issues need
to be resolved as a matter of urgency.
The WHO Task Group reviewing this monograph strongly supported
action and coordination of UV research at the international level.
In particular the Task Group supported the concept of the
International Research Programme on Health, Solar UV Radiation and
Environmental Change (INTERSUN). INTERSUN is a collaborative effort
between WHO, the United Nations Environment Programme (UNEP) and the
International Agency on Cancer Research (IARC). The group recognised
specific research needs in areas of exposure assessment, terrestrial
plants, aquatic ecosystems, and human health effects related to the
skin, immune system and eye. Some of these could be accomplished
under the umbrella of INTERSUN, and some, at least initially, would
require more basic laboratory research to be undertaken.
15.2 INTERSUN
The objectives and approach of INTERSUN are: to accurately
evaluate the quantitative relationship between solar UV at the
surface of the earth and human health effects; develop reliable
predictions of health consequences of changes in UV; provide
baseline estimates of the occurrence of health effects of UV in
representative populations around the world; and develop practical
ways of monitoring change in these effects over time in relation to
environmental and behavioural change.
It is intended to establish field research centres covering a
range of latitudes in both hemispheres in which will be measured:
ground level UV irradiance; incidence of non-melanocytic skin
cancer; incidence and mortality of malignant melanoma; immune
function; biological markers of UV exposure and its' early
carcinogenic effects; constitutional sensitivity to sun exposure;
and present and past sun-related behaviour. In addition, in these
centres there will be undertaken case-control or cross-sectional
studies of relationships between skin cancers, cataract and immune
response and constitutional sensitivity to the sun, lifetime
exposure to the sun, and biological markers of UV exposure and early
carcinogenic effects. Surveys should be repeated at 5-yearly
intervals to allow trends in population measurements to be
established.
Data collected at such centres would be used
- to describe quantitatively the relationship between ground
level solar UV irradiance and the incidence of skin cancers and
other health effects (particularly eye damage and effects on
the immune system) of UV exposure in human populations.
- to estimate the change in occurrence of health effects of UV
radiation that would result from change in ground level solar
UV irradiance due to environmental change.
- to increase understanding of the relationship between personal
risk of health effects of UV radiation and constitutional
sensitivity to the sun and sun-related behaviour.
- to develop and validate appropriate ways of monitoring human
exposure to UV and the occurrence of associated health effects.
- to develop a network of centres monitoring trends in ground
level solar UV irradiance, sun exposure of populations, and the
occurrence of health effects of UV radiation.
- to interpret these trends, as far as is possible, in relation
to environmental change, changes in human behaviour, and the
implementation of public policies aimed at ameliorating
environmental change or human exposure to solar UV radiation.
- to provide a basis for development and evaluation of
interventions to reduce the occurrence of adverse health
effects of solar UV radiation.
In line with the above objectives and the gaps in knowledge
needed to be filled to make a satisfactory health risk assessment,
the following recommendations for future research are provided.
15.3 Solar and Personal UV Monitoring
The quantification of health detriment to an individual or to a
population requires the assessment of exposure of the individual or
the average exposure of a member of the exposed population. In
interpreting epidemiological data the exposures of the groups
comprising the study populations must be known. The requirements
here are precise measurement data of the environment occupied by the
exposed individuals combined with data on their exposure habits,
derived from representative personal exposure measurements, skin
damage or from questionnaires.
15.3.1 Solar monitoring
1. A comprehensive network of spectral and broad band monitoring
stations is needed worldwide giving quality ground measurements
of surface UV, to assess the effects of enhanced UVA and UVB
radiation on exposed populations, and to document the impact of
stratospheric ozone depletion on ambient UV. A long-term data
record is important.
2. Quality and convenient monitoring data are needed as input to
models and for model validation. Current models need to be
further modified to account for cloud cover, gaseous pollutants
and aerosols.
3. Common calibration and audit procedures should be developed and
implemented for national and international monitoring
programmes.
4. Monitoring activities should be coordinated with the efforts of
other groups on a national and international scale. Close
coordination is needed for inter-comparison studies and quality
control/quality assurance efforts.
5. There is a need for global monitoring of UV radiation, with
particular attention to long-term instrument stability and
representative geographical deployment
15.3.2 Personal monitoring
Population studies using personal UV monitoring devices are
needed to determine the fraction of the daily natural UV dose
received by persons at risk. The daily amount of UV received by
human skin varies greatly with occupation, behaviour, and local
climatic and environmental conditions. Little is known about these
factors and this seriously interferes with the interpretation of
existing data on the relationship between UV and the development of
skin cancer and of chronic skin and eye damage. Thus, the
development of personal UV monitoring devices is important.
Small passive personal UV dosimeters or monitoring devices
should be further developed and used to provide data on personal
exposures. Biomarkers for UV exposure and skin cancer risk should be
further developed, e.g. UV-induced mutations in p53 tumour
suppressor genes. Such biomarkers might constitute a person's own
"built in" UV risk monitoring device.
For some people, artificial sources of UV represents a
significant component of their exposure. Occupational exposures and
elective exposures such as from cosmetic sunbeds are important in
this respect. Measurement studies are required in these areas.
Standardized monitoring of exposure to the sun in at risk
populations, particularly children, should be evaluated as a guide
to public health action to control melanoma.
15.4 Terrestrial Plants
Particular attention must be given to the impact on food
production in the developing world and to development of crop
varieties resistant to higher levels of UV radiation.
Substantial research is needed to describe and evaluate the
effects of enhanced UV on plants, specifically in the following
areas:
1. Field validations on crop plants should be extended to
determine whether UV affects yield in other agriculturally
important plants.
2. Studies should be initiated to determine the impacts on
ecosystems. Little is known of the effects of UV in other
ecosystems such as forests, where 80% of net primary production
is currently stored.
3. It is important to study natural plants because they serve in
supplying new drugs, medicines and other natural products. They
also act as a reservoir of genetic diversity for modern crop
breeding programmes.
4. Studies are needed to elucidate how UVB effects are modified by
additional factors such as carbon dioxide, temperature, water
and nutrient stress, heavy metals, diseases, and pests.
5. Of increasing interest are man-made air pollutants such as
sulphur dioxide and nitrogen oxides, which are known to have
damaging effects on crops and forests. Attention should be
focused on whether the negative effects of these air pollutants
may be aggravated by UV radiation.
6. The manner and magnitude of adaptation to increased UV,
especially UVB, such as by increased screening pigments or
enhanced DNA damage repair capacity, need to be investigated
both as adaption capacity of individual plants and as
genetically based changes in plant populations. The latter also
provides a basis for plant breeding interventions for
agricultural and tree species.
15.5 Aquatic Ecosystems
Current data suggest that predicted increases in UV, especially
UVB radiation could have important negative effects in the marine
environment. However, uncertainties regarding the magnitude of these
effects remain large, including problems of extrapolating laboratory
findings to the open sea, and the almost complete absence of data on
long-term effects on ecosystems. Additional information is needed in
several areas before more reliable assessments of risk are possible.
Research is needed to:
1. Determine accurate and appropriate biological action spectra
for selected endpoints of key marine species.
2. Produce dose-response data on a greater variety of ecologically
important primary producers than is now available, as well as
data for key higher organisms within the food web.
3. Determine long-term effects for embryos or larvae exposed to
UVB radiation. Is the survival of the adult population (or
their offspring) affected?
4. Determine effects of enhanced UV on major ecosystems, including
the Antarctic ecosystems.
5. Obtain data on the mechanisms of damage and ranges of possible
adaptation or genetic selection in response to increased UVB
radiation.
6. Relate UVB penetration into the water column and through ice
both in laboratory and field research to determine effects on
phytoplankton and zooplankton.
7. Develop biomarkers to monitor current levels of UV damage in
phytoplankton and zooplankton.
15.6 Human Health
15.6.1 Skin
To a substantial degree, answers are still required to the
following questions regarding the relationship between sun exposure
and both melanoma and non-melanocytic skin cancer. Research is
needed to determine:
1. The quantitative relationship between radiant exposure and
incidence of these cancers; and in particular the shape of the
exposure-response relationship.
2. How the effect of sun exposure is modified by: age at exposure,
time since last exposure and pattern of exposure, particularly
whether a particular dose is received intermittently or
more-or-less continuously.
3. Biomarkers of UV exposure or early effects in the skin, e.g.
UV-induced mutations for UV exposure and skin cancer risk.
4. The importance of UVA in causing skin cancers, both relative to
UVB and comparatively between melanoma and non-melanocytic
cancer.
5. The importance of indirect (e.g. oxidative) as opposed to
direct UV damage in causing these cancers.
6. The contribution of sun exposure to the aetiology of these
cancers in dark-skinned populations and how, in general,
cutaneous sensitivity modifies both quantitatively and
qualitatively the relationship between sun exposure and skin
cancer.
7. If the associations of red hair and freckling with skin cancer
are explained by effects of skin pigmentation or sensitivity to
the sun or mediated by other susceptibility mechanisms.
8. If pigmentary characteristics or cutaneous sun sensitivity
explain most of the ethnic variation in skin cancer incidence
or are there other ethnically related susceptibility
characteristics.
9. Whether the relationships between sun exposure and BCC and SCC
are the same.
10. The role of UV-induced mutations of critical genes in causing
melanoma.
11. The role of sun exposure in causing cancer of the lip. An
epidemiological study is required with adequate control of
potential confounding with alcohol and tobacco use.
12. The relationship between use of sunlamps and sunbeds
(artificial tanning) and both melanoma and non-melanocytic skin
cancer. Adequate control of likely confounding with sun
exposure is required.
13. The effects of the interaction between UVB and the rest of the
solar spectrum in relation to DNA repair, malignant
transformation, and skin tumour development.
14. Additional animal models, particularly for the study of the
experimental induction of malignant melanoma. Further, an
action spectrum for induction of melanoma should be developed
in M. domestica to see if the apparent response of UVA
exposure observed in hybrid fish can be confirmed.
15.6.2 Immune System
Suppression of immune functions results from UV exposure of
humans. The following studies are needed to adequately assess the
consequent risk to human health.
1. Studies to determine whether systemic effects occur in humans
as a result of UV exposure.
2. Establish immunological biomarkers that could identify persons
who are susceptible to specific long-term UV-induced immune
suppression, e.g. to infectious agents.
3. Human and rodent studies are needed to determine the
relationship between UV exposure and susceptibility to
infection and vaccine effectiveness, and quantitative
comparison of the relative sensitivities of mice and humans.
4. Studies of the effects of UVB on immune function in human skin
of all types to determine if all skin types show the same
quality and quantity of effects.
5. Studies to examine whether currently available sunscreens
provide adequate protection and whether dietary supplements are
protective. Specifically:
a) Protective effects for immunosuppression;
b) Systemic toxicity based on penetration of organic
chemicals in sunscreens;
c) Carcinogenicity assays.
6. Studies of the spectral effectiveness of UV in producing immune
function defects and enhanced susceptibility to disease.
7. Research to determine the role of UVB exposure in
immediate-type hypersensitivity (allergic) reactions as well as
specific antibody responses, particularly as it relates to
impact on incidence and severity of asthma.
8. Research on the effects of UV on the development of autoimmune
disease and on underlying mechanisms.
9. Studies on systemic effects caused by release of cytokines
within the epidermis and blood stream.
15.6.3 Eye
Two issues underscore the need for undertaking studies of
health risks of UV exposure of the eye on a priority basis. First,
there is evidence of the depletion of the ozone layer and the
reported consequent increase in ambient UV which will impact on many
eye diseases. Second, the overall ageing of the population worldwide
would further compound the magnitude of the cataract and other eye
problems.
1. There is a need for well-conducted epidemiological studies to
be undertaken to define the strength of the association between
UVA and UVB exposure with cataract and other eye conditions,
including climatic droplet keratopathy and pterygium, at both
the individual and community levels. Ideally, this should be
undertaken in a variety of locations. Careful attention needs
to be given to quantifying, for each study participant, ocular
exposure to UV and exposure to other possible risk factors that
may confound observed associations.
2. Techniques need to be refined for measuring the proportion of
UV of different wavelengths incident upon the human cornea and
lens, and how these are affected by the behavioural response to
avoid direct exposure to bright sunlight, and by different
ground surfaces.
3. Studies should be conducted to provide data on the quantitative
relationship between exposure to UV and cataract and other
lesions of the anterior eye. In relation to this there is an
urgent need to establish an internationally agreed system for
grading the type and severity of cataract.
4. Investigations of the role of personal solar exposure in
causing ocular melanoma, including the protective effect of
ocular devices.
5. The relationship between sunlamp or sunbed exposure and
malignant melanoma of the eye needs to be evaluated.
6. The relative efficacy of different protective devices, headwear
and eye wear needs to be determined.
7. Techniques need to be defined for measuring the proportion of
UV of different wavelengths penetrating the human cornea and
lens in the intact eye, in view of possible retinal effects.
8. The action spectra for chronic exposure leading to ocular
damage needs to be determined in primates and other animals.
This includes action spectra for cataract formation, pterygium,
climatic droplet keratopathy, and macular changes in aphakic
animals.
9. Risks to aphakic people and those with intraocular lens
implants for retinal changes, mainly from UVA and short
wavelengths in the visible part of the spectrum, need to be
determined.
15.7 Laboratory Studies
Little is known about the mechanisms of interaction of UV and
environmental chemical agents on biological systems. Many widely
distributed natural or artificial chemicals (pesticides,
halocarbons, etc.) can be altered by UV, resulting in photoproducts
that may be less or more biologically effective than the parent
compound. Furthermore, many chemicals can be activated by UV in
situ in biological systems and this activation may elicit a
biological effect which neither the chemical nor the radiation alone
exhibits (Psoralens). An international registry of agents that
interact with light to cause adverse health effects would speed
identification of such agents.
Much of the information on the chemical and biological effects
of UV comes from experiments using UVC (particularly 254 nm)
radiation not found in sunlight reaching the earth's surface. There
are studies showing direct and indirect effects on cells and
cellular constituents of UVB, UVA, and visible light that differ
considerably from those of UVC. Thus, the chemical and biological
effects of the wavelengths of UV found in sunlight should be
studied. There is evidence that visible light can, under different
conditions, either help cells to repair UV-induced damage or can
potentiate the detrimental effects of UV. Thus, to better understand
the effects of sunlight on humans and the environment, experiments
should be performed using natural sunlight or artificial lamps with
well-known continuous spectra.
Cellular and molecular studies needed include:
1. Extension of relevant biological action spectra to the
UVA/visible wavelengths, as well as interactions between UVA
and UVB radiation. Crucial spectra required include:
- pre-mutagenic lesions
- gene activation
- activation of viruses specially HIV (activation of DNA
binding, promoter activity, viral replication)
2. Determination of the free radical/oxidative component of UVB
and UVA radiation effects. Determine the reactive intermediates
involved.
3. Determination of adaptation responses in human cells.
4. Determination of endogenous cellular antioxidant defences in
eye tissue and skin.
5. Elucidation of DNA repair pathways in humans using the advanced
technology available for analyses of gene and function.
6. Identification of key endogenous and exogenous sensitizers.
7. Studies to examine the effect of UV on the balance between Th1
and Th2 cells.
15.8 Education
It is essential to educate the general population and workers
concerning the profound importance of sunlight and the possibilities
of either UV deprivation or of acute and chronic UV injury. It is
also important to overcome the lack of respect for the possible
adverse effects on health from overexposure to sunlight, simply
because sunlight is ubiquitous, and the concept that, if something
is natural, it must be totally beneficial and safe. In this context
there is a need to standardize a UV exposure index that can be used
as part of health information/education campaigns.
15.9 Administration
An international panel composed of key national experts, with
its secretariat at WHO, should be formed with the following terms of
reference:
(i) to identify gaps in knowledge and set research priorities
needed for a better health risk assessment of exposure to UV
(ii) to evaluate and monitor progress of research
(iii) to establish mechanisms for funding research and initiate
projects where crucial gaps in knowledge have been identified
(iii) to facilitate international cooperation on UV monitoring
efforts, including instrumentation intercomparisons,
calibration standardization and uniform analysis of data, and
(iv) to facilitate international cooperation in measurement of
human UV exposure, monitoring of trends in occurrence of UV
health effects, and epidemiological studies of the
relationship between UV exposure and health effects
(v) to provide a database of research projects to facilitate
cooperation between researchers and institutions involved in
research.
16. PREVIOUS EVALUATIONS BY INTERNATIONAL BODIES
16.1 United Nations Environment Programme
UNEP has undertaken a number of reviews on the environmental
effects of increased UV intensities resulting from ozone depletion
(1989, 1991, 1992). Their findings on each issue are summarized
below:
16.1.1 Ozone
Significant global scale decreases in total ozone have occurred
over the past ten years. All other factors being constant, there is
no scientific doubt that decreases in total ozone will increase UVB
radiation at ground level. Tropospheric ozone and aerosols may have
masked the consequences of stratospheric ozone depletion for UVB in
some industrialized regions. There are no reliable estimates of the
direction or magnitude of effects of any cloud cover trends on UVB.
Efforts to improve local and regional air quality may bring to light
the increases in UVB associated with the depletion of stratospheric
ozone.
16.1.2 Human health
The induction of immunosuppression by UVB has now been
demonstrated in humans, not only those of light pigmentation, but
also deeply pigmented individuals. This places all of the world's
populations at risk of the potential adverse impacts of UVB on the
immune system, including possible increases in the incidence or
severity of infectious disease.
An increased number of adverse ocular effects have been
associated with exposure to UV. These include age-related
nearsightedness, deformation of the lens capsule, and nuclear
cataract (a form of cataract which previous information excluded
from consideration). These effects appear to be independent of
pigmentation. Estimates of risk would increase slightly if one were
to include nuclear cataract among the forms of cataract increasing
with ozone depletion. It is now predicted that, all other things
being equal, a sustained 10% decrease in ozone will be associated
with between 1.6 and 1.75 million additional cases of cataract per
year world-wide.
Recent information on the relationship of nonmelanoma skin
cancer to UV exposures confirms previous findings and has allowed
refinement of the carcinogenic action spectrum. Incorporation of
this new information into the risk estimation process has led to
slightly lower predictions. It is now predicted that a sustained 10%
decrease in ozone will be associated with 26% increase in
non-melanoma skin cancer. All other things remaining constant, this
would mean an increase in excess of 300,000 cases per year
world-wide.
16.2 International Agency for Research on Cancer
The carcinogenicity of ultraviolet has been evaluated by IARC
(1992, 1993). IARC concluded that "There is sufficient evidence in
humans for the carcinogenicity of solar radiation. Solar radiation
causes cutaneous melanoma and nonmelanocytic skin cancer". With
respect to other potential sources of UV, IARC has concluded that
there is limited evidence for the carcinogenicity of exposure to
fluorescent lighting and there is inadequate evidence for the
carcinogenicity of other artificial sources of UV. Readers are
referred to these excellent monographs for more details -much of
which has been incorporated in this text.
16.3 World Health Organization
WHO and its Regional Office for Europe have completed reviews
on the health effects of ultraviolet (WHO, 1979, 1989). They
concluded that all people are exposed to UV from sunlight, and the
risk to health varies with geographical, genetic and other factors.
Similar risks are involved in the increasing exposure of people to
UV from artificial sources, such as those used for suntanning, in
phototherapy and in industrial processes. The biological effects of
a single exposure differ significantly from the effects of repeated
and cumulative exposures. Both types of risk increase markedly with
excessive exposure.
Most observed biological effects of UVB are extremely
detrimental to living organisms. Much less is known about the
biological effects of UVA. It can augment the biological effects of
UVB, and doses of UVA, which alone do not show any biological
effects, can in the presence of certain chemical agents, result in
injury to tissues (phototoxicity, photoallergy, enhancement of
photocarcinogenesis).
All those who work out of doors are potentially at risk of
overexposure, the consequences of which may be both acute and
long-term effects. The fashion of exposing a large part of the body
to sunlight has during recent years increased the exposure of the
skin, resulting in quite high UV doses. This is true not only for
outdoor work but is now also normal during leisure periods, as
exemplified by the holiday exodus of a large part of the population
of the northern European countries to the Mediterranean coast.
16.4 International Commission on Non-Ionizing Radiation Protection
Through its predecessor committee IRPA/INIRC the ICNIRP was
involved in the publication of the original review of the health
effects of UV (WHO/UNEP/IRPA, 1979). It has recommended
international guidelines on limits (IRPA/INIRC, 1991) as shown in
Chapter 13. It has also recommended against the use of sunlamps for
cosmetic purposes (IRPA/INIRC, 1991a). A review of the studies on
fluorescent lighting with reference to UV levels has suggested that
they do not seem to be associated with an increased risk of
malignant melanoma (IRPA/INIRC, 1991).
REFERENCES
Abrahams PJ, Huitema BA, & van der Eb AJ (1984) Enhanced
reactivation and enhanced mutagenesis of herpes simplex virus in
normal human and xeroderma pigmentosum cells. Mol Cell Biol, 4:
2341-2346.
ACGIH (1993) Threshold limit values for chemical substances and
physical agents and biological exposure indices. Cincinnati, Ohio,
The American Conference of Governmental Industrial Hygienists.
Adam SA, Sheaves JK, Wright NH, Mosser G, Harris RW, & Vessey MP
(1981) A case-control study of the possible association between oral
contraceptives and malignant melanoma. Br J Cancer, 44: 45-50.
Adams JS, Clemens TL, Parrish JA, (1982) Vitamin D synthesis and
metabolism after UV irradiation of normal and vitamin D deficient
subjects. N Engl J Med, 306: 722-725.
Ainsleigh HG (1993) Beneficial effects of sun exposure on cancer
mortality. Prev Med, 22: 132-140.
Akslen LA & Morkve O (1992) Expression of p53 protein in cutaneous
melanoma. Int J Cancer, 52: 13-16.
Albino AP, Nanus DM, Davis ML, & McNutt NS (1991) Lack of evidence
of Ki- ras codon 12 mutations in melanocytic lesions. J Cutan
Pathol, 18: 273-278.
Alcalay J, Freeman SE, Goldberg LH, & Wolf JE (1990) Excision repair
of pyrimidine dimers induced by simulated solar radiation in the
skin of patients with basal cell carcinoma. J Invest Dermatol, 95:
506-509.
Anderson RR & Parrish JA (1981) The optics of human skin. J Invest
Dermatol, 77: 13-19.
Andley UP (1987) Photodamage to the eye. Yearly review. Photochem
Photobiol, 46: 1057-1066.
Angel P, Poting A, Mallick U, Rahmsdorf HJ, Schorpp M, & Herrlich P
(1986) Induction of metallothionein and other mRNA species by
carcinogens and tumour promoters in primary human skin fibroblasts.
Mol Cell Biol, 6: 1760-1766.
Ansel JC, Mountz J, Steinberg AD, DeFabo E, & Green Ira (1985)
Effects if UV radiation on autoimmune strains of mice: Increased
mortality and accelerated autoimmunity in BXSB male mice. J Invest
Dermatol, 85: 181-186.
Araneo BA, Dowell T, Moon HB, & Daynes RA (1989) Regulation of
murine lymphokine production in vivo. Ultraviolet radiation
exposure depresses Il-2 and enhances IL-4 production by T cells
through an Il-1 dependent mechanism. J Immunol, 143: 1737-1744.
Arlett CF & Harcourt SA (1983) Variation in response to mutagens
amongst normal and repair-defective human cells. In: Lawrence CW ed.
Induced mutagenesis: Molecular mechanisms and their implications for
environmental protection. New York, London, Plenum Press, pp
249-267.
Arlett CF, Harcourt SA, Cole J, Green MH, & Anstey AV (1992) A
comparison of the response of unstimulated and stimulated
T-lymphocytes and fibroblasts from normal, Xeroderma pigmentosum and
trichothiodystrophy donors to the lethal action of UVC. Mutat Res,
273: 127-135.
Armstrong BK (1984) Melanoma of the skin. Br Med Bull, 40: 346-350.
Armstrong BK (1988) Epidemiology of malignant melanoma: intermittent
or total accumulated exposure to the sun? J Dermatol Surg Oncol, 14:
835-849.
Armstrong BK (1993) Implications of increased solar UVB for cancer
incidence. In: Chanin ML ed. The role of the stratosphere in global
change. Berlin, Heidelberg, New York, Springer-Verlag, pp 517-540.
Armstrong BK & English DR (1988) The epidemiology of acquired
melanocytic naevi and their relationship to malignant melanoma.
Pigment Cell, 9: 27-47.
Armstrong BK, Woodings TL, Stenhouse NS, & McCall MG (1983)
Mortality from cancer in migrants to Australia 1962 to 1971. Perth,
University of Western Australia.
Armstrong BK, de Klerk NH, & Holman CDJ (1986) Etiology of common
acquired melanocytic nevi: constitutional variables, sun exposure
and diet. J Natl Cancer Inst, 77: 329-335.
Atkin M, Fenning J, Heady JA, Kennaway EL, & Kennaway NM (1949) The
mortality from cancer of the skin and lip in certain occupations. Br
J Cancer, 3: 1-15.
Aubry F & MacGibbon B (1985) Risk factors of squamous cell carcinoma
of the skin. A case-control study in the Montreal region. Cancer,
55: 907-911.
Auerbach H (1961) Geographic variation in incidence of skin cancer
in the United States. Public Health Rep, 76: 345-348.
Augustsson A, Stierner U, Rosdahl I, & Suurküla M (1990) Melanocytic
naevi in sun-exposed and protected skin in melanoma patients and
controls. Acta Dermato-Venereol, 71: 512-517.
Azizi E, Lusky A, Kushelevsky AP, & Schewach-Millet N (1988) Skin
type, hair colour and freckles are predictors of decreased minimal
erythema ultraviolet radiation dose. J Am Acad Dermatol, 19: 32-38.
Azizova AO, Islomov AI, Roshchupkin DI, Predvoditelev DA, Remizov
AN, & Vladimir YA (1980) Free radicals formed on ultraviolet
irradiation of the lipids of biological membrane. Biophysics, 24:
407-414.
Baadsgaard O, Wulf HC, Wantzin GL, & Cooper KD (1987) UVB and UVC,
but not UVA, potently induce the appearance of T6-DR+
antigen-presenting cells in human epidermis. J Invest Dermatol, 89:
113-118.
Baadsgaard O, Lisby S, Wulf HC, Wantzin GL, & Cooper KD (1989) Rapid
recovery of Langerhans cell alloreactivity, without induction of
autoreactivity, after in vivo ultraviolet A, but not ultraviolet B
exposure of human skin. J Immunol, 142: 4213-4218.
Baadsgaard O, Salvo B, Mannie A, Dass B, Fox DA, & Cooper KD (1990)
in vivo ultraviolet-exposed human epidermal cells activate T
suppressor cell pathways that involve CD4+CD45RA+ suppressor-inducer
T cells. J Immunol, 145: 2854-2861.
Baasanhu J, Johnson GJ, Burendei G, & Minassian DC (in press)
Prevalence and causes of blindness and visual impairment in
Mongolia: A survey of populations aged 40 and older. Bull. World
Health Organ.
Bachem A (1956) Ophthalmic ultraviolet action spectra. Am J
Ophthalmol, 41: 969-975.
Baird EA, McHenry PM, & MacKie RM (1992) Effect of maintenance
chemotherapy in childhood on numbers of melanocytic naevi. Br Med J,
305: 799-801.
Baker KS & Smith RC (1982) Spectral irradiance penetration in
natural waters. In: Calkins J ed. The role of solar ultraviolet
radiation in marine ecosystems. New York, London, Plenum Press, pp
223-246.
Barbareschi M, Girlando S, Cristofolini P, Cristofolini M, Togni R,
& Boi S (1992) p53 Protein expression in basal cell carcinoma.
Histopathology, 21: 579-581.
Barker & Brainard (1993) Personal communication.
Barker JNWN & MacDonald DM (1988) Eruptive dysplastic naevi
following renal transplantation. Clin Exp Dermatol, 13: 123-125.
Barnes PW, Jordan PW, Gold WG, Flint SD, & Caldwell MM (1988)
Competition, morphology and canopy structure in wheat (Triticum
aestivum L.) and wild oat (Avena fatua L.) exposed to enhanced
ultraviolet-B radiation. Funct Ecol, 2: 319-330.
Barnes PW, Flint SD, & Caldwell MM (1990) Morphological responses of
crop and weed species of different growth forms to ultraviolet-B
radiation. Am J Bot, 77: 1354-1360.
Barrett SF, Robbins JH, Tarone RE, & Kraemer KH (1991) Evidence for
defective repair of cyclobutane pyrimidine dimers with normal repair
and other DNA photoproducts in a transcriptionally active gene
transfected into Cockayne syndrome cells. Mutat Res, 255: 281-291.
Basu-Modak S & Tyrrell RM (1993) Singlet oxygen. A primary effector
in the UVA/near visible light induction of the human heme oxygenase
gene. Cancer Res. 53: 4505-4510.
Bech-Thomsen N, Wulf HC, Poulsen T, & Lundgren K (1988a)
Pretreatment with long-wave ultraviolet light inhibits
ultraviolet-induced skin tumor development in hairless mice. Arch
Dermatol, 124: 1215-1218.
Bech-Thomsen N, Wulf HC, & Lundgren K (1988b) Pre-treatment with UVA
influences broad-spectrum UV photocarcinogenesis in hairless mice
(Abstract). In: Rikles E ed. Proceedings of the 10th International
Congress on Photobiology, Jerusalem. Jerusalem, Israel,
International Association of Biology, p 34.
Beer JZ & Smudzka BZ (1991) Activation of human immunodeficiency
virus by radiation. In: Seymour CB & Murhersill C ed. New
developments in fundamental and applied radiology. London, Taylor
and Francis, pp 113-123.
Beitner H, Norell SE, Ringborg U, Wennersten G, & Mattson B (1990)
Malignant melanoma: aetiological importance of individual
pigmentation and sun exposure. Br J Dermatol, 122: 43-51.
Beral V & Robinson N (1981) The relationship of malignant melanoma,
basal and squamous skin cancers to indoor and outdoor work. Br J
Cancer, 44: 886-891.
Beral V, Evans S, Shaw H, & Milton G (1982) Malignant melanoma and
exposure to fluorescent lighting at work. Lancet, ii: 290-293.
Berger DS & Urbach F (1982) A climatology of sunburning ultraviolet
radiation. Photochem Photobiol, 35: 187-192.
Bergmanson JP, Pitts DG, & Chu LW (1988) Protection from harmful UV
radiation by contact lens. J Am Optom Assoc, 59:,178-182.
Bergmanson JP, Pitts DG, & Chu LW (1987) The efficacy of a
UV-blocking soft contact lens in protecting cornea against UV
radiation. Acta Opthtalmol, 65: 279-286.
Beukers R & Berends W (1960) Isolation and identification of the
irradiation product of thymine. Biochim Biophys Acta, 41: 550-551.
Bhatnagar R, West KP, Vitale S, Sommer A, Joshi S, & Venkataswamy G
(1991) Risk of cataract and history of severe diarrhoeal disease in
Southern India. Arch Ophthalmol, 109: 696-699.
Bhuyan, K.C. and Bhuyan, D.K. (1983) Molecular mechanism of
caractogenesis I reactive species of oxygen as triggering agents II.
Evidence of lipid peroxidation and membrane damage. In Oxy Radicals,
their scavenger syst. Proc. Int. Conf. Superoxide Dismutase 3rd,
vol. 2 (Ed. G. Cohen and R.A. Greenwald), pp 343-356, Elsevier, New
York.
Bidigare RR (1989) Potential effects of UVB radiation on marine
organisms of the Southern Ocean: distributions of phytoplankton and
krill during Austral spring. Photochem Photobiol, 50: 469-477.
Birch-Hirschfeld A (1914) The pathological effect of radiant energy
on the eye. 1. Blinding by sunlight. Ergebn Allg Path des Menschen
und der Tiere 16, 619-634.
Bishop L (1992) Statistical considerations in network design and
data analysis. Presentation to UVB Workshop: A review of the science
and status of measuring and monitoring programs. Washington, DC,
Science and Policy Associates, Inc.
Bissett DL, Hannon DP, & Orr TV (1989) Wavelength dependence of
histological, physical and visible changes in chronically
UV-irradiated hairless mouse skin. Photochem Photobiol, 50: 763-769.
Bittersmann E, Holzwarth AR, Agel G, & Nultsch W (1988) Picosecond
time-resolved emission spectra of photoinhibited and photobleached
Anabaena variabilis. Photochem Photobiol, 47: 101-105.
Black HS (1987a) Photocarcinogenesis and diet. Fed Proc, 46:
1901-1905.
Black HS (1987b) Potential involvement of free radical reactions in
ultraviolet light mediated cutaneous damage. Photochem Photobiol,
46: 213-221.
Black HS & Chan JT (1975) Suppression of ultraviolet light-induced
tumor formation by dietary antioxidants. J Invest Dermatol, 65:
412-412.
Black HS, Thornby JI, Gerguis J, & Lenger, W (1992) Influence of
dietary omega-6,-3 fatty acid sources on the initiation and
promotion stages of photocarcinognesis. Photochem Photobiol, 56:
195-199.
Blum, HF (1948) Sunlight as a causal factor in cancer of the skin of
man. J Natl Inst Cancer, 9: 247-258.
Blum HF, Butler EG, Dailey TH, Daube JR, Mawe RC, & Soffen GA (1959)
Irradiation of mouse skin with single doses of ultraviolet light. J
Natl Cancer Inst, 22: 979-993.
Blumthaler M, Ambach W, & Daxecker F (1987) On the threshold radiant
exposure for keratitis solaris. Invest Opthalmol Vis Sci, 28:
1713-1716.
Blyth WA, Hill TJ, Field HJ, & Harbour DA (1976) Reactivation of
herpes simples virus infection by ultraviolet light and possible
involvement of prostaglandins. J Gen Virol, 33: 547-550.
Bochow TW, West SK, Azar A, Munoz B, Sommer A, & Taylor HR (1989)
Ultraviolet light exposure and risk of posterior subcapsular
cataracts. Arch Ophthalmol, 107: 369-372.
Boettner EA & Wolter JR (1962) Transmission of the ocular media.
Invest Ophthalmol Vis Sci, 1: 776-783.
Boiteux S, O'Connor TR, & Laval J (1987) Formamido pyrimidine-DNA
glycosylase of Escherichia coli: cloning and sequencing of the fpg
structural gene and overproduction of the protein. EMBO J, 6:
3177-3183.
Booth F (1985) Heredity in one hundred patients admitted for
excision of pterygia. Aust N Z J Ophthalmol, 13: 59-61.
Bornman JF & Vogelmann TC (1991) Effect of UV-B radiation on leaf
optical properties measured with fibre optics. J Exp Vot, 42:
547-554.
Bose B, Agarwal S & Chafterjee SN (1989) UV-A induced lipid
peroxidation in liposomal membrane. Radiat. Environ. Biophys. 28
59-65.
Bose SN & Davies RJH (1984) The photoreactivity of T-A sequences in
oligodeoxyribonucleotides and DNA. Nucl Acids Res, 12: 7903-7913.
Bouwes-Bavinck JN (1992) Risk of skin cancer in renal-transplant
recipients. Leiden, The Netherlands (Thesis).
Boyle J, Mackie RM, Briggs JD, Junor BJR, & Aitchison TC (1984)
Cancer, warts, and sunshine in renal transplant patients: A
case-control study. Lancet, 1: 702-705.
Bradley MO, Hsu IC, & Harris CC (1979) Relationships between sister
chromatid exchange and mutagenicity, toxicity and DNA damage. Nature
(Lond), 282: 318-320.
Brash DE & Haseltine WA (1982) UV-induced hotspots occur at DNA
damage hotspots. Nature (Lond), 298: 189-192.
Brash DE, Rudolph JA, Simon JA, Lin A, McKenna GJ, Baden HP,
Halperin AJ, & Pontén J (1991) A role for sunlight in skin cancer:
UV-induced p53 mutations in squamous cell carcinoma. Proc Natl Acad
Sci (USA), 88: 10124-10128.
Braun J (1991) The protective function of phenolic compounds of rye-
and oat seedlings against UV-B radiation and their biosynthetic
regulation, (Thesis) pp. 1-237 in Karlsr Beitr Entw Okophysiol 9, M.
Tevini (ed.), Bot Inst II, Karlsruhe.
Braun J & Tevini M (1991) Regulation of UV-Protective pigment
synthesis in the epidermal layer of rye seedlings ( Secale cereale
L. cv. Kustro). Photochem Photobiol, 37: 318-321.
Bridges BA (1990) Sunlight, DNA damage and skin cancer: a new
perspective. Jpn J Cancer Res, 81: 105-107.
Brilliant LB, Grasset NC, Pokhrel RP, Kolstad A, Lepkowski JM,
Brilliant GE, Hawks WN, & Pararajasegaram R (1983) Associations
among cataract prevalence, sunlight hours, and altitude in the
Himalayas. Am J Epidemiol, 118: 250-264.
Brodkin RH, Kopf AW, & Andrade R (1969) Basal-cell epithelioma and
elastosis: a comparison of distribution. In: Urbach F ed. Biologic
effects of ultraviolet radiation. Oxford, New York, Pergamon Press,
pp 581-618.
Brühl C & Crutzen PJ (1989) On the disproportionate role of
tropospheric ozone as a filter against solar UV radiation. Geophys
Res Lett, 16: 703-706.
Bruls WAG, Slaper H, Van der Leun JC, & Berrens L (1984)
Transmission of human epidermis and stratum corneum as a function of
thickness in the ultraviolet and visible wavelengths. Photochem
Photobiol, 40(4): 485-494.
Bhuyan KC & Bhuyan DK (1983) Molecular mechanism of
cataractogenesis, I. Reactive species of oxygen as triggering
agents, II. Evidence of lipid peroxidation and membrane damage. In
(ed. Cohen G & Greenwald RA) Oxy Radicals, their scavenger syst.
Proc. Int. Conf. Superoxide Dismutase 3rd, vol 2, Elsevier, New
York, pp 343-356.
Cadet J, Ravanat JC, Buchko GW, Yeo HC, & Ames BN (in press) Singlet
oxygen DNA damage: chromatographic and mass spectrometry analysis of
damage products. Methods Enzymol.
Caldwell MM (1976) The effect of solar UV-B radiation (280-315 nm)
on higher plants: implications of stratosphere ozone reduction. In:
Castellani A ed. Research in photobiology. New York, London, Plenum
Press, pp 597-607.
Caldwell MM, Termura AH, & Tevini M (1989) The changing solar
ultraviolet climate and the ecological consequences for higher
plants. Trends Ecol Evol, 4: 363-366.
Cameron ME (1965) Pterygium throughout the world. Springfield,
Illinois, Charles C. Thomas.
Campbell CC, Quinn AG, & Rees JL (1993) Codon 12 Harvey-ras
mutations are rare events in non-melanoma human skin cancer. Br J
Dermatol, 128: 111-114.
Carretto JJ, Carignana MO, Daleo G, & de Marco SG (1990) Occurrence
of mycosporine-like amino acids in the red tide dinoflagellate
Alexandrium excavarum, UV-photoprotective compounds. J Plankton Res,
12: 909-921.
Cartwright LE & Walter JF (1983) Psoralen-containing sunscreen is
tumorigenic in hairless mice. J Am Acad Dermatol, 8: 830-836.
Cerutti PA & Netrawali M (1979) Formation and repair of DNA damage
induced by indirect action of ultraviolet light in normal and
xeroderma pigmentosum skin fibroblasts. Radiat Res, Suppl: 423-432.
Cervenka J, Witkop CJ, Okoro AN, & King RA (1979) Chromosome breaks
and sister chromatid exchanges in albinos in Nigeria. Clin Genet,
15: 17-21.
Césarini JP & Muel B (1989) Erythema induced by quartz- halogen
sources. Photodermatology, 6: 222-227.
Challoner AVJ, Corless D, & Davis A (1976) Personnel monitoring of
exposure to ultraviolet radiation. Clin Exp Dermatol, 1: 175-179.
Chamberlain J & Moss SH (1987) Lipid peroxidation and other membrane
damage produced in Escherichia coli K1060 by near-UV radiation and
deuterium oxide. Photochem Photobiol, 45: 625-630.
Chamberlin GJ & Chamberlin DG (1980) Colour: It's measurement,
computation and application. London, Heyden & Son, p 46.
Chan GL, Peak MJ, Peak JG, & Haseltine WA (1986) Action spectrum for
the formation of endonuclease-sensitive sites and (6-4)
photoproducts induced in a DNA fragment by ultraviolet radiation.
Int J Radiat Biol, 50: 641-648.
Chatterjee A, Milton RC, & Thyle S (1982) Prevalence and aetiology
of cataract in Punjab. Br J Ophthalmol, 66: 35-42.
Christman MF, Morgan RW, Jacobson FS, & Ames BN (1985) Positive
control of a regulon for defences against oxidative stress and some
heat shock proteins in Salmonella typhimurium. Cell, 41: 753-762.
Chu G & Chang E (1988) Xeroderma pigmentosum group E cells lack a
nuclear factor that binds to damaged DNA. Science, 242: 564-567.
CIE (1987) International lighting vocabulary published jointly by
the International Electrotechnical Commission (IEC) and the
International Commission on Illumination (CIE). Geneva,
International Electrotechnical Commission (ISBN 2-8273-0006-0).
CIE (1990) Report on photosensitising chemicals - Updated in 1993.
Vienna, International Commission on Illumination.
CIE (1991) Sunscreen testing (UVB). Vienna, Commission on
Illumination (CIE Technical report - Publication No. CIE 90).
Cleaver JE (1973) Xeroderma pigmentosum - progress and regress. J
Invest Dermatol, 60: 374-380.
Clemmesen J (1965) Statistical studies in the aetiology of malignant
neoplasms. I. Review and results. Copenhagen, Munksgaard, p 409.
Coffman RL, Seymour WP, Lebman DA, Hirakkki DD, Christiansen JA,
Shrader B, Cherwinski HM, Savelkoul HF, Finkelman FD, Bond MW, &
Mosmann TR (1988) The role of helper T cell products in mouse B cell
differentiation and isotype regulation. Immunol Rev, 102: 5-28.
Cogan DG & Kinsey VE (1946) Action spectrum for keratitis produced
by ultraviolet radiation. Arch Ophthalmol, 35: 670-677.
Cole CA, Forbes PD, & Davies RE (1986) An action spectrum for UV
photocarcinogenesis. Photobiology, 43: 275.
Colin J, Bonissent JF, & Resnikoff S (1985) Epidemiology of the
exfoliation syndrome. Proceedings of the 17th Congress of the
European Society of Ophthalmolology, Helsinki, pp 230-231.
Collmann GW, Shore DL, Shy CM, Checkoway H, & Luria AS (1988)
Sunlight and other risk factors for cataract: an epidemiological
study. Am J Public Health, 78: 1459-1462.
Colston K, Berger U, & Coombes RC (1989) Possible role for vitamin D
in controlling breast cancer cell proliferation. Lancet, 1: 188-191.
Connor MJ & Wheeler LA (1987) Depletion off cutaneous glutathione by
ultraviolet radiation. Photochem Photobiol, 46: 239-245.
Cooke KR & Fraser J (1985) Migration and death from malignant
melanoma. Int J Cancer, 36: 175-178.
Coombs BD, Sharples KJ, Cooke KR, Skegg DCG, & Elwood JM (1992)
Variation and covariates of the number of benign nevi in
adolescents. Am J Epidemiol, 136: 344-355.
Cooper KD (1993) Human studies in photo immunology. Photochem
Photobiol, 57: 745.
Cooper KD, Neises GR, & Katz SI (1986) Antigen-presenting OKM5+
melanophages appear in human epidermis after ultraviolet radiation.
J Invest Dermatol, 861: 363-370.
Cooper KD, Oberhelman BS, Hamilton MS, Baadsgaard O, Terhune M,
LeVee G, Anderson T, & Koren H (1992) UV exposure reduces
immunization rates and promotes tolerance to epicutaneous antigens
in humans; relationship to dose, CD1a-DR+ epidermal macrophage
induction and Langerhans cell depletion. Proc Natl Acad Sci (USA),
89: 8497-8501.
Corominas M, Kamino H, Leon J & Pellicer A (1991) Oncogene
activitation in human benign tumors of the skin (keratoacanthomas):
Is H-ras involved in differentiation as well as proliferation? Proc
Natl Acad Sci (USA), 86: 6372-6376.
Coronel VP, Dai QJ, Vergara BS, & Teramura AH (1990) Preliminary
study on response of rice seedlings to enhanced UV-B radiation. Int
Rice Res Newsl, 15: 37.
Coroneo MT (1993) Pterygium as an early indicator of ultraviolet
insolation: a hypothesis. Br J Ophthalmol, 77: 734-739.
Correll DL, Clark CO, Goldberg B, Goodrich VR, Hayes DR, Klein WH, &
Schecher WD (1992) Spectral ultraviolet-B radiation fluxes at the
earth's surface: long-term variations at 39°N, 77°W. J Geogr Res,
97: 7579-7591.
Cox CWJ (1987) Ultraviolet irradiance levels in welding processes.
In: Passachier WF & Bosnjakovic BFM ed. Human exposure to
ultraviolet radiation: Risks and regulations. Amsterdam, Oxford, New
York, Elsevier Science Publishers, pp 383-386.
Cristofolini M, Franceschi S, Tasin L, Zumiani G, Piscioli F,
Talamini R, & La Vecchia C (1987) Risk factors for cutaneous
malignant melanoma in a northern Italian population. Int J Cancer,
39: 150-154.
Crombie IK (1981) Distribution of malignant melanoma on the body
surface. Br J Cancer, 43: 842-849.
Cruickshanks KJ, Klein R, & Klein BE (1993) Sunlight and age-related
macular degeneration: the Beaver Dam Eye Study. Arch Ophthalmol,
111: 514-518.
Cunningham ML, Johnson JS, Giovanazzi SM, & Peak MJ (1985)
Photosensitized production of superoxide anion by monochromatic
(290-405 nm) ultraviolet irradiation of NADH and NADPH coenzymes.
Photochem Photobiol, 42: 125-128.
Cyrlin MN, Pedris-Leftick A, & Sugar J (1980) Cataract formation in
association with ultraviolet photosensitivity. Ann Ophthalmol, 12:
786-790.
Czochralska B, Bartosz W, & Shugar D (1984) Oxidation of
excited-state NADH and NAD dimer in aqueous medium - involvement of
O2- as a mediator in the presence of oxygen. Biochim Biophys Acta,
801: 403-409.
Danielle RP (1988) Pathophysiology of the asthmatic syndromes. In:
Danielle RP ed. Immunologic diseases of the lung. Boston, Blackwell
Scientific Publishers, pp 503-516.
Dardanoni L, Gafá L, Paterno R, & Pavone G (1984) A case- control
study on lip cancer risk factors in Ragusa (Sicily). Int J Cancer,
34: 335-337.
Dargent JL, Lespagnard L, Heenan M, & Verhest A (1992) Malignant
melanoma occurring in a case of oculocutaneous albinism.
Histopathology, 21: 74-76.
Darrell RW & Bachrach CA (1963) Pterygium among veterans. Arch
Ophthalmol, 70: 158-169.
Davies DM (1985) Calcium metabolism in healthy men deprived of
sunlight. Ann N Y Acad Sci, 453: 21-27.
Daynes RA & Spellman CW (1977) Evidence for the generation of
suppressor cells by UV radiation. Cell Immunol, 31: 182-187.
Daynes RA, Spellman CW, Woodward JG, & Stewart DA (1977) Studies
into the transplantation biology of ultraviolet light-induced
tumours. Transplantation, 23: 343-348.
De Fabo EC & Kripke ML (1979) Dose-response characteristics of
immunologic unresponsiveness to UV-induced tumors produced by UV
irradiation of mice. Photochem Photobiol, 30: 385-390.
De Fabo EC & Noonan FP (1983) Mechanism of immune suppression by
ultraviolet irradiation in vivo. J Exp Med, 157: 84-98.
de Gruijl FR & van der Leun JC (1980) A dose-response model for skin
cancer induction by chronic UV exposure of a human population. J
Theor Biol, 83: 487-504.
de Gruijl FR & van der Leun JC (1982) Systemic influence of
pre-irradiation of a limited skin area on UV-tumorigenesis.
Photochem Photobiol, 35: 379-383.
de Gruijl FR & van der Leun JC (1983) Follow up on systemic
influence of partial pre-irradiation on UV-tumorigenesis. Photochem
Photobiol, 38: 381-383.
de Gruijl FR & van der Leun JC (1991) Action spectra for
carcinogenesis. In: Urbach F ed. Biological responses to UVA.
Overland Park, Kansas, Valdemar Publishing Company, p 91.
de Gruijl FR, van der Meer JB, & van der Leun JC (1983) Dose-time
dependency of tumor formation by chronic UV exposure. Photochem
Photobiol, 37: 53-62.
de Gruijl FR, Sterenborg HJCM, Forbes PD, Davies RE, Cole C,
Kelfkens G, van Weelden H, Slaper H, & van der Leun JC (1993)
Wavelength dependence of skin cancer induction by ultraviolet
radiation of albino hairless mice. Cancer Res, 53: 53-60.
Denkins Y, Fidler IJ, & Kripke ML (1989) Exposure of mice to UV-B
radiation suppresses delayed hypersensitivity to Candida albicans.
Photochem Photobiol, 49: 615-619.
Desai ID, Sawant PL, & Tappel AL (1964) Peroxidative and radiation
damage to isolated lysosomes. Biochim Biophys Acta, 86: 277-285.
Detels R & Dhir SP (1967) Pterygium: a geographical study. Arch
Ophthalmol, 78: 485-491.
Devary Y, Gottlieb RA, Smeal T & Karin M (1992) The mammalian
ultraviolet response is triggered by activation of Src tyrosine
kinases. Cell 71: 1081-1091.
Devary Y, Rosette C, Di Donato JA, & Karin M (1993) NFkB activation
by ultraviolet light not dependent on a nuclear signal. Science,
261: 1442-1445.
Dhir SP, Detels R, & Alexander ER (1967) The role of environmental
factors in cataract, pterygium and trachoma. Am J Ophthalmol, 64:
128-135.
Diffey BL (1977) The calculation of the spectral distribution of
natural ultraviolet radiation under clear sky conditions, Phys Med
Biol, 22: 309-316.
Diffey BL (1986) Use of UVA sunbeds for cosmetic tanning. Br J
Dermatol, 225: 67-76.
Diffey BL (1987) Analysis of the risk of skin cancer from sunlight
and solaria in subjects living in northern Europe. Photodermatology,
4: 118-126.
Diffey BL (1988) The risk of skin cancer from occupational exposure
to ultraviolet radiation in hospitals. Phys Med Biol, 33(10):
1187-1193.
Diffey BL (1989) Ultraviolet radiation and skin cancer: Are
physiotherapists at risk? Physiotherapy, 75(10): 615-616.
Diffey BL (1989a) Ultraviolet radiation dosimetry with polysulphone
film. In: Diffey BL ed. Radiation measurement in photobiology. New
York, London, San Francisco, Academic Press, pp 135-159.
Diffey BL (1990) Human exposure to ultraviolet radiation. Semin
Dermatol, 9: 2-10.
Diffey BL (1993) A photobiological evaluation of lamps used in the
phototherapy of seasonal affective disorder. J Photochem Photobiol,
B17: 203-207.
Diffey BL (1993a) Personal communication.
Diffey BL & Cheeseman J (1992) Sun protection with hats. Br J
Dermatol, 127: 10-12.
Diffey BL & McKinlay AF (1983) The UVB content of 'UVA fluorescent
lamps' and its erythemal effectiveness in human skin. Phys Med Biol,
28: 351-358.
Diffey BL & Robson J (1992) The influence of pigmentation and
illumination on the perception of erythema. Photodermatol
Photoimmunol Photomed, 9: 45-47.
Diffey BL, Tate TJ, & Davis A (1979) Solar dosimetry of the face:
the relationship of natural ultraviolet exposure to basal cell
carcinoma localisation. Phys Med Biol, 24: 931-939.
Diffey BL, Challoner AVJ, & Key PJ (1980) A survey of the
ultraviolet radiation emissions of photochemotherapy units. Br J
Dermatol, 102: 301-306.
Diffey BL, Larko O, & Swanbeck G (1982) UVB doses received during
different outdoor activities and UVB treatment of psoriasis. Br J
Dermatol, 106: 33-41.
Dillon J, Wang RH, & Atherton S (1990) Photochemical and
photophysical studies on human lens constituent. Photochem
Photobiol, 52: 849-854.
Dion M & Hammelin C (1987) Relationship between enhanced
reactivation and mutagenesis of UV-irradiated human cytomegalovirus
in normal human cells. EMBO J, 6: 397-399.
Djavuheri-Mergny M, Maziére JC, Santus R, Mora L, Maziére C, Auclair
M & Dubertet C (1993) Exposure to long wavelength ultraviolet
radiation decreases processing of low density lipoprotein by
cultured human fibroblasts. Photochem Photobiol 57: 302-305.
Döhler G (1990) Effect of UVB (290-320nm) radiation on uptake of
15N-nitrate by marine diatoms. Berlin, Heidelberg, New York,
Springer-Verlag, pp 354-359.
Döhler G & Alt MR (1989) Assimilation of 15N-ammonia during
irradiance with ultraviolet-B and monochromatic light by
Thalassiosira rotula, C R Acad Sci Paris, D308, 513-518.
Dolezal JM, Perkins ES, & Wallace RB (1989) Sunlight, skin
sensitivity, and senile cataract. Am J Epidemiol, 129: 559-568.
Doll R (1991) Urban and rural factors in the aetiology of cancer.
Int J Cancer, 47: 803-810.
Doll R, Muir C, & Waterhouse J ed. (1970) Cancer incidence in five
continents, Volume II. Berlin, Heidelberg, New York,
Springer-Verlag.
Doniger J, Jacobson ED, Krell K, & DiPaolo JA (1981) Ultraviolet
light action spectra for neoplastic transformation and lethality of
syrian hamster embryo cells correlate with spectrum for pyrimidine
dimer formation in cellular DNA. Proc Natl Acad Sci (USA), 78:
2378-2383.
Dorn HF (1944b) Illness from cancer in the United States: IV.
Illness from cancer of specific sites classed in broad groups.
Public Health Rep, 59: 65-77.
Dorn HF (1944a) Illness from cancer in the United States. Public
Health Rep, 59: 33-48.
Dorn CR, Taylor DON, & Schneider R (1971) Sunlight exposure and risk
of developing cutaneous and oral squamous cell carcinomas in white
cats. J Natl Cancer Inst, 46: 1073-1078.
Doughty MJ & Cullen A (1990) Long-term effects of a single dose of
ultraviolet B on albino rabbit cornea. II. Deturgescence and fluid
pump assessed in vitro. Photochem Photobiol, 54: 439-449.
Driscoll CMH (1992) Solar UV trends and distributions. Natl Radiat
Prot Board Bull, 137: 7-13.
Dubin N, Moseson M, & Pasternack BS (1986) Epidemiology of malignant
melanoma: pigmentary traits, ultraviolet radiation, and the
identification of high-risk populations. Recent Results Cancer Res,
102: 56-75.
Duke-Elder WS (1926) The pathological action of light upon the eye.
I. Action on the outer eye: Photophthalmia. Lancet, 1: 1137-1141.
Duvic M, Lowe L, Rapini RP, Rodriguez S, & Levy ML (1989) Eruptive
dysplastic nevi associated with human immunodeficiency virus
infection. Arch Dermatol, 125: 397-401.
Dyall-Smith D & Varigos G (1985) The malignant potential of
papillomavirus. Aust J Dermatol, 26: 102-107.
Eggersdorfer B & Häder DP (1991) Phototaxis, gravitaxis and vertical
migration in the marine dinoflaggelates. Acta Protozool 30(2):
63-71.
Eisenstark A & Perrot G (1987) Catalase has only a minor role in
protection against ultraviolet radiation damage in bacteria. Mol Gen
Genet 207: 68-72.
Eisman JA, MacIntyre I, Martin TJ, Frampton RJ, & King RJB (1980)
Normal and malignant breast tissue is a target organ for 1,25-(OH)2
vitamin D3. Clin Endocrinol, 13: 267-272.
Elliott R (1961) The aetiology of pterygium. Trans Ophthalmol Soc N
Z, 13: 22-41.
Ellison MJ & Childs JD (1981) Pyrimidine dimers induced in
Escherichia coli DNA by ultraviolet radiation present in sunlight.
Photochem Photobiol, 34: 465-469.
Elmets CA, Bergstresser PR, Tigelaar RE, Wood PJ, & Streilein JW
(1983) Analysis of the mechanism of unresponsiveness produced by
haptens painted on skin exposed to low dose ultraviolet radiation. J
Exp Med, 158: 781-794.
Elwood, JM & Gallagher RP (1983) Site distribution of malignant
melanoma. Can Med Assoc J, 128: 1400-1404.
Elwood JM, Gallagher RP, Hill GB, Spinelli JJ, Pearson JCG, &
Threlfall W (1984) Pigmentation and skin reaction to sun as risk
factors for cutaneous melanoma: Western Canada melanoma study. Br
Med J, 288: 99-102.
Elwood JM, Gallagher RP, Davison J, & Hill GB (1985a) Sunburn,
suntan and the risk of cutaneous malignant melanoma: the Western
Canada melanoma study. Br J Cancer, 51: 543-549.
Elwood JM, Gallagher RP, Hill GB, & Pearson JCG (1985b) Cutaneous
melanoma in relation to intermittent and constant sun exposure: the
Western Canada melanoma study. Int J Cancer, 35: 427-433.
Elwood JM, Williamson C, & Stapleton PJ (1986) Malignant melanoma in
relation to moles, pigmentation, and exposure to fluorescent and
other lighting sources. Br J Cancer, 53: 65-74.
Elwood JM, Whitehead SM, Davison J, Stewart M, & Galt M (1990)
Malignant melanoma in England: risk associated with naevi, freckles,
social class, hair colour, and sunburn. Int J Epidemiol, 19:
801-810.
Emmett EA (1973) Ultraviolet radiation as a cause of skin tumors.
Crit Rev Toxicol, 2, 211-255.
Engel A, Johnson M, & Haynes S (1988) Health effects of sunlight
exposure in the United States. Arch Dermatol, 124: 72-79.
English DR & Armstrong BK (in press) Cutaneous malignant melanoma.
In: Schottenfeld, D Fraumeni. J eds, Cancer epidemiology and
prevention, New York, Oxford University Press.
Enninga IC, Groenendijk RTL, Filon AR, van Zeeland AA, & Simons JWIM
(1986) The wavelength dependence of UV-induced pyrimidine dimer
formation, cell killing and mutation induction in human diploid skin
fibroblasts. Carcinogenesis, 7: 1829-1836.
Epe B, Pflaum M, & Boiteux S (1993) DNA damage induced by
photosensitisers in cellular and cell-free systems. Mutat Res 299:
135-145.
Epstein JH (1965) Comparison of the carcinogenic and cocarcinogenic
effects of ultraviolet light on hairless mice. J Natl Cancer Inst,
34: 741-745.
Epstein JH (1985) Animal models for studying photocarcinogenesis.
In: Maibach H & Lowe N ed. Models in Dermatology. Basel, Karger, vol
2, pp 303-312.
Epstein JH (1988) Photocarcinogenesis promotion studies with benzoyl
peroxide (BPO) and croton oil. J Invest Dermatol, 91: 114-116.
Epstein JH & Epstein WL (1962) Cocarcinogenic effect of ultraviolet
light on DMBA tumor initiation in albino mice. J Invest Dermatol,
39: 455-460.
Epstein JH & Roth HL (1968) Experimental ultraviolet light
carcinogenesis. A study of croton oil promoting effects. J Invest
Dermatol, 50: 387-389.
Epstein JH, Tuffanelli DL, & Dubois EL (1965) Light sensitivity and
lupus erythematosus. Arch Dermatol, 91: 483-485.
Epstein JH, Epstein WL, & Nakai T (1967) Production of melanomas
from DMBA-induced 'blue nevi' in hairless mice with ultraviolet
light. J Natl Cancer Inst, 38: 19-30.
Eriksen P (1987) Occupational applications of ultraviolet radiation:
risk evaluation and protection techniques. In: Passchier WF &
Bosnjakovic BFM ed. Human exposure to ultraviolet radiation: Risks
and regulations. Amsterdam, Oxford, New York, Elsevier Science
Publishers, pp 317-331.
Everett MA, Olsen RL, & Sayer RM (1965) Ultraviolet erythema. Arch
Dermatol, 92: 713-719.
Farr PM, Marks JM, Diffey BL, & Ince P (1988) Skin fragility and
blistering due to the use of sunbeds. Br Med J, 296: 1708-1709.
FDA (1988) Quality control guide for sunlamp products, Washington,
DC, US Dept. Health and Human Services, (HHS Publication FDA No.
88-8234).
FDA (1992) Medications that increase sensitivity to light (prepared
by Levine JI). Rockville, Maryland, Food and Drug Administration,
Center of Devices and Radiological Health.
Findlay GM (1928) Ultra-violet light and skin cancer. Lancet, ii:
1070-1073.
Finsen NR (1901) The treatment of lupus vulgaris by concentrated
chemical rays. In: Phototherapy. London, Edward Armold, pp 73-75.
Fisher GJ & Johns HE (1976) Pyrimidine photohydrates. In: (Ed. Wang,
S.) Photochemistry and Photobiology of Nucleic Acids, vol. 1, New
York, Academic Press, pp 169-224.
Fisher MS & Kripke ML (1977) Systemic alteration induced in mice by
ultraviolet light irradiation and its relationship to ultraviolet
carcinogenesis. Proc Natl Acad Sci (USA), 74: 1688-1692.
Fisher MS, Menter JM, & Willis I (1989) Ultraviolet
radiation-induced suppression of contact hypersensitivity in
relation to padimate O and oxybenzone. J Invest Dermatol, 92:
337-341
Fitzpatrick TB & Sober AJ (1985) Sunlight and skin cancer. N Engl J
Med, 313: 818-820.
Fitzpatrick TB, Pathak MA, Magnus IA, & Curwen WL (1963) Abnormal
reactions of man to light. Annu Rev Med, 14: 195-214.
Fitzpatrick TB, Pathak MA, Harber LC, Seiji M, & Kukita A (1974)
Sunlight and man. Tokyo, University of Tokyo Press.
Fleming ID, Barnawell JR, Burlison PE, (1975) Skin cancer in black
patients. Cancer, 35: 600-605.
Forbes PD, Davies RE, Sambuco CP, & Urbach F (1989) Topical urocanic
acid enhances UV-induced skin tumors in hairless mice by topical
application of the sunscreen 2-ethylhexyl-p-methoxycinnamate. J
Toxicol Cutan Ocul Toxicol, 8: 209-226.
Forbes PD, Blum HF, & Davies RE (1981) Photocarcinogenesis in
hairless mice: dose-response and the influence of dose-delivery.
Photochem Photobiol, 34: 361-365.
Forbes PD, Davies RE, Urbach F, Berger D, & Cole C (1982) Simulated
stratospheric ozone depletion and increased ultraviolet radiation:
effects on photocarcinogenesis in hairless mice. Cancer Res, 42:
2796-2803.
Forbes PD, Davies RE, & Urbach F (1978) Experimental ultraviolet
photocarcinogenesis: wavelength interactions and time-dose
relationships. Natl Cancer Inst Monogr, 50: 31-38.
Forsius H (1988) Exfoliation syndrome in various ethnic populations.
Acta Ophthalmol, 68(Suppl 184): 71-85.
Foster HM & Webb SJ (1988) Skin cancer in the North Solomons. Aust N
Z J Surg, 58: 397-401.
Fragu P, Lemarchand-Venencie F, Benhamou S, Francois P, Jeannel D,
Benhamou E, Sezary-Lartigau I, & Avril MF (1991) Long-term effects
in skin and thyroid after radiotherapy for skin angiomas - A French
retrospective cohort study. Eur J Cancer, 27: 1215-1222.
Franklin WA, Lo KM, & Haseltine WA (1982) Alkaline lability of
fluorescent photoproducts produced in ultraviolet light-irradiated
DNA. J Biol Chem, 157: 13535-13543.
Frederick JE, Snell HE, & Haywoood EK (1989) Solar ultraviolet
radiation at the earth's surface. Photochem Photobiol, 51: 443-450.
Freeman RG (1975) Data on the action spectrum for ultraviolet
carcinogenesis. J Natl Cancer Inst, 55: 1119-1122.
Freeman SE, Hacham H, Gange RW, Maytum DJ, Sutherland JC, &
Sutherland BM (1989) Wavelength dependence of pyrimidine dimer
formation in DNA of human skin irradiated in situ with ultraviolet
light. Proc Natl Acad Sci (USA), 86: 5605-5609.
Friedberg EC (1984) DNA repair. New York, Freeman and Company.
Friedmann PS, White SI, Parker S, Moss C, & Matthews JNS (1989)
Antigenic stimulation during ultraviolet therapy in man does not
result in immunological tolerance. Clin Exp Immunol, 76: 68-72.
Fuchs J & Packer L (1991) Photooxidative stress in the skin. In:
Sies H Ed. Oxidative stress: oxidants and antioxidants. New York,
London, San Francisco, Academic Press, pp 554-584.
Fuchs J, Huflejt M, Rothfuss L, Carcamero G, & Packer L (1989)
Impairment of enzymic and nonenzymic antioxidants in skin by UVB
irradiation. J Invest Dermatol 93, 769-773.
Fuller CJ, Faulkner H, Bendich A, Parker RS, & Roe DA (1992) Effect
of ß-carotene supplementation on photosuppression of delayed-type
hypersensitivity in normal young men. Am J Clin Nutr, 56: 684-690.
Funnell SGP & Keast D (1985) The effect of ultraviolet radiation on
the generation of plaque-forming cells and on T-suppressor cell
activity to sheep erythrocytes. Photodermatology, 3: 64-72.
Gaboriau F, Morlière P, Marquis I, Maysan A, Gèze M, & Dubertret L
(1993) Membrane damage induced in cultured human skin fibroblasts by
UVA irradiation. Photochem Photobiol, 58: 515-520.
Gafa L, Filippazzo MG, Tumino R, Dardanoni G, Lanzarone F, &
Dardanoni L (1991) Risk factors of nonmelanoma skin cancer in
Ragusa, Sicily: a case-control study. Cancer Causes Control, 2:
395-399.
Gallagher, RP (1988) Ocular melanoma in farmers (Letter to the
Editor). Am J Ind Med, 13: 523-525.
Gallagher PE & Duker NJ (1986) Detection of UV purine photoproduct
in a defined sequence of human DNA. Mol Cell Biol, 6: 707-709.
Gallagher RP, Elwood JM, & Hill GB (1986) Risk factors for cutaneous
malignant melanoma: the Western Canadian melanoma study, Rec Res
Cancer Res, 102: 38-55.
Gallagher RP, Threlfall WJ, Jeffries E, Band PR, Spinelli J, &
Coldman AJ (1984) Cancer and aplastic anemia in British Columbia
farmers. J Natl Cancer Inst, 72: 1311-1315.
Gallagher CH, Greenoak GE, Reeve VE, Canfield PJ, Baker RSU, & Bonin
AM (1984a) Ultraviolet carcinogenesis in the hairless mouse skin -
influence of the sunscreen 2-ethylhexyl-p-methoxycinnamate. Aust J
Exp Biol Med Sci, 62: 577-588.
Gallagher RP, Elwood JM, Rootman J, Spinelli JJ, Hill GB, Threlfall
WJ, & Birdsell JM (1985) Risk factors for ocular melanoma: Western
Canada melanoma study. J Natl Cancer Inst, 74: 775-778.
Gallagher PE, Weiss RB, Brent TP, & Duker NJ (1989) Wavelength
dependence of DNA incision by a human ultraviolet endonuclease.
Photochem Photobiol, 49: 363-367.
Gallagher RP, Ma B, McLean DI, Yang CP, Ho V, Carruthers JA, &
Warshawski LM (1990a) Trends in basal cell carcinoma, squamous cell
carcinoma, and melanoma of the skin from 1973 through 1987. J Am
Acad Dermatol, 23: 413-421.
Gallagher RP, McLean DI, Yang CP, Coldman AJ, Silver HKB, Spinelli
JJ, & Beagrie M (1990b) Anatomic distribution of acquired
melanocytic nevi in white children. A comparison with melanoma: the
Vancouver mole study. Arch Dermatol, 126: 466-471
Gallagher RP, McLean DI, Yang CP, Coldman AJ, Silver HKB, Spinelli
JJ, & Beagrie M (1990c) Suntan, sunburn and pigmentation factors and
the frequency of acquired melanocytic nevi in children. Similarities
to melanoma: the Vancouver mole study. Arch Dermatol, 126: 770-776
Gallagher RP, Rivers JK, Yang CP, (1991) Melanocytic nevus density
in Asian, Indo-Pakistani, and white children: The Vancouver mole
study. J Am Acad Dermatol, 25: 507-12.
Gange RW, Blackett AD, Matzinger EA, Sutherland BM, & Kochevar IE
(1985) Comparative protection efficiency of UVA-and UVB-induced tans
against erythema and formation of endonuclease-sensitive sites in
DNA by UVB in human skin. J Invest Dermatol, 85: 362-364.
Garbe C, Krüger S, Stadler R, Guggenmoos-Holzmann I, & Orfanos CE
(1989) Markers and relative risk in a German population for
developing malignant melanoma. Int J Dermatol, 28: 517-523.
Garcia-Pichel F & Castenholz RW (1991) Characterization and
biological implications of scytonemin, a cyanobacterial sheath
pigment, J Phycol, 27: 395-409.
Garland C & Garland F (1980) Do sunlight and vitamin D reduce the
likelihood of colon cancer? Int J Epidemiol, 9: 227-231.
Garland CF, Comstock GW, Garland FC, Felsing K, Shaw EK, & Gorham ED
(1989) Serum 25-hydroxyvitamin D and colon cancer: 8-year
prospective study. Lancet, 2: 1176-1178.
Garland FC, Garland CF, Gorham ED, & Young JF (1990) Geographic
variation in breast cancer mortality in the United States: A
hypothesis involving exposure to solar radiation. Prev Med, 19:
614-622.
Garland CF Garland FC, & Gorham ED (1992) Could sunscreens increase
melanoma risk? (Letter) Am J Public Health, 82(4): 614-615.
Garner A (1989) The pathology of tumours at the limbus. Eye, 3:
210-217.
Garsen J, Goettsch W, de Gruy F, & van Loveren H (1993) UVB
suppresses immunity and resistance against systemic infections in
the rat. Photochem Photobiol, 57: 755.
Gasparro F & Fresco J (1986) Ultraviolet-induced 8,8-adenine
dehydromimers in oligo- and polynucleotides. Nucl Acids Res, 14:
4239-4251.
Gellin GA, Kopf AW, & Garfinkel L (1965) Basal cell epithelioma. A
controlled study of associated factors. Arch Dermatol, 91: 38-45.
Gensler HL (1988) Enhancement of chemical carcinogenesis in mice by
systemic effects of ultraviolet radiation. Cancer Res, 48: 620-623.
Gensler HL (1989) Reduction of immunosuppression in UV-irradiated
mice by dietary retinyl palmitate plus canthaxanthin.
Carcinogenesis, 10: 203-207.
Gensler HL & Bowden GT (1987) UVB-Induced modulation of mouse skin
tumor induction by benzo[a]pyrene (Abstract no. 546). Proc Am Assoc
Cancer Res, 28: 137.
Gensler HL & Welch K (1992) Prevalence of tumor prevention rather
than tumor enhancement when repetitive UV radiation treatments
precede initiation and promotion. Carcinogenesis, 13: 9-13.
Gerrish KE & Gensler HL (1993) Prevention of photocarcinogenesis by
dietary vitamin E. Nutr Cancer, 19: 125-133.
Giannini SH (1986a) Suppression of pathogenesis in cutaneous
leishmaniasis by UV irradiation. Infect Immun, 51: 838-843.
Giannini SH (1986b) Effects of UV-B on infectious disease. In: Titus
JG ed. Effects of changes in stratospheric ozone and global climate.
Washington, DC, US Environmental Protection Agency, vol 2, pp
101-112.
Giannini SH (1987) Abrogation of skin lesions in cutaneous
leishmaniasis by ultraviolet irradiation. In: Hart DT ed.
Leishmaniasis: The first centenary (1885-1985) new strategies for
control. New York, London, Plenum Press, pp 677-684 (NATO ASI Series
A: Life Sciences).
Giannini SH (1992) Effects of ultraviolet B irradiation on cutaneous
leishmaniasis. Parasitology Today, 8: 44-48.
Gibbs NK (1993) Is ultraviolet immunosuppression initiated by
photosomerisation of urocanic acid. In: de Gruijl FR ed. The dark
side of sunlight. Utrecht, The Netherlands, Utrecht University.
Gies HP & Roy CR (1990) Bilirubin phototherapy and potential UVR
hazards. Health Phys, 58: 313-320.
Gies HP, Roy CR, & Elliott G (1986) Artificial suntanning: Spectral
irradiance and hazard evaluation of ultraviolet sources. Health
Phys, 50(6): 691-703.
Gies HP, Roy CR, & Elliot G (1990) A proposed UVR protection factor
for sunglasses. Clin Exp Optom, 73: 183-189.
Gies HP, Roy CR, & Elliott G (1992) Ultraviolet radiation protection
factors for personal protection in both occupational and
recreational situations. Radiat Prot Aust, 10(3): 59-66.
Gies HP, Roy CR, Herlihy E, & Rivers J (1992a) Personal dosimetry of
solar UVB using polysulphone film Congress Proceedings (IRPA 8) vol
1, p 791, Montreal, May 1992, International Radiation Protection
Association.
Gilchrest BA (1990) Actinic injury. Annu Rev Med, 41: 199-210.
Gilchrest BA, Soter NA, Stoff JS, & Mihm MC Jr (1981) The human
sunburn reaction: histologic and biochemical studies. J Am Acad
Dermatol, 5: 411-422.
Giles GG, Marks R, & Foley P (1988) Incidence of non- melanocytic
skin cancer treated in Australia. Br Med J, 296: 13-17.
Glass AG & Hoover RN (1989) The emerging epidemic of melanoma and
squamous cell skin cancer. J Am Med Assoc, 262: 2097-2100.
Goldberg LH & Altman A (1984) Benign skin changes associated with
chronic sunlight exposure. Cutis, 34: 33-39.
Goodman GJ, Marks R, Selwood TS, Ponsford MW, & Pakes W (1984)
Non-melanocytic skin cancer and solar keratoses in Victoria -
clinical studies II. Aust J Dermatol, 25: 103-106.
Gorham ED, Garland CF, & Garland FC (1989) Acid haze air pollution
and breast and colon cancer mortality in 20 Canadian cities. Can J
Public Health, 80: 96-100.
Gorham ED, Garland FC, & Garland CF (1990) Sunlight and breast
cancer incidence in the USSR. Int J Epidemiol, 19: 820-824.
Graham S, Marshall J, Haughey B, Stoll H, Zielezny M, Brasure J, &
West D (1985) An inquiry into the epidemiology of melanoma. Am J
Epidemiol, 122: 606-619.
Gray RH, Johnson GJ, & Freedman A (1992) Climatic droplet
keratopathy. Surv Ophthalmol, 36: 241-253.
Greaves MW, Hensby CN, Black AK, Plummer NA, Fincham N, Warin AP, &
Camp R (1978) Inflammatory reactions induced by ultraviolet
irradiation. Bull. Cancer, 65: 299-304.
Green AC (1984) Sun exposure and the risk of melanoma. Aust J
Dermatol, 25: 99-102.
Green AC (1991) Premature ageing of the skin in a Queensland
population. Med J Aust, 155: 473-478.
Green AC & Battistutta D (1990) Incidence and determinants of skin
cancer in a high-risk Australian population. Int J Cancer, 46:
356-361.
Green A & O'Rourke MGE (1985) Cutaneous malignant melanoma in
association with other skin cancers. J Natl Cancer Inst, 74:
977-980.
Green A & Siskind V (1983) Geographical distribution of cutaneous
melanoma in Queensland. Med J Aust, 1: 407-410.
Green AES, Sawada T, & Shettle EP (1974) The middle ultraviolet
reaching the ground. Photochem Photobiol, 19: 251.
Green AC, Siskind V, Bain C, & Alexander J (1985) Sunburn and
malignant melanoma. Br J Cancer, 51: 393-397.
Green AC, Beardmore G, Hart V, Leslie D, Marks R, & Staines D
(1988a) Skin cancer in a Queensland population. J Am Acad Dermatol,
19: 1045-1052.
Green AC, Sorahan T, Pope D, Siskind V, Hansen M, Hanson L, Leech P,
Ball PM, & Grimley RP (1988b) Moles in Australian and British
schoolchildren (Letter to the Editor). Lancet, ii: 1497.
Green C, Diffey BL, & Hawk JLM (1992) Ultraviolet radiation in the
treatment of skin disease. Phys Med Biol, 37(1): 1-20.
Green A, Smith P, McWhirter W, Oregan P, Battistutta D, Yarker ME, &
Lape K (1993a) Melanocytic naevi and melanoma in survivors of
childhood cancer. Br J Cancer, 67: 1053-1057.
Green A, MacLennan R, Youll P, & Martin N (1993b) Site distribution
of cutaneous melanoma in Queensland. Int J Cancer, 53: 232-236.
Greenberg JT, Monach PA, Chou JH, Josephy PD, & Demple B (1990)
Positive control of a global antioxidant defence regulon activated
by superoxide-generating agents in Escherichia coli. Proc Natl Acad
Sci (USA), 87: 6181-6185.
Greene MH & Wilson J (1985) Second cancer following lymphatic and
hematopoietic cancers in Connecticut, 1935-82. Natl Cancer Inst
Monogr, 68: 191-217.
Greene MH, Young TI, & Clark WH Jr (1981) Malignant melanoma in
renal transplant recipients. Lancet, May 30: 1196-1199.
Grob JJ, Gouvernet J, Aymar D, Mostaque A, Romano MH, Collet AM, Noe
MC, Diconstanzo MP, & Bonerandi JJ (1990) Count of benign
melanocytic nevi as a major indicator of risk for nonfamilial
nodular and superficial spreading melanoma. Cancer, 66: 387-395.
Guerry RK, Ham WT, & Mueller HA (1985) Light toxicity in the
posterior segment. In: Duane TD & Jaeger EA ed. Clinical
ophthalmology. Philadelphia, Pennsylvania, Harper and Row, pp 1-17.
Guex-Crosier Y & Herbort CP (1993) Presumed corneal intraepithelial
neoplasia associated with contact lens wear and intense ultraviolet
light exposure. Br J Ophthalmol, 77: 191-192.
Gupta AK, Cardella CJ, & Haberman HF (1986) Cutaneous malignant
neoplasms in patients with renal transplants. Arch Dermatol, 122:
1288-1293.
Gupta AK, Stern RS, Swanson NA, Anderson TF, & Arbor A (1988)
Cutaneous melanomas in patients treated with psoralens plus
ultraviolet A. J Am Acad Dermatol, 19: 67-76.
Gutteridge JMC (1985) Superoxide dismutase inhibits the
superoxide-driven Fenton reaction at two different levels. FEBS
Lett, 185: 19-23.
Häder DP & Worrest RC (1991) Effects of enhanced solar radiation on
aquatic ecosystems. Photochem Photobiol, 53: 717-725.
Häder DP & Lui SM (1991) Mobility and gravitactic orientation of the
flagellate, Euglena gracilis, impaired by artificial and solar UVB
radiation. Curr Microbiol, 21: 161-168.
Haenszel W (1963) Variations in skin cancer incidence within the
United States. Natl Cancer Inst Monogr, 10: 225-243.
Halprin KM, Comerford M, Presser SE, & Taylor JR (1981) Ultraviolet
light treatment delays contact sensitization to nitrogen mustard. Br
J Dermatol, 105: 71-76.
Ham WT, Mueller HA, Ruffolo JJ, Millen JE, Cleary SF, & Guerry RK
(1984) Basic mechanisms underlying the production of photochemical
lesions in the mammalian retina. Curr Eye Res, 3: 165-174.
Hanchette CL & Schwartz GC (1992) Geographic patterns of
prostatecancer mortality. Evidence for a protective effect of
ultraviolet radiation. Cancer, 70: 2861-2869.
Hardie IR, Strong RW, Hartley LCJ, Woodruff PWH, & Clunie GJA (1980)
Skin cancer in Caucasian renal allograft recipients living in a
subtropical climate. Surgery, 87: 177-183.
Hartevelt MM, Bouwes Bavinck JN, Kootte AMM, Vermeer BJ, &
Vandenbroucke JP (1990) Incidence of skin cancer after renal
transplantation in the Netherlands. Transplantation, 49: 506-509.
Hawk, JLM (1984) Photosensitising agents used in the United Kingdom.
Clin Expl Dermatol, 9: 300-302.
Hayashi Y & Aurelian L (1986) Immunity to herpes simplex virus type
2: Viral antigen-presenting capacity of epidermal cells and its
impairment by ultraviolet irradiation. J Immunol, 136: 1087-1092.
Haynes RH (1966) The interpretation of microbiol inactivation and
recovery phenomena. Radiat Res, Suppl 6: 1-29.
Health Council of the Netherlands (1986) UV radiation: Human
exposure to ultraviolet radiation. The Hague, Health Council of the
Netherlands (Report 1986/93).
Hedblom EE (1961) Snowscape eye protection. Arch Environ Health, 2;
685-704.
Hefferren JJ, Cooley RO, Hall JB, Olsen NH, & Lyon HW (1971) Use of
ultraviolet illumination in diagnosis. J Am Dent Assoc, 82:
1353-1360.
Herity B, O'Loughlin G, Moriarty MJ, & Conroy R (1989) Risk factors
for non-melanoma skin cancer. Ir Med J, 82: 151-152.
Hersey P, Hasic E, Edwards A, Bradley M, Maran G, & McCarthy WH
(1983a) Immunological effects of solarium exposure. Lancet, 1:
545-548.
Hersey P, Haran G, Hasic E, & Edwards A (1983b) Alteration of T cell
subsets and induction of suppressor T cell activity in normal
subjects after exposure to sunlight. J Immunol, 31: 171-174.
Hersey P, MacDonald M, Burns C, Schibeci S, Matthews H, & Wilkinson
FJ (1987) Analysis of the effect of sunscreen agent on the
suppression of natural killer cell activity induced in human
subjects by radiation from solarium lamps. J Invest Dermatol, 88:
271-276.
Hersey P, MacDonald M, Henderson C, Schibeci S, D'Alessandro G,
Pryor M, & Wilkinson FJ (1988) Suppression of natural killer cell
activity in humans by radiation from solarium lamps depleted of UVB.
J Invest Dermatol, 90: 305-310.
Hess C (1907) Experiments on the effect of ultraviolet light on the
lens. Arch f Augenheilk 57 185-196.
Higginson J & Oettlé AG (1959) Cancer incidence in the Bantu and
"Cape Colored" races of South Africa: Report of a cancer survey in
the Transvaal (1953-55). J Natl Cancer Inst, 24: 589-671.
Hill D, White V, Marks R, Theobald T, Borland R, & Roy C (1992)
Melanoma prevention: behavioural and nonbehavioural factors in
sunburn among an Australian urban population. Prev Med, 21: 654-669.
Hiller R, Sperduto RD, & Ederer F (1977a) Epidemiological
associations with cataract in the 1971-72 national health and
nutrition examination survey. Am J Epidemiol, 105: 450-459.
Hiller R, Giacometti L, & Yuen K (1977b) Sunlight and cataract: an
epidemiologic investigation. Am J Epidemiol, 105: 450-459.
Hiller R, Sperduto RD, & Ederer F (1983) Epidemiologic associations
with cataract in the 1971-1972 national health and nutrition
examination survey. Am J Epidemiol, 118: 239-249.
Hiller R, Sperduto RD, & Ederer F (1986) Epidemiologic associations
with nuclear, cortical, and posterior subcapsular cataracts. Am J
Epidemiol, 124: 916-925.
Hirschfeld S, Levine AS, Ozato K, & Protic M (1990) A constitutive
damage-specific DNA binding protein is synthesized at higher levels
in UV-irradiated primate cells. Mol Cell Biol, 10: 2041-2048.
Hoeijmakers JHJ & Bootsma D (1990) Molecular genetics of eucaryotic
DNA excision repair. Cancer Cells, 2: 311-320.
Hoffman JS (1987) Ultraviolet radiation and melanoma (with a special
focus on assessing the risks of stratospheric ozone depletion).
Washington, DC, US Environmental Protection Agency, Office of Air
and Radiation (EPA 400/1-87-001D).
Hogan DJ, To T, Gran L, Wong D, & Lane PR (1991) Risk factors for
basal cell carcinoma. Int J Dermatol, 28: 591-594.
Holick MF (1985) The photobiology of vitamin D and its consequences
for human. Ann N Y Acad Sci, 453: 1-3.
Hollows F & Moran D (1981) Cataract - the ultraviolet risk factor.
Lancet, 2: 1249-1250.
Holly EA, Kelly JW, Shpall SN, & Chiu S-H (1987) Number of
melanocytic nevi as a major risk factor for malignant melanoma. J Am
Acad Dermatol, 17: 459-468.
Holly EA, Aston DA, Char DH, Kristiansen JJ, & Ahn DK (1990) Uveal
melanoma in relation to ultraviolet light exposure and host factors.
Cancer Res, 50: 5773-5777.
Holly EA, Aston DA, Ahn DK, Kristiansen JJ, & Char DH (1991) No
excess prior cancer in patients with uveal melanoma. Ophthalmology,
98: 608-611.
Holm-Hansen O (1990) UV radiation in Arctic waters: Effects on rates
of primary production (Appendix G). La Jolla, CA, USA, Scripps
Institution of Oceanography, pp 1-17.
Holman CDJ & Armstrong BK (1984) Pigmentary traits, ethnic origin,
benign nevi, and family history as risk factors for cutaneous
malignant melanoma. J Natl Cancer Inst, 72: 257-266.
Holman CDJ, Gibson IM, Stephenson M, & Armstrong BK (1983)
Ultraviolet irradiation of human body sites in relation to
occupation and outdoor activity: field studies using personal UVR
dosimeters. Clin Exp Dermatol, 8: 269-277.
Holman CDJ, Armstrong BK, Evans PR, Lumsden GJ, Dallimore KJ, Meehan
CJ, Beagley J, & Gibson IM (1984a) Relationship of solar keratosis
and history of skin cancer to objective measures of actinic skin
damage. Br J Dermatol, 110: 129-138.
Holman CD, Evans PR, Lumsden GJ, & Armstrong BK (1984b) The
determinants of actinic skin damage: problems of confounding among
environmental and constitutional variables. Am J Epidemiol, 120:
414-422.
Holman CDJ, Armstrong BK, & Heenan PJ (1986) Relationship of
cutaneous malignant melanoma to individual sunlight-exposure habits.
J Natl Cancer Inst, 76: 403-414.
Hoover R (1977) Effects of Rugs - Immunosuppression. In: Hiatt HH,
Watson JD, & Winsten JA ed. Origins of human cancer. Book A:
Incidence of cancer in humans. Proceedings of the Cold Spring Harbor
Conferences on Cell Proliferation. Cold Spring Harbor, New York,
Cold Spring Harbor Laboratory, vol 4.
Hoppeler Th, Hendrickson Ph, Dietrich C, & Reme (1988) Morphology
and time-course of defined photochemical lesions in the rabbit
retina. Curr Eye Res, 7(9): 849-860.
Howie S, Norval M, & Maingay J (1986) Exposure to low-dose
ultraviolet radiation suppresses delayed-type hypersensitivity to
herpes simplex virus in mice. J Invest Dermatol, 86: 125-128.
Howie SEM, Ross JA, Norval M, & Maingay JP (1986) In vivo
modulation of antigen presentation generates Ts rather than TDH in
HSV-1 infection. Immunology, 60: 419-423.
Hsu J, Forbes PD, Harber LC, & Lakow E (1975) Induction of skin
tumors in hairless mice by a single exposure to UV radiation.
Photochem Photobiol, 21: 185-188.
Hughes BR, Cunliffe WJ, & Bailey CC (1989) Excess benign melanocytic
naevi after chemotherapy for malignancy in childhood. Br Med J, 299:
88-91.
Hunter DJ, Colditz GA, Stampfer MJ, Rosner B, Willett WC, & Speizer
FE (1990) Risk factors for basal cell carcinoma in a prospective
cohort of women. Ann Epidemiol, 1: 13-23.
Hunter DJ, Colditz GA, Stampfer MJ, Rosner B, Willett WC, & Speizer
FE (1992) Diet and risk of basal cell carcinoma of the skin in a
prospective cohort of women. Ann Epidemiol, 2: 231-239.
Husain Z, Yang Q, & Biswas DK (1990) cHa-ras Proto oncogene.
Amplification and overexpression in UVB-induced mouse skin
papillomas and carcinomas. Arch Dermatol, 126: 324-330.
Husain Z, Pathak MA, Flotte T, & Wick MM (1991) Role of ultraviolet
radiation in the induction of melanocytic tumors in hairless mice
following 7,12-dimethylbenz(a)anthracene application and ultraviolet
irradiation. Cancer Res, 51: 4964-4970.
Hyman LG, Lilienfeld AM, Ferris FL, & Fine SL (1983) Senile macular
degeneration: a case-control study. Am J Epidemiol, 118: 213-227.
IARC (1993) In: Kricker A, Armstrong BK, Jones ME & Burton RD ed.
Health, solar UV radiation and environmental change. Lyon,
International Agency for Research on Cancer (IARC Technical Report
No. 13).
IARC (1992) Solar and ultraviolet radiation. Lyon, International
Agency for Research on Cancer (Monographs on the Evaluation of
Carcinogenic Risks to Humans, Volume 55).
IARC (1980) International Agency for Research on Cancer, Monographs
on the Evaluation of the Carcinogenic Risk of Chemicals to Humans,
Vol 24, Some Pharmaceutical Drugs, Lyon, 101-124.
Ignatiades L (1990) Photosynthetic capacity of the surface
microlayer during the mixing period. J Planktonic Res, 12: 851-860.
Imlay JA, Chir SM, & Linn S (1988) Science, 240: 640-642.
IRPA/INIRC (1985) International Radiation Protection Association/
International Non-Ionizing Radiation Committee. Concepts, units and
terminology for NIR protection. Health Phys, 49(6): 1329-1362.
IRPA/INIRC (1991a) International Radiation Protection Association/
International Non-Ionizing Radiation Committee. Health issues of
ultraviolet A sunbeds used for cosmetic purposes. Health Phys,
61(2): 285-288.
IRPA/INIRC (1991b) International Radiation Protection Association/
International Non-Ionizing Radiation Committee. In: Duchêne AS,
Lakey JRA, & Repacholi MH ed. IRPA guidelines on protection against
non-ionizing radiation., New York, McGraw-Hill.
Isaacson C, Selzer G, Kaye V, Greenberg M, Woodruff JD, Davies J,
Ninin D, Vetten D, & Andrew, M (1978) Cancer in the urban blacks of
South Africa. S Afr Cancer Bull, 22: 49-84.
Ishizaki K, Tsujimura T, Nakai M, Nishigori C, Sato K, Katayama S,
Kurimura O, Yoshikawa K, Imamura S, & Ikenaga M (1992) Infrequent
mutation of the ras genes in skin tumors of Xeroderma Pigmentosum
patients in Japan. Int J Cancer, 50: 382-385.
Italian-American Cataract Study Group (1991) Risk factors for
age-related cortical, nuclear, and posterior subcapsular cataracts.
Am J Epidemiol, 133: 541-553.
Ito A & Ito T (1983) Possible involvement of membrane damage in the
inactivation by broad-band near-UV radiation in Saccharomyces
cerevisiae cells. Photochem Photobiol 37: 395-401.
Iversen OH (1988) Skin tumorigenesis and carcinogenesis studies with
7,12-dimethylbenz[a]anthracene, ultraviolet light, benzoyl peroxide
(Panoxyl gel 5%) and ointment gel. Carcinogenesis, 9: 803-809.
Jagger J (1985) Solar-UV actions on living cells. New York, Praeger.
Jeevan A & Kripke ML (1990) Alteration of the immune response to
Mycobacterium bovis BCG in mice exposed chonically to low doses of
UV radiation. Cell Immunol, 130: 32-41.
Jeevan A, Evans R, Brown EL, & Kripke ML (1992a) Effect of local
ultraviolet irradiation on infections of mice with Candida albicans,
Mycobacterium bovis BCG, and Schistosoma mansoni. J Invest Dermatol,
99: 59-64.
Jeevan A, Gilliam K, Heard H, & Kripke ML (1992b) Effects of
ultraviolet radiation on the pathogenesis of Mycobacterium
lepraemurium infection in mice. Exp Dermatol, 1: 152-160.
Jeevan A, Ullrich SE, Dizon V, & Kripke ML (1992c) Supernantants
from ultraviolet-irradiated keratinocytes decrease the resistance
and delayed-type hypersensitivity response to Mycobacterium bovis
bacillus Calmette-Guerin in mice and impair the phagocytic ability
of macrophages. Photodermatol Photoimmunol Photomed, 9: 255-263.
Jeffery SW & Humphrey GH (1975) New spectrophotometric equations for
determining chlorophylls a, b, c1 and C2 in higher plants,
algae, and natural phytoplankton. Biochem Physiol Pflanz, 167:
191-194.
Jensen OM & Bolander (1980) Trends in malignant melanoma of the
skin, WHO Stat Q, 33: 2-26.
Johnson GJ (1981) Aetiology of spheroidal degeneration of the cornea
in Labrador. Br J Ophthalmol, 65: 270-283.
Johnson GJ & Overall M (1978) Histology of spheroidal degeneration
of the cornea in Labrador. Br J Ophthalmol, 62: 53-61.
Johnson GJ, Paterson GD, Green JS, & Perkins ES (1981) Ocular
conditions in a labrador community In: Harvald B & Hansen JP ed.
Circumpolar health 81. Copenhagen, Nordic Council for Arctic Medical
Research.
Johnson GJ, Minassian DC, & Franken S (1989) Alterations of the
anterior lens capsule associated with climatic keratopathy. Br J
Ophthalmol, 73: 229-234
Jones CA, Huberman E, Cunningham ML, & Peak MJ (1987a) Mutagenesis
and cytotoxicity in human epithelial cells by far- and
near-ultraviolet radiations : action spectra. Radiat Res, 110:
244-254.
Jones SK, Moseley H, & Mackie RM (1987b) UVA-induced melanocytic
lesions. Br J Dermatol, 117: 111-115.
Jones ME, Shugg D, Dwyer T, Young B, & Bonett A (1992) Interstate
differences in incidence and mortality from melanoma: a
re-examination of the latitudinal gradient. Med J Aust, 157:
373-377.
Jose JG (1986) Posterior cataract induction by UVB radiation in
albino mice. Exp Eye Res, 42: 11-20.
Jose JG & Pitts DG (1985) Wavelength dependency of cataracts in
albino mice following chronic exposure. Exp Eye Res, 41: 545-563.
Kalimo K, Loulu L, & Jansen CT (1983) Effect of a single UVB or PUVA
exposure on immediate and delayed skin hypersensitivity reactions in
humans. Correlation to erythemal response and Langerhans cell
depletion. Arch Dermatol Res, 275: 374-378.
Karai I & Horiguchi S (1984) Pterygium in welders. Br J Ophthalmol,
68: 347-349.
Karentz D & Lutze LH (1990) Evaluation of biologically harmful
ultraviolet radiation in Antarctica with a biological dosimeter
designed for aquatic environments. Limnol Oceanogr, 35: 549-561.
Karentz D, Mc Euen FS, Land MC, & Dunlap WC (1991) Survey of
microsporine-like amino acid compounds in Antarctic marine
organisms: potential protection from ultraviolet exposure. Mar Biol,
108: 157-166.
Karjalainen S, Salo H, & Teppo L (1989) Basal cell and squamous cell
carcinoma of the skin in Finland Int J Dermatol, 28: 445-450.
Kataoka H & Fujiwara Y (1991) UV damage specific protein in
xeroderma pigmentosum complementation group E. Biochem Biophys Res
Commun, 175: 1139-1143.
Katz L, Ben-Tuvia S, & Steinitz R (1982) Malignant melanoma of the
skin in Israel: effect of migration. In: Magnus K ed Trends in
cancer incidence: Causes and practical implication. Washington, New
York, Hemisphere, pp 419-426.
Kelfkens G, van Weelden H, de Gruijl FR, & van der Leun JC (1991)
Influence of dose rate in ultraviolet tumorigenesis. J Photochem
Photobiol, B10: 41-50.
Kelfkens G & van der Leun JC (1989) Skin temperature changes after
irradiation with UVB or UVC: Implications for the mechanism
underlying ultraviolet erythema. Phys Med Biol, 34: 599-608.
Keller, AZ (1970) Cellular types, survival, race, nativity,
occupations, habits and associated diseases in the pathogenesis of
lip cancers. Am J Epidemiol, 91, 486-499.
Kelly, GE, Meikle, WD, & Moore, DE (1989) Enhancement of UV-induced
skin carcinogenesis by azathioprine: role of photochemical
sensitisation. Photochem Photobiol, 49, 59-65.
Kelly GE, Meikle WD, & Sheil AGR (1987) Effects of immunosuppressive
therapy on the induction of skin tumors by ultraviolet irradiation
in hairless mice. Transplantation, 44, 429-434.
Kennedy JC, Poltier RH, & Pross DC (1990) Photodynamic therapy with
endogenous protoporphyrin. IX. Basic principles and present clinical
experience. J Photochem Photobiol, 6: 143-148.
Keyse SM (1993) The induction of gene expression in mammalian cells
by radiation. Sem Cancer Biol, 4: 119-128.
Keyse SM & Tyrrell RM (1989) Heme oxygenase is the major 32-kDa
stress protein induced in human skin fibroblasts by UVA radiation,
hydrogen peroxide and sodium arsenite. Proc Natl Acad Sci (USA), 86:
99-103.
Khlat M, Vail A, Parkin M, & Green A (1992) Mortality from melanoma
in migrants to Australia: variation by age at arrival and duration
of stay. Am J Epidemiol, 135: 1103-1113.
Kim TY, Kripke ML, & Ullrich SE (1990) Immunosuppression by factors
released from UV-irradiated epidermal cells: Selective effects on
the generation of contact and delayed hypersensitivity after
exposure to UVA or UVB radiation. J Invest Dermatol, 94: 26-32.
Kinlen LJ, Sheil AGR, Peto J, & Doll R (1979) Collaborative United
Kingdom-Australasian study of cancer in patients treated with
immunosuppressive drugs. Br Med J, 2: 1461-1466.
Klamen DK & Tuveson RW (1982) The effect of membrane fatty acid
compositions on the near-UV (300-400 nm) sensitivity of Escherichia
coli K1060. Photochem Photobiol, 35: 167-173.
Klepp O & Magnus K (1979) Some environmental and bodily
characteristics of melanoma patients. A case-control study. Int J
Cancer, 23: 482-486.
Kligman LH, Kaidbey KH, Hitchens VM, & Miller SA (1987) Long
wavelength (>340 nm) ultraviolet-A induced damage in hairless mice
is dose dependent. In: Passchier W & Bosnjakovi, BFM ed., Human
exposure to ultraviolet radiation: Risks and regulations. Amsterdam,
Oxford, New York, Elsevier Science Publishers, pp 77-81.
Knekt P, Aromaa A, Maatela J, Alfthan G, Aaran RK, Nikkari T, Hakama
M, Hakulinen T, & Teppo, L (1991) Serum micronutrients and risk of
cancers of low incidence in Finland. Am J Epidemiol, 134: 356-361.
Kollias, N, Sayre RM, Zeise L, & Cehedekel MR (1991) Photoprotection
by melanin, J Photochem Photobiol, 9: 135-160
Kopecky KE, Pugh GW Jr, Hughes DE, Booth GD, & Cheville, NF (1979)
Biological effect of ultraviolet radiation on cattle: bovine ocular
squamous cell carcinoma. Am J Vet Res, 40: 1783-1788.
Kopf AW, Lazar M, Bart RS, Dubin N, & Bromberg J (1978) Prevalence
of nevocytic nevi on lateral and medial aspects of arms. J Dermatol
Surg Oncol, 4: 153-158.
Kopf AW, Lindsay AC, Rogers GS, Friedman RJ, Rigel DS, & Levenstein
M (1985) Relationship of nevocytic nevi to sun exposure in
dysplastic nevus syndrome. J Am Acad Dermatol, 12: 656-662.
Kraemer KH, Lee MM, & Scotto J (1987) Xeroderma Pigmentosum Arch
Dermatol, 123: 241-50.
Kraemer KH, Seetharam S, Protic-Sabljic M, Brash DE, Bredberg A, &
Seidman MM (1988) Defective DNA repair and mutagenesis by dimer and
non-dimer photoproducts in xeroderma pigmentosum measured with
plasmid vectors. In: Freidberg EC & Hanawalt PC ed. Mechanisms and
consequences of DNA damage processing. New York, Alan R. Liss, pp
325-335.
Kramer GF & Ames BN (1987) Oxidative mechanisms of toxicity of
low-intensity near-UV light in Salmonella typhimurium. J Bacteriol,
164: 2259-2266.
Kremers JJM & van Norren D (1988) Two classes of photochemical
damage to the retina. In. Lasers and light in ophthalmology.
Amsterdam, Berkeley, Milano, Klugler Publications, vpl 2, pp 41-52.
Kress S, Sutter C, Strickland PT, Mukhtar H, Schweizer J, & Schwarz
M (1992) Carcinogen-specific mutational pattern in the p53 gene in
ultraviolet B radiation-induced squamous cell carcinomas of mouse
skin. Cancer Res, 52: 6400-6403.
Kricker A, Armstrong BK, English DR, & Heenan PJ (1991a) Pigmentary
and cutaneous risk factors for non-melanocytic skin cancer - a
case-control study. Int J Cancer, 48: 650-662.
Kricker A, Armstrong BK, English D, Heenan PJ, & Randell PL (1991b)
A case-control study of non-melanocytic skin cancer and sun exposure
in Western Australia (Abstract No. III, P2). Cancer Res Clin Oncol,
117(Suppl II): S75.
Kricker A, English DR, Randell PL, Heenan PJ, Clay CD, Delaney TA, &
Armstrong BK (1990) Skin cancer in Geraldton, Western Australia: a
survey of incidence and prevalence. Med J Aust, 152: 399-407.
Kricker A, Armstrong BK, Jones ME, & Burton RC (1993) Health, solar
UV radiation and environmental change. IARC Technical Report No 13.
Lyon, International Agency for Research on Cancer.
Kripke ML (1974) Antigenicity of murine skin tumors induced by
ultraviolet light. J Natl Cancer Inst, 53: 1333-1336.
Kripke Ml & Fidler IJ (1980) Enhanced experimental metastasis of
ultraviolet light-induced fibrosarcomas in ultraviolet
light-iradiated syngeneic mice. Cancer Res, 40: 625-629.
Kripke ML & Fisher MS (1976) Immunologic parameters of ultraviolet
carcinogenesis. J Natl Cancer Inst, 57: 211-215.
Kripke ML & Sass ER ed. (1978) International Conference on
Ultraviolet Carcinogenesis. Natl Cancer Inst Monogr, 50: 16-17.
Kripke ML, Lofgreen JS, Beard J, Jessup JM, & Fisher MS (1977) in
vivo immune responses of mice during carcinogenesis by ultraviolet
irradiation. J Natl Cancer Inst, 59: 1227-1230.
Kromberg JG & Jenkins T (1982) Prevalence of albinism in the South
African Negro. S Afr Med J, 61: 383-386.
Kromberg JGR, Castle D, Zwane EM, & Jenkins T (1989) Albinism and
skin cancer in southern Africa. Clin Genet, 36: 43-52.
Kulandaivelu G, Neducheshian N & Annamalainathan K (1993)
Ultraviolet-B (280-320) radiation induced changes in photochemical
activities and polypeptide components of C3 and C4 chloroplasts.
Photosynthetica, 25:12-14.
Kune GA, Bannerman S, Field B, Watson LF, Clelan H, Merenstein D, &
Vitetta L (1992) Diet, alcohol, smoking, serum ß-carotene, and
vitamin A in male nonmelanocytic skin cancer patients and controls.
Nutr Cancer, 18: 237-244.
Lancaster HO & Nelson J (1957) Sunlight as a cause of melanoma: a
clinical survey. Med J Aust, 1: 452-456.
Langer B & Wellmann E (1990) Phytochrome induction of
photoreactivation in Phaseolus vulgaris L. seedlings. Photochem
Photobiol, 52: 861-864.
Larko O & Diffey BL (1983) Natural UVB radiation received by people
with outdoor, indoor and mixed occupations and UVB treatment of
psoriasis. Clin Exp Dermatol, 8: 279-285.
Lassam NJ, From L, & Kahn HJ (1993) Overexpression of p53 is a late
event in the development of malignant melanoma. Cancer Res, 53:
2235-2238.
Laycock KA, Lee SF, Brady RH, & Pepose JS (1991) Characterization of
a murine model of recurrent herpes simples viral keratitis induced
by ultraviolet B radiation. Invest Ophthalmol Vis Sci, 32:
2741-2746.
Lê, MG, Cabanes, PA, Desvignes, V, Chanteau, MF, Mlika, N, & Avril,
MF (1992) Oral contraceptive use and risk of cutatneous malignant
melanoma in a case-control study in French women. Cancer Causes
Control, 3, 199-205.
Leach, JF, McLeod, VE, Pingstone, AR, Davis, A and Deane, GHW (1978)
Measurement of the ultraviolet doses received by office workers.
Clinical and Experimental Dermatology, 3,77-79.
Leclerc, JE, Borden, A and Lawrence, CW (1991) The thymine-thymine
pyrimidine-pyrimidine (6-4) ultraviolet light photoproduct is highly
mutagenic and specifically induces 3'-thymine-to-cytosine transition
in Escherichia coli Proc Natl Acad Sci USA, 88: 9685-9689.
Lee JAM (1989) The relationship between malignant melanoma of the
skin and exposure to sunlight. Photochem Photobiol, 50: 493-496.
Lee GA & Hirst LW (1992) Incidence of ocular surface epithelial
dysplasia in metropolitan Brisbane: a 10-year survey. Arch
Ophthalmol, 110: 525-527.
Lerman S (1980) Human ultraviolet radiation cataracts. Ophthalmic
Res, 12: 303-314.
Leske MC, Chylack LT, Wu S, & The Lens Opacities Case-Control Study
Group (1991) The lens opacities case-control study: risk factors for
cataract. Arch Ophthalmol, 109: 244-251.
Levi F, La Vecchia C, Te V-C, & Mezzanotte G (1988) Descriptive
epidemiology of skin cancer in the Swiss canton of Vaud. Int J
Cancer, 42: 811-816.
Levine EA, Ronan SG, Shirali SS, & Das Gupta TK (1992) Malignant
melanoma in a child with oculocutaneous ablinism. J Surg Oncol, 51:
138-142.
Lew RA, Sober AJ, Cook N, Marvell R, & Fitzpatrick TB (1983) Sun
exposure habits in patients with cutaneous melanoma: a case control
study. J Dermatol Surg Oncol, 9: 981-986.
Lewis MG (1967) Malignant melanoma in Uganda (The relationship
between pigmentation and malignant melanoma on the soles of the
feet). Br J Cancer, 21: 483-495.
Ley RD (1993) Photoreactivation in humans. Proc Natl Acad Sci (USA),
90: 4337.
Ley RD Peak MJ, & Lyon LL (1983) Induction of pyrimidine dimers in
epidermal DNA of hairless mice by UVB: an action spectrum. J Invest
Dermatol, 80: 188-191.
Ley RD (1985) Photoreactivation of UV-Induced pyrimidine dimers and
erythema in the marsupial Monodelphis domestica. Proc Natl Acad Sci
(USA), 82: 2409-2411.
Ley RD, Applegate LA, Stuart TD, & Fry RJM (1987) UV
radiation-induced skin tumors in Monodelphis domestica.
Photodermatology, 4: 144-147.
Ley RD, Applegate LA, Padilla RS, & Stuart TD (1989) Ultraviolet
radiation-induced malignant melanoma in Monodelphis domestica.
Photochem Photobiol, 50: 1-5.
Ley RD, Applegate LA, Fry RJM, & Sanchez AB (1991) Photoreactivation
of ultraviolet radiation-induced skin and eye tumors of Monodelphis
domestica. Cancer Res, 51: 6539-6542.
Li YF, Kim ST, & Sancer A (1993) Evidence for lack of DNA
photoreactivity enzyme in humans. Proc Natl Acad Sci (USA), 90:
4389-4393.
Lieu F-M, Yamanishi K, Jonishi K, Kishimoto S, & Yasuno, H (1991)
Low incidence of Ha-ras oncogene mutations in human epidermal
tumors. Cancer Lett, 59: 231-235.
Lill PH (1983) Latent period and antigenicity of murine tumors
induced in C3H mice by short-wavelength ultraviolet radiation. J
Invest Dermatol, 81: 342-346.
Lindberg JG (1917) Clinical study on iris depigmentation and on
transparency of iris in cataract and in normal eyes among elderly
persons. Dissertation, Helsinki, Finland (in Swedish).
Lindelöf B, Sigurgeirsson B, Tegner E, Larkö O, Johannesson A, Berne
B, Christensen OB, Andersson T, Törngren M, Molin L,
Nylander-Lundqvist E, & Emtestam L (1991) PUVA and cancer; a
large-scale epidemiological study. Lancet, 338: 91-93.
Lindqvist C (1979) Risk factors in lip cancer: a questionnaire
survey. Am J Epidemiol, 109: 521-530.
Lippke JA, Gordon LK, Brash D, & Haseltine WA (1981) Distribution of
UV light-induced damage in a defined sequence of human DNA:
Detection of alkaline-sensitive lesions at pyrimidine
nucleoside-cytidine sequences. Proc Natl Acad Sci (USA), 78:
3388-3392.
Lischko AM, Seddon JM, Gragoudas ES, Egan KM, & Glynn RJ (1989)
Evaluation of prior primary malignancy as a determinant of uveal
melanoma. A case-control study. Ophtalmology, 96: 1716-1721.
Logani MK, Sambuco CP, Forbes PD, & Davies RE (1984) Skin-tumour
promoting activity of methyl ethyl ketone peroxide - a potent
lipid-peroxidizing agent. Food chem Toxicol, 22: 879-882.
Luande J, Henschke CI, & Mohammed N (1985) The Tanzanian human
albino skin. Cancer, 55: 1823-1828.
Lubin D, Frederick JE, Booth CR, Lucas T, & Neuschuler D (1989)
Measurements of enhanced springtime ultraviolet radiation at Palmer
Station, Antarctica. Geophys Res Lett, 16: 783-785.
Lynch DH & Daynes RA (1983) Evaluation of naturally occurring
cell-mediated cytotoxic activity in normal and UV-irradiated mice.
Transplantation, 35: 216-223.
Lynge E & Thygesen L (1990) Occupational cancer in Denmark. Cancer
incidence in the 1970 census population. Scand J Work Environ
Health, 16(Suppl 2): 1-35.
Lytle CD, Miller SA, Jacobs ME, Cyr WH, James RH, Kaczmarek RG, Beer
JZ, Landry RJ, Sharkness CM, Gaylor D, & De Gruijl FR (1993) An
estimation of squamous cell carcinoma risk from ultraviolet
radiation emitted by fluorescent lamps. In : Proceedings of the 1993
Meeting of the American Society for Photobiology, Chicago, 26-30
June 1993.
McCormick JP, Fisher JR, Pachlatko JP, & Eisenstark A (1976)
Characterization of a cell lethal product from the photooxidation of
tryptophan: hydrogen peroxide. Science, 198: 468-469.
McCredie M & Coates MS (1989) Cancer incidence in migrants to New
South Wales; 1972 to 1984, New South Wales Central Cancer Registry.
Woolloomooloo, New South Wales Cancer Council, pp 22-23, 62-63.
MacDonald EJ (1948) Malignant melanoma in Connecticut. In: The
biology of melanomas. New York, Academy of Sciences, pp 71-81
(Special publication of the Academy of Sciences No. 4).
McGregor JM, Barker JNWN, & MacDonald DM (1992) The development of
excess numbers of melanocytic naevi in an immunosuppressed identical
twin. Clin Exp Dermatol, 16: 131-132.
Mack TM, & Floderus B (1991) Malignant melanoma risk by nativity,
place of residence at diagnosis, and age at migration. Cancer Causes
Control, 2: 401-411.
Mackenzie FD, Hirst LW, Battistutta D, & Green A (1992) Risk
analysis in the development of pterygia. Ophthalmol, 99: 1056-1061.
MacKie RM & Aitchison T (1982) Severe sunburn and subsequent risk of
primary cutaneous malignant melanoma in Scotland. Br J Cancer, 40:
55-960.
MacKie RM, Freudenberger T, & Aitchison TC (1989) Personal
risk-factor chart for cutaneous melanoma. Lancet, ii: 487-490.
McKinlay AF (1992) Artificial sources of UVA radiation. Loss and
emission characteristics. In: Urbach F ed Biological responses to
UVA radiation. Vandermor Pub Corp Kansas.
McKinlay AF & Diffey BL (1987) A reference action spectrum for
ultra-violet induced erythema in human skin. CIE J, 6: 17-22
McKinlay AF & Whillock MJ (1987) Measurements of ultraviolet
radiation from fluorescent lamps used for general lighting and other
purposes in the UK. In: Passchhier WF & Bosnjakovic BFM ed. Human
exposure to ultraviolet radiation: Risks and regulations.,
Amsterdam, Oxford, New York, Elsevier Science Pubishers, pp 253-258.
McKinlay AF, Whillock MJ, & Meulemans CCE (1989) Ultraviolet
radiation and blue-light emissions from spotlights incorporating
tungsten halogen lamps. London, National Radiological Protection
Board (Report NRPB No. R228).
McKinlay AF, Harlen F, & Whillock MJ (1988) Hazards of optical
radiation: A guide to sources, uses and safety. Bristol,
Philadelphia, Adam Hilger.
Madewell BR, Conroy JD, & Hodgkins EM (1981) Sunlight-skin cancer
association in the dog: a report of three cases. J Cutan Pathol, 8:
434-443.
Magnus K (1977) Incidence of malignant melanoma of the skin in 5
Nordic countries: significance of solar radiation. Int J Cancer, 20:
477.
Magnus K (1986) Malignant melanoma in Norway. Tidsskr Nor
Lacgeforen, 106: 2309-2313.
Magnus K (1991) The Nordic profile of skin cancer incidence. A
comparative epidemiological study of the three main types of skin
cancer. Int J Cancer, 47: 12-19.
Mandal TK & Chatterjee SN (1980) Ultraviolet- and sunlight-induced
lipid peroxidation in liposomal membrane. Radiat Res, 83: 290-302.
Mao W & Hu T (1982) An epidemiologic survey of senile cataract in
China. Chin Med J, 95: 813-818.
Marks R & Hill D ed. (1992) The public health approach to melanoma
control: Prevention and early detection. Geneva, International Union
Against Cancer (UICC Publication).
Marks R, Ponsford MW, Selwood TS, Goodman G, & Mason G (1983)
Non-melanocytic skin cancer and solar keratoses in Victoria. Med J
Aust, 2: 619-622.
Marks R, Jolley D, Dorevitch AP, & Selwood TS (1989) The incidence
of non-melanocytic skin cancers in an Australian population: results
of a five-year prospective study. Med J Aust, 150: 475-478.
Marks R, Staples M, & Giles GG (1993) Trends in non melanocytic skin
cancer treated in Australia: the second national survey. Int J
Cancer, 53: 585-590.
Martin EK (1912) The effects of ultra-violet rays upon the eye. Proc
of the Soc Ser B, 85: 319-334.
Mathews-Roth MM & Krinsky NI (1987) Carotenoids affect development
of UV-B induced skin cancer. Photochem Photobiol, 46: 507-509.
Matsunaga T, Hieda K, & Nikaido O (1991) Wavelength dependent
formation of thymine dimers and (6-4) photoproducts in DNA by
monochromatic ultraviolet light ranging from 150 to 365 nm.
Photochem Photobiol, 54: 403-410.
Matsuoka LY, Wortsman J, Hanifan N, & Holick MF (1988) Chronic
sunscreen use decreases circulating concentration of 25-hydroxy
vitamin D. A preliminary study. Arch Dermatol, 124: 1802-1804.
Mayne LV, Mullenders LHF, & vasn Zeeland AA (1988) Cockayne
syndrome: a UV sensitive disorder with a defect in the repair of
tanscribing DNA but normal overall excision repair. In: Friedberg EC
& Hanawalt PC ed. Mechanisms and consequences of DNA damage
processing. New York, Alan R. Liss, pp 349-353.
Menzies, SW, Greenoak, GE, Reeve, VE, & Gallagher, CH (1991)
Ultraviolet radiation-induced murine tumors produced in the absence
of ultraviolet radiation-induced systemic tumor immunosuppression.
Cancer Res, 51: 2773-2779.
Miguel AG & Tyrrell RM (1983) Induction of oxygen-dependent lethal
damage by monochromatic UVB (313 nm) radiation: strand breakage,
repair and cell death. Carcinogenesis 4, 375-380.
Milham S Jr (1983) Occupational mortality in Washington State
1950-1979. Cincinnati, Ohio, National Institute for Occupational
Safety and Health (DHSS (NIOSH) Publication No. 83-116).
Minassian DC, Mehra V, & Johnson GJ (1992) Mortality and cataract:
findings from a population-based longitudinal study. Bull World
Health Organ, 70: 219-223.
Miskin R & Ben-Ishai R (1981) Induction of placmenogen activator by
UV light in normal and Xeroderma pigmentosum fibroblasts. Proc Natl
Acad Sci (USA), 78: 6236-6240.
Mitchell DL, Pfeifer GP, Taylor JS, Zdienicka MZ, & Nikaido O (1993)
Biological role of (6-4) photoproducts and cyclobutane dimers. In:
Shima A, Ichihashi M, Fujiwara Y, & Takebe H ed. Frontiers of
photobiology. Amsterdam, Oxford, New York, Elsevier Science
Publishers, pp 337-344.
Mitchell DL & Rosenstein BS (1987) The use of specific
radioimmunoassays to determine action spectra for the photolysis of
(6-4) photoproducts. Photochem Photobiol, 45: 781-786.
Mitchell RL & Andersen IC (1965) Catalase Photoinactivation. Science
150: 74.
Moan J & Dahlback A (1992) The relationship between skin cancers,
solar radiation and ozone depletion. Br J Cancer, 65: 916-921.
Modan B, Alfandary E, Shapiro D, Lusky A, Chetrit A, Shewach-Millet
M, & Moushovitz M (1993) Factors affecting the development of skin
cancer after scalp irradiation. Radiat Res, 134: 125-128.
Mohan M, Sperduto RD, Angra SK, Milton RC, Mathur RL, Underwood BA,
Jaffery N, Pandya CB, Chhabra VK, Vajpayee RB, Kalra VK, Sharma YR,
& The Indian-US Case-Control Study Group (1989) India-US
case-control study of age-related cataracts. Arch Ophthalmol, 107:
670-676.
Moan J, Dahlbach A, Henriksen T, & Magnus K (1989) Biological
amplification factor for sunlight-induced nonmelanoma skin cancer at
high latitudes. Cancer Res, 49: 5207-5212.
Molès J-P, Moyret C, Guillot B, Jeanteur P, Guilhou J-J, Theillet C,
& Basset-Sèguin N (1993) p53 Mutations in human epithelial skin
cancers. Oncogene, 8: 583-588.
Molesworth EH (1927) Rodent ulcer. Med J Aust, 1: 878-899.
Moller H, Mellemgaard A, Jacobsen GK, Pedersen D, & Storm HH (1993)
Incidence of second primary cancer following testicular cancer. Eur
J Cancer, 29A: 672-676.
Mooy CM, Van der Helm MJ, Van der Kwast ThH, De Jong PTVM, Ruiter
DJ, & Zwarthoff EC (1991) No N- ras mutations in human uveal
melanoma: the role of ultraviolet light revisited. Br J Cancer, 64:
411-413.
Moran DJ & Hollows FC (1984) Pterygium and ultraviolet radiation: a
positive correlation. Br J Ophthalmol, 68: 343-346.
Morimoto S & Kunihiko Y (1989) Psoriasis and vitamin D: A review of
our experience. Arch Dermatol, 125: 231-234.
Morin RJ, Hu B, Peng S-K, & Sevanian A (1991) Cholesterol oxides and
carcinogenesis. J Clin Lab Anal, 5: 219-225.
Morison WL (1984) The effect of a sunscreen containing
para-aminobenzoic acid on the systemic immunologic alterations
induced in mice by exposure to UVB radiation. J Invest Dermatol, 83:
405-408.
Morlière P, Mayson A, Santus R, Huppe G, Mazière JP, & Dubertet L
(1991) UVA-induced lipid peroxidation in cultured human fibroblasts.
Biochem Biophys Acta, 1084(3): 261-268.
Morlière P, Moysan A, Gaboriou F, Santus R, Mazière JC, & Dubertret
L (1992) Ultraviolet A et la peau: implications d'espèces activées
de l'oxygène, tendances actuelles et résultats récents. Pathol Biol,
40: 160-168.
Moseley H (1988) Hospital Physicists' Association - Non-ionizing
radiation, medical physics handbook 18. Bristol, Philadelphia, Adam
Hilger Publishers.
Mountin JW & Dor H(1939) Some peculiarities in the geography of
cancer. J Am Med Assoc, 113: 2405-2410.
Muir C, Waterhouse J, Mack T, Powell J, & Whelan S ed. (1987) Cancer
incidence in five continents, Vol V (IARC Scientific Publications No
88), Lyon, International Agency for Research on Cancer.
Munkata N (1993) Biological effective dose of solar ultraviolet
radiation estimated by spore dosimetry in Tokyo since 1989.
Photochem Photobiol, 58(3): 386-392.
Munch-Petersen B, Frentz G, Squire B, Wallevik K, Horn CC, Reymann
F, & Faber M (1985) Abnormal lymphocyte response to uv radiation in
multiple skin cancer. Carcinogenesis, 6: 843-845.
Murali NS & Teramura AH (1987) Insensitivity of soyabean
photosynthesis to UVB radiation under phosphorus deficiency. J Plant
Nutr, 10: 501-515.
Murphy GF, Walsh Lj, Kaidby K, & Lavker RM (1991) UV and sunbeds
(Abstract) Clin. Res. 39,195.
Murphy GM, Wright J, Nicholls DSH, McKee PH, Messenger AG, Hawk JLM,
& Levene GM (1989) Sunbed induced pseudoporphyria. Br J Dermatol,
120: 555-562.
NSF (1993) National Science Foundation, Div Polar Programs, Report
to P Penhale (NSF) & CR Booth (Biospherical Instruments), Antarctic
Support Assoc US.
Nachtwey DB & Rundel RD (1981) A photobiological evaluation of
tanning booths. Science, 211: 405-407.
Nakazawa H, English D, Randell PL, Nakazawa K, Martel N, Armstrong
BK, & Yamasaki H (in press) UV and skin cancer; specific p53 gene
mutation in normal skin as a biologically relevant exposure
measurement. Proc Natl Acad Sci (USA).
Naumann GOH & Apple D (1986) Pathology of the eye. Berlin,
Heidelberg, New York, Springer-Verlag.
Nelemans PJ, Groenendal H, Kiemeney LALM, Rampen FHJ, Ruiter DJ, &
Verbeek ALM (1993) Effect of intermittent exposure to sunlight on
melanoma risk among indoor workers and sun sensitive individuals.
Environ Health Perspect, 101(3): 252-255.
Nelson EW, Eichwald EJ, & Shelby J (1987) Increased ultraviolet
radiation-induced skin cancers in cyclosporine-treated mice. Transpl
Proc, 19: 526-527.
NIH (1991) Executive summary: Guidelines for the diagnosis and
management of asthma. Bethesda, Maryland, National Institutes of
Health, US Department of Health and Human Services (Publication No.
91-3042A).
Noell WK, Walker VS, & Kang BS (1966) Retinal damage by light in
rats. Invest Ophthalmol Vis Sci, 5: 450-473.
Noonan FP & De Fabo EC (1990) Ultraviolet-B-dose-response curves for
local and systemic immunosuppression are identical. Photochem
Photobiol, 52, 801-810.
Noonan FP, Kripke ML, Pedersen GM, & Green MI (1981a) Suppression of
contact hypersensitivity in mice by ultraviolet irradiation is
associated with defective antigen presentation. Immunology, 43:
527-533.
Noonan FP, De Fabo EC, & Kripke ML (1981b) Suppression of contact
hypersensitivity by UV radiation and its relationship to UV-induced
suppression of tumor immunity. Photochem Photobiol, 34: 683-689.
Noonan FP, De Fabo EC, & Morrison H (1988) Cis-urocanic acid, a
product formed by ultraviolet B irradiation of the skin, initiates
an antigen presentation defect in splenic dendritic cells in vivo.
J Invest Dermatol, 90: 92-99.
Norbury KC, Kripke ML, & Budmen M (1977) in vitro reactvity of
macrophages and lymphocytes from ultraviolet-irradiated mice. J Natl
Cancer Inst, 59: 1231-1235.
Norn MS (1982) Spheroid degeneration, pinguecula, and pterygium
among Arabs in the Red Sea Territory, Jordan. Acta Ophthalmol, 60:
949-954.
Norval M, Howie SE, Ross JA, & Maingay JP (1987) A murine model of
herpes simplex virus recrudescence. J Gen Virol, 68: 2693-2698.
Nultsch W, Pfan J, & Huppertz K (1990) Photoinhibition of
photosynthetic oxygen production and its recovery in the subtidal
red data Polyneura hilliae, Bot. Acta, 103: 62-67.
O'Beirn SF, Judge P, Urbach F, MacCon CF, & Martin F (1970) Skin
cancer in County Galway, Ireland. Proc Natl Cancer Conf, 6: 489-500.
Oberhelman L, Koren H, & Cooper KD (1992) Depressed capacity to
contact sensitiza women at the onset of menses; elevated contact
sensitization capacity during mid cycle can be inhibited by UV. Clin
Res, 40: 542A.
O'Dell BL, Jessen RT, Becker LE, Jackson RT, & Smith EB (1980)
Diminished immune response in sun-damaged skin. Arch Dermatol, 116:
559-561.
Oettlé,AG (1963) Skin cancer in Africa. Natl Cancer Inst Monogr, 10:
197-214.
Office of Population, Censuses and Surveys (1986) Occupational
mortality: the registrar general's decennial, supplement for Great
Britain 1979-80, 1982-83. London, Office of Population Censuses and
Surveys, Her Majesty's Stationary Office (Report Series DS No. 6).
Oldenburg JB, Gritz DC, & McDonnell PJ (1990) Topical ultraviolet
light-absorbing chromophore protects against experimental
photokeratitis. Arch Ophthalmol, 108: 1142-1144.
O'Loughlin C, Moriarty MJ, Herity B, & Daly, L (1985) A re-
appraisal of risk factors for skin carcinoma in Ireland: a case
control study. Ir J Med Sci, 154: 61-65.
Olsen JH & Jensen OM (1987) Occupation and risk of cancer in
Denmark. An analysis of 93 810 cancer cases, 1970-1979. Scand J Work
Environ Health, 13(Suppl 1): 1-91.
Oluwasanmi JO, Williams AO, & Alli AF (1969) Superficial cancer in
Nigeria. Br J Cancer, 23: 714-728.
Omdahl JR, Garry TJ, & Hunsaker A (1982) Nutritional status in the
healthy elderly population: vitamin D. Am J Clin Nutr, 6: 1225-1233.
Organisciak DT & Winkler BS (1993) Retinal light damage: Practical
and theoretical considerations. In: Osborne N & Chader G ed.
Progress in retinal research. Oxford, New York, Seoul, Tokyo,
Pergamon Press.
Orth AB, Teramura AH, & Sisler HD (1990) Effects of UV-B radiation
on fungal disease development in Cucumis sativus. Am J Bot, 77:
1188-1192.
Ostendfeld-Åkerblom A (1988) Pseudoexfoliation in Eskimos (Inuit) in
Greenland. Acta Ophthalmol, 66: 467-468.
Osterlind A (1987) Trends in incidence of ocular malignant melanoma
in Denmark 1943-1982. Int J Cancer, 40: 161-164.
Osterlind A, Olsen JH, Lynge E, & Ewertz M (1985) Second cancer
following cutaneous melanoma and cancers of the brain, thyroid,
connective tissue, bone, and eye in Denmark, 1943-80. Natl Cancer
Inst Monogr, 68: 361-388.
Osterlind A, Hou-Jensen K, & Jensen OM (1988a) Incidence of
cutaneous malignant melanoma in Denmark 1978-1982. Anatomic site
distribution, histologic types, and comparison with non-melanoma
skin cancer. Br J Cancer, 58: 385-391.
Osterlind A, Tucker MA, Stone BJ, & Jensen OM (1988b) The Danish
case-control study of cutaneous malignant melanoma. II. Importance
of UV-light exposure. Int J Cancer, 42: 319-324.
Otani T & Mori R (1987) The effects of UV irradiation of the skin on
herpes simplex virus infection: Alteration in immune function
mediated by epidermal cells and in the course of infection. Arch
Virol, 96(1-2): 1-15.
Oxholm A, Oxholm P, Staberg B, & Bendtzen K (1988)
Immunohistological detection of interleukin-like molecule and tumor
necrosis factor in human epidermis before and after UVB-irradiation
in vivo. Br J Dermatol, 118: 369-376.
Paffenbarger RS Jr, Wing AL, & Hyde RT (1978) Characteristics in
youth predictive of adult-onset malignant lymphomas, melanomas, and
leukemias: brief communication. J Natl Cancer Inst, 60: 89-92.
Paksoy N, Bouchardy C, & Parkin DM (1991) Cancer incidence in
Western Samoa. Int J Epidemiol, 20: 634-641.
Pang Q & Hays JB (1991) UV-B inducible and temperature- sensitive
photoreactivation of cyclobutane pyrimidine dimers in Arabidopsis
thaliana. Plant Physiol, 95: 536-543.
Parkin DM, Muir CS, Whelan SL, Gao Y-T, Ferlay J, & Powell J ed.
(1992) Cancer incidence in five continents, Vol VI (IARC Scientific
Publications No. 120), Lyon, International Agency for Research on
Cancer.
Parrish JA, Jaenicke KF, & Anderson RR (1982) Erythema and
melanogenesis action spectra of normal human skin. Photochem
Photobiol, 36: 187-191.
Patrick MH (1977) Studies on thymine-derived UV photoproducts in DNA
- I. Formation and biological role of pyrimidine adducts in DNA.
Photochem Photobiol, 25: 357-372.
Peak MJ & Peak JG (1982) Single-strand breaks induced in Bacillus
subtilis DNA by ultraviolet light: action spectrum and properties.
Photochem Photobiol, 35: 675-680.
Peak JG & Peak MJ (1990) Ultraviolet light induces double- strand
breaks in DNA of cultured human P3 cells as measured by neutral
filter elution. Photochem Photobiol, 52: 387-393.
Peak JG & Peak MJ (1991) Comparison of initial yields of
DNA-to-protein crosslinks and single-strand breaks induced in
cultured human cells by far-and near-ultraviolet light, blue light
and x-rays, Mutat Res, 246: 187-191.
Peak MJ, Peak JG, Moehring MP, & Webb RB (1984) Ultrviolet action
spectrum for DNA dimer induction, lethality, and mutagenesis in
Escherichia coli with emphasis on the UVB region. Photochem
Photobiol, 40: 613-620.
Peak MJ, Peak JG, Sikorski RS, & Jones CA (1985a) Induction of
DNA-protein crosslinks in human cells by ultraviolet and visible
radiations: action spectrum. Photochem Photobiol, 41: 295-302.
Peak MJ, Peak JG, & Jones CA (1985b) Different (direct and indirect)
mechanisms for induction of DNA-protein crosslinks in human cells by
far- and near-ultraviolet radiation (290 and 405 nm). Photochem
Photobiol, 42: 141-146.
Peak MJ, Peak JG, & Carnes BA (1987) Induction of direct and
indirect single-strand breaks in human cell DNA by far and
near-ultraviolet radiations: action spectrum and mechanisms.
Photochem Photobiol, 45: 381-387.
Perna JJ, Mannix ML, Ronney JF, Notkins AL, & Straus SE (1987)
Reactivation of latent herpes simplex virus infection by ultraviolet
light: A human model. J Am Acad Dermatol, 17(3): 473-478.
Philips (1983) Lighting, comprehensive handbook including technical
section. Croydon, Philips Lighting.
Pierceall WE, Mukhopadhyay T, Goldberg LH, & Ananthasswamy HN (1991)
Mutations in the P53 tumor suppressor gene in human cutaneous
squamous cell carcinomas. Mol Carcinog, 4: 445-449.
Piltingsrud HV, Fong CW, & Odland LT (1978) An evaluation of
ultraviolet radiation personnel hazards from selected 400-watt high
intensity discharge lamps. Am Ind Hyg Assoc J, 39: 406-413.
Pincus MW, Rollings PK, Craft AB, & Green A (1991) Sunscreen use on
Queensland beaches, Australia. J Dermatol, 32: 21.
Pitcher H & Longtreth J (1991) Melanoma mortality and exposure to
ultraviolet radiation: An empirical relationship. Environ Int, 17:
7-21.
Pitts DG (1974) The human ultraviolet action spectrum. Am J Optom
Arch Am Acad Optom, 51: 946-960.
Pitts DG (1978) The ocular effects of ultraviolet radiation. Am J
Optom Phys Optics, 55: 19-35.
Pitts DG, Cullen AP, & Hacker PD (1977) Ocular effects of
ultraviolet radiation from 295 to 365nm. Invest Ophthalmol Vis Sci,
16: 932-939.
Pound AW (1970) Induced cell proliferation and the initiation of
skin tumour formation in mice by ultraviolet light. Pathology, 2:
269-275.
Prystowsky JH (1988) Photoprotection and the vitamin D status of the
elderly. Arch Dermatol, 124: 1844-1848.
Punnonen K, Puntela A, & Ahotupa M (1991) Effects of ultraviolet A
and B irradiation on lipid peroxidation and activity of the
antioxidant enzymes in keratinocytes in culture. Photochem
Photoimmunol Photomed, 8: 3-6.
Putvinsky AV, Sokolov AI, Roshchupkin DI, & Vladimirov Ya (1979)
Electric breakdown of bilayer phospholipid membranes under
ultraviolet irradiation-induced lipid peroxidation. FEBS Lett, 106:
53-55.
Quan MB & Moy RL (1991) The role of human papillomavirus in
carcinoma. J Am Acad Dermatol, 25: 698-705.
Quintern LE, Horneck G, Eischweiler V, & Büker H (1992) A biofilm
used as ultraviolet dosimeter. Photochem Photobiol, 55: 389-395.
Rady P, Scinicariello F, Wagner RF, & Tyring SK (1992) p53 Mutations
in basal cell carcinomas. Cancer Res, 52: 3804-3806.
Rapp LM & Smith SC (1992) Morphorlogical comparisons between
rhodopsin-mediated and short wavelength classes of retinal light
damage. Invest Ophthalmol Vis Sci, 33: 3367-3377.
Rasanen L, Reunala T, Lehto M, Jansen C, Rantala I, & Leinikki P
(1989) Immediate decrease in antigen-presenting function and delayed
enhancement of interleukin-1 production in human epidermal cells
after in vivo UVB irradiation. Br J Dermatol, 120: 589-596.
Rathbun WB (1989) In: Dolphin D, Poulson R, & Avramoric O ed.
Coenzymes and cofactors - Volume III, Glutathione. New York, John
Wiley and Sons, Inc., pp 467-509.
Raven JA, (1991) Responses of aquatic photosynthetic organisms to
increased solar UVB. J Photochem Photobiol, B9: 239-244.
Reeve VE, Greenoak GE, Gallagher CH, Canfield PJ, & Wilkinson FJ
(1985) Effect of immunosuppressive agents and sunscreens on UV
carcinogenesis in the hairless mouse. Aust J Exp Biol Med Sci, 63:
655-665.
Reeve VE, Matheson M, Greenoak GE, Canfield PJ, Boehm- Wilcox C, &
Gallagher CH (1988) Effect of dietary lipid on UV light
carcinogenesis in the hairless mouse. Photochem Photobiol, 48:
689-696.
Reeve VE, Greenoak GE, Canfield PJ, Boehm-Wilcox C, & Gallagher CH
(1989) Topical urocanic acid enhances UV- induced tumour yield and
malignancy in hairless mouse. Photochem Photobiol, 49: 459-464.
Reeve VE, Bosnic M, & Boehm-Wilcox C (1990) Effect of ultraviolet
(UV) radiation and UVB-absorbing sunscreen ingredients on
7,12-dimethyllbenz(a)anthracene-initiated skin tumorigenesis in
hairless mice. Photodermatol Photoimmunol Photomed, 7: 222-227.
Reeve VE, Bosnic M, Boehm-Wilcos C, & Ley RD (1991) Differential
protection by two sunscreens from UV radiation-induced
immunosuppression. J Invest Dermatol, 97: 624-628.
Reeve VE, Bosnic M, & Rozinova E (1993) Carnosine
(beta-alanylhistidine) protects from the suppression of contact
hypersensitivity by ultraviolet B (280-320 nm) radiation or by cis
urocanic acid. Immunology, 78: 99-104.
Remé CH, Braschler U, Roberts J, & Dillon J (1991) Light damage in
the rat retina: effect of a radioprotective agent (WR-77913) on
acute rod outer segment disk disruption. Photochem Photobiol, 54:
137-142.
Reynolds P, Saunders LD, Layefsky ME, & Lemp GF (1993) The spectrum
of acquired immunodeficiency syndrome (AIDS)- associated
malignancies in San Francisco, 1980-1987. Am J Epidemiol, 137:
19-30.
Rhodes AR, Albert LS, Barnhill RL, & Weinstock MA (1991) Sun-induced
freckles in children and young adults: A correlation of clinical and
histopathologic features. Cancer, 67: 1990-2001.
Rigel DS, Friedman RJ, Levenstein MJ, & Greenwald DI (1983)
Relationship of fluorescent lights to malignant melanoma: another
view. J Dermatol Surg Oncol, 9: 836-838.
Ringvold A & Davangar M (1985) Changes in the rabbit corneal stroma
caused by UV irradiation. Acta Ophthalmol, 63: 601-606.
Ringvold A, Davander M, & Olsen EG (1982) Changes of the cornea
epithelium after ultraviolet radiation. Acta Ophthalmol, 60: 41-52.
Rippey J & Schmaman A (1972) Skin tumours of Africans. In: Marshall
J ed. Essays on tropical dermatology. Amsterdam, Excerpta Medica,
vol 2, pp 98-115.
Rivas JM & Ullrich SE (1992) Systemic suppression of delayed-type
hypersensitivity by supenatants from UV-irradiated keratinocytes. An
essential role for keratinocyte-derived Il-10. J Immunol, 149:
3865-3871.
Rivers JK, Norris PG, Murphy GM, Chu AC, Midgley G, Morris J, Morris
RW, Young AR, & Hawk JLM (1989) Tanning, photoprotection, acute
adverse effects and immunological changes. Br J Dermatol, 120:
767-777.
Ro YS, Cooper PN, Lee JA, Quinn AG, Harrison D, Lane D, Horne CHW
Rees JL, & Angus B (1993) p53 Protein expression in benign and
malignant skin tumours. Br J Dermatol, 128: 237-241.
Roberts DL (1990) Incidence of non-melanoma skin cancer in West
Glamorgan, South Wales. Br J Dermatol, 122: 399-403.
Roberts LK & Daynes RA (1980) Modification of the immunogenic
properties of chemically induced tumors arising in hosts treated
concomitantly with ultraviolet light. J Immunol, 125: 438-447.
Roberts JE, Kinley J, Young A, Jenkins G, Atherton S, & Dillon J
(1991) in vivo and photophysical studies on photooxidative damage
to lens proteins and their protection by radioprotectors. Photochem
Photobiol, 53: 33-38.
Robinson JK & Rademaker AW (1992) Relative importance of prior basal
cell carcinomas, continuing sun exposure, and circulating
T-Lymphocytes on the development of basal cell carcinoma. J Invest
Dermatol, 99: 227-231.
Roffo AH (1934) Cancer et soleil: carcinomes et sarcomes provoqués
par l'action du soleil in toto. Bull Assoc Fr Étude Cancer, 23:
590-616.
Roff AH (1939) Physico-chemical etiology of cancer (with special
emphasis on the association with solar radiation). Strahlentherapie,
66: 328-350 (in German).
Ros J (1990) On the effect of UV-radiation on elongation growth of
sunflower seedlings (Helianthus annuus L.)(Thesis), pp. 1-157 in
Karlst. Beitr. Entw Okophysiol 8, M Tevini (ed), Bot Inst II,
Karlsruhe.
Roschupkin DJ, Pelenitsyn AB, Potapenko AY, Talitsky VV & Vladimirov
YA (1975) Study of the effects of ultraviolet light on biomembranes
- IV. The effect of oxygen on UV-induced hemolysis and lipid
photoperoxidation in rat erythrocytes and liposomes. Photochem.
Photobiol. 21: 63-69.
Rose EF (1973) Pigment variation in relation to protection and
susceptibility to cancer. Pigment Cell, 1:, 236-45.
Rosen ES (1986) Filtration of non-ionizing radiation by the ocular
media. In: Cronley-Dillon J, Rosen ES, & Marshall J ed. Hazards of
light: myths and realities of eye and skin. Oxford, New York,
Pergamon Press, pp 145-152.
Rosenstein BS & Ducore JM (1983) Induction of DNA strand breaks in
normal human fibroblasts exposed to monochromatic ultraviolet and
visible wavelengths in the 240-546 nm range. Photochem Photobiol,
38: 51-55.
Rosenthal F, Phoon C, Bakalian A, & Taylor H (1988) The ocular dose
of ultraviolet radiation to outdoor workers. Invest Ophthalmol Vis
Sci, 29: 649-656.
Rosenthal F, Safran M, & Taylor H (1985) The ocular dose of
ultraviolet radiation from sunlight exposure. Photochem Photobiol,
42: 163-171.
Roshchupkin DI, Pelenitsyn AB, Potapenko AY, Talitsky VV, &
Vladimirov YA (1975) Study of the effects of ultraviolet light on
biomembranes. IV. The effect of oxygen on UV-induced hemolysis and
lipid photoperoxidation in rat erythrocytes and liposomes. Photochem
Photobiol, 21: 63-69.
Ross JA, Howie SE, Norval M, Maingay J, & Simpson TJ (1986)
Ultraviolet-irradiated urocanic acid suppresses delayed-type
hypersensitivity to herpes simplex virus in mice. J Invest Dermatol,
87: 630-633.
Roth M, Müller J, & Boyle JM (1987) Immunochemical determination of
an initial step in thymine dimer excision repair in xeroderma
pigmentosum variant fibroblasts and biopsy material from the normal
population and patients with basal cell carcinoma and melanoma.
Carcinogenesis, 8: 1301-1307.
Roy CR & Gies HP (1993) Personal protection against solar
ultraviolet radiation. In: Proceedings of the International
Symposium on UV, Munich 4-6 May 1993. German Ministry of the
Environment, Munich, Germany, pp 47-52.
Roy CR, Gies HP, & Elliot G (1988) Solar ultraviolet radiation:
personel exposure and protection. J Occup Health Safety (Aust N Z),
4: 133.
Roza L, Baan RA, Van Der Leun JC, Kligman L, & Young AR (1989) UVA
hazards in skin associated with the use of tanning equipment. J
Photochem Photobiol, B3: 281-287.
Russell WO, Wynne ES, Loquvam GS, & Mehl DA (1956) Studies on bovine
ocular squamous carcinoma ('cancer eye'). I. Pathological anatomy
and historical review. Cancer, 9: 1-52.
Ryel RJ, Barnes PW, Beyschlag W, Caldwell MM, & Flint SD (1990)
Plant competition for light analyzed with a multispecies canopy
model. I Model development and influence of enhanced UV-B conditions
on photosynthesis in mixed wheat and wild oat canopies. Oecologia,
82: 304-310.
Sadamori N, Mine M, & Honda T (1991) Incidence of skin cancer among
Nagasaki atomic bomb survivors. J Radiation Res, 32, Suppl 2:
217-225.
Saftlas AF, Blair A, Cantor KP, Hanrahan L, & Anderson HA (1987)
Cancer and other causes of death among Wisconsin farmers. Am J Ind
Med, 11: 119-129.
Schaeffer L, Ray R, Humbert S, Moncollin V, Vermeulen W, Hoeijmakers
JHJ, Chambon P, & Egly JM (1993) The basic transcription factor
BTF2/TFIIH contains a helicase involved in both transcription and
DNA repair. Science, Apr 2; 206(5104):37-38.
Scharffetter K, Wlaschek M, Hogg A, Bolsen K, Schothorst A, Goerz G,
Krieg T, & Plewig G (1991) Arch Dermatol Res, 283: 506-511.
Schmitt C, Schmitt J, Wegener A, & Hockwin O (1988) Effect of an
aldose reductase inhibitor, AL-1576, on the development of UV-B and
X-Ray cataract. Graefe's Arch Clin Exp Ophthlamol, 226: 455-460.
Schothorst AA, Slaper H, Schouten R, & Suurmond D (1985) UVB dose in
maintenance psoriasis phototherapy versus solar UVB exposure.
Photodermatology, 3: 213-220.
Schwartz GG & Hulka BS (1990) Is vitamin D deficiency a risk factor
for prostate cancer? Anticancer Res, 10: 1307-1311.
Schwartz SM & Weiss NS (1988) Place of birth and incidence of ocular
melanoma in the United States. Int J Cancer, 41: 174-177.
SCOPE/UNEP (1993) Effects of increased ultraviolet radiation on
global ecosystems (Workshop proceedings). Paris, Scientific
Committee on Problems of the Environment, United Nations Environment
Programme.
Scotto J, Kopf AW, & Urbach F (1974) Non-melanoma skin cancer among
Caucasians in four areas of the United States. Cancer, 34:
1333-1338.
Scotto J & Fears TR (1987) The association of solar ultraviolet and
skin melanoma incidence among Caucasians in the United States.
Cancer Invest, 5: 275-283.
Scotto J, Fears TR, & Fraumeni JF Jr (1982) Solar radiation. In:
Schottenfeld D & Fraumeni JF Jr ed. Cancer epidemiology and
prevention. Philadelphia, Pennsylvania, W.B. Saunders Company, pp
254-276.
Scotto J, Fears TR, & Fraumeni JF Jr (1983) Incidence of nonmelanoma
skin cancer in the United States. Bethesda, Maryland, National
Cancer Institute (NIH Publication No. 83-2433).
Scotto J, Cotton G, Urbach F, Berger D, & Fears T (1988)
Biologically effective ultraviolet radiation: surface measurements
in the US 1974-1985. Science, 235: 762-764.
Seddon JM, Gragoudas ES, Glynn RJ, Egan KM, Albert DM, & Blitzer PH
(1990) Host factors, UV radiation and risk of uveal melanoma. A
case-control study. Arch Ophthalmol, 108: 1274-1280.
Serrano H, Scotto J, Shornick G, Fears TR, & Greenberg ER (1991)
Incidence of nonmelanoma skin cancer in New Hampshire and Vermont. J
Am Acad Dermatol, 24: 574-579.
Servilla KS, Burnham DK, & Daynes RA (1987) Ability of cyclosporine
to promote the growth of transplanted ultraviolet radiation-induced
tumors in mice. Transplantation, 44; 291-295.
Setlow RB, Grist E, Thompson K, & Woodhead AD (1993) Wavelengths
effective in induction of malignant melanoma. Proc Natl Acad Sci
(USA), 90: 6666-6670.
Setlow RB, Woodhead AD, & Grist E (1989) Animal model for
ultraviolet radiation-induced melanoma: platyfish-swordtail hybrid.
Proc Natl Acad Sci (USA), 86: 8922-8926.
Shea CR, McNutt NS, Volkenandt M, Lugo J, Prioleau PG, & Albino AP
(1992) Overexpression of p53 protein in basal cell carcinoma of
human skin. Am J Pathol, 141: 25-20.
Shearer GM & Clerici M (1992) T-helper cell immune dysfunction in
asymptomatic HIV-1 seropositive individuals: The role of TH1-TH2
cross regulation. In: Coffman RL ed. Regulation and functional
significance of T-cell subsets. Prog Chem Immunol, 54, 21-43
Shetlar MD (1980) Cross-linking of proteins to nucleic acids by
ultraviolet light. Photochem Photobiol Rev, 5: 105-197.
Shibata T, Katoh N, Hatano T, & Sasaki K. Population based
case-control study of cortical cataract in the Noto area, Japan,
Ophthalmic Res. (1993, in press).
Shore RE (1990) Overview of radiation-induced skin cancer in humans.
Int J Radiat Biol, 57: 809-827.
Shore RE, Albert RE, Reed M, Harley N, & Pasternack BS (1984) Skin
cancer incidence among children irradiated for ringworm of the
scalp. Radiat Res, 100: 192-204.
Shukla VK, Hughes DC, Hughes LE, McCormick F, & Padua RA (1989) ras
mutations in human melanotic lesions: K- ras activation is a
frequent and early event in melanoma development. Oncogene Res, 5:
121-127.
Siemiatycki J (1991) Risk factors for cancer in the workplace. Boca
Raton, Florida, CRC Press.
Simon JC, Cruz PD, Bergstresser PR, & Tigelaar RE (1990) Low dose
ultraviolet B-irradiated Langerhans cells preferentially activate
CD4+ cells of the T helper 2 subset. J Immunol, 145: 2087-2091.
Simon JC, Tigelaar RE, Bergstresser PR, Edelbaum D, & Cruz PD (1991)
Ultraviolet B radiation converts Langerhans cells from immunogenic
to tolerogenic antigen-presenting cells. J Immunol, 146: 485-491.
Sjovall P, Christensen OB, & Moller H (1985) Single exposure to
ultraviolet irradiation and elicitation of human allergic contact
dermatitis. Acta Derm Venereol, 65: 93-96.
Slaper H (1987) Skin cancer and UV exposure: Investigations on the
estimation of risks. University of Utrecht, The Netherlands (PhD
Thesis).
Slaper H, Schothorst AA, & Van der Leun JC (1986) Risk evaluation of
UVB-therapy for psoriasis: comparison of calculated risk for
UVB-therapy and observed risk in PUVA- treated patients.
Photodermatology, 3: 271-283.
Sliney, DH (1983) Eye protective techniques for bright light.
Ophthalmology 90(8), 937-944.
Sliney D (1986) Physical factors in cataractogenesis: ambient
ultraviolet radiation and temperature. Invest Ophthalmol Vis Sci,
27(5):, 781-790.
Sliney DH (1987) Estimating the solar ultraviolet radiation exposure
to an intraocular lens implant, J Cataract Refract Surg,
13(5),296-301.
Sliney DH & Wolbarsht ML (1980) Safety with lasers and other optical
sources -A comprehensive handbook. New York, London, Plenum Press.
Smith KC (1976) The radiation-induced addition of proteins and other
molecules to nucleic acids. In: Wang SY ed. Photochemistry and
photobiology of nucleic acids. New York, London, San Francisco,
Academic Press, vol 2, pp 187-218.
Smith GJ & Ryan KG (1993) The effect of changes on differences in
Robertson-Berger radiometer responsitivity on solar ultraviolet-B
measurement. Photochem Photobiol, 58(4): 512-514.
Smith RC, Baker KS, Holm-Hansen O, & Olson R (1980) Photoinhibition
of photosynthesis in natural waters. Photochem Photobiol, 31:
585-592.
Smith EL, Walworth NC, & Holick MF (1986) Effects of 2
alpha-25-dihydrxy vitamin D3 on the morphologic and biological
differentiation of cultured human epidermal keratinocytes grown in
serum-free conditions. J Invest Dermatol, 86: 709-714.
Söderberg PG (1989) Mass alteration in the lens after exposure to
ultraviolet radiation, 300nm. Acta Ophthalmol, 67: 633-644.
Söderberg PG (1991) Na and K in the lens after exposure to radiation
in the 300 nm wavelength region. J Photochem Photobiol, 8: 279-294.
Soparker CN, O'Brien JM, & Albert DM (1993) Investigation of the
role of the ras protooncogene point mutation in human uveal
melanomas. Invest Ophthalmol Vis Sci, 34: 2203-2209.
Sorahan T & Grimley RP (1985) The aetiological significance of
sunlight and fluorescent lighting in malignant melanoma: a
case-control study. Br J Cancer, 52; 765-769.
Spector A (1991) In: Sies H ed. The lens and oxidative stress.
Endogenous antioxidant defences in human blood plasma in oxidative
stress: Oxidants and antioxidants. New York, London, San Francisco,
Academic Press.
Spellman CW & Daynes RA (1978) Properties of ultraviolet
light-induced suppressor lymphocytes within a syngeneic tunour
system. Cell Immunol, 36: 383-387.
Spellman CW, Woodward JG, & Daynes RA (1977) Modification of
immunological potential by ultrviolet radiation. I Immune status of
short-term UV-irradiated mice. Transplantation, 24: 112-119.
Spitzer WO, Hill GB, Chambers LW, Helliwell BE, & Murphy HB (1975)
The occupation of fishing as a risk factor in cancer of the lip. N
Engl J Med, 293: 419-424.
Spruance SL (1985) Pathogenesis of herpes simplex labialis:
Experimental induction of lesions with UV light. J Clin Microbiol,
22: 366-368.
Staberg B, Wulf HC, Poulsen T, Klemp P, & Brodthagen H (1983)
Carcinogenic effect of sequential artificial sunlight and UVA
irradiation in hairless mice. Consequences for solarium 'therapy'.
Arch Dermatol, 119: 641-643.
Stadtman ER (1990) Metal ion catalysed oxidation of
protein-biochemical mechanism and biological consequences. Free
Radic Biol Med, 9: 315-325
Stamnes K, Tsay SC, Wiscombe WJ, & Jayaweera K (1988) Numerically
stable algorithm for discreet-ordinate method radiative transfer in
multiple scattering and emitting media. Appl Optics, 27: 2502-2509.
Stein B, Rahmsdorf HJ, Steffen A, Liffin M, & Herrlich P (1989)
UV-induced damage is an intermediate step in UV-induced expression
of human immunodeficiency virus type 1, collagenase, c-fos and
metallothionein. Mol Cell Biol, 9: 5169-5181.
Steinitz R, Parkin DM, Young JL, Bieber CA, & Katz L ed. (1989)
Cancer incidence in Jewish migrants to Israel, 1961-1981 (IARC
Scientific Publications No 98), Lyon, International Agency for
Research on Cancer, 114-115, 134-135.
Stenbäck F (1975a) Species-specific neoplastic progression by
ultraviolet light on the skin of rats, guinea pigs, hamsters and
mice. Oncology, 31: 209-225.
Stenbäck F (1975b) Ultraviolet light irradiation as initiating agent
in skin tumor formation by the two-stage method. Eur J Cancer, 11:
241-246.
Stephenson TJ, Royds J, Silcocks PB, & Bleehen SS (1992) Mutant p53
oncogene expression in keratoacanthoma and squamous cell carcinoma.
Br J Dermatol, 127: 566-570.
Sterenborg HJCM & van der Leun JC (1990) Tumorigenesis by a long
wavelength UVA source. Photochem Photobiol, 51: 325-330.
Sterenborg HJCM, van der Putte SCJ, & van der Leun JC (1988) The
dose-response relationship of tumorigenesis by ultraviolet radiation
of 254 nm. Photochem Photobiol, 47: 245-253.
Stern RS & Docken W (1986) An exacerbation of SLE after visiting a
tanning salon. J Am Med Assoc, 255: 3120.
Stern RS & Lange R (1988) Non-melanoma skin cancer occurring in
patients treated with PUVA five to ten years after the first
treatment. J Invest Dermatol, 91: 120-124.
Stern RS, Thibodeau LA, & Kleinerman RA (1979) Risk of cutaneous
carcinoma in patients treated with oral methoxsalen
photochemotherapy. N Engl J Med, 300: 809-813.
Stocker R & Frei B (1991) In Sies H ed. Endogenous antioxidant
defences in human blood plasma in oxidative stress: Oxidants and
antioxidants.New York, London, San Francisco, Adacemic Press.
Stretch JR, Gatter KC, Ralfkiaer E, Lane DP, & Harris, AL (1991)
Expression of mutant p53 in melanoma. Cancer Res, 51: 5976-5979.
Strickland PT, Burns FJ, & Albert, RE (1979) Induction of skin
tumors in the rat by single exposure to ultraviolet radiation.
Photochem Photobiol, 30: 683-688.
Strickland PT, Creasia D, & Kripke ML (1985) Enhancement of
two-stage skin carcinogenesis by exposure of distant skin to UV
radiation. J Natl Cancer Inst, 74: 1129-1134.
Strickland PT, Vitasa BC, West SK, Rosenthal FS, Emmett EA, & Taylor
HR (1989) Quantitative carcinogenesis in man: solar ultraviolet B
dose dependence of skin cancer in Maryland watermen. J Natl Cancer
Inst, 81: 1910-1913.
Stryker WS, Stampfer MJ, Stein EA, Kaplan L, Louis TA, Sober A, &
Willett WC (1990) Diet, plasma levels of beta-carotene and
alpha-tocopherol, and risk of malignant melanoma. Am J Epidemiol,
131: 597-611.
Sullivan JH & Teramura AH (1991) The effects of UV-B radiation on
loblolly pine. 2 Growth of field-grown seedlings, Trees (Berl)
6(3):115-120.
Sutherland JC & Griffin K (1981) Absorption spectrum of DNA for
wavelengths greater than 300nm. Radiat Res, 86: 399-410.
Swerdlow AJ, English JSC, MacKie RM, O'Doherty CJ, Hunter JAA, Clark
J, & Hole DJ (1988) Fluorescent lights, ultraviolet lamps, and risk
of cutaneous melanoma. Br Med J, 297: 647-650.
Sydenham MM, Wong CF, Hirst LW, & Collins MJ (1991) Cular UVB
dosimetry made possible for the first time using a CR-39 contact
lens. In: Proceedings of 22nd Session of CIE, Melbourne. Vienna,
International Commission of illumination, vol 1, part 2, pp 27-28.
Talbot G (1948) Pterygium. Trans Ophthalmol Soc N Z, 2: 42-45.
Taylor HR (1979) Pseudoexfoliation, an environmental disease? Trans
Ophthalmol Soc UK, 99: 302-307.
Taylor HR (1980a) Aetiology of climatic droplet keratopathy and
pterygium. Br J Ophthalmol, 64: 154-163.
Taylor HR (1980b) The environment and the lens. Br J Ophthalmol, 64:
303-310.
Taylor HR, West SK, Rosenthal FS, Munoz B, Newland HS, Abbey H, &
Emmett EA (1988) Effect of ultraviolet radiation on cataract
formation. N Engl J Med, 319: 1429-1433.
Taylor HR, West SK, Rosenthal FS, Munoz B, Newland HS, & Emmett EA
(1989) Corneal changes associated with chronic UV irradiation. Arch
Ophthalmol, 107: 1481-1484.
Taylor HR, West S, Munoz B, Rosenthal FS, Bressler SB, & Bressler NM
(1992) The long-term effects of visible light on the eye. Arch
Ophthalmol, 110: 99-104.
Taylor JR, Schmieder GF, Shimizu T, & Streilein JW (1993)
UVB-susceptibility is a risk factor for recurrent herpes labialis.
Photochem Photobiol, 57: 135.
Templeton AC (1967) Tumours of the eye and adnexa in Africans of
Uganda. Cancer, 20: 1689-1698.
Teppo L, Pukkala E, Hakama M, Hakulinen T, Herva A, & Saxén E (1980)
Way of life and cancer incidence in Finland. A municipality-based
ecological analysis. Scand J Soc Med, Suppl 19: 5-84.
Terman M, Reme CE, Rafferty B, Gallin PF, & Terman JS (1990) Bright
light therapy for winter depression: Potential ocular effects and
theoretical implications. Yearly Review. Photochem Photobiol, 51:
781-792.
Tevini M, Braun J, & Fieser G (1991a) The protective function of the
epidermal layer of rye seedlings against ultraviolet-B radiation,
Photochem Photobiol, 53: 329-333.
Tevini M, Mark U, & Saile-Mark M (1991b) Effects of enhanced solar
UV-B radiation on growth and function of crop plant seedlings. In:
Randall D & Blevins D ed. Current topics in plant biochemistry and
physiology. Columbia, University of Missouri.
Tevini M, Mark U, Fieser G, & Saile M (1991c) Effects of enhanced
solar UV-B radiation on growth and function of selected crop plan
seedlings. In: Riklis, D ed. Photobiology. New York, London, Plenum
Press, pp 635-649.
Tobal K, Warren W, Cooper CS, McCartney A, Hungerford J, & Lightman
S (1992) Increased expression and mutation of p53 in choroidal
melanoma. Br J Cancer, 66: 900-904.
Toews GB, Bergstresser PR, & Streilein JW (1980) Epidermal
langerhans cell density determines whether contact hypersensitivity
or unresponsiveness follows skin painting with DNFB. J Immunol, 124:
445-453.
Tomatis, L (1990) ed, Cancer: Causes, Occurrence and Control. Lyon,
International Agency for Research on Cancer (IARC Scientific
Publications No 100).
Travis, LB, Curtis, RE, Boice, JD, Hankey, BF, & Fraumeni Jr, JF
(1991) Second cancers following non-Hodgkin's lymphoma. Cancer, 67:
2002-2009.
Travis, LB, Curtis, RE, Hankey, BF, & Fraumeni Jr, JF (1992) Second
cancers in patients with chronic lymphocytic leukemia. J Natl Cancer
Inst, 84: 1422-1427.
Tucker, MA, Boice, JD, Jr & Hoffman, DA (1985a) Second cancer
following cutaneous melanoma and cancers of the brain, thyroid,
connective tissue, bone and eye in Connecticut, 1935-82. Natl Cancer
Inst Monogr, 68: 161-189.
Tucker, MA, Misfeldt, D, Coleman, N, Clark, WH, & Rosenberg, SA
(1985b) Cutaneous malignant melanoma after Hodgkin's Disease. Ann
Int Med, 102: 37-41.
Tucker, MA, Shields, JA, Hartge, P, Augsberger, J, Hoover, RN, &
Fraumeni, JF, Jr (1985c) Sunlight exposure as risk factor for
intraocular malignant melanoma. New Engl J Med, 313: 789-792.
Turner, BJ, Siatkowski, RM, Augsburger, JJ, Shields, JA, Lustbader,
E, & Mastrangelo, MJ (1989) Other cancers in uveal melanoma patients
and their families. Am J Ophthalmol, 107: 601-608.
Tuveson RW & Sammartano, LJ (1986) Sensitivity of HemA mutant
Escherichia coli cells to inactivation by near-UV light depends on
the level of supplementation with L-aminolevulinic acid. Photochem
Photobiol 43: 621-626.
Tyrrell RM (1973) Induction of pyrimidine dimers in bacterial DNA by
365 nm radiation, Photochem Photobiol 17: 69-73.
Tyrrell RM, Ley RD & Webb RB (1974) Induction of single strand
breaks (Alkali-labile bonds) in bacterial and phage DNA by near-UV
(365 nm) radiation. Photochem. Photobiol. 20: 395-398.
Tyrrell RM (1982) Cell inactivation and mutagenesis by solar
ultraviolet radiation In: Hélène C, Charlier M, Montenay-Garestier
Th & Laustriat G ed. Trends in Photobiology, Plenum Press, New York,
pp 155-172.
Tyrrell, RM (1984) Exposure of nondividing populations of primary
human fibroblasts to UV (254 nm) radiation induces a transient
enhancement in capacity to repair potentially lethal cellular
damage. Proc Natl Acad Sci USA 81: 781-784.
Tyrrell, RM (1991) UVA (320-380 nm) radiation as an oxidative stress
In: Sies H ed. Oxidative Stress : oxidants and antioxidants, London,
Academic Press, pp 57-83.
Tyrrell, RM (1992) Inducible responses to UVA exposure. In: (ed. F.
Urbach) Biological responses to ultraviolet A radiation, Valdenmar
Press, Overland Park, pp. 59-64.
Tyrrell, RM and Pidoux, M (1986) Endogenous glutathione protects
human skin fibroblasts against the cytotoxic action of UVB, UVA and
near-visible radiations. Photochem Photobiol 44: 561-564.
Tyrrell RM & Pidoux M (1987) Action spectra for human skin cells.
Estimates of the relative cytotoxicity of the middle ultraviolet,
near ultraviolet and violet regions of sunlight on epidermal
keratinocytes. Cancer Res 47: 1825-1829.
Tyrrell, RM and Pidoux, M (1988) Correlation between endogenous
glutathione content and sensitivity of cultured human skin cells to
radiation at defined wavelengths in the solar UV range. Photochem
Photobiol 47: 405-412.
Tyrrell, RM & Pidoux M (1989) Singlet oxygen involvement in the
inactivation of cultured human fibroblasts by UVA (334 nm, 365 nm)
and near-visible radiations. Photochem Photobiol 49: 407-412.
Ullrich SE (1985) Suppression of lymphoproliferation by
hapten-specific suppressor T lymphocytes from mice exposed to
ultraviolet radiation. Immunology, 54: 343-352.
Ullrich SE (1986) Suppression of the immune response to allogeneic
histocompatibility antigens by a single exposure to ultraviolet
radiation. Transplantation, 42: 287-291.
Ullrich SE & Kripke ML (1984) Mechanisms in the suppression of tumor
rejection produced in mice by repeated UV irradiation. J Immunol,
133: 2786-2790.
Ullrich SE, Azizi E, Kripke ML (1986a) Suppression of the induction
of delayed-type hypersensitivity reactions in mice by a single
exposure to ultraviolet radiation. Photochem Photobiol, 43: 633-638.
Ullrich SE, Yee GK, & Kripke ML (1986b) Suppressor lymphocytes
induced by epicutaneous sensitization of UV- irradiated mice control
multiple immunological pathways. Immunology, 58: 185-190.
UNEP (1987) The ozone layer, United Nations Environment Programme,
UNEP/GEMS Environmental Library No 2, UNEP, Nairobi.
UNEP (1989) Environmental Effects Panel Report, Van der Leun, JC,
Tevini, M eds. United Nations Environment Programme , Nairobi,
Kenya.
UNEP (1991) Environmental effects of ozone depletion: 1991 Update.
United Nations Environment Programme, Nairobi, Kenya.
UNEP (1992) Effects of increased ultraviolet radiation on biological
systems. Proceedings of meeting, Scientific Committee on Problems of
the Environment (SCOPE). Budapest, United Nations Environment
Programme, Nairobi, Kenya.
UNEP (1993) United Nations Environment Programme. Report of the
ninth meeting of the Open-Ended Working Group of the parties to the
Montreal Protocol, Geneva 30 August-1 September 1993.
UNEP-WMO (1989) Scientific assessment of stratospheric ozone: 1989
WMO Global ozone research and monitoring project, United Nations
Environment Programme, World Meteorological Organization, Geneva
(Report No 29).
Urbach F (1987) Man and ultraviolet radiation. In: Passchier WF &
Bosnjakovic BFM eds. Human exposure to ultraviolet radiation - risks
and regulations. New York, Excerpta Medica.
Urbach F (1987a) Cutaneous photocarcinogenesis. J Environ Sci
Health-Environ Carcinogen Rev, C5, 211-34.
Urbach F (1989) Testing the efficacy of sunscreens: effect of choice
of source and spectral distribution of ultraviolet radiation, and
choice of endpoint. Photodermatology, 6: 177-181.
Urbach F, Davies RE, & Forbes PD (1966) Ultraviolet radiation and
skin cancer in man In: Montagna W & Dobson RL eds. Advances in
Biology of Skin, Oxford, Pergamon Press, Carcinogenesis, Vol VII, pp
195-214.
Urbach F, Epstein JH, & Forbes PD (1974) Ultraviolet carcinogenesis:
experimental, global and genetic aspects. In: Pathak MA, Harber LC,
Seiji M, & Kukita A, eds. Sunlight and Man - Normal and Abnormal
Photobiological Responses, Tokyo, University of Tokyo Press, pp
259-283.
US Food and Drug Administration (1978) Sunscreen drug products for
over-the-counter use. Fed Regis, 43: 38206-28269.
US National Cancer Institute (1989) Sunscreens, Rockville, MD,
Tracor Technological Resources Inc (Class Study Report; Contract No
NO1-CP-71082 (7/89)).
Vågerö D, Ringbäck G, & Kiviranta H (1986) Melanoma and other
tumours of the skin among office, other indoor and outdoor workers
in Sweden 1961-1979. Br J Cancer, 53: 507-512.
Vågerö D, Swerdlow AJ, & Beral V (1990) Occupation and malignant
melanoma: a study based on cancer registration data in England and
Wales and in Sweden. Br J Ind Med, 47: 317-324.
Valerie K, Delers A, Bruck C, Thiriatt C, Rosenberg H, Debouck C &
Rosenberg M (1988) Activation of human immmunodefficiency virus type
1 by DNA damage in human cells. Nature 333: 78-81.
Van der Leun JC (1984) Yearly review: UV-carcinogenesis. Photochem
Photobiol, 39: 861-868.
Van der Leun JC (1987) Principles of risk reduction and protection.
In: Passchier WF & Bosnjakovic BFM, eds. Human Exposure to
Ultraviolet Radiation: Risks and Regulations, Amsterdam, Elsevier,
pp 293-303.
Van der Leun JC (1992) Interactions of UVA and UVB in
photodermatology: what was photoaugmentation? In: Urbach F ed. The
Biological Responses to Ultraviolet A Radiation, Overland Park, KS
Valdenmar, pp 309-319.
Van der Leun JC & de Gruijl FR (1993) Influences of ozone depletion
on human and animal health. In: Tevini M ed. UV-B radiation and
ozone depletion. Effects on humans, animals, plants, microorganisms,
and materials. Boc Raton, Lewis Publishers, pp 95-123.
Van der Schroeff JG, Evers LM, Boot AJM & Bos JL (1990) ras oncogene
mutations in basal cell carcinomas and squamous cell carcinomas of
human skin. J Invest Dermatol, 94: 423-425.
Van Weelden H, de Gruijl FR & van der Leun JC (1986) Carcinogenesis
by UVA, with an attempt to assess the carcinogenic risks of tanning
with UVA and UVB. In: Urbach F & Gange RW eds. The Biological
Effects of UVA Radiation, New York, Praeger, pp 137-146.
Van Weelden H and van der Leun J, (1987) IVA induced tumours in
pigmented and albino hairless mice in Human exposure to ultraviolet
radiation - risks and regulations eds. Passchier WF and Bosnjakovic
BFM (New York: Excerpta Medica).
Van Weelden H, de Gruijl FR, van der Putte SCJ, Toonstra J & van der
Leun JC (1988) The carcinogenic risks of modern tanning equipment:
is UVA safer than UVB? Arch Dermatol Res, 280: 300-307.
Van't Veer LJ, Burgering BMT, Versteeg R, Boot AJM, Ruiter DJ,
Osanto S, Schrier PI & Bos JL (1989) N- ras Mutations in human
cutaneous melanoma from sun-exposed body sites. Mol Cell Biol, 9:
3114-3116.
Varghese AJ & Wang SY (1967) Ultraviolet irradiation of DNA in
vitro and in vivo produces a third thymine-derived product.
Science 156: 955-957.
Verhoeff FH, Bell L & Walker CB (1916) The pathological effects of
radiant energy on the eye. Proc Am Acad Arts Sci, 510: 1-810.
Vermeer M & Streilein JW (1990) Ultraviolet B light-induced
alterations in epidermal Langerhans cells are mediated in part by
tumor necrosis factor-alpha. Photodermatol Photoimmunol Photomed, 7:
258-265.
Vermeer M, Schmieder GJ, Yoshikawa T, Van den Berg J-W, Metzman MS,
Taylor JR, & Streilein JW (1991) Effects of ultraviolet B light on
cutaneous immune responses of humans with deeply pigmented skin. J
Invest Dermatol, 97: 729-734.
Vile GF, Basu-Moda S, Waltner C & Tyrrell RM (1994) Haem oxygenase 1
mediates an adaptive response to oxidative stress in human skin
fibroblasts. Proc Natl Acad Sci USA 91: 2607-2610.
Vincek V, Jurimoto I, Medema JP, Prieto E, & Streilein JW (1993)
Tumor necrosis factor alpha polymorphism correlates with deleterious
effects of ultraviolet B light on cutaneous immunity. Cancer Res,
53: 728-732.
Vitale S, West S, Munoz B, Schein OD, Maguire M, Bressler, N &
Taylor HR (1992) Watermen Study II: mortality and baseline
prevalence of nuclear opacity. Invest Ophthalmol Vi Sci, 33: 1097.
Vitasa BC, Taylor HR, Strickland PT, Rosenthal FS, West S, Abbey H,
Ng SK, Munoz B, & Emmett EA (1990) Association of nonmelanoma skin
cancer and actinic keratosis with cumulative solar ultraviolet
exposure in Maryland watermen. Cancer, 65: 2811-2817.
Volkenandt M, Schlegel U, Nanus DM, & Albino AP (1991) Mutational
analysis of the human p53 gene in malignant melanoma. Pigment Cell
Res, 4: 35-40.
Vosjan, JH, Dohler, G, & Nieuwland G (1990) Effect of UVB irradiance
on the ATP content of microorganisms of the Weddell Sea Antarctica,
Neth J Sea Res 25, 391-394.
Vuillaume M, Daya-Grosjean L, Vincens P, Pennetier JL, Tarroux P,
Baret A, Calvayrac R, Taieb A & Satorin, A (1992) Striking
differences in cellular catalase activity between two DNA
repair-deficient diseases. Xeroderma pigmentosum and
trichothiodystophy. Carcinogenesis 13: 321-328.
Walker GC (1987) The SOS response of Escherichia coli In: Niedhardt
FC, Ingraham JL, Low KB, Magasanik B, Shaechter M, Umbarger HE eds.
Escherichia coli and Salmonella typhimurium. Cellular and Molecular
Biology, American Society of Microbiology, Washington DC pp
1346-1357.
Walter SD, Marrett LD, From L, Hertzman C, Shannon HS, & Roy P
(1990) The association of cutaneous malignant melanoma with the use
of sunbeds and sunlamps. Am J Epidemiol, 131: 232-243.
Warfel AH, Moy JA, Meola T, Sanchez M, Soter NA, & Belsito DV (1993)
Effect of ultraviolet B (UVB) on the expression of human
immunodeficiency virus (HIV) in mice and humans, Photochem
Photobiol, 57: 755.
Waring GO, Roth AM, & Ekins MB (1984) Clinical and pathological
description of 17 cases of corneal intraepithelial neoplasia. Am J
Ophthalmol, 97: 547-559.
Waterhouse J, Muir C, Correa P, & Powell J eds (1976) Cancer
Incidence in Five Continents, Lyon, International Agency for
Research on Cancer, Vol III (IARC Scientific Publications No 15).
Waterhouse J, Muir C, Shanmugaratnam K, & Powell J eds (1982) Cancer
Incidence in Five Continents, Lyon, International Agency for
Research on Cancer, Vol IV (IARC Scientific Publications No 42).
Webb RB (1977) Lethal and mutagenic effects of near-ultraviolet
radiation Photochem Photobiol Rev 2: 169-261.
Wei Q, Matanoski GM, Farmer ER, Hedayati MA, & Grossman L (1993) DNA
repair and aging in basal cell carcinoma: A molecular epidemiology
study. Proc Natl Acad Sci USA, 90: 1614-1618.
Weinstock MA, Colditz GA, Willett WC, Stampfer MJ, Bronstein BR,
Mihm MC Jr, & Speizer FE (1989) Nonfamilial cutaneous melanoma
incidence in women associated with sun exposure before 20 years of
age. Pediatrics, 84: 199-204.
Wellmann E, (1971) Phytochrome mediated flavone glycoside synthesis
in cell suspension cultures of Petroselinum hortense after
preirradiation with ultraviolet light, Planta, 101: 283-286.
Wellmann E, (1991) Specific ultraviolet effects in plant
development, J Exp Bot, (Suppl), 32: 42.
Welsh D & Diffey B (1981) Protection against solar actinic radiation
afforded by common clothing fabrics. Clinical and Experimental
Dermatology 6: 577-582.
Werner JS (1991) The damaging effects of light on the eye and
implications for understanding changes in vision across the life
span, in (P Bagnoli and W Hodos) The Changing Visual System, New
York, Plenum Press.
West SK, Rosenthal FS, Bressler NM, Bressler SB, Munoz B, Fine SL, &
Taylor HR (1989) Exposure to sunlight and other risk factors for
age-related macular degeneration. Arch Ophthalmol, 107: 875-879.
Wester A (1987) Ultraviolet transmission properties of sunscreens
and sunglasses. In: Passchier W & Bosnjakovic B eds. Human exposure
to ultraviolet radiation: risks and regulations, Elsevier,
Amsterdam.
Westerveld A, Hoeijmakers JHJ, van Duin M, de Wit J, Odijk H,
Pavlink A, Wood RD & Bootsma D (1984) Molecular cloning of a human
DNA repair gene. Nature 310: 425-529.
Whillock, MJ, Clark, IE, McKinlay, AF, Todd, CD & Mundy, SJ (1988)
Ultraviolet radiation levels associated with the use of fluorescent
general lighting, UVA and UVB lamps in th eworkplace and home.
National radiological Protection Board, Research Report R221,
London, HMSO.
Whillock MJ, McKinlay AF, Kemmlert J & Forsgren PG (1990)
Ultraviolet radiation emissions from miniature [compact] fluorescent
lamps. Lighting Res Technol 22: 125-128.
Whitaker CJ, Lee WR, & Downes JE (1979) Squamous cell skin cancer in
the north-west of England, 1967-69, and its relation to occupation.
Br J Ind Med, 36: 43-51.
WHO (1948) International Statistical Classification of Diseases,
Injuries and Causes of Death, Sixth Revision, Geneva.
WHO (1977) Manual of the International Statistical Classification of
Diseases, Injuries, and Causes of Death. International
Classification of Diseases, 1975 rev, Vol 1, Geneva, p 102.
WHO (1989) Non-ionizing radiation protection. MJ Suess & DA
Benwell-Morison eds. World Health Organization Regional Office for
Europe, European series No 25, WHO, Copenhagen.
WHO/UNEP/IRPA (1979) Ultraviolet Radiation, Environmental Health
Criteria 14, World Health Organization, United Nations Environment
Programme, WHO, Geneva.
Widmark (1889) Ueber den Einfluss des Lichtes auf die vorderen
medien des auges. Skand Arch f physiol 1, 264-330.
Widmark J (1901) Ueber den Einfluss des Lichtes auf die Linse.
Mitteil aus d Augenklin d Carol med chir Inst zu Stockholm 3:
135-149.
Wilesmith JW, Wells GA, Cranwell MP and Ryan JB (1988) Bovine
spongiform encephalopathy : epidemiological studies. Vet Rec, 123,
638-644.
Willis I, Menter JM, & Whyte HJ (1981) The rapid induction of
cancers in the hairless mouse utilizing the principle of
photoaugmentation. J Invest Dermatol, 76: 404-408.
Wittenberg S (1986) Solar radiation and the eye: a review of
knowledge relevant to eye care. Am J Optom Physiol Optics, 63:
676-689.
WMO (1993) World Meteorological Organization Report of the second
meeting of the ozone research managers of the parties to the Vienna
Convention for the protection of the ozone layer. Geneva 10-12
March.
Wolf P, Donawho CK, & Kripke ML (1993a) Analysis of the protective
effect of different sunscreens on ultraviolet radiation-induced
local and systemic suppression of contact hypersensitivity and
inflammatory responses in mice. J Invest Dermatol 100: 254-259.
Wolf P, Yarosh DB, & Kripke ML (1993b) Effects of sunscreens and a
DNA excision repair enzyme on ultraviolet radiation - induced
inflammation, immune suppression and cyclobutane pyrimidine dimer
formation in mice. J invest Dermatol 101: 523-527.
Wong CF, Fleming R, & Carter SJ (1989) A new dosimeter for
ultraviolet B radiation. Photochem Photobiol 50: 611-615.
Wong L, Ho SC, Coggon D, Cruddas AM, Hwang CH, Ho CP, Robertshaw AM,
& MacDonald DM (1993) Sunlight exposure, antioxidant status, and
cataract in Hong Kong fishermen. J Epidemiol Comm Health, 47: 46-49.
Wood RD, Robbins P & Lindahl T (1988) Complementation of the
Xeroderma pigmentosum DNA repair defect in cell-free extracts. Cell
53: 91-106.
Wucherpfenning V (1931) Biologie und prackische verwenbarkeit der
erythemschwelle des UV. Strahlentherapie 40: 201-244.
Yannuzzi L, Fisher Y, Slakter J, Krueger A (1989) Solar retinipathy.
A photobiologic and geophysical analysis, Retina, 9, 28-43.
Yarosh D, Alas LG, Yee V, Oberyszyn A, Kibitel JT, Mitchell D,
Rosenstein R, Spinowitz A, & Citron M (1992) Pyrimidine dimer
removal enhanced by DNA repair liposomes reduces incidence of UV
skin cancer in mice. Cancer Res, 52: 4227-4231.
Yasumoto S, Yoshinobu H, & Aurelian L (1987) Immunity to herpes
simplex virus type 2: Suppression of virus-induced responses in
ultraviolet B-irradiated mice. J Immunol, 139: 2788-2793.
Yoshida A, Ishiguro S, & Tamai M (1993) Expression of glial
fibrillary acidic protein in muoller cells after
lensectomy-vitrectomy. Invest. Ophthalmol. Vis. Sci. 34: 3154-3160.
Yoshikawa T & Streilein JW (1990) On the genetic basis of the
effects of ultraviolet B on cutaneous immunity. Evidence that
polymorphisms at the Tnfý and Lps loci govern susceptibility.
Immunogenetics, 32: 398-405.
Yoshikawa T, Rae V, Bruins-Slot W, Van den Berg J-W, Taylor Jr, &
Streilein JW (1990) Susceptibility to effects of UVB radiation on
induction of contact hypersensitivity as a risk factor for skin
cancer in humans. J Invest Dermatol, 95: 530-536.
Young AR, Magnus IA, Davies AC, & Smith NP (1983) A comparison of
the phototumorigenic potential of 8-MOP and 5-MOP in hairless albino
mice exposed to solar simulated radiation. Br J Dermatol, 108:
507-518.
Young AR, Walker SL, Kinley JS, Plastow SR, Averbeck D, Morlière P,
& Dubertret L (1990) Phototumorigenesis studies of 5-methoxypsoralen
in bergamot oil: evaluation and modification of risk of human use in
an albino mouse skin model. J Photochem Photobiol B Biol, 7:
231-250.
Young AR, Potten CS, Chadwick CA, Murphy GM, Hawk JLM & Cohen AJ
(1991) Photo-protection and 5-MOP photochemoprotection from
UVR-induced DNA damage in humans: the role of skin type. J Invest
Dermatol, 97: 942-948.
Young RW (1991) Age-related cataract. New York, Oxford University
Press.
Yu CC, MacGregor JM, Dublin EA, Barnes DM, MacDonald DM, & Levison
DA (1992) Patterns of immunostaining for p53 in benign and malignant
melanocytic lesions (meeting abstract). J Pathol, 167: (Suppl),
130A.
Zadjela E & Bisagni E (1981) 5-Methoxypsoralen, the melanogenic
additive in sun-tan preparations, is tumorigenic in mice exposed to
365 nm uv radiation. Carcinogenesis, 2: 121-127.
Zamansky G & Chou, I-N (1987) Environmental wavelength of
ultraviolet light induce cytoskeletal damage. J Inv Dermatol 89:
603-606.
Zanetti R, Rosso S, Faggiano F, Roffino R, Colonna S, & Martina G
(1988) A case-control study on cutaneous malignant melanoma in the
province of Torino, Italy (Fr). Rev Epidemiol Santé publ, 36:
309-317.
Zanetti R, Franceschi S, Rosso S, Colonna S, & Bidoli E (1992)
Cutaneous melanoma and sunburns in childhood in a southern European
population. Eur J Cancer, 28A: 1172-1176.
Zaridze D, Mukeria A, & Duffy SW (1992) Risk factors for skin
melanoma in Moscow. Int J Cancer, 52: 159-161.
Zaunuddin D & Sasaki K (1991) Risk factor analysis in a cataract
epidemiology survey in West Samatara, Indonesia. Dev Ophthalmol, 21:
78-86.
Ziegler A-M, Leffell DJ, Kunala S, Sharma HW, Gailani M, Simon JA,
Halperin AJ, Baden HP, Shapiro PE, Bale AE, & Brash DE (1993)
Mutation hotspots due to sunlight in the p53 gene of nonmelanoma
skin cancers. Proc Natl Acad Sci USA, 90: 4216-4220.
Zigman S (1993) Yearly review: ocular light damage. Photochem.
Photobiol. 57: 1060-1068.
Zigman S, Yulo T, & Schultz J (1974) Cataract induction in mice
exposed to near UV light. Ophthalmol Res, 6: 259-270.
Zigman S, Graff J, Yulo T, & Vaughen T (1975) The response of mouse
ocular tissue to continuous near-UV light exposure. Invest
Ophthalmol, 14: 710-713.
Zmudzka BZ & Beer JZ (1990) Activation of human immunodeficiency
virus by ultraviolet radiation. Photochem Photobiol, 52: 1153-1162.
Zuclich JA (1989) Ultraviolet induced photochemical damage in ocular
tissues. Health Physics 56: 671-682.
Zuclich J & Kurtin W (1977) Oxygen dependence of near-ultraviolet
induced corneal damage. Photochem Photobiol 25: 133-135.
Zundorf, I, & Hader, DP (1991) Biochemical and spectroscopic
analysis of UV effects on the marine flagellate, cryoptomonas
maculata, Arch Microbiol.
15. FUTURE RESEARCH
15.1 Introduction
At the United Nations Conference on the Environment and
Development (UNCED) in 1992 it was declared under Agenda 21 that
there should be activities on the effects of UV. Specifically:
(i) Undertake as a matter of urgency, research on the effects on
human health of increasing UV reaching the earth's surface as a
consequence of depletion of the stratospheric ozone layer;
(ii) On the basis of the outcome of this research, consider taking
appropriate remedial measures to mitigate the above mentioned
effects on human beings.
This report gives a thorough review of the health hazards of UV
exposure. However, although it is known that the burden of
UV-related diseases on human populations is high, the exact nature
and extent of these diseases is still largely unknown.
There is great uncertainty about future trends in atmospheric
ozone. For example the Antarctic holes and large depletions of ozone
that have occurred recently were not predicted in any of the ozone
depletion models. While agreements have been reached to reduce
releases of CFCs into the environment, and this will have future
benefit on the ozone layer, there is uncertainty about the extent of
ozone depletion caused by chemical pollutants. What is apparent is
that decreased ozone levels will persist for many years to come and
the corresponding increases in UV intensities will result in more
significant adverse health effects on all populations of the world
for many decades to come (WMO 1993).
The health effects of UV are not restricted to fair skinned
populations. UV exposure is thought to cause diseases of the eye and
suppression of the immune system in all populations of the world. UV
induced immune suppression may have adverse consequences on
infectious disease immunization programmes, particularly in areas
where the UV intensities are high. The possibility that UV will
cause progression of various diseases such as for HIV positive
patients still has to be elucidated. Many such important issues need
to be resolved as a matter of urgency.
The WHO Task Group reviewing this monograph strongly supported
action and coordination of UV research at the international level.
In particular the Task Group supported the concept of the
International Research Programme on Health, Solar UV Radiation and
Environmental Change (INTERSUN). INTERSUN is a collaborative effort
between WHO, the United Nations Environment Programme (UNEP) and the
International Agency on Cancer Research (IARC). The group recognised
specific research needs in areas of exposure assessment, terrestrial
plants, aquatic ecosystems, and human health effects related to the
skin, immune system and eye. Some of these could be accomplished
under the umbrella of INTERSUN, and some, at least initially, would
require more basic laboratory research to be undertaken.
15.2 INTERSUN
The objectives and approach of INTERSUN are: to accurately
evaluate the quantitative relationship between solar UV at the
surface of the earth and human health effects; develop reliable
predictions of health consequences of changes in UV; provide
baseline estimates of the occurrence of health effects of UV in
representative populations around the world; and develop practical
ways of monitoring change in these effects over time in relation to
environmental and behavioural change.
It is intended to establish field research centres covering a
range of latitudes in both hemispheres in which will be measured:
ground level UV irradiance; incidence of non-melanocytic skin
cancer; incidence and mortality of malignant melanoma; immune
function; biological markers of UV exposure and its' early
carcinogenic effects; constitutional sensitivity to sun exposure;
and present and past sun-related behaviour. In addition, in these
centres there will be undertaken case-control or cross-sectional
studies of relationships between skin cancers, cataract and immune
response and constitutional sensitivity to the sun, lifetime
exposure to the sun, and biological markers of UV exposure and early
carcinogenic effects. Surveys should be repeated at 5-yearly
intervals to allow trends in population measurements to be
established.
Data collected at such centres would be used
- to describe quantitatively the relationship between ground
level solar UV irradiance and the incidence of skin cancers and
other health effects (particularly eye damage and effects on
the immune system) of UV exposure in human populations.
- to estimate the change in occurrence of health effects of UV
radiation that would result from change in ground level solar
UV irradiance due to environmental change.
- to increase understanding of the relationship between personal
risk of health effects of UV radiation and constitutional
sensitivity to the sun and sun-related behaviour.
- to develop and validate appropriate ways of monitoring human
exposure to UV and the occurrence of associated health effects.
- to develop a network of centres monitoring trends in ground
level solar UV irradiance, sun exposure of populations, and the
occurrence of health effects of UV radiation.
- to interpret these trends, as far as is possible, in relation
to environmental change, changes in human behaviour, and the
implementation of public policies aimed at ameliorating
environmental change or human exposure to solar UV radiation.
- to provide a basis for development and evaluation of
interventions to reduce the occurrence of adverse health
effects of solar UV radiation.
In line with the above objectives and the gaps in knowledge
needed to be filled to make a satisfactory health risk assessment,
the following recommendations for future research are provided.
15.3 Solar and Personal UV Monitoring
The quantification of health detriment to an individual or to a
population requires the assessment of exposure of the individual or
the average exposure of a member of the exposed population. In
interpreting epidemiological data the exposures of the groups
comprising the study populations must be known. The requirements
here are precise measurement data of the environment occupied by the
exposed individuals combined with data on their exposure habits,
derived from representative personal exposure measurements, skin
damage or from questionnaires.
15.3.1 Solar monitoring
1. A comprehensive network of spectral and broad band monitoring
stations is needed worldwide giving quality ground measurements
of surface UV, to assess the effects of enhanced UVA and UVB
radiation on exposed populations, and to document the impact of
stratospheric ozone depletion on ambient UV. A long-term data
record is important.
2. Quality and convenient monitoring data are needed as input to
models and for model validation. Current models need to be
further modified to account for cloud cover, gaseous pollutants
and aerosols.
3. Common calibration and audit procedures should be developed and
implemented for national and international monitoring
programmes.
4. Monitoring activities should be coordinated with the efforts of
other groups on a national and international scale. Close
coordination is needed for inter-comparison studies and quality
control/quality assurance efforts.
5. There is a need for global monitoring of UV radiation, with
particular attention to long-term instrument stability and
representative geographical deployment
15.3.2 Personal monitoring
Population studies using personal UV monitoring devices are
needed to determine the fraction of the daily natural UV dose
received by persons at risk. The daily amount of UV received by
human skin varies greatly with occupation, behaviour, and local
climatic and environmental conditions. Little is known about these
factors and this seriously interferes with the interpretation of
existing data on the relationship between UV and the development of
skin cancer and of chronic skin and eye damage. Thus, the
development of personal UV monitoring devices is important.
Small passive personal UV dosimeters or monitoring devices
should be further developed and used to provide data on personal
exposures. Biomarkers for UV exposure and skin cancer risk should be
further developed, e.g. UV-induced mutations in p53 tumour
suppressor genes. Such biomarkers might constitute a person's own
"built in" UV risk monitoring device.
For some people, artificial sources of UV represents a
significant component of their exposure. Occupational exposures and
elective exposures such as from cosmetic sunbeds are important in
this respect. Measurement studies are required in these areas.
Standardized monitoring of exposure to the sun in at risk
populations, particularly children, should be evaluated as a guide
to public health action to control melanoma.
15.4 Terrestrial Plants
Particular attention must be given to the impact on food
production in the developing world and to development of crop
varieties resistant to higher levels of UV radiation.
Substantial research is needed to describe and evaluate the
effects of enhanced UV on plants, specifically in the following
areas:
1. Field validations on crop plants should be extended to
determine whether UV affects yield in other agriculturally
important plants.
2. Studies should be initiated to determine the impacts on
ecosystems. Little is known of the effects of UV in other
ecosystems such as forests, where 80% of net primary production
is currently stored.
3. It is important to study natural plants because they serve in
supplying new drugs, medicines and other natural products. They
also act as a reservoir of genetic diversity for modern crop
breeding programmes.
4. Studies are needed to elucidate how UVB effects are modified by
additional factors such as carbon dioxide, temperature, water
and nutrient stress, heavy metals, diseases, and pests.
5. Of increasing interest are man-made air pollutants such as
sulphur dioxide and nitrogen oxides, which are known to have
damaging effects on crops and forests. Attention should be
focused on whether the negative effects of these air pollutants
may be aggravated by UV radiation.
6. The manner and magnitude of adaptation to increased UV,
especially UVB, such as by increased screening pigments or
enhanced DNA damage repair capacity, need to be investigated
both as adaption capacity of individual plants and as
genetically based changes in plant populations. The latter also
provides a basis for plant breeding interventions for
agricultural and tree species.
15.5 Aquatic Ecosystems
Current data suggest that predicted increases in UV, especially
UVB radiation could have important negative effects in the marine
environment. However, uncertainties regarding the magnitude of these
effects remain large, including problems of extrapolating laboratory
findings to the open sea, and the almost complete absence of data on
long-term effects on ecosystems. Additional information is needed in
several areas before more reliable assessments of risk are possible.
Research is needed to:
1. Determine accurate and appropriate biological action spectra
for selected endpoints of key marine species.
2. Produce dose-response data on a greater variety of ecologically
important primary producers than is now available, as well as
data for key higher organisms within the food web.
3. Determine long-term effects for embryos or larvae exposed to
UVB radiation. Is the survival of the adult population (or
their offspring) affected?
4. Determine effects of enhanced UV on major ecosystems, including
the Antarctic ecosystems.
5. Obtain data on the mechanisms of damage and ranges of possible
adaptation or genetic selection in response to increased UVB
radiation.
6. Relate UVB penetration into the water column and through ice
both in laboratory and field research to determine effects on
phytoplankton and zooplankton.
7. Develop biomarkers to monitor current levels of UV damage in
phytoplankton and zooplankton.
15.6 Human Health
15.6.1 Skin
To a substantial degree, answers are still required to the
following questions regarding the relationship between sun exposure
and both melanoma and non-melanocytic skin cancer. Research is
needed to determine:
1. The quantitative relationship between radiant exposure and
incidence of these cancers; and in particular the shape of the
exposure-response relationship.
2. How the effect of sun exposure is modified by: age at exposure,
time since last exposure and pattern of exposure, particularly
whether a particular dose is received intermittently or
more-or-less continuously.
3. Biomarkers of UV exposure or early effects in the skin, e.g.
UV-induced mutations for UV exposure and skin cancer risk.
4. The importance of UVA in causing skin cancers, both relative to
UVB and comparatively between melanoma and non-melanocytic
cancer.
5. The importance of indirect (e.g. oxidative) as opposed to
direct UV damage in causing these cancers.
6. The contribution of sun exposure to the aetiology of these
cancers in dark-skinned populations and how, in general,
cutaneous sensitivity modifies both quantitatively and
qualitatively the relationship between sun exposure and skin
cancer.
7. If the associations of red hair and freckling with skin cancer
are explained by effects of skin pigmentation or sensitivity to
the sun or mediated by other susceptibility mechanisms.
8. If pigmentary characteristics or cutaneous sun sensitivity
explain most of the ethnic variation in skin cancer incidence
or are there other ethnically related susceptibility
characteristics.
9. Whether the relationships between sun exposure and BCC and SCC
are the same.
10. The role of UV-induced mutations of critical genes in causing
melanoma.
11. The role of sun exposure in causing cancer of the lip. An
epidemiological study is required with adequate control of
potential confounding with alcohol and tobacco use.
12. The relationship between use of sunlamps and sunbeds
(artificial tanning) and both melanoma and non-melanocytic skin
cancer. Adequate control of likely confounding with sun
exposure is required.
13. The effects of the interaction between UVB and the rest of the
solar spectrum in relation to DNA repair, malignant
transformation, and skin tumour development.
14. Additional animal models, particularly for the study of the
experimental induction of malignant melanoma. Further, an
action spectrum for induction of melanoma should be developed
in M. domestica to see if the apparent response of UVA
exposure observed in hybrid fish can be confirmed.
15.6.2 Immune System
Suppression of immune functions results from UV exposure of
humans. The following studies are needed to adequately assess the
consequent risk to human health.
1. Studies to determine whether systemic effects occur in humans
as a result of UV exposure.
2. Establish immunological biomarkers that could identify persons
who are susceptible to specific long-term UV-induced immune
suppression, e.g. to infectious agents.
3. Human and rodent studies are needed to determine the
relationship between UV exposure and susceptibility to
infection and vaccine effectiveness, and quantitative
comparison of the relative sensitivities of mice and humans.
4. Studies of the effects of UVB on immune function in human skin
of all types to determine if all skin types show the same
quality and quantity of effects.
5. Studies to examine whether currently available sunscreens
provide adequate protection and whether dietary supplements are
protective. Specifically:
a) Protective effects for immunosuppression;
b) Systemic toxicity based on penetration of organic
chemicals in sunscreens;
c) Carcinogenicity assays.
6. Studies of the spectral effectiveness of UV in producing immune
function defects and enhanced susceptibility to disease.
7. Research to determine the role of UVB exposure in
immediate-type hypersensitivity (allergic) reactions as well as
specific antibody responses, particularly as it relates to
impact on incidence and severity of asthma.
8. Research on the effects of UV on the development of autoimmune
disease and on underlying mechanisms.
9. Studies on systemic effects caused by release of cytokines
within the epidermis and blood stream.
15.6.3 Eye
Two issues underscore the need for undertaking studies of
health risks of UV exposure of the eye on a priority basis. First,
there is evidence of the depletion of the ozone layer and the
reported consequent increase in ambient UV which will impact on many
eye diseases. Second, the overall ageing of the population worldwide
would further compound the magnitude of the cataract and other eye
problems.
1. There is a need for well-conducted epidemiological studies to
be undertaken to define the strength of the association between
UVA and UVB exposure with cataract and other eye conditions,
including climatic droplet keratopathy and pterygium, at both
the individual and community levels. Ideally, this should be
undertaken in a variety of locations. Careful attention needs
to be given to quantifying, for each study participant, ocular
exposure to UV and exposure to other possible risk factors that
may confound observed associations.
2. Techniques need to be refined for measuring the proportion of
UV of different wavelengths incident upon the human cornea and
lens, and how these are affected by the behavioural response to
avoid direct exposure to bright sunlight, and by different
ground surfaces.
3. Studies should be conducted to provide data on the quantitative
relationship between exposure to UV and cataract and other
lesions of the anterior eye. In relation to this there is an
urgent need to establish an internationally agreed system for
grading the type and severity of cataract.
4. Investigations of the role of personal solar exposure in
causing ocular melanoma, including the protective effect of
ocular devices.
5. The relationship between sunlamp or sunbed exposure and
malignant melanoma of the eye needs to be evaluated.
6. The relative efficacy of different protective devices, headwear
and eye wear needs to be determined.
7. Techniques need to be defined for measuring the proportion of
UV of different wavelengths penetrating the human cornea and
lens in the intact eye, in view of possible retinal effects.
8. The action spectra for chronic exposure leading to ocular
damage needs to be determined in primates and other animals.
This includes action spectra for cataract formation, pterygium,
climatic droplet keratopathy, and macular changes in aphakic
animals.
9. Risks to aphakic people and those with intraocular lens
implants for retinal changes, mainly from UVA and short
wavelengths in the visible part of the spectrum, need to be
determined.
15.7 Laboratory Studies
Little is known about the mechanisms of interaction of UV and
environmental chemical agents on biological systems. Many widely
distributed natural or artificial chemicals (pesticides,
halocarbons, etc.) can be altered by UV, resulting in photoproducts
that may be less or more biologically effective than the parent
compound. Furthermore, many chemicals can be activated by UV in
situ in biological systems and this activation may elicit a
biological effect which neither the chemical nor the radiation alone
exhibits (Psoralens). An international registry of agents that
interact with light to cause adverse health effects would speed
identification of such agents.
Much of the information on the chemical and biological effects
of UV comes from experiments using UVC (particularly 254 nm)
radiation not found in sunlight reaching the earth's surface. There
are studies showing direct and indirect effects on cells and
cellular constituents of UVB, UVA, and visible light that differ
considerably from those of UVC. Thus, the chemical and biological
effects of the wavelengths of UV found in sunlight should be
studied. There is evidence that visible light can, under different
conditions, either help cells to repair UV-induced damage or can
potentiate the detrimental effects of UV. Thus, to better understand
the effects of sunlight on humans and the environment, experiments
should be performed using natural sunlight or artificial lamps with
well-known continuous spectra.
Cellular and molecular studies needed include:
1. Extension of relevant biological action spectra to the
UVA/visible wavelengths, as well as interactions between UVA
and UVB radiation. Crucial spectra required include:
- pre-mutagenic lesions
- gene activation
- activation of viruses specially HIV (activation of DNA
binding, promoter activity, viral replication)
2. Determination of the free radical/oxidative component of UVB
and UVA radiation effects. Determine the reactive intermediates
involved.
3. Determination of adaptation responses in human cells.
4. Determination of endogenous cellular antioxidant defences in
eye tissue and skin.
5. Elucidation of DNA repair pathways in humans using the advanced
technology available for analyses of gene and function.
6. Identification of key endogenous and exogenous sensitizers.
7. Studies to examine the effect of UV on the balance between Th1
and Th2 cells.
15.8 Education
It is essential to educate the general population and workers
concerning the profound importance of sunlight and the possibilities
of either UV deprivation or of acute and chronic UV injury. It is
also important to overcome the lack of respect for the possible
adverse effects on health from overexposure to sunlight, simply
because sunlight is ubiquitous, and the concept that, if something
is natural, it must be totally beneficial and safe. In this context
there is a need to standardize a UV exposure index that can be used
as part of health information/education campaigns.
15.9 Administration
An international panel composed of key national experts, with
its secretariat at WHO, should be formed with the following terms of
reference:
(i) to identify gaps in knowledge and set research priorities
needed for a better health risk assessment of exposure to UV
(ii) to evaluate and monitor progress of research
(iii) to establish mechanisms for funding research and initiate
projects where crucial gaps in knowledge have been identified
(iii) to facilitate international cooperation on UV monitoring
efforts, including instrumentation intercomparisons,
calibration standardization and uniform analysis of data, and
(iv) to facilitate international cooperation in measurement of
human UV exposure, monitoring of trends in occurrence of UV
health effects, and epidemiological studies of the
relationship between UV exposure and health effects
(v) to provide a database of research projects to facilitate
cooperation between researchers and institutions involved in
research.
16. PREVIOUS EVALUATIONS BY INTERNATIONAL BODIES
16.1 United Nations Environment Programme
UNEP has undertaken a number of reviews on the environmental
effects of increased UV intensities resulting from ozone depletion
(1989, 1991, 1992). Their findings on each issue are summarized
below:
16.1.1 Ozone
Significant global scale decreases in total ozone have occurred
over the past ten years. All other factors being constant, there is
no scientific doubt that decreases in total ozone will increase UVB
radiation at ground level. Tropospheric ozone and aerosols may have
masked the consequences of stratospheric ozone depletion for UVB in
some industrialized regions. There are no reliable estimates of the
direction or magnitude of effects of any cloud cover trends on UVB.
Efforts to improve local and regional air quality may bring to light
the increases in UVB associated with the depletion of stratospheric
ozone.
16.1.2 Human health
The induction of immunosuppression by UVB has now been
demonstrated in humans, not only those of light pigmentation, but
also deeply pigmented individuals. This places all of the world's
populations at risk of the potential adverse impacts of UVB on the
immune system, including possible increases in the incidence or
severity of infectious disease.
An increased number of adverse ocular effects have been
associated with exposure to UV. These include age-related
nearsightedness, deformation of the lens capsule, and nuclear
cataract (a form of cataract which previous information excluded
from consideration). These effects appear to be independent of
pigmentation. Estimates of risk would increase slightly if one were
to include nuclear cataract among the forms of cataract increasing
with ozone depletion. It is now predicted that, all other things
being equal, a sustained 10% decrease in ozone will be associated
with between 1.6 and 1.75 million additional cases of cataract per
year world-wide.
Recent information on the relationship of nonmelanoma skin
cancer to UV exposures confirms previous findings and has allowed
refinement of the carcinogenic action spectrum. Incorporation of
this new information into the risk estimation process has led to
slightly lower predictions. It is now predicted that a sustained 10%
decrease in ozone will be associated with 26% increase in
non-melanoma skin cancer. All other things remaining constant, this
would mean an increase in excess of 300,000 cases per year
world-wide.
16.2 International Agency for Research on Cancer
The carcinogenicity of ultraviolet has been evaluated by IARC
(1992, 1993). IARC concluded that "There is sufficient evidence in
humans for the carcinogenicity of solar radiation. Solar radiation
causes cutaneous melanoma and nonmelanocytic skin cancer". With
respect to other potential sources of UV, IARC has concluded that
there is limited evidence for the carcinogenicity of exposure to
fluorescent lighting and there is inadequate evidence for the
carcinogenicity of other artificial sources of UV. Readers are
referred to these excellent monographs for more details -much of
which has been incorporated in this text.
16.3 World Health Organization
WHO and its Regional Office for Europe have completed reviews
on the health effects of ultraviolet (WHO, 1979, 1989). They
concluded that all people are exposed to UV from sunlight, and the
risk to health varies with geographical, genetic and other factors.
Similar risks are involved in the increasing exposure of people to
UV from artificial sources, such as those used for suntanning, in
phototherapy and in industrial processes. The biological effects of
a single exposure differ significantly from the effects of repeated
and cumulative exposures. Both types of risk increase markedly with
excessive exposure.
Most observed biological effects of UVB are extremely
detrimental to living organisms. Much less is known about the
biological effects of UVA. It can augment the biological effects of
UVB, and doses of UVA, which alone do not show any biological
effects, can in the presence of certain chemical agents, result in
injury to tissues (phototoxicity, photoallergy, enhancement of
photocarcinogenesis).
All those who work out of doors are potentially at risk of
overexposure, the consequences of which may be both acute and
long-term effects. The fashion of exposing a large part of the body
to sunlight has during recent years increased the exposure of the
skin, resulting in quite high UV doses. This is true not only for
outdoor work but is now also normal during leisure periods, as
exemplified by the holiday exodus of a large part of the population
of the northern European countries to the Mediterranean coast.
16.4 International Commission on Non-Ionizing Radiation Protection
Through its predecessor committee IRPA/INIRC the ICNIRP was
involved in the publication of the original review of the health
effects of UV (WHO/UNEP/IRPA, 1979). It has recommended
international guidelines on limits (IRPA/INIRC, 1991) as shown in
Chapter 13. It has also recommended against the use of sunlamps for
cosmetic purposes (IRPA/INIRC, 1991a). A review of the studies on
fluorescent lighting with reference to UV levels has suggested that
they do not seem to be associated with an increased risk of
malignant melanoma (IRPA/INIRC, 1991).
REFERENCES
Abrahams PJ, Huitema BA, & van der Eb AJ (1984) Enhanced
reactivation and enhanced mutagenesis of herpes simplex virus in
normal human and xeroderma pigmentosum cells. Mol Cell Biol, 4:
2341-2346.
ACGIH (1993) Threshold limit values for chemical substances and
physical agents and biological exposure indices. Cincinnati, Ohio,
The American Conference of Governmental Industrial Hygienists.
Adam SA, Sheaves JK, Wright NH, Mosser G, Harris RW, & Vessey MP
(1981) A case-control study of the possible association between oral
contraceptives and malignant melanoma. Br J Cancer, 44: 45-50.
Adams JS, Clemens TL, Parrish JA, (1982) Vitamin D synthesis and
metabolism after UV irradiation of normal and vitamin D deficient
subjects. N Engl J Med, 306: 722-725.
Ainsleigh HG (1993) Beneficial effects of sun exposure on cancer
mortality. Prev Med, 22: 132-140.
Akslen LA & Morkve O (1992) Expression of p53 protein in cutaneous
melanoma. Int J Cancer, 52: 13-16.
Albino AP, Nanus DM, Davis ML, & McNutt NS (1991) Lack of evidence
of Ki- ras codon 12 mutations in melanocytic lesions. J Cutan
Pathol, 18: 273-278.
Alcalay J, Freeman SE, Goldberg LH, & Wolf JE (1990) Excision repair
of pyrimidine dimers induced by simulated solar radiation in the
skin of patients with basal cell carcinoma. J Invest Dermatol, 95:
506-509.
Anderson RR & Parrish JA (1981) The optics of human skin. J Invest
Dermatol, 77: 13-19.
Andley UP (1987) Photodamage to the eye. Yearly review. Photochem
Photobiol, 46: 1057-1066.
Angel P, Poting A, Mallick U, Rahmsdorf HJ, Schorpp M, & Herrlich P
(1986) Induction of metallothionein and other mRNA species by
carcinogens and tumour promoters in primary human skin fibroblasts.
Mol Cell Biol, 6: 1760-1766.
Ansel JC, Mountz J, Steinberg AD, DeFabo E, & Green Ira (1985)
Effects if UV radiation on autoimmune strains of mice: Increased
mortality and accelerated autoimmunity in BXSB male mice. J Invest
Dermatol, 85: 181-186.
Araneo BA, Dowell T, Moon HB, & Daynes RA (1989) Regulation of
murine lymphokine production in vivo. Ultraviolet radiation
exposure depresses Il-2 and enhances IL-4 production by T cells
through an Il-1 dependent mechanism. J Immunol, 143: 1737-1744.
Arlett CF & Harcourt SA (1983) Variation in response to mutagens
amongst normal and repair-defective human cells. In: Lawrence CW ed.
Induced mutagenesis: Molecular mechanisms and their implications for
environmental protection. New York, London, Plenum Press, pp
249-267.
Arlett CF, Harcourt SA, Cole J, Green MH, & Anstey AV (1992) A
comparison of the response of unstimulated and stimulated
T-lymphocytes and fibroblasts from normal, Xeroderma pigmentosum and
trichothiodystrophy donors to the lethal action of UVC. Mutat Res,
273: 127-135.
Armstrong BK (1984) Melanoma of the skin. Br Med Bull, 40: 346-350.
Armstrong BK (1988) Epidemiology of malignant melanoma: intermittent
or total accumulated exposure to the sun? J Dermatol Surg Oncol, 14:
835-849.
Armstrong BK (1993) Implications of increased solar UVB for cancer
incidence. In: Chanin ML ed. The role of the stratosphere in global
change. Berlin, Heidelberg, New York, Springer-Verlag, pp 517-540.
Armstrong BK & English DR (1988) The epidemiology of acquired
melanocytic naevi and their relationship to malignant melanoma.
Pigment Cell, 9: 27-47.
Armstrong BK, Woodings TL, Stenhouse NS, & McCall MG (1983)
Mortality from cancer in migrants to Australia 1962 to 1971. Perth,
University of Western Australia.
Armstrong BK, de Klerk NH, & Holman CDJ (1986) Etiology of common
acquired melanocytic nevi: constitutional variables, sun exposure
and diet. J Natl Cancer Inst, 77: 329-335.
Atkin M, Fenning J, Heady JA, Kennaway EL, & Kennaway NM (1949) The
mortality from cancer of the skin and lip in certain occupations. Br
J Cancer, 3: 1-15.
Aubry F & MacGibbon B (1985) Risk factors of squamous cell carcinoma
of the skin. A case-control study in the Montreal region. Cancer,
55: 907-911.
Auerbach H (1961) Geographic variation in incidence of skin cancer
in the United States. Public Health Rep, 76: 345-348.
Augustsson A, Stierner U, Rosdahl I, & Suurküla M (1990) Melanocytic
naevi in sun-exposed and protected skin in melanoma patients and
controls. Acta Dermato-Venereol, 71: 512-517.
Azizi E, Lusky A, Kushelevsky AP, & Schewach-Millet N (1988) Skin
type, hair colour and freckles are predictors of decreased minimal
erythema ultraviolet radiation dose. J Am Acad Dermatol, 19: 32-38.
Azizova AO, Islomov AI, Roshchupkin DI, Predvoditelev DA, Remizov
AN, & Vladimir YA (1980) Free radicals formed on ultraviolet
irradiation of the lipids of biological membrane. Biophysics, 24:
407-414.
Baadsgaard O, Wulf HC, Wantzin GL, & Cooper KD (1987) UVB and UVC,
but not UVA, potently induce the appearance of T6-DR+
antigen-presenting cells in human epidermis. J Invest Dermatol, 89:
113-118.
Baadsgaard O, Lisby S, Wulf HC, Wantzin GL, & Cooper KD (1989) Rapid
recovery of Langerhans cell alloreactivity, without induction of
autoreactivity, after in vivo ultraviolet A, but not ultraviolet B
exposure of human skin. J Immunol, 142: 4213-4218.
Baadsgaard O, Salvo B, Mannie A, Dass B, Fox DA, & Cooper KD (1990)
in vivo ultraviolet-exposed human epidermal cells activate T
suppressor cell pathways that involve CD4+CD45RA+ suppressor-inducer
T cells. J Immunol, 145: 2854-2861.
Baasanhu J, Johnson GJ, Burendei G, & Minassian DC (in press)
Prevalence and causes of blindness and visual impairment in
Mongolia: A survey of populations aged 40 and older. Bull. World
Health Organ.
Bachem A (1956) Ophthalmic ultraviolet action spectra. Am J
Ophthalmol, 41: 969-975.
Baird EA, McHenry PM, & MacKie RM (1992) Effect of maintenance
chemotherapy in childhood on numbers of melanocytic naevi. Br Med J,
305: 799-801.
Baker KS & Smith RC (1982) Spectral irradiance penetration in
natural waters. In: Calkins J ed. The role of solar ultraviolet
radiation in marine ecosystems. New York, London, Plenum Press, pp
223-246.
Barbareschi M, Girlando S, Cristofolini P, Cristofolini M, Togni R,
& Boi S (1992) p53 Protein expression in basal cell carcinoma.
Histopathology, 21: 579-581.
Barker & Brainard (1993) Personal communication.
Barker JNWN & MacDonald DM (1988) Eruptive dysplastic naevi
following renal transplantation. Clin Exp Dermatol, 13: 123-125.
Barnes PW, Jordan PW, Gold WG, Flint SD, & Caldwell MM (1988)
Competition, morphology and canopy structure in wheat (Triticum
aestivum L.) and wild oat (Avena fatua L.) exposed to enhanced
ultraviolet-B radiation. Funct Ecol, 2: 319-330.
Barnes PW, Flint SD, & Caldwell MM (1990) Morphological responses of
crop and weed species of different growth forms to ultraviolet-B
radiation. Am J Bot, 77: 1354-1360.
Barrett SF, Robbins JH, Tarone RE, & Kraemer KH (1991) Evidence for
defective repair of cyclobutane pyrimidine dimers with normal repair
and other DNA photoproducts in a transcriptionally active gene
transfected into Cockayne syndrome cells. Mutat Res, 255: 281-291.
Basu-Modak S & Tyrrell RM (1993) Singlet oxygen. A primary effector
in the UVA/near visible light induction of the human heme oxygenase
gene. Cancer Res. 53: 4505-4510.
Bech-Thomsen N, Wulf HC, Poulsen T, & Lundgren K (1988a)
Pretreatment with long-wave ultraviolet light inhibits
ultraviolet-induced skin tumor development in hairless mice. Arch
Dermatol, 124: 1215-1218.
Bech-Thomsen N, Wulf HC, & Lundgren K (1988b) Pre-treatment with UVA
influences broad-spectrum UV photocarcinogenesis in hairless mice
(Abstract). In: Rikles E ed. Proceedings of the 10th International
Congress on Photobiology, Jerusalem. Jerusalem, Israel,
International Association of Biology, p 34.
Beer JZ & Smudzka BZ (1991) Activation of human immunodeficiency
virus by radiation. In: Seymour CB & Murhersill C ed. New
developments in fundamental and applied radiology. London, Taylor
and Francis, pp 113-123.
Beitner H, Norell SE, Ringborg U, Wennersten G, & Mattson B (1990)
Malignant melanoma: aetiological importance of individual
pigmentation and sun exposure. Br J Dermatol, 122: 43-51.
Beral V & Robinson N (1981) The relationship of malignant melanoma,
basal and squamous skin cancers to indoor and outdoor work. Br J
Cancer, 44: 886-891.
Beral V, Evans S, Shaw H, & Milton G (1982) Malignant melanoma and
exposure to fluorescent lighting at work. Lancet, ii: 290-293.
Berger DS & Urbach F (1982) A climatology of sunburning ultraviolet
radiation. Photochem Photobiol, 35: 187-192.
Bergmanson JP, Pitts DG, & Chu LW (1988) Protection from harmful UV
radiation by contact lens. J Am Optom Assoc, 59:,178-182.
Bergmanson JP, Pitts DG, & Chu LW (1987) The efficacy of a
UV-blocking soft contact lens in protecting cornea against UV
radiation. Acta Opthtalmol, 65: 279-286.
Beukers R & Berends W (1960) Isolation and identification of the
irradiation product of thymine. Biochim Biophys Acta, 41: 550-551.
Bhatnagar R, West KP, Vitale S, Sommer A, Joshi S, & Venkataswamy G
(1991) Risk of cataract and history of severe diarrhoeal disease in
Southern India. Arch Ophthalmol, 109: 696-699.
Bhuyan, K.C. and Bhuyan, D.K. (1983) Molecular mechanism of
caractogenesis I reactive species of oxygen as triggering agents II.
Evidence of lipid peroxidation and membrane damage. In Oxy Radicals,
their scavenger syst. Proc. Int. Conf. Superoxide Dismutase 3rd,
vol. 2 (Ed. G. Cohen and R.A. Greenwald), pp 343-356, Elsevier, New
York.
Bidigare RR (1989) Potential effects of UVB radiation on marine
organisms of the Southern Ocean: distributions of phytoplankton and
krill during Austral spring. Photochem Photobiol, 50: 469-477.
Birch-Hirschfeld A (1914) The pathological effect of radiant energy
on the eye. 1. Blinding by sunlight. Ergebn Allg Path des Menschen
und der Tiere 16, 619-634.
Bishop L (1992) Statistical considerations in network design and
data analysis. Presentation to UVB Workshop: A review of the science
and status of measuring and monitoring programs. Washington, DC,
Science and Policy Associates, Inc.
Bissett DL, Hannon DP, & Orr TV (1989) Wavelength dependence of
histological, physical and visible changes in chronically
UV-irradiated hairless mouse skin. Photochem Photobiol, 50: 763-769.
Bittersmann E, Holzwarth AR, Agel G, & Nultsch W (1988) Picosecond
time-resolved emission spectra of photoinhibited and photobleached
Anabaena variabilis. Photochem Photobiol, 47: 101-105.
Black HS (1987a) Photocarcinogenesis and diet. Fed Proc, 46:
1901-1905.
Black HS (1987b) Potential involvement of free radical reactions in
ultraviolet light mediated cutaneous damage. Photochem Photobiol,
46: 213-221.
Black HS & Chan JT (1975) Suppression of ultraviolet light-induced
tumor formation by dietary antioxidants. J Invest Dermatol, 65:
412-412.
Black HS, Thornby JI, Gerguis J, & Lenger, W (1992) Influence of
dietary omega-6,-3 fatty acid sources on the initiation and
promotion stages of photocarcinognesis. Photochem Photobiol, 56:
195-199.
Blum, HF (1948) Sunlight as a causal factor in cancer of the skin of
man. J Natl Inst Cancer, 9: 247-258.
Blum HF, Butler EG, Dailey TH, Daube JR, Mawe RC, & Soffen GA (1959)
Irradiation of mouse skin with single doses of ultraviolet light. J
Natl Cancer Inst, 22: 979-993.
Blumthaler M, Ambach W, & Daxecker F (1987) On the threshold radiant
exposure for keratitis solaris. Invest Opthalmol Vis Sci, 28:
1713-1716.
Blyth WA, Hill TJ, Field HJ, & Harbour DA (1976) Reactivation of
herpes simples virus infection by ultraviolet light and possible
involvement of prostaglandins. J Gen Virol, 33: 547-550.
Bochow TW, West SK, Azar A, Munoz B, Sommer A, & Taylor HR (1989)
Ultraviolet light exposure and risk of posterior subcapsular
cataracts. Arch Ophthalmol, 107: 369-372.
Boettner EA & Wolter JR (1962) Transmission of the ocular media.
Invest Ophthalmol Vis Sci, 1: 776-783.
Boiteux S, O'Connor TR, & Laval J (1987) Formamido pyrimidine-DNA
glycosylase of Escherichia coli: cloning and sequencing of the fpg
structural gene and overproduction of the protein. EMBO J, 6:
3177-3183.
Booth F (1985) Heredity in one hundred patients admitted for
excision of pterygia. Aust N Z J Ophthalmol, 13: 59-61.
Bornman JF & Vogelmann TC (1991) Effect of UV-B radiation on leaf
optical properties measured with fibre optics. J Exp Vot, 42:
547-554.
Bose B, Agarwal S & Chafterjee SN (1989) UV-A induced lipid
peroxidation in liposomal membrane. Radiat. Environ. Biophys. 28
59-65.
Bose SN & Davies RJH (1984) The photoreactivity of T-A sequences in
oligodeoxyribonucleotides and DNA. Nucl Acids Res, 12: 7903-7913.
Bouwes-Bavinck JN (1992) Risk of skin cancer in renal-transplant
recipients. Leiden, The Netherlands (Thesis).
Boyle J, Mackie RM, Briggs JD, Junor BJR, & Aitchison TC (1984)
Cancer, warts, and sunshine in renal transplant patients: A
case-control study. Lancet, 1: 702-705.
Bradley MO, Hsu IC, & Harris CC (1979) Relationships between sister
chromatid exchange and mutagenicity, toxicity and DNA damage. Nature
(Lond), 282: 318-320.
Brash DE & Haseltine WA (1982) UV-induced hotspots occur at DNA
damage hotspots. Nature (Lond), 298: 189-192.
Brash DE, Rudolph JA, Simon JA, Lin A, McKenna GJ, Baden HP,
Halperin AJ, & Pontén J (1991) A role for sunlight in skin cancer:
UV-induced p53 mutations in squamous cell carcinoma. Proc Natl Acad
Sci (USA), 88: 10124-10128.
Braun J (1991) The protective function of phenolic compounds of rye-
and oat seedlings against UV-B radiation and their biosynthetic
regulation, (Thesis) pp. 1-237 in Karlsr Beitr Entw Okophysiol 9, M.
Tevini (ed.), Bot Inst II, Karlsruhe.
Braun J & Tevini M (1991) Regulation of UV-Protective pigment
synthesis in the epidermal layer of rye seedlings ( Secale cereale
L. cv. Kustro). Photochem Photobiol, 37: 318-321.
Bridges BA (1990) Sunlight, DNA damage and skin cancer: a new
perspective. Jpn J Cancer Res, 81: 105-107.
Brilliant LB, Grasset NC, Pokhrel RP, Kolstad A, Lepkowski JM,
Brilliant GE, Hawks WN, & Pararajasegaram R (1983) Associations
among cataract prevalence, sunlight hours, and altitude in the
Himalayas. Am J Epidemiol, 118: 250-264.
Brodkin RH, Kopf AW, & Andrade R (1969) Basal-cell epithelioma and
elastosis: a comparison of distribution. In: Urbach F ed. Biologic
effects of ultraviolet radiation. Oxford, New York, Pergamon Press,
pp 581-618.
Brühl C & Crutzen PJ (1989) On the disproportionate role of
tropospheric ozone as a filter against solar UV radiation. Geophys
Res Lett, 16: 703-706.
Bruls WAG, Slaper H, Van der Leun JC, & Berrens L (1984)
Transmission of human epidermis and stratum corneum as a function of
thickness in the ultraviolet and visible wavelengths. Photochem
Photobiol, 40(4): 485-494.
Bhuyan KC & Bhuyan DK (1983) Molecular mechanism of
cataractogenesis, I. Reactive species of oxygen as triggering
agents, II. Evidence of lipid peroxidation and membrane damage. In
(ed. Cohen G & Greenwald RA) Oxy Radicals, their scavenger syst.
Proc. Int. Conf. Superoxide Dismutase 3rd, vol 2, Elsevier, New
York, pp 343-356.
Cadet J, Ravanat JC, Buchko GW, Yeo HC, & Ames BN (in press) Singlet
oxygen DNA damage: chromatographic and mass spectrometry analysis of
damage products. Methods Enzymol.
Caldwell MM (1976) The effect of solar UV-B radiation (280-315 nm)
on higher plants: implications of stratosphere ozone reduction. In:
Castellani A ed. Research in photobiology. New York, London, Plenum
Press, pp 597-607.
Caldwell MM, Termura AH, & Tevini M (1989) The changing solar
ultraviolet climate and the ecological consequences for higher
plants. Trends Ecol Evol, 4: 363-366.
Cameron ME (1965) Pterygium throughout the world. Springfield,
Illinois, Charles C. Thomas.
Campbell CC, Quinn AG, & Rees JL (1993) Codon 12 Harvey-ras
mutations are rare events in non-melanoma human skin cancer. Br J
Dermatol, 128: 111-114.
Carretto JJ, Carignana MO, Daleo G, & de Marco SG (1990) Occurrence
of mycosporine-like amino acids in the red tide dinoflagellate
Alexandrium excavarum, UV-photoprotective compounds. J Plankton Res,
12: 909-921.
Cartwright LE & Walter JF (1983) Psoralen-containing sunscreen is
tumorigenic in hairless mice. J Am Acad Dermatol, 8: 830-836.
Cerutti PA & Netrawali M (1979) Formation and repair of DNA damage
induced by indirect action of ultraviolet light in normal and
xeroderma pigmentosum skin fibroblasts. Radiat Res, Suppl: 423-432.
Cervenka J, Witkop CJ, Okoro AN, & King RA (1979) Chromosome breaks
and sister chromatid exchanges in albinos in Nigeria. Clin Genet,
15: 17-21.
Césarini JP & Muel B (1989) Erythema induced by quartz- halogen
sources. Photodermatology, 6: 222-227.
Challoner AVJ, Corless D, & Davis A (1976) Personnel monitoring of
exposure to ultraviolet radiation. Clin Exp Dermatol, 1: 175-179.
Chamberlain J & Moss SH (1987) Lipid peroxidation and other membrane
damage produced in Escherichia coli K1060 by near-UV radiation and
deuterium oxide. Photochem Photobiol, 45: 625-630.
Chamberlin GJ & Chamberlin DG (1980) Colour: It's measurement,
computation and application. London, Heyden & Son, p 46.
Chan GL, Peak MJ, Peak JG, & Haseltine WA (1986) Action spectrum for
the formation of endonuclease-sensitive sites and (6-4)
photoproducts induced in a DNA fragment by ultraviolet radiation.
Int J Radiat Biol, 50: 641-648.
Chatterjee A, Milton RC, & Thyle S (1982) Prevalence and aetiology
of cataract in Punjab. Br J Ophthalmol, 66: 35-42.
Christman MF, Morgan RW, Jacobson FS, & Ames BN (1985) Positive
control of a regulon for defences against oxidative stress and some
heat shock proteins in Salmonella typhimurium. Cell, 41: 753-762.
Chu G & Chang E (1988) Xeroderma pigmentosum group E cells lack a
nuclear factor that binds to damaged DNA. Science, 242: 564-567.
CIE (1987) International lighting vocabulary published jointly by
the International Electrotechnical Commission (IEC) and the
International Commission on Illumination (CIE). Geneva,
International Electrotechnical Commission (ISBN 2-8273-0006-0).
CIE (1990) Report on photosensitising chemicals - Updated in 1993.
Vienna, International Commission on Illumination.
CIE (1991) Sunscreen testing (UVB). Vienna, Commission on
Illumination (CIE Technical report - Publication No. CIE 90).
Cleaver JE (1973) Xeroderma pigmentosum - progress and regress. J
Invest Dermatol, 60: 374-380.
Clemmesen J (1965) Statistical studies in the aetiology of malignant
neoplasms. I. Review and results. Copenhagen, Munksgaard, p 409.
Coffman RL, Seymour WP, Lebman DA, Hirakkki DD, Christiansen JA,
Shrader B, Cherwinski HM, Savelkoul HF, Finkelman FD, Bond MW, &
Mosmann TR (1988) The role of helper T cell products in mouse B cell
differentiation and isotype regulation. Immunol Rev, 102: 5-28.
Cogan DG & Kinsey VE (1946) Action spectrum for keratitis produced
by ultraviolet radiation. Arch Ophthalmol, 35: 670-677.
Cole CA, Forbes PD, & Davies RE (1986) An action spectrum for UV
photocarcinogenesis. Photobiology, 43: 275.
Colin J, Bonissent JF, & Resnikoff S (1985) Epidemiology of the
exfoliation syndrome. Proceedings of the 17th Congress of the
European Society of Ophthalmolology, Helsinki, pp 230-231.
Collmann GW, Shore DL, Shy CM, Checkoway H, & Luria AS (1988)
Sunlight and other risk factors for cataract: an epidemiological
study. Am J Public Health, 78: 1459-1462.
Colston K, Berger U, & Coombes RC (1989) Possible role for vitamin D
in controlling breast cancer cell proliferation. Lancet, 1: 188-191.
Connor MJ & Wheeler LA (1987) Depletion off cutaneous glutathione by
ultraviolet radiation. Photochem Photobiol, 46: 239-245.
Cooke KR & Fraser J (1985) Migration and death from malignant
melanoma. Int J Cancer, 36: 175-178.
Coombs BD, Sharples KJ, Cooke KR, Skegg DCG, & Elwood JM (1992)
Variation and covariates of the number of benign nevi in
adolescents. Am J Epidemiol, 136: 344-355.
Cooper KD (1993) Human studies in photo immunology. Photochem
Photobiol, 57: 745.
Cooper KD, Neises GR, & Katz SI (1986) Antigen-presenting OKM5+
melanophages appear in human epidermis after ultraviolet radiation.
J Invest Dermatol, 861: 363-370.
Cooper KD, Oberhelman BS, Hamilton MS, Baadsgaard O, Terhune M,
LeVee G, Anderson T, & Koren H (1992) UV exposure reduces
immunization rates and promotes tolerance to epicutaneous antigens
in humans; relationship to dose, CD1a-DR+ epidermal macrophage
induction and Langerhans cell depletion. Proc Natl Acad Sci (USA),
89: 8497-8501.
Corominas M, Kamino H, Leon J & Pellicer A (1991) Oncogene
activitation in human benign tumors of the skin (keratoacanthomas):
Is H-ras involved in differentiation as well as proliferation? Proc
Natl Acad Sci (USA), 86: 6372-6376.
Coronel VP, Dai QJ, Vergara BS, & Teramura AH (1990) Preliminary
study on response of rice seedlings to enhanced UV-B radiation. Int
Rice Res Newsl, 15: 37.
Coroneo MT (1993) Pterygium as an early indicator of ultraviolet
insolation: a hypothesis. Br J Ophthalmol, 77: 734-739.
Correll DL, Clark CO, Goldberg B, Goodrich VR, Hayes DR, Klein WH, &
Schecher WD (1992) Spectral ultraviolet-B radiation fluxes at the
earth's surface: long-term variations at 39°N, 77°W. J Geogr Res,
97: 7579-7591.
Cox CWJ (1987) Ultraviolet irradiance levels in welding processes.
In: Passachier WF & Bosnjakovic BFM ed. Human exposure to
ultraviolet radiation: Risks and regulations. Amsterdam, Oxford, New
York, Elsevier Science Publishers, pp 383-386.
Cristofolini M, Franceschi S, Tasin L, Zumiani G, Piscioli F,
Talamini R, & La Vecchia C (1987) Risk factors for cutaneous
malignant melanoma in a northern Italian population. Int J Cancer,
39: 150-154.
Crombie IK (1981) Distribution of malignant melanoma on the body
surface. Br J Cancer, 43: 842-849.
Cruickshanks KJ, Klein R, & Klein BE (1993) Sunlight and age-related
macular degeneration: the Beaver Dam Eye Study. Arch Ophthalmol,
111: 514-518.
Cunningham ML, Johnson JS, Giovanazzi SM, & Peak MJ (1985)
Photosensitized production of superoxide anion by monochromatic
(290-405 nm) ultraviolet irradiation of NADH and NADPH coenzymes.
Photochem Photobiol, 42: 125-128.
Cyrlin MN, Pedris-Leftick A, & Sugar J (1980) Cataract formation in
association with ultraviolet photosensitivity. Ann Ophthalmol, 12:
786-790.
Czochralska B, Bartosz W, & Shugar D (1984) Oxidation of
excited-state NADH and NAD dimer in aqueous medium - involvement of
O2- as a mediator in the presence of oxygen. Biochim Biophys Acta,
801: 403-409.
Danielle RP (1988) Pathophysiology of the asthmatic syndromes. In:
Danielle RP ed. Immunologic diseases of the lung. Boston, Blackwell
Scientific Publishers, pp 503-516.
Dardanoni L, Gafá L, Paterno R, & Pavone G (1984) A case- control
study on lip cancer risk factors in Ragusa (Sicily). Int J Cancer,
34: 335-337.
Dargent JL, Lespagnard L, Heenan M, & Verhest A (1992) Malignant
melanoma occurring in a case of oculocutaneous albinism.
Histopathology, 21: 74-76.
Darrell RW & Bachrach CA (1963) Pterygium among veterans. Arch
Ophthalmol, 70: 158-169.
Davies DM (1985) Calcium metabolism in healthy men deprived of
sunlight. Ann N Y Acad Sci, 453: 21-27.
Daynes RA & Spellman CW (1977) Evidence for the generation of
suppressor cells by UV radiation. Cell Immunol, 31: 182-187.
Daynes RA, Spellman CW, Woodward JG, & Stewart DA (1977) Studies
into the transplantation biology of ultraviolet light-induced
tumours. Transplantation, 23: 343-348.
De Fabo EC & Kripke ML (1979) Dose-response characteristics of
immunologic unresponsiveness to UV-induced tumors produced by UV
irradiation of mice. Photochem Photobiol, 30: 385-390.
De Fabo EC & Noonan FP (1983) Mechanism of immune suppression by
ultraviolet irradiation in vivo. J Exp Med, 157: 84-98.
de Gruijl FR & van der Leun JC (1980) A dose-response model for skin
cancer induction by chronic UV exposure of a human population. J
Theor Biol, 83: 487-504.
de Gruijl FR & van der Leun JC (1982) Systemic influence of
pre-irradiation of a limited skin area on UV-tumorigenesis.
Photochem Photobiol, 35: 379-383.
de Gruijl FR & van der Leun JC (1983) Follow up on systemic
influence of partial pre-irradiation on UV-tumorigenesis. Photochem
Photobiol, 38: 381-383.
de Gruijl FR & van der Leun JC (1991) Action spectra for
carcinogenesis. In: Urbach F ed. Biological responses to UVA.
Overland Park, Kansas, Valdemar Publishing Company, p 91.
de Gruijl FR, van der Meer JB, & van der Leun JC (1983) Dose-time
dependency of tumor formation by chronic UV exposure. Photochem
Photobiol, 37: 53-62.
de Gruijl FR, Sterenborg HJCM, Forbes PD, Davies RE, Cole C,
Kelfkens G, van Weelden H, Slaper H, & van der Leun JC (1993)
Wavelength dependence of skin cancer induction by ultraviolet
radiation of albino hairless mice. Cancer Res, 53: 53-60.
Denkins Y, Fidler IJ, & Kripke ML (1989) Exposure of mice to UV-B
radiation suppresses delayed hypersensitivity to Candida albicans.
Photochem Photobiol, 49: 615-619.
Desai ID, Sawant PL, & Tappel AL (1964) Peroxidative and radiation
damage to isolated lysosomes. Biochim Biophys Acta, 86: 277-285.
Detels R & Dhir SP (1967) Pterygium: a geographical study. Arch
Ophthalmol, 78: 485-491.
Devary Y, Gottlieb RA, Smeal T & Karin M (1992) The mammalian
ultraviolet response is triggered by activation of Src tyrosine
kinases. Cell 71: 1081-1091.
Devary Y, Rosette C, Di Donato JA, & Karin M (1993) NFkB activation
by ultraviolet light not dependent on a nuclear signal. Science,
261: 1442-1445.
Dhir SP, Detels R, & Alexander ER (1967) The role of environmental
factors in cataract, pterygium and trachoma. Am J Ophthalmol, 64:
128-135.
Diffey BL (1977) The calculation of the spectral distribution of
natural ultraviolet radiation under clear sky conditions, Phys Med
Biol, 22: 309-316.
Diffey BL (1986) Use of UVA sunbeds for cosmetic tanning. Br J
Dermatol, 225: 67-76.
Diffey BL (1987) Analysis of the risk of skin cancer from sunlight
and solaria in subjects living in northern Europe. Photodermatology,
4: 118-126.
Diffey BL (1988) The risk of skin cancer from occupational exposure
to ultraviolet radiation in hospitals. Phys Med Biol, 33(10):
1187-1193.
Diffey BL (1989) Ultraviolet radiation and skin cancer: Are
physiotherapists at risk? Physiotherapy, 75(10): 615-616.
Diffey BL (1989a) Ultraviolet radiation dosimetry with polysulphone
film. In: Diffey BL ed. Radiation measurement in photobiology. New
York, London, San Francisco, Academic Press, pp 135-159.
Diffey BL (1990) Human exposure to ultraviolet radiation. Semin
Dermatol, 9: 2-10.
Diffey BL (1993) A photobiological evaluation of lamps used in the
phototherapy of seasonal affective disorder. J Photochem Photobiol,
B17: 203-207.
Diffey BL (1993a) Personal communication.
Diffey BL & Cheeseman J (1992) Sun protection with hats. Br J
Dermatol, 127: 10-12.
Diffey BL & McKinlay AF (1983) The UVB content of 'UVA fluorescent
lamps' and its erythemal effectiveness in human skin. Phys Med Biol,
28: 351-358.
Diffey BL & Robson J (1992) The influence of pigmentation and
illumination on the perception of erythema. Photodermatol
Photoimmunol Photomed, 9: 45-47.
Diffey BL, Tate TJ, & Davis A (1979) Solar dosimetry of the face:
the relationship of natural ultraviolet exposure to basal cell
carcinoma localisation. Phys Med Biol, 24: 931-939.
Diffey BL, Challoner AVJ, & Key PJ (1980) A survey of the
ultraviolet radiation emissions of photochemotherapy units. Br J
Dermatol, 102: 301-306.
Diffey BL, Larko O, & Swanbeck G (1982) UVB doses received during
different outdoor activities and UVB treatment of psoriasis. Br J
Dermatol, 106: 33-41.
Dillon J, Wang RH, & Atherton S (1990) Photochemical and
photophysical studies on human lens constituent. Photochem
Photobiol, 52: 849-854.
Dion M & Hammelin C (1987) Relationship between enhanced
reactivation and mutagenesis of UV-irradiated human cytomegalovirus
in normal human cells. EMBO J, 6: 397-399.
Djavuheri-Mergny M, Maziére JC, Santus R, Mora L, Maziére C, Auclair
M & Dubertet C (1993) Exposure to long wavelength ultraviolet
radiation decreases processing of low density lipoprotein by
cultured human fibroblasts. Photochem Photobiol 57: 302-305.
Döhler G (1990) Effect of UVB (290-320nm) radiation on uptake of
15N-nitrate by marine diatoms. Berlin, Heidelberg, New York,
Springer-Verlag, pp 354-359.
Döhler G & Alt MR (1989) Assimilation of 15N-ammonia during
irradiance with ultraviolet-B and monochromatic light by
Thalassiosira rotula, C R Acad Sci Paris, D308, 513-518.
Dolezal JM, Perkins ES, & Wallace RB (1989) Sunlight, skin
sensitivity, and senile cataract. Am J Epidemiol, 129: 559-568.
Doll R (1991) Urban and rural factors in the aetiology of cancer.
Int J Cancer, 47: 803-810.
Doll R, Muir C, & Waterhouse J ed. (1970) Cancer incidence in five
continents, Volume II. Berlin, Heidelberg, New York,
Springer-Verlag.
Doniger J, Jacobson ED, Krell K, & DiPaolo JA (1981) Ultraviolet
light action spectra for neoplastic transformation and lethality of
syrian hamster embryo cells correlate with spectrum for pyrimidine
dimer formation in cellular DNA. Proc Natl Acad Sci (USA), 78:
2378-2383.
Dorn HF (1944b) Illness from cancer in the United States: IV.
Illness from cancer of specific sites classed in broad groups.
Public Health Rep, 59: 65-77.
Dorn HF (1944a) Illness from cancer in the United States. Public
Health Rep, 59: 33-48.
Dorn CR, Taylor DON, & Schneider R (1971) Sunlight exposure and risk
of developing cutaneous and oral squamous cell carcinomas in white
cats. J Natl Cancer Inst, 46: 1073-1078.
Doughty MJ & Cullen A (1990) Long-term effects of a single dose of
ultraviolet B on albino rabbit cornea. II. Deturgescence and fluid
pump assessed in vitro. Photochem Photobiol, 54: 439-449.
Driscoll CMH (1992) Solar UV trends and distributions. Natl Radiat
Prot Board Bull, 137: 7-13.
Dubin N, Moseson M, & Pasternack BS (1986) Epidemiology of malignant
melanoma: pigmentary traits, ultraviolet radiation, and the
identification of high-risk populations. Recent Results Cancer Res,
102: 56-75.
Duke-Elder WS (1926) The pathological action of light upon the eye.
I. Action on the outer eye: Photophthalmia. Lancet, 1: 1137-1141.
Duvic M, Lowe L, Rapini RP, Rodriguez S, & Levy ML (1989) Eruptive
dysplastic nevi associated with human immunodeficiency virus
infection. Arch Dermatol, 125: 397-401.
Dyall-Smith D & Varigos G (1985) The malignant potential of
papillomavirus. Aust J Dermatol, 26: 102-107.
Eggersdorfer B & Häder DP (1991) Phototaxis, gravitaxis and vertical
migration in the marine dinoflaggelates. Acta Protozool 30(2):
63-71.
Eisenstark A & Perrot G (1987) Catalase has only a minor role in
protection against ultraviolet radiation damage in bacteria. Mol Gen
Genet 207: 68-72.
Eisman JA, MacIntyre I, Martin TJ, Frampton RJ, & King RJB (1980)
Normal and malignant breast tissue is a target organ for 1,25-(OH)2
vitamin D3. Clin Endocrinol, 13: 267-272.
Elliott R (1961) The aetiology of pterygium. Trans Ophthalmol Soc N
Z, 13: 22-41.
Ellison MJ & Childs JD (1981) Pyrimidine dimers induced in
Escherichia coli DNA by ultraviolet radiation present in sunlight.
Photochem Photobiol, 34: 465-469.
Elmets CA, Bergstresser PR, Tigelaar RE, Wood PJ, & Streilein JW
(1983) Analysis of the mechanism of unresponsiveness produced by
haptens painted on skin exposed to low dose ultraviolet radiation. J
Exp Med, 158: 781-794.
Elwood, JM & Gallagher RP (1983) Site distribution of malignant
melanoma. Can Med Assoc J, 128: 1400-1404.
Elwood JM, Gallagher RP, Hill GB, Spinelli JJ, Pearson JCG, &
Threlfall W (1984) Pigmentation and skin reaction to sun as risk
factors for cutaneous melanoma: Western Canada melanoma study. Br
Med J, 288: 99-102.
Elwood JM, Gallagher RP, Davison J, & Hill GB (1985a) Sunburn,
suntan and the risk of cutaneous malignant melanoma: the Western
Canada melanoma study. Br J Cancer, 51: 543-549.
Elwood JM, Gallagher RP, Hill GB, & Pearson JCG (1985b) Cutaneous
melanoma in relation to intermittent and constant sun exposure: the
Western Canada melanoma study. Int J Cancer, 35: 427-433.
Elwood JM, Williamson C, & Stapleton PJ (1986) Malignant melanoma in
relation to moles, pigmentation, and exposure to fluorescent and
other lighting sources. Br J Cancer, 53: 65-74.
Elwood JM, Whitehead SM, Davison J, Stewart M, & Galt M (1990)
Malignant melanoma in England: risk associated with naevi, freckles,
social class, hair colour, and sunburn. Int J Epidemiol, 19:
801-810.
Emmett EA (1973) Ultraviolet radiation as a cause of skin tumors.
Crit Rev Toxicol, 2, 211-255.
Engel A, Johnson M, & Haynes S (1988) Health effects of sunlight
exposure in the United States. Arch Dermatol, 124: 72-79.
English DR & Armstrong BK (in press) Cutaneous malignant melanoma.
In: Schottenfeld, D Fraumeni. J eds, Cancer epidemiology and
prevention, New York, Oxford University Press.
Enninga IC, Groenendijk RTL, Filon AR, van Zeeland AA, & Simons JWIM
(1986) The wavelength dependence of UV-induced pyrimidine dimer
formation, cell killing and mutation induction in human diploid skin
fibroblasts. Carcinogenesis, 7: 1829-1836.
Epe B, Pflaum M, & Boiteux S (1993) DNA damage induced by
photosensitisers in cellular and cell-free systems. Mutat Res 299:
135-145.
Epstein JH (1965) Comparison of the carcinogenic and cocarcinogenic
effects of ultraviolet light on hairless mice. J Natl Cancer Inst,
34: 741-745.
Epstein JH (1985) Animal models for studying photocarcinogenesis.
In: Maibach H & Lowe N ed. Models in Dermatology. Basel, Karger, vol
2, pp 303-312.
Epstein JH (1988) Photocarcinogenesis promotion studies with benzoyl
peroxide (BPO) and croton oil. J Invest Dermatol, 91: 114-116.
Epstein JH & Epstein WL (1962) Cocarcinogenic effect of ultraviolet
light on DMBA tumor initiation in albino mice. J Invest Dermatol,
39: 455-460.
Epstein JH & Roth HL (1968) Experimental ultraviolet light
carcinogenesis. A study of croton oil promoting effects. J Invest
Dermatol, 50: 387-389.
Epstein JH, Tuffanelli DL, & Dubois EL (1965) Light sensitivity and
lupus erythematosus. Arch Dermatol, 91: 483-485.
Epstein JH, Epstein WL, & Nakai T (1967) Production of melanomas
from DMBA-induced 'blue nevi' in hairless mice with ultraviolet
light. J Natl Cancer Inst, 38: 19-30.
Eriksen P (1987) Occupational applications of ultraviolet radiation:
risk evaluation and protection techniques. In: Passchier WF &
Bosnjakovic BFM ed. Human exposure to ultraviolet radiation: Risks
and regulations. Amsterdam, Oxford, New York, Elsevier Science
Publishers, pp 317-331.
Everett MA, Olsen RL, & Sayer RM (1965) Ultraviolet erythema. Arch
Dermatol, 92: 713-719.
Farr PM, Marks JM, Diffey BL, & Ince P (1988) Skin fragility and
blistering due to the use of sunbeds. Br Med J, 296: 1708-1709.
FDA (1988) Quality control guide for sunlamp products, Washington,
DC, US Dept. Health and Human Services, (HHS Publication FDA No.
88-8234).
FDA (1992) Medications that increase sensitivity to light (prepared
by Levine JI). Rockville, Maryland, Food and Drug Administration,
Center of Devices and Radiological Health.
Findlay GM (1928) Ultra-violet light and skin cancer. Lancet, ii:
1070-1073.
Finsen NR (1901) The treatment of lupus vulgaris by concentrated
chemical rays. In: Phototherapy. London, Edward Armold, pp 73-75.
Fisher GJ & Johns HE (1976) Pyrimidine photohydrates. In: (Ed. Wang,
S.) Photochemistry and Photobiology of Nucleic Acids, vol. 1, New
York, Academic Press, pp 169-224.
Fisher MS & Kripke ML (1977) Systemic alteration induced in mice by
ultraviolet light irradiation and its relationship to ultraviolet
carcinogenesis. Proc Natl Acad Sci (USA), 74: 1688-1692.
Fisher MS, Menter JM, & Willis I (1989) Ultraviolet
radiation-induced suppression of contact hypersensitivity in
relation to padimate O and oxybenzone. J Invest Dermatol, 92:
337-341
Fitzpatrick TB & Sober AJ (1985) Sunlight and skin cancer. N Engl J
Med, 313: 818-820.
Fitzpatrick TB, Pathak MA, Magnus IA, & Curwen WL (1963) Abnormal
reactions of man to light. Annu Rev Med, 14: 195-214.
Fitzpatrick TB, Pathak MA, Harber LC, Seiji M, & Kukita A (1974)
Sunlight and man. Tokyo, University of Tokyo Press.
Fleming ID, Barnawell JR, Burlison PE, (1975) Skin cancer in black
patients. Cancer, 35: 600-605.
Forbes PD, Davies RE, Sambuco CP, & Urbach F (1989) Topical urocanic
acid enhances UV-induced skin tumors in hairless mice by topical
application of the sunscreen 2-ethylhexyl-p-methoxycinnamate. J
Toxicol Cutan Ocul Toxicol, 8: 209-226.
Forbes PD, Blum HF, & Davies RE (1981) Photocarcinogenesis in
hairless mice: dose-response and the influence of dose-delivery.
Photochem Photobiol, 34: 361-365.
Forbes PD, Davies RE, Urbach F, Berger D, & Cole C (1982) Simulated
stratospheric ozone depletion and increased ultraviolet radiation:
effects on photocarcinogenesis in hairless mice. Cancer Res, 42:
2796-2803.
Forbes PD, Davies RE, & Urbach F (1978) Experimental ultraviolet
photocarcinogenesis: wavelength interactions and time-dose
relationships. Natl Cancer Inst Monogr, 50: 31-38.
Forsius H (1988) Exfoliation syndrome in various ethnic populations.
Acta Ophthalmol, 68(Suppl 184): 71-85.
Foster HM & Webb SJ (1988) Skin cancer in the North Solomons. Aust N
Z J Surg, 58: 397-401.
Fragu P, Lemarchand-Venencie F, Benhamou S, Francois P, Jeannel D,
Benhamou E, Sezary-Lartigau I, & Avril MF (1991) Long-term effects
in skin and thyroid after radiotherapy for skin angiomas - A French
retrospective cohort study. Eur J Cancer, 27: 1215-1222.
Franklin WA, Lo KM, & Haseltine WA (1982) Alkaline lability of
fluorescent photoproducts produced in ultraviolet light-irradiated
DNA. J Biol Chem, 157: 13535-13543.
Frederick JE, Snell HE, & Haywoood EK (1989) Solar ultraviolet
radiation at the earth's surface. Photochem Photobiol, 51: 443-450.
Freeman RG (1975) Data on the action spectrum for ultraviolet
carcinogenesis. J Natl Cancer Inst, 55: 1119-1122.
Freeman SE, Hacham H, Gange RW, Maytum DJ, Sutherland JC, &
Sutherland BM (1989) Wavelength dependence of pyrimidine dimer
formation in DNA of human skin irradiated in situ with ultraviolet
light. Proc Natl Acad Sci (USA), 86: 5605-5609.
Friedberg EC (1984) DNA repair. New York, Freeman and Company.
Friedmann PS, White SI, Parker S, Moss C, & Matthews JNS (1989)
Antigenic stimulation during ultraviolet therapy in man does not
result in immunological tolerance. Clin Exp Immunol, 76: 68-72.
Fuchs J & Packer L (1991) Photooxidative stress in the skin. In:
Sies H Ed. Oxidative stress: oxidants and antioxidants. New York,
London, San Francisco, Academic Press, pp 554-584.
Fuchs J, Huflejt M, Rothfuss L, Carcamero G, & Packer L (1989)
Impairment of enzymic and nonenzymic antioxidants in skin by UVB
irradiation. J Invest Dermatol 93, 769-773.
Fuller CJ, Faulkner H, Bendich A, Parker RS, & Roe DA (1992) Effect
of ß-carotene supplementation on photosuppression of delayed-type
hypersensitivity in normal young men. Am J Clin Nutr, 56: 684-690.
Funnell SGP & Keast D (1985) The effect of ultraviolet radiation on
the generation of plaque-forming cells and on T-suppressor cell
activity to sheep erythrocytes. Photodermatology, 3: 64-72.
Gaboriau F, Morlière P, Marquis I, Maysan A, Gèze M, & Dubertret L
(1993) Membrane damage induced in cultured human skin fibroblasts by
UVA irradiation. Photochem Photobiol, 58: 515-520.
Gafa L, Filippazzo MG, Tumino R, Dardanoni G, Lanzarone F, &
Dardanoni L (1991) Risk factors of nonmelanoma skin cancer in
Ragusa, Sicily: a case-control study. Cancer Causes Control, 2:
395-399.
Gallagher, RP (1988) Ocular melanoma in farmers (Letter to the
Editor). Am J Ind Med, 13: 523-525.
Gallagher PE & Duker NJ (1986) Detection of UV purine photoproduct
in a defined sequence of human DNA. Mol Cell Biol, 6: 707-709.
Gallagher RP, Elwood JM, & Hill GB (1986) Risk factors for cutaneous
malignant melanoma: the Western Canadian melanoma study, Rec Res
Cancer Res, 102: 38-55.
Gallagher RP, Threlfall WJ, Jeffries E, Band PR, Spinelli J, &
Coldman AJ (1984) Cancer and aplastic anemia in British Columbia
farmers. J Natl Cancer Inst, 72: 1311-1315.
Gallagher CH, Greenoak GE, Reeve VE, Canfield PJ, Baker RSU, & Bonin
AM (1984a) Ultraviolet carcinogenesis in the hairless mouse skin -
influence of the sunscreen 2-ethylhexyl-p-methoxycinnamate. Aust J
Exp Biol Med Sci, 62: 577-588.
Gallagher RP, Elwood JM, Rootman J, Spinelli JJ, Hill GB, Threlfall
WJ, & Birdsell JM (1985) Risk factors for ocular melanoma: Western
Canada melanoma study. J Natl Cancer Inst, 74: 775-778.
Gallagher PE, Weiss RB, Brent TP, & Duker NJ (1989) Wavelength
dependence of DNA incision by a human ultraviolet endonuclease.
Photochem Photobiol, 49: 363-367.
Gallagher RP, Ma B, McLean DI, Yang CP, Ho V, Carruthers JA, &
Warshawski LM (1990a) Trends in basal cell carcinoma, squamous cell
carcinoma, and melanoma of the skin from 1973 through 1987. J Am
Acad Dermatol, 23: 413-421.
Gallagher RP, McLean DI, Yang CP, Coldman AJ, Silver HKB, Spinelli
JJ, & Beagrie M (1990b) Anatomic distribution of acquired
melanocytic nevi in white children. A comparison with melanoma: the
Vancouver mole study. Arch Dermatol, 126: 466-471
Gallagher RP, McLean DI, Yang CP, Coldman AJ, Silver HKB, Spinelli
JJ, & Beagrie M (1990c) Suntan, sunburn and pigmentation factors and
the frequency of acquired melanocytic nevi in children. Similarities
to melanoma: the Vancouver mole study. Arch Dermatol, 126: 770-776
Gallagher RP, Rivers JK, Yang CP, (1991) Melanocytic nevus density
in Asian, Indo-Pakistani, and white children: The Vancouver mole
study. J Am Acad Dermatol, 25: 507-12.
Gange RW, Blackett AD, Matzinger EA, Sutherland BM, & Kochevar IE
(1985) Comparative protection efficiency of UVA-and UVB-induced tans
against erythema and formation of endonuclease-sensitive sites in
DNA by UVB in human skin. J Invest Dermatol, 85: 362-364.
Garbe C, Krüger S, Stadler R, Guggenmoos-Holzmann I, & Orfanos CE
(1989) Markers and relative risk in a German population for
developing malignant melanoma. Int J Dermatol, 28: 517-523.
Garcia-Pichel F & Castenholz RW (1991) Characterization and
biological implications of scytonemin, a cyanobacterial sheath
pigment, J Phycol, 27: 395-409.
Garland C & Garland F (1980) Do sunlight and vitamin D reduce the
likelihood of colon cancer? Int J Epidemiol, 9: 227-231.
Garland CF, Comstock GW, Garland FC, Felsing K, Shaw EK, & Gorham ED
(1989) Serum 25-hydroxyvitamin D and colon cancer: 8-year
prospective study. Lancet, 2: 1176-1178.
Garland FC, Garland CF, Gorham ED, & Young JF (1990) Geographic
variation in breast cancer mortality in the United States: A
hypothesis involving exposure to solar radiation. Prev Med, 19:
614-622.
Garland CF Garland FC, & Gorham ED (1992) Could sunscreens increase
melanoma risk? (Letter) Am J Public Health, 82(4): 614-615.
Garner A (1989) The pathology of tumours at the limbus. Eye, 3:
210-217.
Garsen J, Goettsch W, de Gruy F, & van Loveren H (1993) UVB
suppresses immunity and resistance against systemic infections in
the rat. Photochem Photobiol, 57: 755.
Gasparro F & Fresco J (1986) Ultraviolet-induced 8,8-adenine
dehydromimers in oligo- and polynucleotides. Nucl Acids Res, 14:
4239-4251.
Gellin GA, Kopf AW, & Garfinkel L (1965) Basal cell epithelioma. A
controlled study of associated factors. Arch Dermatol, 91: 38-45.
Gensler HL (1988) Enhancement of chemical carcinogenesis in mice by
systemic effects of ultraviolet radiation. Cancer Res, 48: 620-623.
Gensler HL (1989) Reduction of immunosuppression in UV-irradiated
mice by dietary retinyl palmitate plus canthaxanthin.
Carcinogenesis, 10: 203-207.
Gensler HL & Bowden GT (1987) UVB-Induced modulation of mouse skin
tumor induction by benzo[a]pyrene (Abstract no. 546). Proc Am Assoc
Cancer Res, 28: 137.
Gensler HL & Welch K (1992) Prevalence of tumor prevention rather
than tumor enhancement when repetitive UV radiation treatments
precede initiation and promotion. Carcinogenesis, 13: 9-13.
Gerrish KE & Gensler HL (1993) Prevention of photocarcinogenesis by
dietary vitamin E. Nutr Cancer, 19: 125-133.
Giannini SH (1986a) Suppression of pathogenesis in cutaneous
leishmaniasis by UV irradiation. Infect Immun, 51: 838-843.
Giannini SH (1986b) Effects of UV-B on infectious disease. In: Titus
JG ed. Effects of changes in stratospheric ozone and global climate.
Washington, DC, US Environmental Protection Agency, vol 2, pp
101-112.
Giannini SH (1987) Abrogation of skin lesions in cutaneous
leishmaniasis by ultraviolet irradiation. In: Hart DT ed.
Leishmaniasis: The first centenary (1885-1985) new strategies for
control. New York, London, Plenum Press, pp 677-684 (NATO ASI Series
A: Life Sciences).
Giannini SH (1992) Effects of ultraviolet B irradiation on cutaneous
leishmaniasis. Parasitology Today, 8: 44-48.
Gibbs NK (1993) Is ultraviolet immunosuppression initiated by
photosomerisation of urocanic acid. In: de Gruijl FR ed. The dark
side of sunlight. Utrecht, The Netherlands, Utrecht University.
Gies HP & Roy CR (1990) Bilirubin phototherapy and potential UVR
hazards. Health Phys, 58: 313-320.
Gies HP, Roy CR, & Elliott G (1986) Artificial suntanning: Spectral
irradiance and hazard evaluation of ultraviolet sources. Health
Phys, 50(6): 691-703.
Gies HP, Roy CR, & Elliot G (1990) A proposed UVR protection factor
for sunglasses. Clin Exp Optom, 73: 183-189.
Gies HP, Roy CR, & Elliott G (1992) Ultraviolet radiation protection
factors for personal protection in both occupational and
recreational situations. Radiat Prot Aust, 10(3): 59-66.
Gies HP, Roy CR, Herlihy E, & Rivers J (1992a) Personal dosimetry of
solar UVB using polysulphone film Congress Proceedings (IRPA 8) vol
1, p 791, Montreal, May 1992, International Radiation Protection
Association.
Gilchrest BA (1990) Actinic injury. Annu Rev Med, 41: 199-210.
Gilchrest BA, Soter NA, Stoff JS, & Mihm MC Jr (1981) The human
sunburn reaction: histologic and biochemical studies. J Am Acad
Dermatol, 5: 411-422.
Giles GG, Marks R, & Foley P (1988) Incidence of non- melanocytic
skin cancer treated in Australia. Br Med J, 296: 13-17.
Glass AG & Hoover RN (1989) The emerging epidemic of melanoma and
squamous cell skin cancer. J Am Med Assoc, 262: 2097-2100.
Goldberg LH & Altman A (1984) Benign skin changes associated with
chronic sunlight exposure. Cutis, 34: 33-39.
Goodman GJ, Marks R, Selwood TS, Ponsford MW, & Pakes W (1984)
Non-melanocytic skin cancer and solar keratoses in Victoria -
clinical studies II. Aust J Dermatol, 25: 103-106.
Gorham ED, Garland CF, & Garland FC (1989) Acid haze air pollution
and breast and colon cancer mortality in 20 Canadian cities. Can J
Public Health, 80: 96-100.
Gorham ED, Garland FC, & Garland CF (1990) Sunlight and breast
cancer incidence in the USSR. Int J Epidemiol, 19: 820-824.
Graham S, Marshall J, Haughey B, Stoll H, Zielezny M, Brasure J, &
West D (1985) An inquiry into the epidemiology of melanoma. Am J
Epidemiol, 122: 606-619.
Gray RH, Johnson GJ, & Freedman A (1992) Climatic droplet
keratopathy. Surv Ophthalmol, 36: 241-253.
Greaves MW, Hensby CN, Black AK, Plummer NA, Fincham N, Warin AP, &
Camp R (1978) Inflammatory reactions induced by ultraviolet
irradiation. Bull. Cancer, 65: 299-304.
Green AC (1984) Sun exposure and the risk of melanoma. Aust J
Dermatol, 25: 99-102.
Green AC (1991) Premature ageing of the skin in a Queensland
population. Med J Aust, 155: 473-478.
Green AC & Battistutta D (1990) Incidence and determinants of skin
cancer in a high-risk Australian population. Int J Cancer, 46:
356-361.
Green A & O'Rourke MGE (1985) Cutaneous malignant melanoma in
association with other skin cancers. J Natl Cancer Inst, 74:
977-980.
Green A & Siskind V (1983) Geographical distribution of cutaneous
melanoma in Queensland. Med J Aust, 1: 407-410.
Green AES, Sawada T, & Shettle EP (1974) The middle ultraviolet
reaching the ground. Photochem Photobiol, 19: 251.
Green AC, Siskind V, Bain C, & Alexander J (1985) Sunburn and
malignant melanoma. Br J Cancer, 51: 393-397.
Green AC, Beardmore G, Hart V, Leslie D, Marks R, & Staines D
(1988a) Skin cancer in a Queensland population. J Am Acad Dermatol,
19: 1045-1052.
Green AC, Sorahan T, Pope D, Siskind V, Hansen M, Hanson L, Leech P,
Ball PM, & Grimley RP (1988b) Moles in Australian and British
schoolchildren (Letter to the Editor). Lancet, ii: 1497.
Green C, Diffey BL, & Hawk JLM (1992) Ultraviolet radiation in the
treatment of skin disease. Phys Med Biol, 37(1): 1-20.
Green A, Smith P, McWhirter W, Oregan P, Battistutta D, Yarker ME, &
Lape K (1993a) Melanocytic naevi and melanoma in survivors of
childhood cancer. Br J Cancer, 67: 1053-1057.
Green A, MacLennan R, Youll P, & Martin N (1993b) Site distribution
of cutaneous melanoma in Queensland. Int J Cancer, 53: 232-236.
Greenberg JT, Monach PA, Chou JH, Josephy PD, & Demple B (1990)
Positive control of a global antioxidant defence regulon activated
by superoxide-generating agents in Escherichia coli. Proc Natl Acad
Sci (USA), 87: 6181-6185.
Greene MH & Wilson J (1985) Second cancer following lymphatic and
hematopoietic cancers in Connecticut, 1935-82. Natl Cancer Inst
Monogr, 68: 191-217.
Greene MH, Young TI, & Clark WH Jr (1981) Malignant melanoma in
renal transplant recipients. Lancet, May 30: 1196-1199.
Grob JJ, Gouvernet J, Aymar D, Mostaque A, Romano MH, Collet AM, Noe
MC, Diconstanzo MP, & Bonerandi JJ (1990) Count of benign
melanocytic nevi as a major indicator of risk for nonfamilial
nodular and superficial spreading melanoma. Cancer, 66: 387-395.
Guerry RK, Ham WT, & Mueller HA (1985) Light toxicity in the
posterior segment. In: Duane TD & Jaeger EA ed. Clinical
ophthalmology. Philadelphia, Pennsylvania, Harper and Row, pp 1-17.
Guex-Crosier Y & Herbort CP (1993) Presumed corneal intraepithelial
neoplasia associated with contact lens wear and intense ultraviolet
light exposure. Br J Ophthalmol, 77: 191-192.
Gupta AK, Cardella CJ, & Haberman HF (1986) Cutaneous malignant
neoplasms in patients with renal transplants. Arch Dermatol, 122:
1288-1293.
Gupta AK, Stern RS, Swanson NA, Anderson TF, & Arbor A (1988)
Cutaneous melanomas in patients treated with psoralens plus
ultraviolet A. J Am Acad Dermatol, 19: 67-76.
Gutteridge JMC (1985) Superoxide dismutase inhibits the
superoxide-driven Fenton reaction at two different levels. FEBS
Lett, 185: 19-23.
Häder DP & Worrest RC (1991) Effects of enhanced solar radiation on
aquatic ecosystems. Photochem Photobiol, 53: 717-725.
Häder DP & Lui SM (1991) Mobility and gravitactic orientation of the
flagellate, Euglena gracilis, impaired by artificial and solar UVB
radiation. Curr Microbiol, 21: 161-168.
Haenszel W (1963) Variations in skin cancer incidence within the
United States. Natl Cancer Inst Monogr, 10: 225-243.
Halprin KM, Comerford M, Presser SE, & Taylor JR (1981) Ultraviolet
light treatment delays contact sensitization to nitrogen mustard. Br
J Dermatol, 105: 71-76.
Ham WT, Mueller HA, Ruffolo JJ, Millen JE, Cleary SF, & Guerry RK
(1984) Basic mechanisms underlying the production of photochemical
lesions in the mammalian retina. Curr Eye Res, 3: 165-174.
Hanchette CL & Schwartz GC (1992) Geographic patterns of
prostatecancer mortality. Evidence for a protective effect of
ultraviolet radiation. Cancer, 70: 2861-2869.
Hardie IR, Strong RW, Hartley LCJ, Woodruff PWH, & Clunie GJA (1980)
Skin cancer in Caucasian renal allograft recipients living in a
subtropical climate. Surgery, 87: 177-183.
Hartevelt MM, Bouwes Bavinck JN, Kootte AMM, Vermeer BJ, &
Vandenbroucke JP (1990) Incidence of skin cancer after renal
transplantation in the Netherlands. Transplantation, 49: 506-509.
Hawk, JLM (1984) Photosensitising agents used in the United Kingdom.
Clin Expl Dermatol, 9: 300-302.
Hayashi Y & Aurelian L (1986) Immunity to herpes simplex virus type
2: Viral antigen-presenting capacity of epidermal cells and its
impairment by ultraviolet irradiation. J Immunol, 136: 1087-1092.
Haynes RH (1966) The interpretation of microbiol inactivation and
recovery phenomena. Radiat Res, Suppl 6: 1-29.
Health Council of the Netherlands (1986) UV radiation: Human
exposure to ultraviolet radiation. The Hague, Health Council of the
Netherlands (Report 1986/93).
Hedblom EE (1961) Snowscape eye protection. Arch Environ Health, 2;
685-704.
Hefferren JJ, Cooley RO, Hall JB, Olsen NH, & Lyon HW (1971) Use of
ultraviolet illumination in diagnosis. J Am Dent Assoc, 82:
1353-1360.
Herity B, O'Loughlin G, Moriarty MJ, & Conroy R (1989) Risk factors
for non-melanoma skin cancer. Ir Med J, 82: 151-152.
Hersey P, Hasic E, Edwards A, Bradley M, Maran G, & McCarthy WH
(1983a) Immunological effects of solarium exposure. Lancet, 1:
545-548.
Hersey P, Haran G, Hasic E, & Edwards A (1983b) Alteration of T cell
subsets and induction of suppressor T cell activity in normal
subjects after exposure to sunlight. J Immunol, 31: 171-174.
Hersey P, MacDonald M, Burns C, Schibeci S, Matthews H, & Wilkinson
FJ (1987) Analysis of the effect of sunscreen agent on the
suppression of natural killer cell activity induced in human
subjects by radiation from solarium lamps. J Invest Dermatol, 88:
271-276.
Hersey P, MacDonald M, Henderson C, Schibeci S, D'Alessandro G,
Pryor M, & Wilkinson FJ (1988) Suppression of natural killer cell
activity in humans by radiation from solarium lamps depleted of UVB.
J Invest Dermatol, 90: 305-310.
Hess C (1907) Experiments on the effect of ultraviolet light on the
lens. Arch f Augenheilk 57 185-196.
Higginson J & Oettlé AG (1959) Cancer incidence in the Bantu and
"Cape Colored" races of South Africa: Report of a cancer survey in
the Transvaal (1953-55). J Natl Cancer Inst, 24: 589-671.
Hill D, White V, Marks R, Theobald T, Borland R, & Roy C (1992)
Melanoma prevention: behavioural and nonbehavioural factors in
sunburn among an Australian urban population. Prev Med, 21: 654-669.
Hiller R, Sperduto RD, & Ederer F (1977a) Epidemiological
associations with cataract in the 1971-72 national health and
nutrition examination survey. Am J Epidemiol, 105: 450-459.
Hiller R, Giacometti L, & Yuen K (1977b) Sunlight and cataract: an
epidemiologic investigation. Am J Epidemiol, 105: 450-459.
Hiller R, Sperduto RD, & Ederer F (1983) Epidemiologic associations
with cataract in the 1971-1972 national health and nutrition
examination survey. Am J Epidemiol, 118: 239-249.
Hiller R, Sperduto RD, & Ederer F (1986) Epidemiologic associations
with nuclear, cortical, and posterior subcapsular cataracts. Am J
Epidemiol, 124: 916-925.
Hirschfeld S, Levine AS, Ozato K, & Protic M (1990) A constitutive
damage-specific DNA binding protein is synthesized at higher levels
in UV-irradiated primate cells. Mol Cell Biol, 10: 2041-2048.
Hoeijmakers JHJ & Bootsma D (1990) Molecular genetics of eucaryotic
DNA excision repair. Cancer Cells, 2: 311-320.
Hoffman JS (1987) Ultraviolet radiation and melanoma (with a special
focus on assessing the risks of stratospheric ozone depletion).
Washington, DC, US Environmental Protection Agency, Office of Air
and Radiation (EPA 400/1-87-001D).
Hogan DJ, To T, Gran L, Wong D, & Lane PR (1991) Risk factors for
basal cell carcinoma. Int J Dermatol, 28: 591-594.
Holick MF (1985) The photobiology of vitamin D and its consequences
for human. Ann N Y Acad Sci, 453: 1-3.
Hollows F & Moran D (1981) Cataract - the ultraviolet risk factor.
Lancet, 2: 1249-1250.
Holly EA, Kelly JW, Shpall SN, & Chiu S-H (1987) Number of
melanocytic nevi as a major risk factor for malignant melanoma. J Am
Acad Dermatol, 17: 459-468.
Holly EA, Aston DA, Char DH, Kristiansen JJ, & Ahn DK (1990) Uveal
melanoma in relation to ultraviolet light exposure and host factors.
Cancer Res, 50: 5773-5777.
Holly EA, Aston DA, Ahn DK, Kristiansen JJ, & Char DH (1991) No
excess prior cancer in patients with uveal melanoma. Ophthalmology,
98: 608-611.
Holm-Hansen O (1990) UV radiation in Arctic waters: Effects on rates
of primary production (Appendix G). La Jolla, CA, USA, Scripps
Institution of Oceanography, pp 1-17.
Holman CDJ & Armstrong BK (1984) Pigmentary traits, ethnic origin,
benign nevi, and family history as risk factors for cutaneous
malignant melanoma. J Natl Cancer Inst, 72: 257-266.
Holman CDJ, Gibson IM, Stephenson M, & Armstrong BK (1983)
Ultraviolet irradiation of human body sites in relation to
occupation and outdoor activity: field studies using personal UVR
dosimeters. Clin Exp Dermatol, 8: 269-277.
Holman CDJ, Armstrong BK, Evans PR, Lumsden GJ, Dallimore KJ, Meehan
CJ, Beagley J, & Gibson IM (1984a) Relationship of solar keratosis
and history of skin cancer to objective measures of actinic skin
damage. Br J Dermatol, 110: 129-138.
Holman CD, Evans PR, Lumsden GJ, & Armstrong BK (1984b) The
determinants of actinic skin damage: problems of confounding among
environmental and constitutional variables. Am J Epidemiol, 120:
414-422.
Holman CDJ, Armstrong BK, & Heenan PJ (1986) Relationship of
cutaneous malignant melanoma to individual sunlight-exposure habits.
J Natl Cancer Inst, 76: 403-414.
Hoover R (1977) Effects of Rugs - Immunosuppression. In: Hiatt HH,
Watson JD, & Winsten JA ed. Origins of human cancer. Book A:
Incidence of cancer in humans. Proceedings of the Cold Spring Harbor
Conferences on Cell Proliferation. Cold Spring Harbor, New York,
Cold Spring Harbor Laboratory, vol 4.
Hoppeler Th, Hendrickson Ph, Dietrich C, & Reme (1988) Morphology
and time-course of defined photochemical lesions in the rabbit
retina. Curr Eye Res, 7(9): 849-860.
Howie S, Norval M, & Maingay J (1986) Exposure to low-dose
ultraviolet radiation suppresses delayed-type hypersensitivity to
herpes simplex virus in mice. J Invest Dermatol, 86: 125-128.
Howie SEM, Ross JA, Norval M, & Maingay JP (1986) In vivo
modulation of antigen presentation generates Ts rather than TDH in
HSV-1 infection. Immunology, 60: 419-423.
Hsu J, Forbes PD, Harber LC, & Lakow E (1975) Induction of skin
tumors in hairless mice by a single exposure to UV radiation.
Photochem Photobiol, 21: 185-188.
Hughes BR, Cunliffe WJ, & Bailey CC (1989) Excess benign melanocytic
naevi after chemotherapy for malignancy in childhood. Br Med J, 299:
88-91.
Hunter DJ, Colditz GA, Stampfer MJ, Rosner B, Willett WC, & Speizer
FE (1990) Risk factors for basal cell carcinoma in a prospective
cohort of women. Ann Epidemiol, 1: 13-23.
Hunter DJ, Colditz GA, Stampfer MJ, Rosner B, Willett WC, & Speizer
FE (1992) Diet and risk of basal cell carcinoma of the skin in a
prospective cohort of women. Ann Epidemiol, 2: 231-239.
Husain Z, Yang Q, & Biswas DK (1990) cHa-ras Proto oncogene.
Amplification and overexpression in UVB-induced mouse skin
papillomas and carcinomas. Arch Dermatol, 126: 324-330.
Husain Z, Pathak MA, Flotte T, & Wick MM (1991) Role of ultraviolet
radiation in the induction of melanocytic tumors in hairless mice
following 7,12-dimethylbenz(a)anthracene application and ultraviolet
irradiation. Cancer Res, 51: 4964-4970.
Hyman LG, Lilienfeld AM, Ferris FL, & Fine SL (1983) Senile macular
degeneration: a case-control study. Am J Epidemiol, 118: 213-227.
IARC (1993) In: Kricker A, Armstrong BK, Jones ME & Burton RD ed.
Health, solar UV radiation and environmental change. Lyon,
International Agency for Research on Cancer (IARC Technical Report
No. 13).
IARC (1992) Solar and ultraviolet radiation. Lyon, International
Agency for Research on Cancer (Monographs on the Evaluation of
Carcinogenic Risks to Humans, Volume 55).
IARC (1980) International Agency for Research on Cancer, Monographs
on the Evaluation of the Carcinogenic Risk of Chemicals to Humans,
Vol 24, Some Pharmaceutical Drugs, Lyon, 101-124.
Ignatiades L (1990) Photosynthetic capacity of the surface
microlayer during the mixing period. J Planktonic Res, 12: 851-860.
Imlay JA, Chir SM, & Linn S (1988) Science, 240: 640-642.
IRPA/INIRC (1985) International Radiation Protection Association/
International Non-Ionizing Radiation Committee. Concepts, units and
terminology for NIR protection. Health Phys, 49(6): 1329-1362.
IRPA/INIRC (1991a) International Radiation Protection Association/
International Non-Ionizing Radiation Committee. Health issues of
ultraviolet A sunbeds used for cosmetic purposes. Health Phys,
61(2): 285-288.
IRPA/INIRC (1991b) International Radiation Protection Association/
International Non-Ionizing Radiation Committee. In: Duchêne AS,
Lakey JRA, & Repacholi MH ed. IRPA guidelines on protection against
non-ionizing radiation., New York, McGraw-Hill.
Isaacson C, Selzer G, Kaye V, Greenberg M, Woodruff JD, Davies J,
Ninin D, Vetten D, & Andrew, M (1978) Cancer in the urban blacks of
South Africa. S Afr Cancer Bull, 22: 49-84.
Ishizaki K, Tsujimura T, Nakai M, Nishigori C, Sato K, Katayama S,
Kurimura O, Yoshikawa K, Imamura S, & Ikenaga M (1992) Infrequent
mutation of the ras genes in skin tumors of Xeroderma Pigmentosum
patients in Japan. Int J Cancer, 50: 382-385.
Italian-American Cataract Study Group (1991) Risk factors for
age-related cortical, nuclear, and posterior subcapsular cataracts.
Am J Epidemiol, 133: 541-553.
Ito A & Ito T (1983) Possible involvement of membrane damage in the
inactivation by broad-band near-UV radiation in Saccharomyces
cerevisiae cells. Photochem Photobiol 37: 395-401.
Iversen OH (1988) Skin tumorigenesis and carcinogenesis studies with
7,12-dimethylbenz[a]anthracene, ultraviolet light, benzoyl peroxide
(Panoxyl gel 5%) and ointment gel. Carcinogenesis, 9: 803-809.
Jagger J (1985) Solar-UV actions on living cells. New York, Praeger.
Jeevan A & Kripke ML (1990) Alteration of the immune response to
Mycobacterium bovis BCG in mice exposed chonically to low doses of
UV radiation. Cell Immunol, 130: 32-41.
Jeevan A, Evans R, Brown EL, & Kripke ML (1992a) Effect of local
ultraviolet irradiation on infections of mice with Candida albicans,
Mycobacterium bovis BCG, and Schistosoma mansoni. J Invest Dermatol,
99: 59-64.
Jeevan A, Gilliam K, Heard H, & Kripke ML (1992b) Effects of
ultraviolet radiation on the pathogenesis of Mycobacterium
lepraemurium infection in mice. Exp Dermatol, 1: 152-160.
Jeevan A, Ullrich SE, Dizon V, & Kripke ML (1992c) Supernantants
from ultraviolet-irradiated keratinocytes decrease the resistance
and delayed-type hypersensitivity response to Mycobacterium bovis
bacillus Calmette-Guerin in mice and impair the phagocytic ability
of macrophages. Photodermatol Photoimmunol Photomed, 9: 255-263.
Jeffery SW & Humphrey GH (1975) New spectrophotometric equations for
determining chlorophylls a, b, c1 and C2 in higher plants,
algae, and natural phytoplankton. Biochem Physiol Pflanz, 167:
191-194.
Jensen OM & Bolander (1980) Trends in malignant melanoma of the
skin, WHO Stat Q, 33: 2-26.
Johnson GJ (1981) Aetiology of spheroidal degeneration of the cornea
in Labrador. Br J Ophthalmol, 65: 270-283.
Johnson GJ & Overall M (1978) Histology of spheroidal degeneration
of the cornea in Labrador. Br J Ophthalmol, 62: 53-61.
Johnson GJ, Paterson GD, Green JS, & Perkins ES (1981) Ocular
conditions in a labrador community In: Harvald B & Hansen JP ed.
Circumpolar health 81. Copenhagen, Nordic Council for Arctic Medical
Research.
Johnson GJ, Minassian DC, & Franken S (1989) Alterations of the
anterior lens capsule associated with climatic keratopathy. Br J
Ophthalmol, 73: 229-234
Jones CA, Huberman E, Cunningham ML, & Peak MJ (1987a) Mutagenesis
and cytotoxicity in human epithelial cells by far- and
near-ultraviolet radiations : action spectra. Radiat Res, 110:
244-254.
Jones SK, Moseley H, & Mackie RM (1987b) UVA-induced melanocytic
lesions. Br J Dermatol, 117: 111-115.
Jones ME, Shugg D, Dwyer T, Young B, & Bonett A (1992) Interstate
differences in incidence and mortality from melanoma: a
re-examination of the latitudinal gradient. Med J Aust, 157:
373-377.
Jose JG (1986) Posterior cataract induction by UVB radiation in
albino mice. Exp Eye Res, 42: 11-20.
Jose JG & Pitts DG (1985) Wavelength dependency of cataracts in
albino mice following chronic exposure. Exp Eye Res, 41: 545-563.
Kalimo K, Loulu L, & Jansen CT (1983) Effect of a single UVB or PUVA
exposure on immediate and delayed skin hypersensitivity reactions in
humans. Correlation to erythemal response and Langerhans cell
depletion. Arch Dermatol Res, 275: 374-378.
Karai I & Horiguchi S (1984) Pterygium in welders. Br J Ophthalmol,
68: 347-349.
Karentz D & Lutze LH (1990) Evaluation of biologically harmful
ultraviolet radiation in Antarctica with a biological dosimeter
designed for aquatic environments. Limnol Oceanogr, 35: 549-561.
Karentz D, Mc Euen FS, Land MC, & Dunlap WC (1991) Survey of
microsporine-like amino acid compounds in Antarctic marine
organisms: potential protection from ultraviolet exposure. Mar Biol,
108: 157-166.
Karjalainen S, Salo H, & Teppo L (1989) Basal cell and squamous cell
carcinoma of the skin in Finland Int J Dermatol, 28: 445-450.
Kataoka H & Fujiwara Y (1991) UV damage specific protein in
xeroderma pigmentosum complementation group E. Biochem Biophys Res
Commun, 175: 1139-1143.
Katz L, Ben-Tuvia S, & Steinitz R (1982) Malignant melanoma of the
skin in Israel: effect of migration. In: Magnus K ed Trends in
cancer incidence: Causes and practical implication. Washington, New
York, Hemisphere, pp 419-426.
Kelfkens G, van Weelden H, de Gruijl FR, & van der Leun JC (1991)
Influence of dose rate in ultraviolet tumorigenesis. J Photochem
Photobiol, B10: 41-50.
Kelfkens G & van der Leun JC (1989) Skin temperature changes after
irradiation with UVB or UVC: Implications for the mechanism
underlying ultraviolet erythema. Phys Med Biol, 34: 599-608.
Keller, AZ (1970) Cellular types, survival, race, nativity,
occupations, habits and associated diseases in the pathogenesis of
lip cancers. Am J Epidemiol, 91, 486-499.
Kelly, GE, Meikle, WD, & Moore, DE (1989) Enhancement of UV-induced
skin carcinogenesis by azathioprine: role of photochemical
sensitisation. Photochem Photobiol, 49, 59-65.
Kelly GE, Meikle WD, & Sheil AGR (1987) Effects of immunosuppressive
therapy on the induction of skin tumors by ultraviolet irradiation
in hairless mice. Transplantation, 44, 429-434.
Kennedy JC, Poltier RH, & Pross DC (1990) Photodynamic therapy with
endogenous protoporphyrin. IX. Basic principles and present clinical
experience. J Photochem Photobiol, 6: 143-148.
Keyse SM (1993) The induction of gene expression in mammalian cells
by radiation. Sem Cancer Biol, 4: 119-128.
Keyse SM & Tyrrell RM (1989) Heme oxygenase is the major 32-kDa
stress protein induced in human skin fibroblasts by UVA radiation,
hydrogen peroxide and sodium arsenite. Proc Natl Acad Sci (USA), 86:
99-103.
Khlat M, Vail A, Parkin M, & Green A (1992) Mortality from melanoma
in migrants to Australia: variation by age at arrival and duration
of stay. Am J Epidemiol, 135: 1103-1113.
Kim TY, Kripke ML, & Ullrich SE (1990) Immunosuppression by factors
released from UV-irradiated epidermal cells: Selective effects on
the generation of contact and delayed hypersensitivity after
exposure to UVA or UVB radiation. J Invest Dermatol, 94: 26-32.
Kinlen LJ, Sheil AGR, Peto J, & Doll R (1979) Collaborative United
Kingdom-Australasian study of cancer in patients treated with
immunosuppressive drugs. Br Med J, 2: 1461-1466.
Klamen DK & Tuveson RW (1982) The effect of membrane fatty acid
compositions on the near-UV (300-400 nm) sensitivity of Escherichia
coli K1060. Photochem Photobiol, 35: 167-173.
Klepp O & Magnus K (1979) Some environmental and bodily
characteristics of melanoma patients. A case-control study. Int J
Cancer, 23: 482-486.
Kligman LH, Kaidbey KH, Hitchens VM, & Miller SA (1987) Long
wavelength (>340 nm) ultraviolet-A induced damage in hairless mice
is dose dependent. In: Passchier W & Bosnjakovi, BFM ed., Human
exposure to ultraviolet radiation: Risks and regulations. Amsterdam,
Oxford, New York, Elsevier Science Publishers, pp 77-81.
Knekt P, Aromaa A, Maatela J, Alfthan G, Aaran RK, Nikkari T, Hakama
M, Hakulinen T, & Teppo, L (1991) Serum micronutrients and risk of
cancers of low incidence in Finland. Am J Epidemiol, 134: 356-361.
Kollias, N, Sayre RM, Zeise L, & Cehedekel MR (1991) Photoprotection
by melanin, J Photochem Photobiol, 9: 135-160
Kopecky KE, Pugh GW Jr, Hughes DE, Booth GD, & Cheville, NF (1979)
Biological effect of ultraviolet radiation on cattle: bovine ocular
squamous cell carcinoma. Am J Vet Res, 40: 1783-1788.
Kopf AW, Lazar M, Bart RS, Dubin N, & Bromberg J (1978) Prevalence
of nevocytic nevi on lateral and medial aspects of arms. J Dermatol
Surg Oncol, 4: 153-158.
Kopf AW, Lindsay AC, Rogers GS, Friedman RJ, Rigel DS, & Levenstein
M (1985) Relationship of nevocytic nevi to sun exposure in
dysplastic nevus syndrome. J Am Acad Dermatol, 12: 656-662.
Kraemer KH, Lee MM, & Scotto J (1987) Xeroderma Pigmentosum Arch
Dermatol, 123: 241-50.
Kraemer KH, Seetharam S, Protic-Sabljic M, Brash DE, Bredberg A, &
Seidman MM (1988) Defective DNA repair and mutagenesis by dimer and
non-dimer photoproducts in xeroderma pigmentosum measured with
plasmid vectors. In: Freidberg EC & Hanawalt PC ed. Mechanisms and
consequences of DNA damage processing. New York, Alan R. Liss, pp
325-335.
Kramer GF & Ames BN (1987) Oxidative mechanisms of toxicity of
low-intensity near-UV light in Salmonella typhimurium. J Bacteriol,
164: 2259-2266.
Kremers JJM & van Norren D (1988) Two classes of photochemical
damage to the retina. In. Lasers and light in ophthalmology.
Amsterdam, Berkeley, Milano, Klugler Publications, vpl 2, pp 41-52.
Kress S, Sutter C, Strickland PT, Mukhtar H, Schweizer J, & Schwarz
M (1992) Carcinogen-specific mutational pattern in the p53 gene in
ultraviolet B radiation-induced squamous cell carcinomas of mouse
skin. Cancer Res, 52: 6400-6403.
Kricker A, Armstrong BK, English DR, & Heenan PJ (1991a) Pigmentary
and cutaneous risk factors for non-melanocytic skin cancer - a
case-control study. Int J Cancer, 48: 650-662.
Kricker A, Armstrong BK, English D, Heenan PJ, & Randell PL (1991b)
A case-control study of non-melanocytic skin cancer and sun exposure
in Western Australia (Abstract No. III, P2). Cancer Res Clin Oncol,
117(Suppl II): S75.
Kricker A, English DR, Randell PL, Heenan PJ, Clay CD, Delaney TA, &
Armstrong BK (1990) Skin cancer in Geraldton, Western Australia: a
survey of incidence and prevalence. Med J Aust, 152: 399-407.
Kricker A, Armstrong BK, Jones ME, & Burton RC (1993) Health, solar
UV radiation and environmental change. IARC Technical Report No 13.
Lyon, International Agency for Research on Cancer.
Kripke ML (1974) Antigenicity of murine skin tumors induced by
ultraviolet light. J Natl Cancer Inst, 53: 1333-1336.
Kripke Ml & Fidler IJ (1980) Enhanced experimental metastasis of
ultraviolet light-induced fibrosarcomas in ultraviolet
light-iradiated syngeneic mice. Cancer Res, 40: 625-629.
Kripke ML & Fisher MS (1976) Immunologic parameters of ultraviolet
carcinogenesis. J Natl Cancer Inst, 57: 211-215.
Kripke ML & Sass ER ed. (1978) International Conference on
Ultraviolet Carcinogenesis. Natl Cancer Inst Monogr, 50: 16-17.
Kripke ML, Lofgreen JS, Beard J, Jessup JM, & Fisher MS (1977) in
vivo immune responses of mice during carcinogenesis by ultraviolet
irradiation. J Natl Cancer Inst, 59: 1227-1230.
Kromberg JG & Jenkins T (1982) Prevalence of albinism in the South
African Negro. S Afr Med J, 61: 383-386.
Kromberg JGR, Castle D, Zwane EM, & Jenkins T (1989) Albinism and
skin cancer in southern Africa. Clin Genet, 36: 43-52.
Kulandaivelu G, Neducheshian N & Annamalainathan K (1993)
Ultraviolet-B (280-320) radiation induced changes in photochemical
activities and polypeptide components of C3 and C4 chloroplasts.
Photosynthetica, 25:12-14.
Kune GA, Bannerman S, Field B, Watson LF, Clelan H, Merenstein D, &
Vitetta L (1992) Diet, alcohol, smoking, serum ß-carotene, and
vitamin A in male nonmelanocytic skin cancer patients and controls.
Nutr Cancer, 18: 237-244.
Lancaster HO & Nelson J (1957) Sunlight as a cause of melanoma: a
clinical survey. Med J Aust, 1: 452-456.
Langer B & Wellmann E (1990) Phytochrome induction of
photoreactivation in Phaseolus vulgaris L. seedlings. Photochem
Photobiol, 52: 861-864.
Larko O & Diffey BL (1983) Natural UVB radiation received by people
with outdoor, indoor and mixed occupations and UVB treatment of
psoriasis. Clin Exp Dermatol, 8: 279-285.
Lassam NJ, From L, & Kahn HJ (1993) Overexpression of p53 is a late
event in the development of malignant melanoma. Cancer Res, 53:
2235-2238.
Laycock KA, Lee SF, Brady RH, & Pepose JS (1991) Characterization of
a murine model of recurrent herpes simples viral keratitis induced
by ultraviolet B radiation. Invest Ophthalmol Vis Sci, 32:
2741-2746.
Lê, MG, Cabanes, PA, Desvignes, V, Chanteau, MF, Mlika, N, & Avril,
MF (1992) Oral contraceptive use and risk of cutatneous malignant
melanoma in a case-control study in French women. Cancer Causes
Control, 3, 199-205.
Leach, JF, McLeod, VE, Pingstone, AR, Davis, A and Deane, GHW (1978)
Measurement of the ultraviolet doses received by office workers.
Clinical and Experimental Dermatology, 3,77-79.
Leclerc, JE, Borden, A and Lawrence, CW (1991) The thymine-thymine
pyrimidine-pyrimidine (6-4) ultraviolet light photoproduct is highly
mutagenic and specifically induces 3'-thymine-to-cytosine transition
in Escherichia coli Proc Natl Acad Sci USA, 88: 9685-9689.
Lee JAM (1989) The relationship between malignant melanoma of the
skin and exposure to sunlight. Photochem Photobiol, 50: 493-496.
Lee GA & Hirst LW (1992) Incidence of ocular surface epithelial
dysplasia in metropolitan Brisbane: a 10-year survey. Arch
Ophthalmol, 110: 525-527.
Lerman S (1980) Human ultraviolet radiation cataracts. Ophthalmic
Res, 12: 303-314.
Leske MC, Chylack LT, Wu S, & The Lens Opacities Case-Control Study
Group (1991) The lens opacities case-control study: risk factors for
cataract. Arch Ophthalmol, 109: 244-251.
Levi F, La Vecchia C, Te V-C, & Mezzanotte G (1988) Descriptive
epidemiology of skin cancer in the Swiss canton of Vaud. Int J
Cancer, 42: 811-816.
Levine EA, Ronan SG, Shirali SS, & Das Gupta TK (1992) Malignant
melanoma in a child with oculocutaneous ablinism. J Surg Oncol, 51:
138-142.
Lew RA, Sober AJ, Cook N, Marvell R, & Fitzpatrick TB (1983) Sun
exposure habits in patients with cutaneous melanoma: a case control
study. J Dermatol Surg Oncol, 9: 981-986.
Lewis MG (1967) Malignant melanoma in Uganda (The relationship
between pigmentation and malignant melanoma on the soles of the
feet). Br J Cancer, 21: 483-495.
Ley RD (1993) Photoreactivation in humans. Proc Natl Acad Sci (USA),
90: 4337.
Ley RD Peak MJ, & Lyon LL (1983) Induction of pyrimidine dimers in
epidermal DNA of hairless mice by UVB: an action spectrum. J Invest
Dermatol, 80: 188-191.
Ley RD (1985) Photoreactivation of UV-Induced pyrimidine dimers and
erythema in the marsupial Monodelphis domestica. Proc Natl Acad Sci
(USA), 82: 2409-2411.
Ley RD, Applegate LA, Stuart TD, & Fry RJM (1987) UV
radiation-induced skin tumors in Monodelphis domestica.
Photodermatology, 4: 144-147.
Ley RD, Applegate LA, Padilla RS, & Stuart TD (1989) Ultraviolet
radiation-induced malignant melanoma in Monodelphis domestica.
Photochem Photobiol, 50: 1-5.
Ley RD, Applegate LA, Fry RJM, & Sanchez AB (1991) Photoreactivation
of ultraviolet radiation-induced skin and eye tumors of Monodelphis
domestica. Cancer Res, 51: 6539-6542.
Li YF, Kim ST, & Sancer A (1993) Evidence for lack of DNA
photoreactivity enzyme in humans. Proc Natl Acad Sci (USA), 90:
4389-4393.
Lieu F-M, Yamanishi K, Jonishi K, Kishimoto S, & Yasuno, H (1991)
Low incidence of Ha-ras oncogene mutations in human epidermal
tumors. Cancer Lett, 59: 231-235.
Lill PH (1983) Latent period and antigenicity of murine tumors
induced in C3H mice by short-wavelength ultraviolet radiation. J
Invest Dermatol, 81: 342-346.
Lindberg JG (1917) Clinical study on iris depigmentation and on
transparency of iris in cataract and in normal eyes among elderly
persons. Dissertation, Helsinki, Finland (in Swedish).
Lindelöf B, Sigurgeirsson B, Tegner E, Larkö O, Johannesson A, Berne
B, Christensen OB, Andersson T, Törngren M, Molin L,
Nylander-Lundqvist E, & Emtestam L (1991) PUVA and cancer; a
large-scale epidemiological study. Lancet, 338: 91-93.
Lindqvist C (1979) Risk factors in lip cancer: a questionnaire
survey. Am J Epidemiol, 109: 521-530.
Lippke JA, Gordon LK, Brash D, & Haseltine WA (1981) Distribution of
UV light-induced damage in a defined sequence of human DNA:
Detection of alkaline-sensitive lesions at pyrimidine
nucleoside-cytidine sequences. Proc Natl Acad Sci (USA), 78:
3388-3392.
Lischko AM, Seddon JM, Gragoudas ES, Egan KM, & Glynn RJ (1989)
Evaluation of prior primary malignancy as a determinant of uveal
melanoma. A case-control study. Ophtalmology, 96: 1716-1721.
Logani MK, Sambuco CP, Forbes PD, & Davies RE (1984) Skin-tumour
promoting activity of methyl ethyl ketone peroxide - a potent
lipid-peroxidizing agent. Food chem Toxicol, 22: 879-882.
Luande J, Henschke CI, & Mohammed N (1985) The Tanzanian human
albino skin. Cancer, 55: 1823-1828.
Lubin D, Frederick JE, Booth CR, Lucas T, & Neuschuler D (1989)
Measurements of enhanced springtime ultraviolet radiation at Palmer
Station, Antarctica. Geophys Res Lett, 16: 783-785.
Lynch DH & Daynes RA (1983) Evaluation of naturally occurring
cell-mediated cytotoxic activity in normal and UV-irradiated mice.
Transplantation, 35: 216-223.
Lynge E & Thygesen L (1990) Occupational cancer in Denmark. Cancer
incidence in the 1970 census population. Scand J Work Environ
Health, 16(Suppl 2): 1-35.
Lytle CD, Miller SA, Jacobs ME, Cyr WH, James RH, Kaczmarek RG, Beer
JZ, Landry RJ, Sharkness CM, Gaylor D, & De Gruijl FR (1993) An
estimation of squamous cell carcinoma risk from ultraviolet
radiation emitted by fluorescent lamps. In : Proceedings of the 1993
Meeting of the American Society for Photobiology, Chicago, 26-30
June 1993.
McCormick JP, Fisher JR, Pachlatko JP, & Eisenstark A (1976)
Characterization of a cell lethal product from the photooxidation of
tryptophan: hydrogen peroxide. Science, 198: 468-469.
McCredie M & Coates MS (1989) Cancer incidence in migrants to New
South Wales; 1972 to 1984, New South Wales Central Cancer Registry.
Woolloomooloo, New South Wales Cancer Council, pp 22-23, 62-63.
MacDonald EJ (1948) Malignant melanoma in Connecticut. In: The
biology of melanomas. New York, Academy of Sciences, pp 71-81
(Special publication of the Academy of Sciences No. 4).
McGregor JM, Barker JNWN, & MacDonald DM (1992) The development of
excess numbers of melanocytic naevi in an immunosuppressed identical
twin. Clin Exp Dermatol, 16: 131-132.
Mack TM, & Floderus B (1991) Malignant melanoma risk by nativity,
place of residence at diagnosis, and age at migration. Cancer Causes
Control, 2: 401-411.
Mackenzie FD, Hirst LW, Battistutta D, & Green A (1992) Risk
analysis in the development of pterygia. Ophthalmol, 99: 1056-1061.
MacKie RM & Aitchison T (1982) Severe sunburn and subsequent risk of
primary cutaneous malignant melanoma in Scotland. Br J Cancer, 40:
55-960.
MacKie RM, Freudenberger T, & Aitchison TC (1989) Personal
risk-factor chart for cutaneous melanoma. Lancet, ii: 487-490.
McKinlay AF (1992) Artificial sources of UVA radiation. Loss and
emission characteristics. In: Urbach F ed Biological responses to
UVA radiation. Vandermor Pub Corp Kansas.
McKinlay AF & Diffey BL (1987) A reference action spectrum for
ultra-violet induced erythema in human skin. CIE J, 6: 17-22
McKinlay AF & Whillock MJ (1987) Measurements of ultraviolet
radiation from fluorescent lamps used for general lighting and other
purposes in the UK. In: Passchhier WF & Bosnjakovic BFM ed. Human
exposure to ultraviolet radiation: Risks and regulations.,
Amsterdam, Oxford, New York, Elsevier Science Pubishers, pp 253-258.
McKinlay AF, Whillock MJ, & Meulemans CCE (1989) Ultraviolet
radiation and blue-light emissions from spotlights incorporating
tungsten halogen lamps. London, National Radiological Protection
Board (Report NRPB No. R228).
McKinlay AF, Harlen F, & Whillock MJ (1988) Hazards of optical
radiation: A guide to sources, uses and safety. Bristol,
Philadelphia, Adam Hilger.
Madewell BR, Conroy JD, & Hodgkins EM (1981) Sunlight-skin cancer
association in the dog: a report of three cases. J Cutan Pathol, 8:
434-443.
Magnus K (1977) Incidence of malignant melanoma of the skin in 5
Nordic countries: significance of solar radiation. Int J Cancer, 20:
477.
Magnus K (1986) Malignant melanoma in Norway. Tidsskr Nor
Lacgeforen, 106: 2309-2313.
Magnus K (1991) The Nordic profile of skin cancer incidence. A
comparative epidemiological study of the three main types of skin
cancer. Int J Cancer, 47: 12-19.
Mandal TK & Chatterjee SN (1980) Ultraviolet- and sunlight-induced
lipid peroxidation in liposomal membrane. Radiat Res, 83: 290-302.
Mao W & Hu T (1982) An epidemiologic survey of senile cataract in
China. Chin Med J, 95: 813-818.
Marks R & Hill D ed. (1992) The public health approach to melanoma
control: Prevention and early detection. Geneva, International Union
Against Cancer (UICC Publication).
Marks R, Ponsford MW, Selwood TS, Goodman G, & Mason G (1983)
Non-melanocytic skin cancer and solar keratoses in Victoria. Med J
Aust, 2: 619-622.
Marks R, Jolley D, Dorevitch AP, & Selwood TS (1989) The incidence
of non-melanocytic skin cancers in an Australian population: results
of a five-year prospective study. Med J Aust, 150: 475-478.
Marks R, Staples M, & Giles GG (1993) Trends in non melanocytic skin
cancer treated in Australia: the second national survey. Int J
Cancer, 53: 585-590.
Martin EK (1912) The effects of ultra-violet rays upon the eye. Proc
of the Soc Ser B, 85: 319-334.
Mathews-Roth MM & Krinsky NI (1987) Carotenoids affect development
of UV-B induced skin cancer. Photochem Photobiol, 46: 507-509.
Matsunaga T, Hieda K, & Nikaido O (1991) Wavelength dependent
formation of thymine dimers and (6-4) photoproducts in DNA by
monochromatic ultraviolet light ranging from 150 to 365 nm.
Photochem Photobiol, 54: 403-410.
Matsuoka LY, Wortsman J, Hanifan N, & Holick MF (1988) Chronic
sunscreen use decreases circulating concentration of 25-hydroxy
vitamin D. A preliminary study. Arch Dermatol, 124: 1802-1804.
Mayne LV, Mullenders LHF, & vasn Zeeland AA (1988) Cockayne
syndrome: a UV sensitive disorder with a defect in the repair of
tanscribing DNA but normal overall excision repair. In: Friedberg EC
& Hanawalt PC ed. Mechanisms and consequences of DNA damage
processing. New York, Alan R. Liss, pp 349-353.
Menzies, SW, Greenoak, GE, Reeve, VE, & Gallagher, CH (1991)
Ultraviolet radiation-induced murine tumors produced in the absence
of ultraviolet radiation-induced systemic tumor immunosuppression.
Cancer Res, 51: 2773-2779.
Miguel AG & Tyrrell RM (1983) Induction of oxygen-dependent lethal
damage by monochromatic UVB (313 nm) radiation: strand breakage,
repair and cell death. Carcinogenesis 4, 375-380.
Milham S Jr (1983) Occupational mortality in Washington State
1950-1979. Cincinnati, Ohio, National Institute for Occupational
Safety and Health (DHSS (NIOSH) Publication No. 83-116).
Minassian DC, Mehra V, & Johnson GJ (1992) Mortality and cataract:
findings from a population-based longitudinal study. Bull World
Health Organ, 70: 219-223.
Miskin R & Ben-Ishai R (1981) Induction of placmenogen activator by
UV light in normal and Xeroderma pigmentosum fibroblasts. Proc Natl
Acad Sci (USA), 78: 6236-6240.
Mitchell DL, Pfeifer GP, Taylor JS, Zdienicka MZ, & Nikaido O (1993)
Biological role of (6-4) photoproducts and cyclobutane dimers. In:
Shima A, Ichihashi M, Fujiwara Y, & Takebe H ed. Frontiers of
photobiology. Amsterdam, Oxford, New York, Elsevier Science
Publishers, pp 337-344.
Mitchell DL & Rosenstein BS (1987) The use of specific
radioimmunoassays to determine action spectra for the photolysis of
(6-4) photoproducts. Photochem Photobiol, 45: 781-786.
Mitchell RL & Andersen IC (1965) Catalase Photoinactivation. Science
150: 74.
Moan J & Dahlback A (1992) The relationship between skin cancers,
solar radiation and ozone depletion. Br J Cancer, 65: 916-921.
Modan B, Alfandary E, Shapiro D, Lusky A, Chetrit A, Shewach-Millet
M, & Moushovitz M (1993) Factors affecting the development of skin
cancer after scalp irradiation. Radiat Res, 134: 125-128.
Mohan M, Sperduto RD, Angra SK, Milton RC, Mathur RL, Underwood BA,
Jaffery N, Pandya CB, Chhabra VK, Vajpayee RB, Kalra VK, Sharma YR,
& The Indian-US Case-Control Study Group (1989) India-US
case-control study of age-related cataracts. Arch Ophthalmol, 107:
670-676.
Moan J, Dahlbach A, Henriksen T, & Magnus K (1989) Biological
amplification factor for sunlight-induced nonmelanoma skin cancer at
high latitudes. Cancer Res, 49: 5207-5212.
Molès J-P, Moyret C, Guillot B, Jeanteur P, Guilhou J-J, Theillet C,
& Basset-Sèguin N (1993) p53 Mutations in human epithelial skin
cancers. Oncogene, 8: 583-588.
Molesworth EH (1927) Rodent ulcer. Med J Aust, 1: 878-899.
Moller H, Mellemgaard A, Jacobsen GK, Pedersen D, & Storm HH (1993)
Incidence of second primary cancer following testicular cancer. Eur
J Cancer, 29A: 672-676.
Mooy CM, Van der Helm MJ, Van der Kwast ThH, De Jong PTVM, Ruiter
DJ, & Zwarthoff EC (1991) No N- ras mutations in human uveal
melanoma: the role of ultraviolet light revisited. Br J Cancer, 64:
411-413.
Moran DJ & Hollows FC (1984) Pterygium and ultraviolet radiation: a
positive correlation. Br J Ophthalmol, 68: 343-346.
Morimoto S & Kunihiko Y (1989) Psoriasis and vitamin D: A review of
our experience. Arch Dermatol, 125: 231-234.
Morin RJ, Hu B, Peng S-K, & Sevanian A (1991) Cholesterol oxides and
carcinogenesis. J Clin Lab Anal, 5: 219-225.
Morison WL (1984) The effect of a sunscreen containing
para-aminobenzoic acid on the systemic immunologic alterations
induced in mice by exposure to UVB radiation. J Invest Dermatol, 83:
405-408.
Morlière P, Mayson A, Santus R, Huppe G, Mazière JP, & Dubertet L
(1991) UVA-induced lipid peroxidation in cultured human fibroblasts.
Biochem Biophys Acta, 1084(3): 261-268.
Morlière P, Moysan A, Gaboriou F, Santus R, Mazière JC, & Dubertret
L (1992) Ultraviolet A et la peau: implications d'espèces activées
de l'oxygène, tendances actuelles et résultats récents. Pathol Biol,
40: 160-168.
Moseley H (1988) Hospital Physicists' Association - Non-ionizing
radiation, medical physics handbook 18. Bristol, Philadelphia, Adam
Hilger Publishers.
Mountin JW & Dor H(1939) Some peculiarities in the geography of
cancer. J Am Med Assoc, 113: 2405-2410.
Muir C, Waterhouse J, Mack T, Powell J, & Whelan S ed. (1987) Cancer
incidence in five continents, Vol V (IARC Scientific Publications No
88), Lyon, International Agency for Research on Cancer.
Munkata N (1993) Biological effective dose of solar ultraviolet
radiation estimated by spore dosimetry in Tokyo since 1989.
Photochem Photobiol, 58(3): 386-392.
Munch-Petersen B, Frentz G, Squire B, Wallevik K, Horn CC, Reymann
F, & Faber M (1985) Abnormal lymphocyte response to uv radiation in
multiple skin cancer. Carcinogenesis, 6: 843-845.
Murali NS & Teramura AH (1987) Insensitivity of soyabean
photosynthesis to UVB radiation under phosphorus deficiency. J Plant
Nutr, 10: 501-515.
Murphy GF, Walsh Lj, Kaidby K, & Lavker RM (1991) UV and sunbeds
(Abstract) Clin. Res. 39,195.
Murphy GM, Wright J, Nicholls DSH, McKee PH, Messenger AG, Hawk JLM,
& Levene GM (1989) Sunbed induced pseudoporphyria. Br J Dermatol,
120: 555-562.
NSF (1993) National Science Foundation, Div Polar Programs, Report
to P Penhale (NSF) & CR Booth (Biospherical Instruments), Antarctic
Support Assoc US.
Nachtwey DB & Rundel RD (1981) A photobiological evaluation of
tanning booths. Science, 211: 405-407.
Nakazawa H, English D, Randell PL, Nakazawa K, Martel N, Armstrong
BK, & Yamasaki H (in press) UV and skin cancer; specific p53 gene
mutation in normal skin as a biologically relevant exposure
measurement. Proc Natl Acad Sci (USA).
Naumann GOH & Apple D (1986) Pathology of the eye. Berlin,
Heidelberg, New York, Springer-Verlag.
Nelemans PJ, Groenendal H, Kiemeney LALM, Rampen FHJ, Ruiter DJ, &
Verbeek ALM (1993) Effect of intermittent exposure to sunlight on
melanoma risk among indoor workers and sun sensitive individuals.
Environ Health Perspect, 101(3): 252-255.
Nelson EW, Eichwald EJ, & Shelby J (1987) Increased ultraviolet
radiation-induced skin cancers in cyclosporine-treated mice. Transpl
Proc, 19: 526-527.
NIH (1991) Executive summary: Guidelines for the diagnosis and
management of asthma. Bethesda, Maryland, National Institutes of
Health, US Department of Health and Human Services (Publication No.
91-3042A).
Noell WK, Walker VS, & Kang BS (1966) Retinal damage by light in
rats. Invest Ophthalmol Vis Sci, 5: 450-473.
Noonan FP & De Fabo EC (1990) Ultraviolet-B-dose-response curves for
local and systemic immunosuppression are identical. Photochem
Photobiol, 52, 801-810.
Noonan FP, Kripke ML, Pedersen GM, & Green MI (1981a) Suppression of
contact hypersensitivity in mice by ultraviolet irradiation is
associated with defective antigen presentation. Immunology, 43:
527-533.
Noonan FP, De Fabo EC, & Kripke ML (1981b) Suppression of contact
hypersensitivity by UV radiation and its relationship to UV-induced
suppression of tumor immunity. Photochem Photobiol, 34: 683-689.
Noonan FP, De Fabo EC, & Morrison H (1988) Cis-urocanic acid, a
product formed by ultraviolet B irradiation of the skin, initiates
an antigen presentation defect in splenic dendritic cells in vivo.
J Invest Dermatol, 90: 92-99.
Norbury KC, Kripke ML, & Budmen M (1977) in vitro reactvity of
macrophages and lymphocytes from ultraviolet-irradiated mice. J Natl
Cancer Inst, 59: 1231-1235.
Norn MS (1982) Spheroid degeneration, pinguecula, and pterygium
among Arabs in the Red Sea Territory, Jordan. Acta Ophthalmol, 60:
949-954.
Norval M, Howie SE, Ross JA, & Maingay JP (1987) A murine model of
herpes simplex virus recrudescence. J Gen Virol, 68: 2693-2698.
Nultsch W, Pfan J, & Huppertz K (1990) Photoinhibition of
photosynthetic oxygen production and its recovery in the subtidal
red data Polyneura hilliae, Bot. Acta, 103: 62-67.
O'Beirn SF, Judge P, Urbach F, MacCon CF, & Martin F (1970) Skin
cancer in County Galway, Ireland. Proc Natl Cancer Conf, 6: 489-500.
Oberhelman L, Koren H, & Cooper KD (1992) Depressed capacity to
contact sensitiza women at the onset of menses; elevated contact
sensitization capacity during mid cycle can be inhibited by UV. Clin
Res, 40: 542A.
O'Dell BL, Jessen RT, Becker LE, Jackson RT, & Smith EB (1980)
Diminished immune response in sun-damaged skin. Arch Dermatol, 116:
559-561.
Oettlé,AG (1963) Skin cancer in Africa. Natl Cancer Inst Monogr, 10:
197-214.
Office of Population, Censuses and Surveys (1986) Occupational
mortality: the registrar general's decennial, supplement for Great
Britain 1979-80, 1982-83. London, Office of Population Censuses and
Surveys, Her Majesty's Stationary Office (Report Series DS No. 6).
Oldenburg JB, Gritz DC, & McDonnell PJ (1990) Topical ultraviolet
light-absorbing chromophore protects against experimental
photokeratitis. Arch Ophthalmol, 108: 1142-1144.
O'Loughlin C, Moriarty MJ, Herity B, & Daly, L (1985) A re-
appraisal of risk factors for skin carcinoma in Ireland: a case
control study. Ir J Med Sci, 154: 61-65.
Olsen JH & Jensen OM (1987) Occupation and risk of cancer in
Denmark. An analysis of 93 810 cancer cases, 1970-1979. Scand J Work
Environ Health, 13(Suppl 1): 1-91.
Oluwasanmi JO, Williams AO, & Alli AF (1969) Superficial cancer in
Nigeria. Br J Cancer, 23: 714-728.
Omdahl JR, Garry TJ, & Hunsaker A (1982) Nutritional status in the
healthy elderly population: vitamin D. Am J Clin Nutr, 6: 1225-1233.
Organisciak DT & Winkler BS (1993) Retinal light damage: Practical
and theoretical considerations. In: Osborne N & Chader G ed.
Progress in retinal research. Oxford, New York, Seoul, Tokyo,
Pergamon Press.
Orth AB, Teramura AH, & Sisler HD (1990) Effects of UV-B radiation
on fungal disease development in Cucumis sativus. Am J Bot, 77:
1188-1192.
Ostendfeld-Åkerblom A (1988) Pseudoexfoliation in Eskimos (Inuit) in
Greenland. Acta Ophthalmol, 66: 467-468.
Osterlind A (1987) Trends in incidence of ocular malignant melanoma
in Denmark 1943-1982. Int J Cancer, 40: 161-164.
Osterlind A, Olsen JH, Lynge E, & Ewertz M (1985) Second cancer
following cutaneous melanoma and cancers of the brain, thyroid,
connective tissue, bone, and eye in Denmark, 1943-80. Natl Cancer
Inst Monogr, 68: 361-388.
Osterlind A, Hou-Jensen K, & Jensen OM (1988a) Incidence of
cutaneous malignant melanoma in Denmark 1978-1982. Anatomic site
distribution, histologic types, and comparison with non-melanoma
skin cancer. Br J Cancer, 58: 385-391.
Osterlind A, Tucker MA, Stone BJ, & Jensen OM (1988b) The Danish
case-control study of cutaneous malignant melanoma. II. Importance
of UV-light exposure. Int J Cancer, 42: 319-324.
Otani T & Mori R (1987) The effects of UV irradiation of the skin on
herpes simplex virus infection: Alteration in immune function
mediated by epidermal cells and in the course of infection. Arch
Virol, 96(1-2): 1-15.
Oxholm A, Oxholm P, Staberg B, & Bendtzen K (1988)
Immunohistological detection of interleukin-like molecule and tumor
necrosis factor in human epidermis before and after UVB-irradiation
in vivo. Br J Dermatol, 118: 369-376.
Paffenbarger RS Jr, Wing AL, & Hyde RT (1978) Characteristics in
youth predictive of adult-onset malignant lymphomas, melanomas, and
leukemias: brief communication. J Natl Cancer Inst, 60: 89-92.
Paksoy N, Bouchardy C, & Parkin DM (1991) Cancer incidence in
Western Samoa. Int J Epidemiol, 20: 634-641.
Pang Q & Hays JB (1991) UV-B inducible and temperature- sensitive
photoreactivation of cyclobutane pyrimidine dimers in Arabidopsis
thaliana. Plant Physiol, 95: 536-543.
Parkin DM, Muir CS, Whelan SL, Gao Y-T, Ferlay J, & Powell J ed.
(1992) Cancer incidence in five continents, Vol VI (IARC Scientific
Publications No. 120), Lyon, International Agency for Research on
Cancer.
Parrish JA, Jaenicke KF, & Anderson RR (1982) Erythema and
melanogenesis action spectra of normal human skin. Photochem
Photobiol, 36: 187-191.
Patrick MH (1977) Studies on thymine-derived UV photoproducts in DNA
- I. Formation and biological role of pyrimidine adducts in DNA.
Photochem Photobiol, 25: 357-372.
Peak MJ & Peak JG (1982) Single-strand breaks induced in Bacillus
subtilis DNA by ultraviolet light: action spectrum and properties.
Photochem Photobiol, 35: 675-680.
Peak JG & Peak MJ (1990) Ultraviolet light induces double- strand
breaks in DNA of cultured human P3 cells as measured by neutral
filter elution. Photochem Photobiol, 52: 387-393.
Peak JG & Peak MJ (1991) Comparison of initial yields of
DNA-to-protein crosslinks and single-strand breaks induced in
cultured human cells by far-and near-ultraviolet light, blue light
and x-rays, Mutat Res, 246: 187-191.
Peak MJ, Peak JG, Moehring MP, & Webb RB (1984) Ultrviolet action
spectrum for DNA dimer induction, lethality, and mutagenesis in
Escherichia coli with emphasis on the UVB region. Photochem
Photobiol, 40: 613-620.
Peak MJ, Peak JG, Sikorski RS, & Jones CA (1985a) Induction of
DNA-protein crosslinks in human cells by ultraviolet and visible
radiations: action spectrum. Photochem Photobiol, 41: 295-302.
Peak MJ, Peak JG, & Jones CA (1985b) Different (direct and indirect)
mechanisms for induction of DNA-protein crosslinks in human cells by
far- and near-ultraviolet radiation (290 and 405 nm). Photochem
Photobiol, 42: 141-146.
Peak MJ, Peak JG, & Carnes BA (1987) Induction of direct and
indirect single-strand breaks in human cell DNA by far and
near-ultraviolet radiations: action spectrum and mechanisms.
Photochem Photobiol, 45: 381-387.
Perna JJ, Mannix ML, Ronney JF, Notkins AL, & Straus SE (1987)
Reactivation of latent herpes simplex virus infection by ultraviolet
light: A human model. J Am Acad Dermatol, 17(3): 473-478.
Philips (1983) Lighting, comprehensive handbook including technical
section. Croydon, Philips Lighting.
Pierceall WE, Mukhopadhyay T, Goldberg LH, & Ananthasswamy HN (1991)
Mutations in the P53 tumor suppressor gene in human cutaneous
squamous cell carcinomas. Mol Carcinog, 4: 445-449.
Piltingsrud HV, Fong CW, & Odland LT (1978) An evaluation of
ultraviolet radiation personnel hazards from selected 400-watt high
intensity discharge lamps. Am Ind Hyg Assoc J, 39: 406-413.
Pincus MW, Rollings PK, Craft AB, & Green A (1991) Sunscreen use on
Queensland beaches, Australia. J Dermatol, 32: 21.
Pitcher H & Longtreth J (1991) Melanoma mortality and exposure to
ultraviolet radiation: An empirical relationship. Environ Int, 17:
7-21.
Pitts DG (1974) The human ultraviolet action spectrum. Am J Optom
Arch Am Acad Optom, 51: 946-960.
Pitts DG (1978) The ocular effects of ultraviolet radiation. Am J
Optom Phys Optics, 55: 19-35.
Pitts DG, Cullen AP, & Hacker PD (1977) Ocular effects of
ultraviolet radiation from 295 to 365nm. Invest Ophthalmol Vis Sci,
16: 932-939.
Pound AW (1970) Induced cell proliferation and the initiation of
skin tumour formation in mice by ultraviolet light. Pathology, 2:
269-275.
Prystowsky JH (1988) Photoprotection and the vitamin D status of the
elderly. Arch Dermatol, 124: 1844-1848.
Punnonen K, Puntela A, & Ahotupa M (1991) Effects of ultraviolet A
and B irradiation on lipid peroxidation and activity of the
antioxidant enzymes in keratinocytes in culture. Photochem
Photoimmunol Photomed, 8: 3-6.
Putvinsky AV, Sokolov AI, Roshchupkin DI, & Vladimirov Ya (1979)
Electric breakdown of bilayer phospholipid membranes under
ultraviolet irradiation-induced lipid peroxidation. FEBS Lett, 106:
53-55.
Quan MB & Moy RL (1991) The role of human papillomavirus in
carcinoma. J Am Acad Dermatol, 25: 698-705.
Quintern LE, Horneck G, Eischweiler V, & Büker H (1992) A biofilm
used as ultraviolet dosimeter. Photochem Photobiol, 55: 389-395.
Rady P, Scinicariello F, Wagner RF, & Tyring SK (1992) p53 Mutations
in basal cell carcinomas. Cancer Res, 52: 3804-3806.
Rapp LM & Smith SC (1992) Morphorlogical comparisons between
rhodopsin-mediated and short wavelength classes of retinal light
damage. Invest Ophthalmol Vis Sci, 33: 3367-3377.
Rasanen L, Reunala T, Lehto M, Jansen C, Rantala I, & Leinikki P
(1989) Immediate decrease in antigen-presenting function and delayed
enhancement of interleukin-1 production in human epidermal cells
after in vivo UVB irradiation. Br J Dermatol, 120: 589-596.
Rathbun WB (1989) In: Dolphin D, Poulson R, & Avramoric O ed.
Coenzymes and cofactors - Volume III, Glutathione. New York, John
Wiley and Sons, Inc., pp 467-509.
Raven JA, (1991) Responses of aquatic photosynthetic organisms to
increased solar UVB. J Photochem Photobiol, B9: 239-244.
Reeve VE, Greenoak GE, Gallagher CH, Canfield PJ, & Wilkinson FJ
(1985) Effect of immunosuppressive agents and sunscreens on UV
carcinogenesis in the hairless mouse. Aust J Exp Biol Med Sci, 63:
655-665.
Reeve VE, Matheson M, Greenoak GE, Canfield PJ, Boehm- Wilcox C, &
Gallagher CH (1988) Effect of dietary lipid on UV light
carcinogenesis in the hairless mouse. Photochem Photobiol, 48:
689-696.
Reeve VE, Greenoak GE, Canfield PJ, Boehm-Wilcox C, & Gallagher CH
(1989) Topical urocanic acid enhances UV- induced tumour yield and
malignancy in hairless mouse. Photochem Photobiol, 49: 459-464.
Reeve VE, Bosnic M, & Boehm-Wilcox C (1990) Effect of ultraviolet
(UV) radiation and UVB-absorbing sunscreen ingredients on
7,12-dimethyllbenz(a)anthracene-initiated skin tumorigenesis in
hairless mice. Photodermatol Photoimmunol Photomed, 7: 222-227.
Reeve VE, Bosnic M, Boehm-Wilcos C, & Ley RD (1991) Differential
protection by two sunscreens from UV radiation-induced
immunosuppression. J Invest Dermatol, 97: 624-628.
Reeve VE, Bosnic M, & Rozinova E (1993) Carnosine
(beta-alanylhistidine) protects from the suppression of contact
hypersensitivity by ultraviolet B (280-320 nm) radiation or by cis
urocanic acid. Immunology, 78: 99-104.
Remé CH, Braschler U, Roberts J, & Dillon J (1991) Light damage in
the rat retina: effect of a radioprotective agent (WR-77913) on
acute rod outer segment disk disruption. Photochem Photobiol, 54:
137-142.
Reynolds P, Saunders LD, Layefsky ME, & Lemp GF (1993) The spectrum
of acquired immunodeficiency syndrome (AIDS)- associated
malignancies in San Francisco, 1980-1987. Am J Epidemiol, 137:
19-30.
Rhodes AR, Albert LS, Barnhill RL, & Weinstock MA (1991) Sun-induced
freckles in children and young adults: A correlation of clinical and
histopathologic features. Cancer, 67: 1990-2001.
Rigel DS, Friedman RJ, Levenstein MJ, & Greenwald DI (1983)
Relationship of fluorescent lights to malignant melanoma: another
view. J Dermatol Surg Oncol, 9: 836-838.
Ringvold A & Davangar M (1985) Changes in the rabbit corneal stroma
caused by UV irradiation. Acta Ophthalmol, 63: 601-606.
Ringvold A, Davander M, & Olsen EG (1982) Changes of the cornea
epithelium after ultraviolet radiation. Acta Ophthalmol, 60: 41-52.
Rippey J & Schmaman A (1972) Skin tumours of Africans. In: Marshall
J ed. Essays on tropical dermatology. Amsterdam, Excerpta Medica,
vol 2, pp 98-115.
Rivas JM & Ullrich SE (1992) Systemic suppression of delayed-type
hypersensitivity by supenatants from UV-irradiated keratinocytes. An
essential role for keratinocyte-derived Il-10. J Immunol, 149:
3865-3871.
Rivers JK, Norris PG, Murphy GM, Chu AC, Midgley G, Morris J, Morris
RW, Young AR, & Hawk JLM (1989) Tanning, photoprotection, acute
adverse effects and immunological changes. Br J Dermatol, 120:
767-777.
Ro YS, Cooper PN, Lee JA, Quinn AG, Harrison D, Lane D, Horne CHW
Rees JL, & Angus B (1993) p53 Protein expression in benign and
malignant skin tumours. Br J Dermatol, 128: 237-241.
Roberts DL (1990) Incidence of non-melanoma skin cancer in West
Glamorgan, South Wales. Br J Dermatol, 122: 399-403.
Roberts LK & Daynes RA (1980) Modification of the immunogenic
properties of chemically induced tumors arising in hosts treated
concomitantly with ultraviolet light. J Immunol, 125: 438-447.
Roberts JE, Kinley J, Young A, Jenkins G, Atherton S, & Dillon J
(1991) in vivo and photophysical studies on photooxidative damage
to lens proteins and their protection by radioprotectors. Photochem
Photobiol, 53: 33-38.
Robinson JK & Rademaker AW (1992) Relative importance of prior basal
cell carcinomas, continuing sun exposure, and circulating
T-Lymphocytes on the development of basal cell carcinoma. J Invest
Dermatol, 99: 227-231.
Roffo AH (1934) Cancer et soleil: carcinomes et sarcomes provoqués
par l'action du soleil in toto. Bull Assoc Fr Étude Cancer, 23:
590-616.
Roff AH (1939) Physico-chemical etiology of cancer (with special
emphasis on the association with solar radiation). Strahlentherapie,
66: 328-350 (in German).
Ros J (1990) On the effect of UV-radiation on elongation growth of
sunflower seedlings (Helianthus annuus L.)(Thesis), pp. 1-157 in
Karlst. Beitr. Entw Okophysiol 8, M Tevini (ed), Bot Inst II,
Karlsruhe.
Roschupkin DJ, Pelenitsyn AB, Potapenko AY, Talitsky VV & Vladimirov
YA (1975) Study of the effects of ultraviolet light on biomembranes
- IV. The effect of oxygen on UV-induced hemolysis and lipid
photoperoxidation in rat erythrocytes and liposomes. Photochem.
Photobiol. 21: 63-69.
Rose EF (1973) Pigment variation in relation to protection and
susceptibility to cancer. Pigment Cell, 1:, 236-45.
Rosen ES (1986) Filtration of non-ionizing radiation by the ocular
media. In: Cronley-Dillon J, Rosen ES, & Marshall J ed. Hazards of
light: myths and realities of eye and skin. Oxford, New York,
Pergamon Press, pp 145-152.
Rosenstein BS & Ducore JM (1983) Induction of DNA strand breaks in
normal human fibroblasts exposed to monochromatic ultraviolet and
visible wavelengths in the 240-546 nm range. Photochem Photobiol,
38: 51-55.
Rosenthal F, Phoon C, Bakalian A, & Taylor H (1988) The ocular dose
of ultraviolet radiation to outdoor workers. Invest Ophthalmol Vis
Sci, 29: 649-656.
Rosenthal F, Safran M, & Taylor H (1985) The ocular dose of
ultraviolet radiation from sunlight exposure. Photochem Photobiol,
42: 163-171.
Roshchupkin DI, Pelenitsyn AB, Potapenko AY, Talitsky VV, &
Vladimirov YA (1975) Study of the effects of ultraviolet light on
biomembranes. IV. The effect of oxygen on UV-induced hemolysis and
lipid photoperoxidation in rat erythrocytes and liposomes. Photochem
Photobiol, 21: 63-69.
Ross JA, Howie SE, Norval M, Maingay J, & Simpson TJ (1986)
Ultraviolet-irradiated urocanic acid suppresses delayed-type
hypersensitivity to herpes simplex virus in mice. J Invest Dermatol,
87: 630-633.
Roth M, Müller J, & Boyle JM (1987) Immunochemical determination of
an initial step in thymine dimer excision repair in xeroderma
pigmentosum variant fibroblasts and biopsy material from the normal
population and patients with basal cell carcinoma and melanoma.
Carcinogenesis, 8: 1301-1307.
Roy CR & Gies HP (1993) Personal protection against solar
ultraviolet radiation. In: Proceedings of the International
Symposium on UV, Munich 4-6 May 1993. German Ministry of the
Environment, Munich, Germany, pp 47-52.
Roy CR, Gies HP, & Elliot G (1988) Solar ultraviolet radiation:
personel exposure and protection. J Occup Health Safety (Aust N Z),
4: 133.
Roza L, Baan RA, Van Der Leun JC, Kligman L, & Young AR (1989) UVA
hazards in skin associated with the use of tanning equipment. J
Photochem Photobiol, B3: 281-287.
Russell WO, Wynne ES, Loquvam GS, & Mehl DA (1956) Studies on bovine
ocular squamous carcinoma ('cancer eye'). I. Pathological anatomy
and historical review. Cancer, 9: 1-52.
Ryel RJ, Barnes PW, Beyschlag W, Caldwell MM, & Flint SD (1990)
Plant competition for light analyzed with a multispecies canopy
model. I Model development and influence of enhanced UV-B conditions
on photosynthesis in mixed wheat and wild oat canopies. Oecologia,
82: 304-310.
Sadamori N, Mine M, & Honda T (1991) Incidence of skin cancer among
Nagasaki atomic bomb survivors. J Radiation Res, 32, Suppl 2:
217-225.
Saftlas AF, Blair A, Cantor KP, Hanrahan L, & Anderson HA (1987)
Cancer and other causes of death among Wisconsin farmers. Am J Ind
Med, 11: 119-129.
Schaeffer L, Ray R, Humbert S, Moncollin V, Vermeulen W, Hoeijmakers
JHJ, Chambon P, & Egly JM (1993) The basic transcription factor
BTF2/TFIIH contains a helicase involved in both transcription and
DNA repair. Science, Apr 2; 206(5104):37-38.
Scharffetter K, Wlaschek M, Hogg A, Bolsen K, Schothorst A, Goerz G,
Krieg T, & Plewig G (1991) Arch Dermatol Res, 283: 506-511.
Schmitt C, Schmitt J, Wegener A, & Hockwin O (1988) Effect of an
aldose reductase inhibitor, AL-1576, on the development of UV-B and
X-Ray cataract. Graefe's Arch Clin Exp Ophthlamol, 226: 455-460.
Schothorst AA, Slaper H, Schouten R, & Suurmond D (1985) UVB dose in
maintenance psoriasis phototherapy versus solar UVB exposure.
Photodermatology, 3: 213-220.
Schwartz GG & Hulka BS (1990) Is vitamin D deficiency a risk factor
for prostate cancer? Anticancer Res, 10: 1307-1311.
Schwartz SM & Weiss NS (1988) Place of birth and incidence of ocular
melanoma in the United States. Int J Cancer, 41: 174-177.
SCOPE/UNEP (1993) Effects of increased ultraviolet radiation on
global ecosystems (Workshop proceedings). Paris, Scientific
Committee on Problems of the Environment, United Nations Environment
Programme.
Scotto J, Kopf AW, & Urbach F (1974) Non-melanoma skin cancer among
Caucasians in four areas of the United States. Cancer, 34:
1333-1338.
Scotto J & Fears TR (1987) The association of solar ultraviolet and
skin melanoma incidence among Caucasians in the United States.
Cancer Invest, 5: 275-283.
Scotto J, Fears TR, & Fraumeni JF Jr (1982) Solar radiation. In:
Schottenfeld D & Fraumeni JF Jr ed. Cancer epidemiology and
prevention. Philadelphia, Pennsylvania, W.B. Saunders Company, pp
254-276.
Scotto J, Fears TR, & Fraumeni JF Jr (1983) Incidence of nonmelanoma
skin cancer in the United States. Bethesda, Maryland, National
Cancer Institute (NIH Publication No. 83-2433).
Scotto J, Cotton G, Urbach F, Berger D, & Fears T (1988)
Biologically effective ultraviolet radiation: surface measurements
in the US 1974-1985. Science, 235: 762-764.
Seddon JM, Gragoudas ES, Glynn RJ, Egan KM, Albert DM, & Blitzer PH
(1990) Host factors, UV radiation and risk of uveal melanoma. A
case-control study. Arch Ophthalmol, 108: 1274-1280.
Serrano H, Scotto J, Shornick G, Fears TR, & Greenberg ER (1991)
Incidence of nonmelanoma skin cancer in New Hampshire and Vermont. J
Am Acad Dermatol, 24: 574-579.
Servilla KS, Burnham DK, & Daynes RA (1987) Ability of cyclosporine
to promote the growth of transplanted ultraviolet radiation-induced
tumors in mice. Transplantation, 44; 291-295.
Setlow RB, Grist E, Thompson K, & Woodhead AD (1993) Wavelengths
effective in induction of malignant melanoma. Proc Natl Acad Sci
(USA), 90: 6666-6670.
Setlow RB, Woodhead AD, & Grist E (1989) Animal model for
ultraviolet radiation-induced melanoma: platyfish-swordtail hybrid.
Proc Natl Acad Sci (USA), 86: 8922-8926.
Shea CR, McNutt NS, Volkenandt M, Lugo J, Prioleau PG, & Albino AP
(1992) Overexpression of p53 protein in basal cell carcinoma of
human skin. Am J Pathol, 141: 25-20.
Shearer GM & Clerici M (1992) T-helper cell immune dysfunction in
asymptomatic HIV-1 seropositive individuals: The role of TH1-TH2
cross regulation. In: Coffman RL ed. Regulation and functional
significance of T-cell subsets. Prog Chem Immunol, 54, 21-43
Shetlar MD (1980) Cross-linking of proteins to nucleic acids by
ultraviolet light. Photochem Photobiol Rev, 5: 105-197.
Shibata T, Katoh N, Hatano T, & Sasaki K. Population based
case-control study of cortical cataract in the Noto area, Japan,
Ophthalmic Res. (1993, in press).
Shore RE (1990) Overview of radiation-induced skin cancer in humans.
Int J Radiat Biol, 57: 809-827.
Shore RE, Albert RE, Reed M, Harley N, & Pasternack BS (1984) Skin
cancer incidence among children irradiated for ringworm of the
scalp. Radiat Res, 100: 192-204.
Shukla VK, Hughes DC, Hughes LE, McCormick F, & Padua RA (1989) ras
mutations in human melanotic lesions: K- ras activation is a
frequent and early event in melanoma development. Oncogene Res, 5:
121-127.
Siemiatycki J (1991) Risk factors for cancer in the workplace. Boca
Raton, Florida, CRC Press.
Simon JC, Cruz PD, Bergstresser PR, & Tigelaar RE (1990) Low dose
ultraviolet B-irradiated Langerhans cells preferentially activate
CD4+ cells of the T helper 2 subset. J Immunol, 145: 2087-2091.
Simon JC, Tigelaar RE, Bergstresser PR, Edelbaum D, & Cruz PD (1991)
Ultraviolet B radiation converts Langerhans cells from immunogenic
to tolerogenic antigen-presenting cells. J Immunol, 146: 485-491.
Sjovall P, Christensen OB, & Moller H (1985) Single exposure to
ultraviolet irradiation and elicitation of human allergic contact
dermatitis. Acta Derm Venereol, 65: 93-96.
Slaper H (1987) Skin cancer and UV exposure: Investigations on the
estimation of risks. University of Utrecht, The Netherlands (PhD
Thesis).
Slaper H, Schothorst AA, & Van der Leun JC (1986) Risk evaluation of
UVB-therapy for psoriasis: comparison of calculated risk for
UVB-therapy and observed risk in PUVA- treated patients.
Photodermatology, 3: 271-283.
Sliney, DH (1983) Eye protective techniques for bright light.
Ophthalmology 90(8), 937-944.
Sliney D (1986) Physical factors in cataractogenesis: ambient
ultraviolet radiation and temperature. Invest Ophthalmol Vis Sci,
27(5):, 781-790.
Sliney DH (1987) Estimating the solar ultraviolet radiation exposure
to an intraocular lens implant, J Cataract Refract Surg,
13(5),296-301.
Sliney DH & Wolbarsht ML (1980) Safety with lasers and other optical
sources -A comprehensive handbook. New York, London, Plenum Press.
Smith KC (1976) The radiation-induced addition of proteins and other
molecules to nucleic acids. In: Wang SY ed. Photochemistry and
photobiology of nucleic acids. New York, London, San Francisco,
Academic Press, vol 2, pp 187-218.
Smith GJ & Ryan KG (1993) The effect of changes on differences in
Robertson-Berger radiometer responsitivity on solar ultraviolet-B
measurement. Photochem Photobiol, 58(4): 512-514.
Smith RC, Baker KS, Holm-Hansen O, & Olson R (1980) Photoinhibition
of photosynthesis in natural waters. Photochem Photobiol, 31:
585-592.
Smith EL, Walworth NC, & Holick MF (1986) Effects of 2
alpha-25-dihydrxy vitamin D3 on the morphologic and biological
differentiation of cultured human epidermal keratinocytes grown in
serum-free conditions. J Invest Dermatol, 86: 709-714.
Söderberg PG (1989) Mass alteration in the lens after exposure to
ultraviolet radiation, 300nm. Acta Ophthalmol, 67: 633-644.
Söderberg PG (1991) Na and K in the lens after exposure to radiation
in the 300 nm wavelength region. J Photochem Photobiol, 8: 279-294.
Soparker CN, O'Brien JM, & Albert DM (1993) Investigation of the
role of the ras protooncogene point mutation in human uveal
melanomas. Invest Ophthalmol Vis Sci, 34: 2203-2209.
Sorahan T & Grimley RP (1985) The aetiological significance of
sunlight and fluorescent lighting in malignant melanoma: a
case-control study. Br J Cancer, 52; 765-769.
Spector A (1991) In: Sies H ed. The lens and oxidative stress.
Endogenous antioxidant defences in human blood plasma in oxidative
stress: Oxidants and antioxidants. New York, London, San Francisco,
Academic Press.
Spellman CW & Daynes RA (1978) Properties of ultraviolet
light-induced suppressor lymphocytes within a syngeneic tunour
system. Cell Immunol, 36: 383-387.
Spellman CW, Woodward JG, & Daynes RA (1977) Modification of
immunological potential by ultrviolet radiation. I Immune status of
short-term UV-irradiated mice. Transplantation, 24: 112-119.
Spitzer WO, Hill GB, Chambers LW, Helliwell BE, & Murphy HB (1975)
The occupation of fishing as a risk factor in cancer of the lip. N
Engl J Med, 293: 419-424.
Spruance SL (1985) Pathogenesis of herpes simplex labialis:
Experimental induction of lesions with UV light. J Clin Microbiol,
22: 366-368.
Staberg B, Wulf HC, Poulsen T, Klemp P, & Brodthagen H (1983)
Carcinogenic effect of sequential artificial sunlight and UVA
irradiation in hairless mice. Consequences for solarium 'therapy'.
Arch Dermatol, 119: 641-643.
Stadtman ER (1990) Metal ion catalysed oxidation of
protein-biochemical mechanism and biological consequences. Free
Radic Biol Med, 9: 315-325
Stamnes K, Tsay SC, Wiscombe WJ, & Jayaweera K (1988) Numerically
stable algorithm for discreet-ordinate method radiative transfer in
multiple scattering and emitting media. Appl Optics, 27: 2502-2509.
Stein B, Rahmsdorf HJ, Steffen A, Liffin M, & Herrlich P (1989)
UV-induced damage is an intermediate step in UV-induced expression
of human immunodeficiency virus type 1, collagenase, c-fos and
metallothionein. Mol Cell Biol, 9: 5169-5181.
Steinitz R, Parkin DM, Young JL, Bieber CA, & Katz L ed. (1989)
Cancer incidence in Jewish migrants to Israel, 1961-1981 (IARC
Scientific Publications No 98), Lyon, International Agency for
Research on Cancer, 114-115, 134-135.
Stenbäck F (1975a) Species-specific neoplastic progression by
ultraviolet light on the skin of rats, guinea pigs, hamsters and
mice. Oncology, 31: 209-225.
Stenbäck F (1975b) Ultraviolet light irradiation as initiating agent
in skin tumor formation by the two-stage method. Eur J Cancer, 11:
241-246.
Stephenson TJ, Royds J, Silcocks PB, & Bleehen SS (1992) Mutant p53
oncogene expression in keratoacanthoma and squamous cell carcinoma.
Br J Dermatol, 127: 566-570.
Sterenborg HJCM & van der Leun JC (1990) Tumorigenesis by a long
wavelength UVA source. Photochem Photobiol, 51: 325-330.
Sterenborg HJCM, van der Putte SCJ, & van der Leun JC (1988) The
dose-response relationship of tumorigenesis by ultraviolet radiation
of 254 nm. Photochem Photobiol, 47: 245-253.
Stern RS & Docken W (1986) An exacerbation of SLE after visiting a
tanning salon. J Am Med Assoc, 255: 3120.
Stern RS & Lange R (1988) Non-melanoma skin cancer occurring in
patients treated with PUVA five to ten years after the first
treatment. J Invest Dermatol, 91: 120-124.
Stern RS, Thibodeau LA, & Kleinerman RA (1979) Risk of cutaneous
carcinoma in patients treated with oral methoxsalen
photochemotherapy. N Engl J Med, 300: 809-813.
Stocker R & Frei B (1991) In Sies H ed. Endogenous antioxidant
defences in human blood plasma in oxidative stress: Oxidants and
antioxidants.New York, London, San Francisco, Adacemic Press.
Stretch JR, Gatter KC, Ralfkiaer E, Lane DP, & Harris, AL (1991)
Expression of mutant p53 in melanoma. Cancer Res, 51: 5976-5979.
Strickland PT, Burns FJ, & Albert, RE (1979) Induction of skin
tumors in the rat by single exposure to ultraviolet radiation.
Photochem Photobiol, 30: 683-688.
Strickland PT, Creasia D, & Kripke ML (1985) Enhancement of
two-stage skin carcinogenesis by exposure of distant skin to UV
radiation. J Natl Cancer Inst, 74: 1129-1134.
Strickland PT, Vitasa BC, West SK, Rosenthal FS, Emmett EA, & Taylor
HR (1989) Quantitative carcinogenesis in man: solar ultraviolet B
dose dependence of skin cancer in Maryland watermen. J Natl Cancer
Inst, 81: 1910-1913.
Stryker WS, Stampfer MJ, Stein EA, Kaplan L, Louis TA, Sober A, &
Willett WC (1990) Diet, plasma levels of beta-carotene and
alpha-tocopherol, and risk of malignant melanoma. Am J Epidemiol,
131: 597-611.
Sullivan JH & Teramura AH (1991) The effects of UV-B radiation on
loblolly pine. 2 Growth of field-grown seedlings, Trees (Berl)
6(3):115-120.
Sutherland JC & Griffin K (1981) Absorption spectrum of DNA for
wavelengths greater than 300nm. Radiat Res, 86: 399-410.
Swerdlow AJ, English JSC, MacKie RM, O'Doherty CJ, Hunter JAA, Clark
J, & Hole DJ (1988) Fluorescent lights, ultraviolet lamps, and risk
of cutaneous melanoma. Br Med J, 297: 647-650.
Sydenham MM, Wong CF, Hirst LW, & Collins MJ (1991) Cular UVB
dosimetry made possible for the first time using a CR-39 contact
lens. In: Proceedings of 22nd Session of CIE, Melbourne. Vienna,
International Commission of illumination, vol 1, part 2, pp 27-28.
Talbot G (1948) Pterygium. Trans Ophthalmol Soc N Z, 2: 42-45.
Taylor HR (1979) Pseudoexfoliation, an environmental disease? Trans
Ophthalmol Soc UK, 99: 302-307.
Taylor HR (1980a) Aetiology of climatic droplet keratopathy and
pterygium. Br J Ophthalmol, 64: 154-163.
Taylor HR (1980b) The environment and the lens. Br J Ophthalmol, 64:
303-310.
Taylor HR, West SK, Rosenthal FS, Munoz B, Newland HS, Abbey H, &
Emmett EA (1988) Effect of ultraviolet radiation on cataract
formation. N Engl J Med, 319: 1429-1433.
Taylor HR, West SK, Rosenthal FS, Munoz B, Newland HS, & Emmett EA
(1989) Corneal changes associated with chronic UV irradiation. Arch
Ophthalmol, 107: 1481-1484.
Taylor HR, West S, Munoz B, Rosenthal FS, Bressler SB, & Bressler NM
(1992) The long-term effects of visible light on the eye. Arch
Ophthalmol, 110: 99-104.
Taylor JR, Schmieder GF, Shimizu T, & Streilein JW (1993)
UVB-susceptibility is a risk factor for recurrent herpes labialis.
Photochem Photobiol, 57: 135.
Templeton AC (1967) Tumours of the eye and adnexa in Africans of
Uganda. Cancer, 20: 1689-1698.
Teppo L, Pukkala E, Hakama M, Hakulinen T, Herva A, & Saxén E (1980)
Way of life and cancer incidence in Finland. A municipality-based
ecological analysis. Scand J Soc Med, Suppl 19: 5-84.
Terman M, Reme CE, Rafferty B, Gallin PF, & Terman JS (1990) Bright
light therapy for winter depression: Potential ocular effects and
theoretical implications. Yearly Review. Photochem Photobiol, 51:
781-792.
Tevini M, Braun J, & Fieser G (1991a) The protective function of the
epidermal layer of rye seedlings against ultraviolet-B radiation,
Photochem Photobiol, 53: 329-333.
Tevini M, Mark U, & Saile-Mark M (1991b) Effects of enhanced solar
UV-B radiation on growth and function of crop plant seedlings. In:
Randall D & Blevins D ed. Current topics in plant biochemistry and
physiology. Columbia, University of Missouri.
Tevini M, Mark U, Fieser G, & Saile M (1991c) Effects of enhanced
solar UV-B radiation on growth and function of selected crop plan
seedlings. In: Riklis, D ed. Photobiology. New York, London, Plenum
Press, pp 635-649.
Tobal K, Warren W, Cooper CS, McCartney A, Hungerford J, & Lightman
S (1992) Increased expression and mutation of p53 in choroidal
melanoma. Br J Cancer, 66: 900-904.
Toews GB, Bergstresser PR, & Streilein JW (1980) Epidermal
langerhans cell density determines whether contact hypersensitivity
or unresponsiveness follows skin painting with DNFB. J Immunol, 124:
445-453.
Tomatis, L (1990) ed, Cancer: Causes, Occurrence and Control. Lyon,
International Agency for Research on Cancer (IARC Scientific
Publications No 100).
Travis, LB, Curtis, RE, Boice, JD, Hankey, BF, & Fraumeni Jr, JF
(1991) Second cancers following non-Hodgkin's lymphoma. Cancer, 67:
2002-2009.
Travis, LB, Curtis, RE, Hankey, BF, & Fraumeni Jr, JF (1992) Second
cancers in patients with chronic lymphocytic leukemia. J Natl Cancer
Inst, 84: 1422-1427.
Tucker, MA, Boice, JD, Jr & Hoffman, DA (1985a) Second cancer
following cutaneous melanoma and cancers of the brain, thyroid,
connective tissue, bone and eye in Connecticut, 1935-82. Natl Cancer
Inst Monogr, 68: 161-189.
Tucker, MA, Misfeldt, D, Coleman, N, Clark, WH, & Rosenberg, SA
(1985b) Cutaneous malignant melanoma after Hodgkin's Disease. Ann
Int Med, 102: 37-41.
Tucker, MA, Shields, JA, Hartge, P, Augsberger, J, Hoover, RN, &
Fraumeni, JF, Jr (1985c) Sunlight exposure as risk factor for
intraocular malignant melanoma. New Engl J Med, 313: 789-792.
Turner, BJ, Siatkowski, RM, Augsburger, JJ, Shields, JA, Lustbader,
E, & Mastrangelo, MJ (1989) Other cancers in uveal melanoma patients
and their families. Am J Ophthalmol, 107: 601-608.
Tuveson RW & Sammartano, LJ (1986) Sensitivity of HemA mutant
Escherichia coli cells to inactivation by near-UV light depends on
the level of supplementation with L-aminolevulinic acid. Photochem
Photobiol 43: 621-626.
Tyrrell RM (1973) Induction of pyrimidine dimers in bacterial DNA by
365 nm radiation, Photochem Photobiol 17: 69-73.
Tyrrell RM, Ley RD & Webb RB (1974) Induction of single strand
breaks (Alkali-labile bonds) in bacterial and phage DNA by near-UV
(365 nm) radiation. Photochem. Photobiol. 20: 395-398.
Tyrrell RM (1982) Cell inactivation and mutagenesis by solar
ultraviolet radiation In: Hélène C, Charlier M, Montenay-Garestier
Th & Laustriat G ed. Trends in Photobiology, Plenum Press, New York,
pp 155-172.
Tyrrell, RM (1984) Exposure of nondividing populations of primary
human fibroblasts to UV (254 nm) radiation induces a transient
enhancement in capacity to repair potentially lethal cellular
damage. Proc Natl Acad Sci USA 81: 781-784.
Tyrrell, RM (1991) UVA (320-380 nm) radiation as an oxidative stress
In: Sies H ed. Oxidative Stress : oxidants and antioxidants, London,
Academic Press, pp 57-83.
Tyrrell, RM (1992) Inducible responses to UVA exposure. In: (ed. F.
Urbach) Biological responses to ultraviolet A radiation, Valdenmar
Press, Overland Park, pp. 59-64.
Tyrrell, RM and Pidoux, M (1986) Endogenous glutathione protects
human skin fibroblasts against the cytotoxic action of UVB, UVA and
near-visible radiations. Photochem Photobiol 44: 561-564.
Tyrrell RM & Pidoux M (1987) Action spectra for human skin cells.
Estimates of the relative cytotoxicity of the middle ultraviolet,
near ultraviolet and violet regions of sunlight on epidermal
keratinocytes. Cancer Res 47: 1825-1829.
Tyrrell, RM and Pidoux, M (1988) Correlation between endogenous
glutathione content and sensitivity of cultured human skin cells to
radiation at defined wavelengths in the solar UV range. Photochem
Photobiol 47: 405-412.
Tyrrell, RM & Pidoux M (1989) Singlet oxygen involvement in the
inactivation of cultured human fibroblasts by UVA (334 nm, 365 nm)
and near-visible radiations. Photochem Photobiol 49: 407-412.
Ullrich SE (1985) Suppression of lymphoproliferation by
hapten-specific suppressor T lymphocytes from mice exposed to
ultraviolet radiation. Immunology, 54: 343-352.
Ullrich SE (1986) Suppression of the immune response to allogeneic
histocompatibility antigens by a single exposure to ultraviolet
radiation. Transplantation, 42: 287-291.
Ullrich SE & Kripke ML (1984) Mechanisms in the suppression of tumor
rejection produced in mice by repeated UV irradiation. J Immunol,
133: 2786-2790.
Ullrich SE, Azizi E, Kripke ML (1986a) Suppression of the induction
of delayed-type hypersensitivity reactions in mice by a single
exposure to ultraviolet radiation. Photochem Photobiol, 43: 633-638.
Ullrich SE, Yee GK, & Kripke ML (1986b) Suppressor lymphocytes
induced by epicutaneous sensitization of UV- irradiated mice control
multiple immunological pathways. Immunology, 58: 185-190.
UNEP (1987) The ozone layer, United Nations Environment Programme,
UNEP/GEMS Environmental Library No 2, UNEP, Nairobi.
UNEP (1989) Environmental Effects Panel Report, Van der Leun, JC,
Tevini, M eds. United Nations Environment Programme , Nairobi,
Kenya.
UNEP (1991) Environmental effects of ozone depletion: 1991 Update.
United Nations Environment Programme, Nairobi, Kenya.
UNEP (1992) Effects of increased ultraviolet radiation on biological
systems. Proceedings of meeting, Scientific Committee on Problems of
the Environment (SCOPE). Budapest, United Nations Environment
Programme, Nairobi, Kenya.
UNEP (1993) United Nations Environment Programme. Report of the
ninth meeting of the Open-Ended Working Group of the parties to the
Montreal Protocol, Geneva 30 August-1 September 1993.
UNEP-WMO (1989) Scientific assessment of stratospheric ozone: 1989
WMO Global ozone research and monitoring project, United Nations
Environment Programme, World Meteorological Organization, Geneva
(Report No 29).
Urbach F (1987) Man and ultraviolet radiation. In: Passchier WF &
Bosnjakovic BFM eds. Human exposure to ultraviolet radiation - risks
and regulations. New York, Excerpta Medica.
Urbach F (1987a) Cutaneous photocarcinogenesis. J Environ Sci
Health-Environ Carcinogen Rev, C5, 211-34.
Urbach F (1989) Testing the efficacy of sunscreens: effect of choice
of source and spectral distribution of ultraviolet radiation, and
choice of endpoint. Photodermatology, 6: 177-181.
Urbach F, Davies RE, & Forbes PD (1966) Ultraviolet radiation and
skin cancer in man In: Montagna W & Dobson RL eds. Advances in
Biology of Skin, Oxford, Pergamon Press, Carcinogenesis, Vol VII, pp
195-214.
Urbach F, Epstein JH, & Forbes PD (1974) Ultraviolet carcinogenesis:
experimental, global and genetic aspects. In: Pathak MA, Harber LC,
Seiji M, & Kukita A, eds. Sunlight and Man - Normal and Abnormal
Photobiological Responses, Tokyo, University of Tokyo Press, pp
259-283.
US Food and Drug Administration (1978) Sunscreen drug products for
over-the-counter use. Fed Regis, 43: 38206-28269.
US National Cancer Institute (1989) Sunscreens, Rockville, MD,
Tracor Technological Resources Inc (Class Study Report; Contract No
NO1-CP-71082 (7/89)).
Vågerö D, Ringbäck G, & Kiviranta H (1986) Melanoma and other
tumours of the skin among office, other indoor and outdoor workers
in Sweden 1961-1979. Br J Cancer, 53: 507-512.
Vågerö D, Swerdlow AJ, & Beral V (1990) Occupation and malignant
melanoma: a study based on cancer registration data in England and
Wales and in Sweden. Br J Ind Med, 47: 317-324.
Valerie K, Delers A, Bruck C, Thiriatt C, Rosenberg H, Debouck C &
Rosenberg M (1988) Activation of human immmunodefficiency virus type
1 by DNA damage in human cells. Nature 333: 78-81.
Van der Leun JC (1984) Yearly review: UV-carcinogenesis. Photochem
Photobiol, 39: 861-868.
Van der Leun JC (1987) Principles of risk reduction and protection.
In: Passchier WF & Bosnjakovic BFM, eds. Human Exposure to
Ultraviolet Radiation: Risks and Regulations, Amsterdam, Elsevier,
pp 293-303.
Van der Leun JC (1992) Interactions of UVA and UVB in
photodermatology: what was photoaugmentation? In: Urbach F ed. The
Biological Responses to Ultraviolet A Radiation, Overland Park, KS
Valdenmar, pp 309-319.
Van der Leun JC & de Gruijl FR (1993) Influences of ozone depletion
on human and animal health. In: Tevini M ed. UV-B radiation and
ozone depletion. Effects on humans, animals, plants, microorganisms,
and materials. Boc Raton, Lewis Publishers, pp 95-123.
Van der Schroeff JG, Evers LM, Boot AJM & Bos JL (1990) ras oncogene
mutations in basal cell carcinomas and squamous cell carcinomas of
human skin. J Invest Dermatol, 94: 423-425.
Van Weelden H, de Gruijl FR & van der Leun JC (1986) Carcinogenesis
by UVA, with an attempt to assess the carcinogenic risks of tanning
with UVA and UVB. In: Urbach F & Gange RW eds. The Biological
Effects of UVA Radiation, New York, Praeger, pp 137-146.
Van Weelden H and van der Leun J, (1987) IVA induced tumours in
pigmented and albino hairless mice in Human exposure to ultraviolet
radiation - risks and regulations eds. Passchier WF and Bosnjakovic
BFM (New York: Excerpta Medica).
Van Weelden H, de Gruijl FR, van der Putte SCJ, Toonstra J & van der
Leun JC (1988) The carcinogenic risks of modern tanning equipment:
is UVA safer than UVB? Arch Dermatol Res, 280: 300-307.
Van't Veer LJ, Burgering BMT, Versteeg R, Boot AJM, Ruiter DJ,
Osanto S, Schrier PI & Bos JL (1989) N- ras Mutations in human
cutaneous melanoma from sun-exposed body sites. Mol Cell Biol, 9:
3114-3116.
Varghese AJ & Wang SY (1967) Ultraviolet irradiation of DNA in
vitro and in vivo produces a third thymine-derived product.
Science 156: 955-957.
Verhoeff FH, Bell L & Walker CB (1916) The pathological effects of
radiant energy on the eye. Proc Am Acad Arts Sci, 510: 1-810.
Vermeer M & Streilein JW (1990) Ultraviolet B light-induced
alterations in epidermal Langerhans cells are mediated in part by
tumor necrosis factor-alpha. Photodermatol Photoimmunol Photomed, 7:
258-265.
Vermeer M, Schmieder GJ, Yoshikawa T, Van den Berg J-W, Metzman MS,
Taylor JR, & Streilein JW (1991) Effects of ultraviolet B light on
cutaneous immune responses of humans with deeply pigmented skin. J
Invest Dermatol, 97: 729-734.
Vile GF, Basu-Moda S, Waltner C & Tyrrell RM (1994) Haem oxygenase 1
mediates an adaptive response to oxidative stress in human skin
fibroblasts. Proc Natl Acad Sci USA 91: 2607-2610.
Vincek V, Jurimoto I, Medema JP, Prieto E, & Streilein JW (1993)
Tumor necrosis factor alpha polymorphism correlates with deleterious
effects of ultraviolet B light on cutaneous immunity. Cancer Res,
53: 728-732.
Vitale S, West S, Munoz B, Schein OD, Maguire M, Bressler, N &
Taylor HR (1992) Watermen Study II: mortality and baseline
prevalence of nuclear opacity. Invest Ophthalmol Vi Sci, 33: 1097.
Vitasa BC, Taylor HR, Strickland PT, Rosenthal FS, West S, Abbey H,
Ng SK, Munoz B, & Emmett EA (1990) Association of nonmelanoma skin
cancer and actinic keratosis with cumulative solar ultraviolet
exposure in Maryland watermen. Cancer, 65: 2811-2817.
Volkenandt M, Schlegel U, Nanus DM, & Albino AP (1991) Mutational
analysis of the human p53 gene in malignant melanoma. Pigment Cell
Res, 4: 35-40.
Vosjan, JH, Dohler, G, & Nieuwland G (1990) Effect of UVB irradiance
on the ATP content of microorganisms of the Weddell Sea Antarctica,
Neth J Sea Res 25, 391-394.
Vuillaume M, Daya-Grosjean L, Vincens P, Pennetier JL, Tarroux P,
Baret A, Calvayrac R, Taieb A & Satorin, A (1992) Striking
differences in cellular catalase activity between two DNA
repair-deficient diseases. Xeroderma pigmentosum and
trichothiodystophy. Carcinogenesis 13: 321-328.
Walker GC (1987) The SOS response of Escherichia coli In: Niedhardt
FC, Ingraham JL, Low KB, Magasanik B, Shaechter M, Umbarger HE eds.
Escherichia coli and Salmonella typhimurium. Cellular and Molecular
Biology, American Society of Microbiology, Washington DC pp
1346-1357.
Walter SD, Marrett LD, From L, Hertzman C, Shannon HS, & Roy P
(1990) The association of cutaneous malignant melanoma with the use
of sunbeds and sunlamps. Am J Epidemiol, 131: 232-243.
Warfel AH, Moy JA, Meola T, Sanchez M, Soter NA, & Belsito DV (1993)
Effect of ultraviolet B (UVB) on the expression of human
immunodeficiency virus (HIV) in mice and humans, Photochem
Photobiol, 57: 755.
Waring GO, Roth AM, & Ekins MB (1984) Clinical and pathological
description of 17 cases of corneal intraepithelial neoplasia. Am J
Ophthalmol, 97: 547-559.
Waterhouse J, Muir C, Correa P, & Powell J eds (1976) Cancer
Incidence in Five Continents, Lyon, International Agency for
Research on Cancer, Vol III (IARC Scientific Publications No 15).
Waterhouse J, Muir C, Shanmugaratnam K, & Powell J eds (1982) Cancer
Incidence in Five Continents, Lyon, International Agency for
Research on Cancer, Vol IV (IARC Scientific Publications No 42).
Webb RB (1977) Lethal and mutagenic effects of near-ultraviolet
radiation Photochem Photobiol Rev 2: 169-261.
Wei Q, Matanoski GM, Farmer ER, Hedayati MA, & Grossman L (1993) DNA
repair and aging in basal cell carcinoma: A molecular epidemiology
study. Proc Natl Acad Sci USA, 90: 1614-1618.
Weinstock MA, Colditz GA, Willett WC, Stampfer MJ, Bronstein BR,
Mihm MC Jr, & Speizer FE (1989) Nonfamilial cutaneous melanoma
incidence in women associated with sun exposure before 20 years of
age. Pediatrics, 84: 199-204.
Wellmann E, (1971) Phytochrome mediated flavone glycoside synthesis
in cell suspension cultures of Petroselinum hortense after
preirradiation with ultraviolet light, Planta, 101: 283-286.
Wellmann E, (1991) Specific ultraviolet effects in plant
development, J Exp Bot, (Suppl), 32: 42.
Welsh D & Diffey B (1981) Protection against solar actinic radiation
afforded by common clothing fabrics. Clinical and Experimental
Dermatology 6: 577-582.
Werner JS (1991) The damaging effects of light on the eye and
implications for understanding changes in vision across the life
span, in (P Bagnoli and W Hodos) The Changing Visual System, New
York, Plenum Press.
West SK, Rosenthal FS, Bressler NM, Bressler SB, Munoz B, Fine SL, &
Taylor HR (1989) Exposure to sunlight and other risk factors for
age-related macular degeneration. Arch Ophthalmol, 107: 875-879.
Wester A (1987) Ultraviolet transmission properties of sunscreens
and sunglasses. In: Passchier W & Bosnjakovic B eds. Human exposure
to ultraviolet radiation: risks and regulations, Elsevier,
Amsterdam.
Westerveld A, Hoeijmakers JHJ, van Duin M, de Wit J, Odijk H,
Pavlink A, Wood RD & Bootsma D (1984) Molecular cloning of a human
DNA repair gene. Nature 310: 425-529.
Whillock, MJ, Clark, IE, McKinlay, AF, Todd, CD & Mundy, SJ (1988)
Ultraviolet radiation levels associated with the use of fluorescent
general lighting, UVA and UVB lamps in th eworkplace and home.
National radiological Protection Board, Research Report R221,
London, HMSO.
Whillock MJ, McKinlay AF, Kemmlert J & Forsgren PG (1990)
Ultraviolet radiation emissions from miniature [compact] fluorescent
lamps. Lighting Res Technol 22: 125-128.
Whitaker CJ, Lee WR, & Downes JE (1979) Squamous cell skin cancer in
the north-west of England, 1967-69, and its relation to occupation.
Br J Ind Med, 36: 43-51.
WHO (1948) International Statistical Classification of Diseases,
Injuries and Causes of Death, Sixth Revision, Geneva.
WHO (1977) Manual of the International Statistical Classification of
Diseases, Injuries, and Causes of Death. International
Classification of Diseases, 1975 rev, Vol 1, Geneva, p 102.
WHO (1989) Non-ionizing radiation protection. MJ Suess & DA
Benwell-Morison eds. World Health Organization Regional Office for
Europe, European series No 25, WHO, Copenhagen.
WHO/UNEP/IRPA (1979) Ultraviolet Radiation, Environmental Health
Criteria 14, World Health Organization, United Nations Environment
Programme, WHO, Geneva.
Widmark (1889) Ueber den Einfluss des Lichtes auf die vorderen
medien des auges. Skand Arch f physiol 1, 264-330.
Widmark J (1901) Ueber den Einfluss des Lichtes auf die Linse.
Mitteil aus d Augenklin d Carol med chir Inst zu Stockholm 3:
135-149.
Wilesmith JW, Wells GA, Cranwell MP and Ryan JB (1988) Bovine
spongiform encephalopathy : epidemiological studies. Vet Rec, 123,
638-644.
Willis I, Menter JM, & Whyte HJ (1981) The rapid induction of
cancers in the hairless mouse utilizing the principle of
photoaugmentation. J Invest Dermatol, 76: 404-408.
Wittenberg S (1986) Solar radiation and the eye: a review of
knowledge relevant to eye care. Am J Optom Physiol Optics, 63:
676-689.
WMO (1993) World Meteorological Organization Report of the second
meeting of the ozone research managers of the parties to the Vienna
Convention for the protection of the ozone layer. Geneva 10-12
March.
Wolf P, Donawho CK, & Kripke ML (1993a) Analysis of the protective
effect of different sunscreens on ultraviolet radiation-induced
local and systemic suppression of contact hypersensitivity and
inflammatory responses in mice. J Invest Dermatol 100: 254-259.
Wolf P, Yarosh DB, & Kripke ML (1993b) Effects of sunscreens and a
DNA excision repair enzyme on ultraviolet radiation - induced
inflammation, immune suppression and cyclobutane pyrimidine dimer
formation in mice. J invest Dermatol 101: 523-527.
Wong CF, Fleming R, & Carter SJ (1989) A new dosimeter for
ultraviolet B radiation. Photochem Photobiol 50: 611-615.
Wong L, Ho SC, Coggon D, Cruddas AM, Hwang CH, Ho CP, Robertshaw AM,
& MacDonald DM (1993) Sunlight exposure, antioxidant status, and
cataract in Hong Kong fishermen. J Epidemiol Comm Health, 47: 46-49.
Wood RD, Robbins P & Lindahl T (1988) Complementation of the
Xeroderma pigmentosum DNA repair defect in cell-free extracts. Cell
53: 91-106.
Wucherpfenning V (1931) Biologie und prackische verwenbarkeit der
erythemschwelle des UV. Strahlentherapie 40: 201-244.
Yannuzzi L, Fisher Y, Slakter J, Krueger A (1989) Solar retinipathy.
A photobiologic and geophysical analysis, Retina, 9, 28-43.
Yarosh D, Alas LG, Yee V, Oberyszyn A, Kibitel JT, Mitchell D,
Rosenstein R, Spinowitz A, & Citron M (1992) Pyrimidine dimer
removal enhanced by DNA repair liposomes reduces incidence of UV
skin cancer in mice. Cancer Res, 52: 4227-4231.
Yasumoto S, Yoshinobu H, & Aurelian L (1987) Immunity to herpes
simplex virus type 2: Suppression of virus-induced responses in
ultraviolet B-irradiated mice. J Immunol, 139: 2788-2793.
Yoshida A, Ishiguro S, & Tamai M (1993) Expression of glial
fibrillary acidic protein in muoller cells after
lensectomy-vitrectomy. Invest. Ophthalmol. Vis. Sci. 34: 3154-3160.
Yoshikawa T & Streilein JW (1990) On the genetic basis of the
effects of ultraviolet B on cutaneous immunity. Evidence that
polymorphisms at the Tnfý and Lps loci govern susceptibility.
Immunogenetics, 32: 398-405.
Yoshikawa T, Rae V, Bruins-Slot W, Van den Berg J-W, Taylor Jr, &
Streilein JW (1990) Susceptibility to effects of UVB radiation on
induction of contact hypersensitivity as a risk factor for skin
cancer in humans. J Invest Dermatol, 95: 530-536.
Young AR, Magnus IA, Davies AC, & Smith NP (1983) A comparison of
the phototumorigenic potential of 8-MOP and 5-MOP in hairless albino
mice exposed to solar simulated radiation. Br J Dermatol, 108:
507-518.
Young AR, Walker SL, Kinley JS, Plastow SR, Averbeck D, Morlière P,
& Dubertret L (1990) Phototumorigenesis studies of 5-methoxypsoralen
in bergamot oil: evaluation and modification of risk of human use in
an albino mouse skin model. J Photochem Photobiol B Biol, 7:
231-250.
Young AR, Potten CS, Chadwick CA, Murphy GM, Hawk JLM & Cohen AJ
(1991) Photo-protection and 5-MOP photochemoprotection from
UVR-induced DNA damage in humans: the role of skin type. J Invest
Dermatol, 97: 942-948.
Young RW (1991) Age-related cataract. New York, Oxford University
Press.
Yu CC, MacGregor JM, Dublin EA, Barnes DM, MacDonald DM, & Levison
DA (1992) Patterns of immunostaining for p53 in benign and malignant
melanocytic lesions (meeting abstract). J Pathol, 167: (Suppl),
130A.
Zadjela E & Bisagni E (1981) 5-Methoxypsoralen, the melanogenic
additive in sun-tan preparations, is tumorigenic in mice exposed to
365 nm uv radiation. Carcinogenesis, 2: 121-127.
Zamansky G & Chou, I-N (1987) Environmental wavelength of
ultraviolet light induce cytoskeletal damage. J Inv Dermatol 89:
603-606.
Zanetti R, Rosso S, Faggiano F, Roffino R, Colonna S, & Martina G
(1988) A case-control study on cutaneous malignant melanoma in the
province of Torino, Italy (Fr). Rev Epidemiol Santé publ, 36:
309-317.
Zanetti R, Franceschi S, Rosso S, Colonna S, & Bidoli E (1992)
Cutaneous melanoma and sunburns in childhood in a southern European
population. Eur J Cancer, 28A: 1172-1176.
Zaridze D, Mukeria A, & Duffy SW (1992) Risk factors for skin
melanoma in Moscow. Int J Cancer, 52: 159-161.
Zaunuddin D & Sasaki K (1991) Risk factor analysis in a cataract
epidemiology survey in West Samatara, Indonesia. Dev Ophthalmol, 21:
78-86.
Ziegler A-M, Leffell DJ, Kunala S, Sharma HW, Gailani M, Simon JA,
Halperin AJ, Baden HP, Shapiro PE, Bale AE, & Brash DE (1993)
Mutation hotspots due to sunlight in the p53 gene of nonmelanoma
skin cancers. Proc Natl Acad Sci USA, 90: 4216-4220.
Zigman S (1993) Yearly review: ocular light damage. Photochem.
Photobiol. 57: 1060-1068.
Zigman S, Yulo T, & Schultz J (1974) Cataract induction in mice
exposed to near UV light. Ophthalmol Res, 6: 259-270.
Zigman S, Graff J, Yulo T, & Vaughen T (1975) The response of mouse
ocular tissue to continuous near-UV light exposure. Invest
Ophthalmol, 14: 710-713.
Zmudzka BZ & Beer JZ (1990) Activation of human immunodeficiency
virus by ultraviolet radiation. Photochem Photobiol, 52: 1153-1162.
Zuclich JA (1989) Ultraviolet induced photochemical damage in ocular
tissues. Health Physics 56: 671-682.
Zuclich J & Kurtin W (1977) Oxygen dependence of near-ultraviolet
induced corneal damage. Photochem Photobiol 25: 133-135.
Zundorf, I, & Hader, DP (1991) Biochemical and spectroscopic
analysis of UV effects on the marine flagellate, cryoptomonas
maculata, Arch Microbiol.