
First draft prepared by
E. Vavasour
Health Canada, Ottawa, Canada
and Junshi Chen
Chinese Centers for Disease Control and Prevention, Beijing, China
|
Contribution from added mineral hydrocarbons and naturally occurring hydrocarbons |
|
Intake due to migration from coating and packaging materials |
Mineral oils (medium- and low-viscosity) comprise a subgroup of the family of mineral oils and waxes. They are manufactured from crude mineral oils in various refining steps, such as distillation, extraction and crystallization, and are subsequently purified by acid treatment (oleum method) and/or hydrotreatment (catalytic hydrogenation). Mineral oils (medium- and low-viscosity) are mixtures of highly refined paraffinic and naphthenic liquid hydrocarbons with boiling-points greater than 200 °C.
In order to define the materials more clearly, the Committee at its forty-fourth meeting (Annex 1, reference 116) prepared individual specifications for micro-crystalline wax and for two groups of mineral oils: mineral oils (high-viscosity) and mineral oils (medium- and low-viscosity). The latter group was furthermore divided into three classes. For the purpose of characterization of the different types of oils and waxes, the criteria viscosity, average relative molecular mass and carbon number at 5% distillation-point were used in the specifications (see Table 1).
Table 1. Classification of mineral hydrocarbons
|
Name |
ADI |
Viscosity at |
Average relative molecular mass |
Carbon number at 5% distillation-point |
|
Microcrystalline wax |
0–20a,b |
>11 |
>500 |
>25 |
|
High-melting-point wax |
Not included in present studies |
|||
|
Low-melting-point wax |
Withdrawnc |
No JECFA specification |
||
|
Low-melting-point wax |
3.3 |
380 |
22 |
|
|
Mineral oil (high viscosity) |
0–20a |
> 11 |
> 500 |
> 28 |
|
P100 |
11 |
520 |
29 |
|
|
Mineral oil (medium and low viscosity) |
0–10d |
8.5–11 |
480–500 |
>25 |
|
class I |
||||
|
P70 |
9.0 |
480 |
27 |
|
|
Medium-viscosity liquid petroleum |
8.7 |
480 |
25 |
|
|
P70(H) |
8.6 |
480 |
27 |
|
|
Mineral oil (medium and low viscosity) |
0–0.01e |
7.0–8.5 |
400–480 |
>22 |
|
class II |
||||
|
N70(H) |
7.7 |
420 |
23 |
|
|
Mineral oil (medium and low viscosity) |
0–0.01e |
3.0–7.0 |
300–400 |
>17 |
|
class III |
||||
|
P15(H) |
3.5 |
350 |
17 |
|
|
N15(H) |
3.5 |
330 |
17 |
|
|
P100 oil, crude: paraffinic, viscosity (40 °C): 100 mm2/s |
|
|
P70 oil, crude: paraffinic, viscosity (40 °C): 70 mm2/s |
|
|
P70(H) oil, crude: paraffinic, viscosity (40 °C): 70 mm2/s, hydrotreated (catalytic hydrogenation) |
|
|
N70(H) oil, crude: naphthenic, viscosity (40 °C): 70 mm2/s, hydrotreated (catalytic hydrogenation |
|
|
P15(H) oil, crude: paraffinic, viscosity (40 °C): 15 mm2/s, hydrotreated (catalytic hydrogenation |
|
|
N15(H) oil, crude: naphthenic, viscosity (40 °C): 15 mm2/s, hydrotreated (catalytic hydrogenation) |
|
|
a |
Established by the Committee at its forty-fourth meeting |
|
b |
Group ADI |
|
c |
Previous ADI ‘not specified’ |
|
d |
Established at the present meeting |
|
e |
Temporary group ADI, extended at the present meeting |
Food-grade mineral oils were last evaluated by the Committee at its forty-fourth meeting (Annex 1, reference 116), when four 90-day studies conducted in Fischer 344 rats with a range of mineral oils and waxes representative of materials in commercial use were reviewed. The materials tested included low-, medium- and high-viscosity mineral oils (N10(A), N15(H), P15(H), N70(A), N70(H), P70(H) and P100(H)); paraffin waxes (low-melting-point wax and intermediate-melting-point wax); and microcrystalline waxes (high-melting-point wax and high-sulfur wax). All the substances, with the exception of the microcrystalline waxes, appeared to accumulate in the tissues of the animals to varying degrees, depending on the material and dose. Except for P70(H) and P100(H) oils, there was evidence of accumulation of the mineral hydrocarbons and effects indicative of a reaction to a foreign body at one or more doses. The types of effects seen were similar and included focal histiocytosis, increased weights of liver, lymph nodes, spleen and kidneys, granulomas or microgranulomas of the liver, haematological changes typical of a mild, chronic inflammatory reaction and biochemical changes indicative of mild hepatic damage.
The ADI of 0–20 mg/kg bw for mineral oils with the specifications of high-viscosity oils and of high-melting-point and high-sulfur waxes was based on NOELs at the highest dose tested (2% in the diet) in 90-day studies in Fischer 344 rats. The NOELs for all the other materials except P70(H) oil (i.e. class II and III medium- and low-viscosity oils and low-melting-point wax) were based on an increased incidence of histiocytosis in the lymph nodes at the next highest dose. For P70(H) oil, the NOEL was based on an increased incidence of pigmented macrophages in male rats at a dietary concentration of 2%, a minor effect considered of doubtful biological significance. Accordingly, because effects were observed at all doses, the ADIs for low-melting-point and intermediate-melting-point waxes were withdrawn. A group temporary ADI of 0–0.01mg/kg bw was allocated for class II and III medium- and low-viscosity mineral oils, the temporary nature of the ADI being due to uncertainty about the long-term significance of the inflammatory response to accumulated dietary mineral hydrocarbons. A temporary ADI of 0–1 mg/kg bw was allocated for P70(H) oil. These temporary ADIs were extended at the fifty-first meeting (Annex 1, reference 137).
At its forty-fourth meeting, the Committee considered that, although the types of effects seen were essentially reactions to a foreign body, it was possible that a prolonged inflammatory response of the type observed could result in functional changes in the immune system and that this aspect required further investigation. It also noted that the oils and waxes for which high NOELs were observed contained a greater proportion of hydrocarbon components of high relative molecular mass (high carbon number) and had higher viscosities than those materials with a low NOEL, which contained a greater proportion of hydrocarbon components of lower relative molecular mass (low carbon number). The Committee required information about the compositional factors in mineral oils that influence their absorption and toxicity. It also required a study of at least 1 year’s duration on one of these materials in Fischer 344 rats, including an assessment of immune function at appropriate times and an investigation of the kinetics of accumulation of the material, to determine whether a plateau is reached. It recommended that a reversal period of 1 year be included, in order to determine whether the granulomatous hepatic lesions observed in rats in the 90-day studies were fully reversible.
At its present meeting, the Committee reviewed a number of new studies, including the results of a combined 2-year study of toxicity and carcinogenicity and a 1-year study of toxicity with a 1-year recovery period with P70(H) and P100(H) oils (with the specifications of class-I medium- and low-viscosity mineral oils and of high-viscosity mineral oil, respectively) conducted in parallel. In addition, the Committee received and reviewed a number of studies conducted with low- and medium-viscosity mineral oils, including: a 2-year study of the carcinogenicity of a medium-viscosity liquid petroleum (class I medium- and low-viscosity mineral oil); studies of pharmacokinetics and studies of humoral immune function after administration to P15(H) mineral oil (class III medium- and low-viscosity mineral oil) in Fischer 344 and Sprague-Dawley rats; and a 90-day study of histopathological responses and compositional analysis of absorbed hydrocarbons with N15(H), N70(H) and P70(H) oils (classes III, II and I medium- and low-viscosity mineral oils, respectively). Several studies of low-melting-point paraffin wax were also reviewed by the Committee, as they were considered to provide information relevant for the evaluation of low- and medium-viscosity mineral oils relating to the difference in response to mineral hydrocarbons in Fischer 344 and Sprague-Dawley rats.
Differences in the disposition and pharmacokinetics of a low-viscosity mineral oil were assessed in rats of two strains given a representative mineral hydrocarbon component. For the study of disposition, fasted female Fischer 344 and Sprague-Dawley rats received a single dose by gavage of 2 ml/kg bw of a 4:1 mixture (v/v) of olive oil and a food-grade paraffinic white mineral oil meeting the specifications for medium- and low-viscosity mineral oil class III, containing [1-14C]-1-eicosanylcyclo-hexane and a non-absorbable marker, [1,2-3H]polyethylene glycol 4000. The dose of mineral oil was 340 mg/kg bw. The rats were placed in metabolism cages for collection of urine, faeces and expired air at regular intervals over 96 h, at which time they were killed, and liver and mesenteric lymph nodes were collected. For the study of pharmacokinetics, the rats received the same single dose of the olive oil:mineral oil mixture containing [1-14C]-1-eicosanylcyclohexane by gavage, and serial blood samples were collected from in-dwelling jugular vein catheters at regular intervals for up to 96 h, in addition to the collection of urine and faeces. Three rats of each strain were killed at each time, and blood, liver and mesenteric lymph nodes and the contents of the bladder and intestines were analysed for total radioactivity.
A comparison of the values for the area under the curve of the plasma concentration versus time (AUC), an indicator of overall systemic exposure, indicated that the bioavailability of total 14C label was greater in Fischer 344 rats than in Sprague-Dawley rats. The maximum blood concentration attained (Cmax), and the time to reach Cmax were also greater in Fischer 344 rats. Faecal excretion of 14C-labelled material was the major route of elimination for both strains of rat, although there were strain differences in the rate of faecal excretion. The majority of 14C label (70%) had been eliminated by 16 h in the Sprague-Dawley rats, which corresponded to the gastrointestinal transit time in both strains. In contrast, only 11% of the 14C label had been eliminated by 16 h in the Fischer 344 rats, and the majority (75%) was eliminated within 48 h. The authors speculated that the longer delay in faecal excretion of labelled hydrocarbon in Fischer 344 than in Sprague-Dawley rats was the result of biliary excretion of absorbed 14C label or sloughing of gastrointestinal endothelial cells containing the label. Urinary excretion was the second route of elimination of 14C-containing products and reached near-maximal values (10%) by 24 h in Sprague-Dawley rats. By contrast, urinary excretion of radioactivity by Fischer 344 rats was linear over time during the 96-h monitoring period, attaining 7% of the administered 14C label. The authors indicated that this was characteristic of a depot effect in which the rate of elimination is determined by the export of 14C from a peripheral compartment before urinary excretion, in this case, probably the liver. In the livers of Sprague-Dawley rats, the maximum amount of radioactivity (2%) occurred at 8 h and decreased thereafter to reach 0.1% by 96 h. By contrast, the maximum amount of radioactivity in the livers of Fischer 344 rats (4%) was not reached until 24 h, and 3% of the administered radioactivity was still present at 96 h. The authors concluded that Sprague-Dawley rats can efficiently metabolize and excrete 14C-containing compounds, whereas the inability of Fischer 344 rats to clear these compounds from the liver probably resulted in deposition and possible retention. The amount of radioactivity in the mesenteric lymph nodes was similar in Fischer 344 and Sprague-Dawley rats until the 96-h sampling time, when the percentage of the administered radioactivity increased by nearly 10-fold, from 0.002% to 0.02%, in Sprague-Dawley rats (Sipes & Halladay, undated).
The above study was repeated with a 10-fold lower dose of the olive oil:mineral oil mixture. Fasted female Fischer 344 and Sprague-Dawley rats received 0.2 ml/kg bw of the oil mixture, to deliver a single oral dose of 34 mg/kg bw of the low-viscosity oil (mineral oil, medium and low viscosity, class III), by gavage. As in the previous study, [1-14C]-1-eicosanylcyclohexane (0.34 mg/kg bw) was used as a tracer. The animals were placed in metabolism cages for the collection of urine and faeces at regular intervals up to sacrifice at 96 h. Blood samples were collected from an in-dwelling jugular vein catheter, also at regular intervals. Liver and mesenteric lymph nodes were collected and analysed for radioactivity at sacrifice. In contrast to the results with the high dose, the radioactivity in blood of Fischer 344 rats showed a biphasic profile, with two Cmax values at 4 and 16 h (one at 18 h with the high dose). The Cmax in Sprague-Dawley rats was at 5–5.6 h in both studies. As in the previous study, the AUC was greater in Fischer 344 than in Sprague-Dawley rats, indicating greater bioavailability. Faecal excretion was the major route of excretion, with 76% (Fischer 344) and 72% (Sprague-Dawley) of the administered radioactivity eliminated by 96 h. The delay in faecal elimination of radioactivity seen in Fischer 344 rats was less apparent with the low dose. The differences in patterns of urinary excretion and liver retention of radioactivity by Fischer 344 and Sprague-Dawley rats were similar to those observed with the high dose. As in the study with the high dose, more radioactivity (approximately 2.5% of the administered dose) was retained in the liver of Fischer 344 than Sprague-Dawley rats (0.5%), and the radioactivity retained in lymph nodes in Fischer 344 rats exceeded that in Sprague-Dawley rats over the 96-h observation period by twofold (approximately 0.02% versus 0.01%) (Sipes & Halladay, 2001a).
The study of disposition of low-viscosity mineral oil, described above, was repeated with the addition of a 2-week dietary pretreatment before the single gavage dose of radiolabelled tracer. Female Fischer 344 and Sprague-Dawley rats were fed standard diet containing 2% white oil (mineral oil, medium and low viscosity, class III) or untreated diet for 2 weeks, during which time food consumption and body weight were monitored. At the end of the 2-week period, the treated animals received a single oral dose of the white oil (340 mg/kg bw) in a 1:4 ratio with olive oil (2 ml/kg) containing [1-14C]-1-eicosanylcyclohexane (2.1 mg/kg bw). The animals were placed in metabolism cages for the collection of urine and faeces at regular intervals up to 96 h. At sacrifice, liver and mesenteric lymph nodes were collected for determination of total radioactivity.
Consumption of mineral oil in the diet during the pretreatment period did not affect weight gain or food consumption, although Sprague-Dawley rats had a higher rate of food consumption and greater weight gain than Fischer 344 rats. Faecal excretion was the major route of excretion, with 84% (Fischer 344) and 94% (Sprague-Dawley) of the administered radioactivity eliminated by 96 h, compared with 92% and 88%, respectively, in un-pretreated control animals and 82% and 87% in the study with a high single dose by gavage. The delay in faecal elimination of radioactivity by Fischer 344 rats in the study with the single high dose was shorter. The differences in patterns of urinary excretion of radioactivity between Fischer 344 and Sprague-Dawley rats were similar to those observed with the single high dose (nearly complete by 16 h in Sprague-Dawley rats, linear over the 96-h observation period in Fischer 344 rats). However, more 14C label was excreted in the urine of Fischer 344 rats after dietary pretreatment with mineral oil than in un-pretreated controls (19% versus 11%), and slightly less label was excreted in the urine of Sprague-Dawley rats (8% versus 11%) than in un-pretreated controls.
As the data on faecal excretion by rats that did not receive pretreatment in this study and that with the single high dose are inconsistent, no conclusion could be reached about whether the increased urinary excretion of radioactive tracer in pretreated Fischer 344 rats indicated increased absorption of mineral hydrocarbon or reduced retention. The proportion of administered radioactivity recovered in the liver was not altered by mineral oil pretreatment and, as observed previously, was greater in Fischer 344 than in Sprague-Dawley rats (about 2% versus 0.1%). In the mesenteric lymph nodes, Fischer 344 rats again retained more radioactivity than Sprague-Dawley rats (0.03% versus 0.003%). A threefold increase in retention of 14C label in mesenteric lymph nodes was seen in pretreated Fischer 344 rats as compared with un-pretreated controls after 96 h, although no difference in retention was seen from that of rats given a single dose of radioactive tracer by gavage in the previous study. Inconsistencies between the results obtained for Fischer 344 rats in the control group in this study and those receiving a single high dose by gavage in the previous study make it difficult to draw any conclusion about the possible effects of mineral oil pretreatment on the disposition of 14C-labelled tracer in the mesenteric lymph nodes. These results confirm the observation in the previous studies, that Fischer 344 and Sprague-Dawley rats differ in terms of the amount of mineral hydrocarbon retained in liver and mesenteric lymph nodes (Sipes & Halladay, 2001b).
A qualitative investigation was conducted to determine whether low-melting-point paraffin wax accumulated in the liver and Kupffer cells of female Fischer 344 and Sprague-Dawley rats fed 0 or 2% in the diet for 14, 30 or 60 days. Extracts of livers from Fischer 344 rats fed 2% in the diet for 60 days and of Kupffer cells from Fischer 344 rats treated for 14 and 30 days contained detectable amounts of low-melting-point paraffin wax. In contrast, Sprague-Dawley rats did not accumulate detectable amounts in liver or Kupffer cells, as determined by gas chromatography and mass spectrometry (Sipes & Hoglen, 1998).
The comparative efficiency of N-hydrocarbon metabolism in vitro was investigated in liver or small intestinal microsomes prepared from Wistar, Sprague-Dawley and Fischer 344 rats. Two linear saturated hydrocarbons, N-octadecane (C18) and N-heptadecane (C17), were used as representative substrates. For the assay, 2 mg of microsomal protein were incubated for 2 h with 84 µmol/l [14C]heptadecane or 17, 54 or 84 µmol/l [14C]octadecane. The extracts from the incubations were analysed for oxidative metabolites by high-performance liquid chromatography. No hydrocarbon metabolism was detected in the small intestinal microsomal preparations with 84 µmol/l heptadecane or 54 µmol/l octadecane. No significant hydroxylation of octadecane was seen after incubation of octadecane with liver microsomes at any of the three concentrations. Incubation of liver microsomes with 84 µmol/l heptadecane resulted in the production of three metabolites: the corresponding alcohol and acid, heptadecanol and heptadecanoic acid. There was no apparent sex-related difference in the rate of metabolism. The overall rate of hydroxylation of heptadecane was calculated to be 3.6%, 1.4% and 0.75% for Wistar, Sprague-Dawley and Fischer 344 rats, respectively (Institut National de la Recherche Agronomique, 2000).
The effects of low-melting point paraffin wax were compared in a 90-day study in female Fischer 344 and Sprague-Dawley rats. This mineral hydrocarbon was selected since it was reported to have more severe effects in Fischer 344 rats than the other oils and waxes examined. The study was conducted in compliance with US Food and Drug Administration Good Laboratory Practice Regulations and the OECD Principles of Good Laboratory Practice.
The study was divided into a component for determination of toxicity and one for analysis of mineral hydrocarbon concentrations. For the study of toxicity, groups of 10 female rats of each strain were fed a standard diet containing 0.0, 0.2 or 2.0% low-melting-point paraffin wax (100% pure) (equal to 0, 160 and 1600 mg/kg bw per day) for 90 days or 0.0 or 2.0% for 30 or 60 days. For the analysis of mineral hydrocarbon, groups of five female rats of each strain were fed diets containing 0.0, 0.2 or 2.0% low-melting-point paraffin wax for 90 days or 2.0% for 30 or 60 days. The food consumption and body weights of all groups were recorded weekly. The animals were killed at end of the respective feeding periods. For the study of toxicity, blood was collected from all animals before sacrifice for haematological and clinical chemical evaluations. All animals were subjected to a macroscopic post-mortem examination, and the weights of the heart, kidneys, liver, mesenteric lymph nodes, spleen and ovaries were determined. Histological examinations were performed on the heart (with mitral valve), kidneys, liver (caudal and medial lobes), cervical and mesenteric lymph nodes, ovaries and spleen from control and treated animals killed after 90 days, and on the heart (with mitral valve), liver (caudal and medial lobes) and mesenteric lymph nodes from controls and animals at the high dose killed at 30 and 60 days. In addition, sections of the duodenum, jejunum, ileum and caecum from three controls and three rats of each strain at the high dose killed at 90 days were examined histologically. For analysis of mineral hydrocarbon, heart, kidney, liver, mesenteric lymph node and spleen were collected from all animals at the 90-day sacrifice for measurement of total mineral hydrocarbon, and liver and mesenteric lymph nodes were also collected from animals at the high dose at the 30- and 60-day sacrifices. In addition, liver tissue from selected groups was collected and preserved for electron microscopic analysis, while other tissues were used to evaluate immunological and pathological end-points, as described in sections 2.2.5 and 2.2.6.
No deaths occurred during the study. Treatment did not adversely affect body-weight gain, food consumption or physical parameters in either strain. In the Fischer 344 rats, statistically significant, treatment-related effects were observed in: haematological parameters (decreased erythrocyte count, erythrocyte volume fraction, haemoglobin concentration and platelet count; increased neutrophil count; values for monocytes were not presented); clinical chemical parameters (increased serum activity of alanine and aspartate aminotransferases, alkaline phosphatase and gamma-glutamyl transpeptidase; decreased serum albumin); organ weights (increased liver, mesenteric lymph node, spleen and ovary weights), macroscopic findings (discoloured liver, enlarged mesenteric lymph nodes); and histological changes in the liver (increased incidence and severity of vacuolated hepatocytes, time-related development of microgranulomas, lymphoid-cell infiltrates or aggregates and scattered foci of necrosis), mesenteric lymph nodes (increased incidence and severity of microgranulomas and reticuloendothelial-cell hyperplasia), cervical lymph node (development of microgranulomas in 3/9 animals at the high dose), spleen (increased severity of extramedullary haematopoiesis) and heart (time-related increase in incidence of lymphoid-cell infiltrates in the base of the mitral valve). In animals at the high dose, many of these effects were seen as early as 30 days into the study. With the exception of inflammation of the mitral valve (affecting 1/10, 2/10 and 5/10 rats in the control group and at the low and high doses, respectively) and microgranulomas in the cervical lymph node (affecting 0/10, 2/9 and 3/9 rats in the three groups, respectively), the histopathological changes affected all rats at the high dose (10/10). Slight reticuloendothelial cell hyperplasia was also observed in the Peyer patch of the jejunum of one of three Fischer 344 female rats at the high dose. The only treatment-related effects observed in the Sprague-Dawley animals were increased weight of the mesenteric lymph nodes accompanied by microscopic changes (microgranulomas and reticuloendothelial-cell hyperplasia).
The mineral hydrocarbon concentrations in tissues measured over the course of the study were also indicative of a clear strain difference. The Fischer 344 rats showed a significant, time- and dose-related accumulation of mineral hydrocarbon in the mesenteric lymph nodes and liver. No plateau was seen during the 90 days of the study. In Sprague-Dawley rats, slight accumulation of mineral hydrocarbon was observed only in the mesenteric lymph nodes and only after 90 days of treatment. There was no notable accumulation in the heart, kidney or spleen in either strain of rat, regardless of dose and treatment duration (Thornton et al., 2000).
A 28-day study and a 90-day study were conducted to confirm the findings of previous feeding studies with mineral oils and waxes and to extend these observations to include a synthetic wax and a wider range of tissue samples for histopathological examination and chemical analysis. Groups of 12 female Fischer 344 rats were fed diets for either 28 or 90 days containing 2% of low-melting-point wax, C80 synthetic wax or one of three white oils, N15H (mineral oil, medium and low viscosity, class III), N70H (mineral oil, medium and low viscosity, class II) or P70H (mineral oil, medium and low viscosity, class I), providing doses of 2500, 2600, 2500, 2600 and 2600 mg/kg bw per day, respectively, for the groups treated for 28 days and 2100, 2100, 2000, 2000 and 2100 mg/kg bw per day, respectively, for those treated for 90 days. Respective control groups received standard diet containing < 0.01% mineral hydrocarbon by weight. Four animals in each group at each time were designated for chemical analysis of tissues, and the remaining eight animals per group were designated for histological examination. Body weight and food intake were determined twice a week throughout the study. On day 14 of the study and on the day before necropsy, urine and faeces were collected during 4 h from the animals in each group designated for chemical analysis; blood samples were collected from these animals at necropsy. At scheduled necropsy, the weights of the brain, heart, kidney, liver, mesenteric lymph nodes and spleen from all the animals were recorded. In addition, the kidney, liver, mesenteric and cervical lymph nodes, small intestine and spleen from all animals were evaluated histopathologically, and the mineral hydrocarbon content was measured in the kidney, liver, mesenteric and cervical lymph nodes and small intestine. Mineral hydrocarbon accumulation was assessed as part of the histopathological examination.
No deaths were reported. Consumption of 2% mineral hydrocarbon in the diet did not affect body-weight gain. Food consumption was statistically significantly increased in the groups receiving the white oils (N15H, N70H and P70H) throughout most of the treatment period of each study. While increased food consumption was noted with diets containing low-melting-point paraffin wax at various times during the first 10 weeks of the 13-week study, no difference from controls was observed during the 4-week study. Animals on the C80 wax diets had increased food intake during weeks 1–4 of both studies. Increases in the absolute and relative (to body weight) weights of liver, spleen and mesenteric lymph nodes were observed with some of the mineral hydrocarbons at both 4 and 13 weeks. The liver weights were statistically significantly increased in the group given N15H low-viscosity oil at 4 weeks and in those given N15H and low-melting-point paraffin wax at 13 weeks. The weights of the mesenteric lymph nodes were increased in the groups given low-melting-point paraffin wax, C80 wax and N15H oil at 4 and 13 weeks and in those given N70H, but not the P70H medium-viscosity oil, at 13 weeks. At 4 weeks, spleen weights were increased only with the N70H and P70H medium-viscosity oils, while at 13 weeks increases in spleen weights were observed only with the low-melting-point paraffin wax, C80 wax and N15H oil. The weights of the kidney, heart and brain were not affected by treatment. Significant histopathological alterations were found in the cervical and mesenteric lymph nodes, liver, heart and small intestine. The incidence and severity of the findings were more marked at 13 than at 4 weeks and were associated primarily with ingestion of low-melting-point paraffin wax. Histiocytosis was seen in the lymph nodes of rats in all groups except those receiving N70H medium-viscosity oil for 4 weeks or P70H medium-viscosity oil for either 4 or 13 weeks. The incidence, severity (progression to individual cell necrosis) and onset were greater in mesenteric than in cervical nodes and in proximal mesenteric than in distal mesenteric nodes. The combined incidence and severity scores indicated a rank order of low-melting-point paraffin wax > C80 wax, N15H oil > N70H oil > P70H oil. Other changes identified at 13 weeks in nearly all (6–8/8) rats receiving low-melting-point paraffin wax in the diet included: hepatic focal necrosis with inflammation, periportal vacuolation and granuloma; and focal inflammation of the mitral valve of the heart and macrophage accumulation, particularly in Peyer patches of the small intestine. Periportal vacuolation was also observed in the livers of 7/8 rats receiving dietary C80 wax; vacuolation of macrophages was found in the lamina propria of the small intestine; granuloma of lower severity was seen in the livers of 7/8 rats; and focal inflammation of the cardiac mitral valve was recorded in 3/8 rats receiving N15H low-viscosity oil. Calcification of the renal medulla was exacerbated at both 28 and 90 days by dietary treatment with the white oils and marginally by treatment with the waxes.
No hydrocarbons with carbon numbers between C16 and C56 were detected in urine samples. The hydrocarbon profiles of faecal extracts were very similar to those of the corresponding mineral hydrocarbon materials, but with small differences for some materials, suggesting selective uptake and/or metabolism. A distortion of the faecal hydrocarbon profile was observed in extracts from the faeces of rats fed the C80 wax (loss of hydrocarbons C47–C56) and N15H oil (loss of hydrocarbons below C22). A very slight change was observed in faecal extracts from rats fed low-melting-point paraffin wax (loss of hydrocarbons from both the high and low ends of the carbon range). The authors indicated that apparent loss of alkanes with high carbon numbers was due to poor solubility during extraction and analysis. No significant changes were observed in the hydrocarbon profile in faecal extracts from rats consuming the N70H or P70H medium-viscosity mineral oils. Mineral hydrocarbon was detected in samples of small intestine, heart and kidney from all treated rats and in the livers of rats given low-melting-point paraffin wax or C80 wax, the only treatments for which this tissue was analysed. In addition, mineral hydrocarbon was detected in mesenteric lymph nodes of rats from all treated groups except that given P70H oil, and in cervical lymph nodes and aortic blood of rats treated with low-melting-point paraffin wax or C80 wax. The extent of accumulation of mineral hydrocarbons in tissues ranged from 0.01% to nearly 3%; the tissues with the greatest accumulation of hydrocarbon were not those with the greatest likelihood of histopathological alterations. Qualitative analysis of the carbon number range of the accumulated mineral hydrocarbons in tissue samples from rats treated with low-melting-point paraffin wax or C80 wax suggested that material in the alkane range C20–C35 was preferentially retained, confirming observations from previous studies (Cunninghame et al., 2001).
Parallel studies were conducted to assess the long-term toxic and carcinogenic effects of P70(H) mineral oil (medium and low viscosity, class I) and P100(H) mineral oil (high viscosity) in male and female Fischer 344 rats after 2 years’ administration in the diet. The study design also included an evaluation of the reversibility or persistence of the biological effects associated with 12 months’ exposure, after a 12-month recovery period. The studies were conducted in compliance with US Food and Drug Administration Good Laboratory Practice Regulations and the OECD Principles of Good Laboratory Practice.
Five groups were used for each study: a control group and groups given concentrations in the diet corresponding to a dose of 60, 120, 240 or 1200 mg/kg bw per day. The concentrations in the diet were adjusted to achieve a constant target dose throughout the study. Groups of 50 male and 50 female animals were used in the main (24-month) study, with additional groups of 30 male and 30 female animals for the reversibility phase (treatment for 12 months followed by 12 months on control diet). Of the 30 animals of each sex per group in the reversibility phase, 10 animals of each sex per group were killed after the 12-month feeding period.
Additional small groups of at least five female animals were added to evaluate tissue hydrocarbon concentrations at 3, 6, 12, 18 and 24 months in both phases of the study. The parameters investigated included body weight, food consumption, clinical observations, serum chemistry, haematology, ophthalmology, urine parameters and organ weights, including mesenteric lymph nodes. Analyses for mineral hydrocarbon were performed on liver, kidneys, mesenteric lymph nodes and spleen from female animals. Detailed histopathological examination of 48 tissues, including the liver, spleen, mesenteric and mandibular lymph nodes, Peyer patches, kidney and bone marrow, was conducted for all animals in the control group and at the highest dose in the main (2-year) study and at the 12-month sacrifice. From animals at 60, 120 or 240 mg/kg bw per day in the main study, only the lungs, liver, mesenteric lymph nodes, spleen and kidneys were examined; the mesenteric lymph nodes and livers of animals in all groups in the recovery study were also examined. Immune function was not tested, but standard end-points considered to reflect immune function (i.e. total and differential leukocyte count, albumin:globulin ratio, the weights and histological appearance of the thymus, spleen and mesenteric lymph nodes, histopathological evaluation of Peyer patches and bone-marrow cellularity) were assessed.
Administration of either oil to Fischer 344 rats in the diet for 24 months did not affect survival. The small decrease in survival rate among females at the high dose of P100H oil, starting at 15 months of the study, was within the rates of other control groups and was considered by the authors not to be biologically significant. No treatment-related effects were seen on clinical signs, body weight, food consumption, food conversion efficiency, ophthalmic, haematological, serum chemical or urinary parameters, and no treatment-related changes were seen at gross necropsy. Dietary administration of both oils was associated with increased weight of mesenteric lymph nodes and increased grade of infiltrating cell histiocytosis; increased incidence and grade of vacuolation of periportal hepatocytes; increased incidence of combined cystic degeneration or angiectasis of the liver of male rats, with no dose–response relationship; and a quantifiable, reversible accumulation of mineral hydrocarbons in the liver to a similar level regardless of dose but dependent on the type of mineral oil. The effects were more marked in rats treated with P70H oil than with P100H oil, and there were sex-related differences in response. Details of these findings are described separately for each oil. Neither oil was carcinogenic in this assay.
Treatment for 24 months with medium-viscosity white mineral oil (P70 (H)) resulted in a dose-related, statistically significant increase in mean mesenteric lymph node weight, both absolute and relative to body and brain weight, at all doses in female rats and at the highest dose, 1200 mg/kg bw per day, in males. Mesenteric lymph node weights were also statistically significantly increased in males at the highest dose after 12 months’ feeding; this effect was no longer significant after the 12-month recovery period owing to an increased value for this parameter in the control group. No significant differences in mesenteric lymph node weights were observed in females after 12 months of treatment, although, after the 12-month recovery, a slight but significant increase was seen in females at the two higher doses, 240 and 1200 mg/kg bw per day. The grade, but not the incidence, of infiltrating cell histiocytosis of the mesenteric lymph nodes was increased in all treated groups after 24 months of treatment. In addition, there was a slight increase in the severity of this lesion in animals at the highest dose, the only group assessed, at the 12-month sacrifice. After the 12-month recovery period, these changes were apparent in all groups, at a severity greater than or comparable to that in the animals treated for 24 months.
A dose-related increase in the incidence and grade of vacuolation of periportal hepatocytes was observed in the livers of males in all treated groups. A smaller increase in grade only was seen in treated female rats, because the incidence and grade of portal vacuolation were already high in female, compared with male, controls. In males allowed to recover, the increased incidence and severity of vacuolation of periportal hepatocytes was still evident after 12 months without treatment. The investigators did not consider this indicative of an adverse effect but rather a marker of prolonged exposure to mineral oil. An increased incidence of combined angiectasis and cystic degeneration (focal sinusoidal dilatation) was also observed in all treated male groups compared with the control group at the 24-month sacrifice. This lesion was of minimal grade, and the incidence was similar in all treated groups. While the incidence of this lesion was not increased over that in controls in males allowed to recover for 12 months, data from the 12-month sacrifice were not available to assess the significance of this observation.
An increased incidence of mononuclear cell leukaemia was observed in treated females. This was not considered treatment-related, however, as the incidence in treated groups was not dose-related and was within the range for other control female Fischer 344 rats.
Histological analysis revealed quantifiable amounts of hydrocarbon in the livers of treated animals; the hepatic concentrations of mineral hydrocarbon in animals at the highest dose reached nearly a maximum within 3 months and increased slowly up to 24 months of treatment. The values for animals at 60, 120 and 240 mg/kg bw per day at 12 and 24 months were 1200–1500 µg/g tissue, which were similar to each other and were approximately 60% of those of animals at 1200 mg/kg bw per day (1800 and 2300 µg/g tissue, respectively). After cessation of treatment, the hepatic concentrations had dropped substantially by 6 months and had returned to background levels by 12 months. The amount of mineral hydrocarbon in the spleen and mesenteric lymph nodes of most treated animals at higher doses was below the practical limit of reliable quantification.
After 24 months of treatment with high viscosity white mineral oil (P100 (H)), statistically significant increases in mean mesenteric lymph node weights (absolute and relative to body weight and brain weight) were observed in females at 1200 mg/kg bw per day. This effect was not evident at lower doses or in male rats or after the recovery period. A marginal increase in the grade, but not the incidence, of infiltrating cell histiocytosis in the mesenteric lymph nodes was observed in all treated groups at 24 months. No clear dose–response relationship was apparent in the males, and no effects were observed at 12 months of treatment.
A slight increase in the incidence and grade of vacuolation of periportal hepatocytes was observed in the livers of treated animals of each sex in all treated groups compared with controls. An increase in the incidence of combined angiectasis and cystic degeneration (focal sinusoidal dilatation) was observed in all treated male groups, without a dose–response relationship, which was statistically significant in animals given 1200 mg/kg bw per day. This lesion was not apparent at the 12-month sacrifice. In view of the nature and severity of the response, the investigators did not consider the increased grade of vacuolation to be indicative of an adverse toxicological effect but rather a marker of prolonged administration of white oil.
Histopathological examination revealed an increased incidence of adenoma of the pars distalis of the pituitary in female rats at the highest dose (42% in controls and 60% at the highest dose). This finding was not considered treatment-related as the incidence was well within the range reported for other control female Fischer 344 rats.
Histological analysis revealed quantifiable amounts of hydrocarbon in the livers of treated animals; the hepatic concentrations of mineral hydrocarbon in animals at the highest dose reached nearly a maximum within 3 months (900 µg/g tissue) and increased slowly up to 24 months of treatment. The concentrations in the liver at 12 and 24 months were similar in all treated groups (800–900 and 1200–1400 µg/g tissue, respectively), suggesting that a steady-state level had been reached. After cessation of treatment, the hepatic levels had dropped substantially by 6 months and had returned to background levels by 12 months. Mineral hydrocarbon was found in mesenteric lymph nodes and spleen at some times. The concentrations in the spleen and mesenteric lymph nodes of most treated animals at the higher doses were below the practical limit of reliable quantification. Detectable residues of mineral hydrocarbon were not found consistently in the kidneys (Exxon Biomedical Sciences, 2001).
A medium-viscosity white mineral oil (mineral oil, medium- and low-viscosity, class I), a blend of equal quantities of eight commercially available paraffinic white mineral oils obtained from eight member companies of the Japan Liquid Paraffin Industry, was fed in the diet to Fischer 344 rats. The oils also complied with the requirements of the Japanese food additive standards and the Japanese pharmacopoeia. Five of the component white mineral oils had been derived from petroleum by acid treatment, and the other three had been derived by hydrotreatment. The physical properties of the blended mineral oil were intermediate between those of P70H and N70H.
Groups of 50 male and 50 female Fischer 344 rats were fed diets containing 2.5% or 5% of the composite medium-viscosity white mineral oil (equivalent to 1250 and 2500 mg/kg bw per day), continually for 104 weeks. Body weights and food consumption were measured throughout the study. At the end of the study, the animals were killed and blood samples were collected for haematological and clinical chemical measurements. A full necropsy was performed on all animals; the major organs were weighed, and a range of tissues, including liver, mesenteric lymph node, heart and spleen, were taken for histological examination.
The food consumption and body weights of animals of each sex given 5% mineral oil were slightly increased. The frequency of clinical signs, mortality and haemato-logical parameters were unaffected by treatment. In the group given 5%, the absolute weights of the liver and kidney were increased in males and the absolute and relative weights of the submaxillary gland were reduced in females. The increased absolute organ weights were attributed to the slightly increased body weights of males at this concentration. The absolute and relative weights of the heart and spleen were unaffected by treatment. A variety of tumours developed in all groups, including the control group, but all the neoplastic lesions were histologically similar to those known to occur spontaneously in Fischer 344 rats, and no statistically significant increase in the incidence of any tumour type was found for either sex in the treated groups. An increased grade of small granulomatous foci of macrophages was observed in the mesenteric lymph nodes of both sexes at 2.5 and 5% in comparison with the respective control groups (Shoda et al. 1997).
In a comparison of the effects of dietary administration of low melting-point paraffin wax on Kupffer-cell function and morphology in female Fischer 344 and Sprague-Dawley rats, the animals received either control diet or a diet containing 2% low melting-point paraffin wax for 60 days in two separate studies. In the first study, blood was collected for analysis of serum enzymes (alanine and aspartate aminotransferases and gamma-glutamyl transpeptidase) and total leukocyte and neutrophil counts, and the liver was collected for histopathological examination and determination of its wax content. Kupffer cells were isolated from the liver and assayed for production of superoxide anion, nitrite ion, tumour necrosis factor-alpha (TNF-alpha and leukotriene B4, phagocytic activity and morphology.
Mean body weights were not affected by consumption of diets containing low melting-point paraffin wax. In treated female Fischer 344 rats, the serum activity of the liver enzymes was increased and the total leukocyte and neutrophil counts were significantly higher than those of untreated controls. No such differences were seen in Sprague-Dawley rats. The concentration of mineral hydrocarbon in the liver of treated Fischer 344 rats was 3.6 ± 0.65 mg/g liver, but none was detected in the livers of untreated control Fischer 344 rats or treated or untreated Sprague-Dawley rats. Microgranulomas were observed in the livers of all treated Fischer 344 rats but in only one treated Sprague-Dawley rat, in addition to small areas of focal necrosis, lymphoid cell aggregates and increased frequency of cytoplasmic vacuoles.
Over half the Kupffer cells isolated from Fischer 344 rats treated with low melting-point paraffin wax contained large, irregularly shaped vacuoles, and another 10–20% had numerous smaller vacuoles. The vacuoles were membrane-associated, suggesting phagocytic uptake. Similar vacuoles were detected only rarely in Kupffer cells isolated from Sprague-Dawley rats. The indices of Kupffer cell function—production of superoxide anion and nitric oxide and phagocytic activity—were increased, and TNF-alpha and leukotriene B4 production were decreased in treated Fischer 344 rats compared with untreated controls. No significant changes in these functions were observed in Kupffer cells isolated from treated Sprague-Dawley rats (Hoglen et al., 1998).
Serum immunooglobin (Ig) levels were measured in samples from Fischer 344 and Sprague-Dawley rats that had been fed diets containing 0.0, 0.2 or 2.0% low melting-point paraffin wax for 90 days (see section 2.2.2), in order to determine whether the differences in response to mineral hydrocarbons in these two rat strains were due to a difference in immunological response. The study was conducted according to the Good Laboratory Practices for Nonclinical Laboratory Studies of the Food and Drug Administration in the USA. Serum immunoglobin classes (IgG, IgM, IgA) and IgG subclasses (IgG1, IgG2a, IgG2b, IgG2c) were measured in frozen serum samples and related to total serum protein in rats dosed for 30 and 60 days (control and high dose) and for 90 days (control, low and high dose). Treatment did not affect serum immunoglobin levels in Sprague-Dawley rats but increased IgA, IgM and IgG2c and decreased IgG and IgG2a levels in Fischer 344 rats. The changes were dose-related for IgG and IgG2c and both dose- and time-related for IgA, IgM and IgG2a (Burleson, 2001).
The humoral immune response to the T-dependent antigen, sheep erythrocytes, was evaluated in female Fischer 344 and Sprague-Dawley rats given P15 mineral oil (medium- and low-viscosity, class III) in the diet for 90 days. The study was conducted in compliance with the Good Laboratory Practice Regulations of the Food and Drug Administration in the USA and was divided into two phases. In phase I, groups of eight female Fischer 344 rats were fed diets containing P15 mineral oil at 0%, 0.02% or 2.0% and groups of eight female Sprague-Dawley rats were fed diets containing 0%, 1.0% or 2.0% for 90 days. In phase II of the study, groups of eight female Fischer 344 were fed diets containing P15 mineral oil at 0%, 0.02% or 2.0% and Sprague-Dawley rats were fed diets containing 0%, 1.0% or 2.0% for 120 days, followed by a 30-day recovery period. A positive control group for each strain received cyclophosphamide at a dose of 50 mg/kg bw intraperitoneally on each of 4 days after sensitization (see below). Four days before sacrifice, the rats in phase I were sensitized by intravenous injection of 2 x 108 sheep erythrocytes. On day 91, the day of sacrifice, spleen-cell suspensions were prepared for evaluation of the IgM response with a modified haemolytic plaque assay. The animals in phase II were sensitized with 100 µg of dinitrophenol–human serum albumin intravenously on day 85, and on day 91 a blood sample was taken for determination of the IgM antibody titres to this antigen. Depending on the results of the assay for immune function in phase I, a decision was taken whether to continue phase II past day 91. The sensitization procedure and blood collection were repeated on days 115 and 121, respectively, for determination of secondary antibody (IgG) to dinitrophenol–human serum albumin. The treated diet was discontinued at this time. Serum IgM and IgG antibody titres were determined by enzyme-linked immunosorbent assay. On day 147, the animals received 2 x 108 sheep erythrocytes intravenously, and on day 151 they were killed and their spleens removed for determination of the primary IgM response to sheep erythrocytes. Body weights and food consumption were monitored for 90 days during each study phase, and, at sacrifice, the weights of the brain, liver, spleen and lymph nodes were recorded. The liver and mesenteric lymph nodes with associated adipose tissue from all animals and only the spleens from Sprague-Dawley rats killed at day 91 in the phase II study were examined histopathologically.
A diet containing P15 mineral oil for 90 days had no consistent effect on body weight or food consumption in female Fischer 344 or Sprague-Dawley rats in phase I or II. In the Sprague-Dawley rats, the absolute and relative liver weights were statistically significantly increased over those of controls, by approximately the same amount after treatment at 1% and 2% in both phase I and phase II. Lymph nodes (type not specified) showed a slight, non-significant, dose-related increase in weight over those of controls. At 90 days, the absolute and relative weights of the liver in Fischer 344 rats at 0.02% and 2% in the diet were statistically significantly increased in a dose-related manner; and the weights of the spleen and mesenteric lymph nodes of animals at the highest concentration were also statistically significantly increased compared with controls. After 120 days’ treatment and 30 days’ recovery, the livers of the Fischer 344 rats were still heavier than those of controls, but only the difference in relative weights was statistically significant; the absolute and relative spleen weights remained statistically significantly increased in rats at the highest concentration. The weights of mesenteric lymph nodes after the recovery period were not provided.
Granulomatous inflammation of the mesenteric lymph nodes was noted in nearly all control and treated Fischer 344 rats (6/8 controls, 8/8 at the low concentration and 8/8 at the high concentration); the grade of the lesion was higher at the higher dietary level than at the lower or in controls, and the granulomatous inflammation extended to the attached mesenteric adipose tissue. The incidence and severity of granulomatous inflammation of the liver were also greater in rats at the higher concentration (1/8 at the low concentration and 7/8 at the high concentration, with none in controls). After 120 days’ treatment and 30 days’ recovery, granulomatous inflammation of the mesenteric lymph nodes was observed in all control and treated animals, with slightly greater severity in rats at the lower concentration and no change in severity in animals at the higher concentration or in controls from that in Fischer 344 rats at the 91-day sacrifice. Granulomatous inflammation of the liver found after longer exposure and after the recovery period was also found after the 91-day sacrifice. The spleens of Fischer 344 rats were not available for examination. In the Sprague-Dawley rats killed at 91 days, granulomatous inflammation of the mesenteric lymph nodes was seen, which was generally of minimal severity, in 1/16 controls, 15/16 at the lower concentration and 15/16 at the higher; the incidence and severity of this finding were similar in the groups at 1% and 2% in the diet. No treatment-related effects were seen in the spleen or liver.
Consumption of a diet containing mineral oil for 90 days had no effect on the spleen IgM antibody response in Sprague-Dawley rats, as assessed in the plaque assay, although the number of spleen cells was increased in both treated groups, statistically significantly at 1% in the diet. The Fischer 344 rats showed a dose-related decrease (40%) in the antibody-forming cell response, expressed per 106 spleen cells, which was statistically significant at the higher concentration; no effect was seen when the data were expressed on the basis of total spleen activity, owing to the significant increase in spleen cell number observed at the higher concentration. Increased spleen weights were also seen in these animals. No flow cytometry data were provided to allow identification of the specific spleen cell population affected; in addition, no histopathological examination of the spleen was conducted in Fischer 344 rats to find an explanation for the increase in spleen cell number in animals at the higher concentration. After 120 days’ exposure and 30 days’ recovery, the responses for all end-points (spleen weight, number of spleen cells, IgM antibody-forming cells per 106 spleen cells, IgM antibody-forming cells per spleen x 103) were similar to those observed at 90 days, except that the control value for the number of antibody-forming cells per 106 spleen cells was lower, and consequently the decrease in this parameter with treatment was not statistically significant. The positive control, cyclophosphamide, markedly suppressed the spleen IgM antibody response. Owing to the lack of effect of mineral oil on the spleen IgM T-dependent antibody-forming cell response in Sprague-Dawley rats, the primary IgM and secondary IgG responses to dinitrophenol–human serum albumin were not assessed in this strain. Treatment had no effect on the IgM response at 90 days, and a small, dose-related but non-significant effect on the IgG response was seen at 120 days in this strain. The positive control markedly reduced both the IgM and the IgG response (ImmunoTox, 2001).
The effects of low melting-point paraffin wax on cell infiltrates in the liver, mesenteric lymph node and selected cardiac mitral valves were evaluated in Fischer 344 and Sprague-Dawley rats. Replicate sections of liver, mesenteric lymph nodes and cardiac mitral valves from rats in the 90-day study described above were stained either with haematoxylin and eosin or immunohistochemically to demonstrate the presence of cells expressing CD3 (T cells), CD45RA (B cells), CD8 (CD8a, suppressor/cytotoxic T cells) or ED2 (resident macrophages) antibodies and lysozyme (activated macrophages). In addition, sections of the liver were evaluated by electron microscopy to assess Kupffer cell activation and the nature of the accumulated lipid material.
Increased grades of hepatocellular and Kupffer cell vacuolation were apparent in both rat strains in response to a diet containing low melting-point paraffin wax. The earliest observed change in the Fischer 344 rats was small, mostly centrilobular vacuoles at day 30, which increased in size and distribution with duration of treatment. Lipid-like pseudocysts and granulomatous inflammation associated with the lipid-like material developed with increasing grade as a function of duration of treatment. Occasional single-cell necrosis was observed in hepatocytes, both in the presence and absence of inflammatory infiltrates. Distinct crystalloid structures were present within lysosomes of many Kupffer cells and hepatocytes. The lipid-like pseudocysts were considered to be consistent with accumulation of the lipid-like material in crystalline form. Many activated (lysozyme-positive) Kupffer cells were observed. The granulomas were characterized by a predominantly centrilobular–midzonal distribution and were composed of large epithelioid ED-2-negative (non-resident), lysozyme-positive macrophages. Hepatic granulomatous inflammation was accompanied by a T-cell (lymphocyte) response (CD3+/CD4+/CD8+). Vacuolar changes were also observed in hepatocytes of Sprague-Dawley rats but to a much lesser extent than in Fischer 344 rats. In Sprague-Dawley rats, activation of Kupffer cells was much less evident and granuloma formation was observed in only one treated animal. In contrast to Fischer 344 rats, the crystalloid structures within lysosomes of hepatocytes and Kupffer cells were not observed. A higher threshold number of ED2-positive Kupffer cells was seen in control Sprague-Dawley rats than in control Fischer 344 rats.
The primary change in mesenteric lymph nodes of both strains of rat in response to a diet containing low melting-point paraffin wax was formation of granulomas composed of epithelioid macrophages. The severity and incidence of the changes was greater in the Fischer 344 rats. The granulomas were initially small and subcapsular and cortical in location. With increasing dose and duration of treatment, they increased in size and ultimately coalesced to occupy most of the nodal cortex and parts of the medulla. Macrophages of the granulomas were predominantly ED2-negative and lysozyme-positive, consistent with the non-resident nature of the cells. Resident lymphocytes of the node surrounded the granuloma; most were CD3+, with equal numbers of CD4+ and CD8a+ cells.
The inflammatory changes in the mitral heart valve of Fischer 344 rats were slight and consisted of phenotypically mixed cell populations. They were also observed in one control animal, leading the investigators to conclude that the observation was of questionable significance. Only limited numbers of tissues were available for examination. The authors suggested that the data were consistent with a difference in pharmacokinetics between the strains, related to metabolic capacity, resulting in saturation of enzyme systems and accumulation of lipid-like crystalloids, cell disruption and inflammation (Francke et al., 2001).
Mineral hydrocarbons found in food include both added mineral oils and naturally occurring hydrocarbons. The former category includes white mineral oils (various viscosities), paraffin waxes (low melting-point), microcrystalline wax (high melting-point) and petroleum jelly, which are used in coating and packaging materials, during bakery and confectionery manufacture and in grain and seed dust control. These petroleum-based products are mixtures of saturated hydrocarbons with carbon numbers of 18–50; however, only hydrocarbons with a chain length 32 are of concern because they can be absorbed in the gastrointestinal tract. Many plants and animals also synthesize long-chain hydrocarbons, and humans are thus exposed to these naturally occurring hydrocarbons.
The dietary intakes of mineral oils of various viscosities and paraffin wax (low melting-point) and microcrystalline wax (high melting-point) were assessed; the intake of petroleum jelly was not included in this assessment.
Data on mineral oils from a number of studies conducted in Europe and USA were submitted by CONCAWE, the European Wax Federation and the American Petroleum Institute and included national estimates for the countries of the European Union, the United Kingdom and the USA. Data from food basket surveys, household economic surveys or sales were provided for the European Union, and data from individual dietary records were provided for the United Kingdom and the USA. The estimated dietary intakes of various categories of mineral hydrocarbons were calculated on the basis of their concentrations (added, migrated or naturally existing) in a number of food groups and combined with food consumption data from food balance sheets for countries of the European Union and national surveys, such as the National Diet and Nutrition Survey in the United Kingdom (adults, 1986–87; preschool children, 1992–93) and the Continuing Survey of Food Intake by Individuals (1989–92) in the USA.
The main use of mineral oils in foods (both direct and indirect) is in the form of polystyrene, which represents 74% of total sales in the USA. Paraffin wax is used primarily in corrugated cartons as a saturating wax (54% of total sales), in paper cups and containers for cold drinks (22%) and in wax-coated, flexible packaging (10%). Microcrystalline wax is used primarily in wax-laminated flexible packaging (23%), curtain coating for corrugated cartons (22%) and flexible packaging with wax-coated substrates (22%).
The potential concentrations of white mineral oils, paraffin wax and microcrystalline wax in various categories of foods in the United Kingdom and the USA, based on the amounts added to foods and studies of migration from coatings and packaging materials are presented in Tables 2–4. These concentrations were used to derive the intake estimates. The data show that: (1) the principal dietary sources of mineral oil in the United Kingdom were bread divider oils (high- and low-viscosity oils) and grain dust control (low-viscosity oils) (Food Chemical Risk Analysis, 2001), while those in the USA were fruit and vegetable coatings (low-viscosity oils), confectionery (low-viscosity oils), grain dust control (low-viscosity oils), and bakery pan release oils (high-viscosity oils) (Reich et al., 1998); and (2) the sole dietary source of paraffin wax in Europe was wax paper packaging, and the principal source of microcrystalline wax was use as a confectionery glazing agent (Food Chemical Risk Analysis, 2001), while the main sources of both waxes in the USA were fruit and vegetable coatings and flexible packaging (Reich et al., 1998).
In addition, data were available from other surveys on the concentrations in five major food groups.
Table 2. Potential concentrations (mg/kg) of mineral oils in food in the United Kingdom
|
Application and medium |
Viscosity at 40 °C (mm2/s) |
Comment |
|
|
< 65a |
> 65b |
||
|
Polystyrene |
|||
|
Drink containers |
– |
< 1 |
|
|
Milk containers |
– |
< 1 |
|
|
Individual milk/cream containers |
– |
< 5–93 |
Occasional use |
|
Polyethylene |
|||
|
Containers |
– |
Not significant |
|
|
Delicatessen meats |
– |
7 |
|
|
Bakery release agent or divider oils |
|||
|
Bread |
243 |
243 |
Very limited use |
|
Confectionery |
|||
|
Sugar confectionery |
250 |
250 |
Now obsolete |
|
Chocolate |
50 |
50 |
Now obsolete |
|
Jute batching oil |
|||
|
Chocolate |
7 |
– |
|
|
Temporary sausage skins |
|||
|
Skinless sausages |
28 |
– |
|
|
Grain and seed de-dusting |
|||
|
Flours and cereals |
9–42 |
– |
|
|
Vegetables |
110 |
– |
|
From Food Chemical Risk Analysis (2001)
a Includes class III low-viscosity oils
b Includes high-viscosity and class I and II medium-viscosity oils
Table 3. Potential concentrations (mg/kg) of mineral waxes in food in the United Kingdom
|
Application and medium |
Concentration (mg/kg) |
Comment |
|
|
Paraffin wax |
Microcrystalline wax |
||
|
Flexible wax-coated packaging |
|||
|
Sliced cheese |
7 |
7 |
|
|
Sliced meats |
< 1 |
< 1 |
|
|
Biscuits |
105 |
105 |
Obsolete |
|
Cereals |
105 |
105 |
|
|
Bread |
29 |
29 |
|
|
Sugar |
52 |
52 |
|
|
Cartons with saturating wax |
|||
|
Milk/cream |
< 1 |
– |
No direct contact |
|
Cheese coating |
|||
|
e.g. Gouda, Edam cheese |
4.4 |
4.4 |
|
|
Chewing-gum base |
Not significant |
Not significant |
|
|
Fruit coating |
|||
|
Tropical fruit peel |
– |
200 |
No direct contact |
|
Poultry de-feathering |
|||
|
Duck meat |
9 |
9 |
|
|
Confectionery glazing agent |
|||
|
Chewing-gum |
– |
200 |
|
|
Non-chocolate confectionery |
– |
200 |
|
From Food Chemical Risk Analysis (2001)
Table 4. Potential concentrations of mineral oils and waxes in food in the USA
|
Food group |
Source |
Concentration |
|
Cheese |
Coating: paraffin wax |
2.3 |
|
Edam and Gouda |
Coating: microcrystalline wax |
3.1 |
|
Barley |
Dust control: mineral oil |
40 |
|
Corn bran |
Dust control: mineral oil |
33 |
|
Corn endosperm |
Dust control: mineral oil |
33 |
|
Oats |
Dust control: mineral oil |
40 |
|
Oat bran |
Dust control: mineral oil |
40 |
|
Rice rough |
Dust control: mineral oil |
40 |
|
Rice bran |
Dust control: mineral oil |
40 |
|
Rice milled |
Dust control: mineral oil |
40 |
|
Rye rough |
Dust control: mineral oil |
40 |
|
Rye flour |
Dust control: mineral oil |
9 |
|
Wheat bran |
Dust control: mineral oil |
42 |
|
Wheat flour |
Dust control: mineral oil |
9 |
|
Wheat rough |
Dust control: mineral oil |
42 |
|
Soybean flour |
Dust control: mineral oil |
25 |
|
Sunflower seeds |
Dust control: mineral oil |
40 |
|
Corn oil |
Dust control: mineral oil |
600 |
|
Sunflower oil |
Dust control: mineral oil |
110 |
|
Soybean oil |
Dust control: mineral oil |
110 |
|
Pan breads, rolls |
Pan release oils: mineral oil |
210 |
|
Fruits and vegetables |
Edible coating: paraffin wax |
44 |
|
Edible coating: microcrystalline wax |
44 |
|
|
Edible coating: mineral oil |
160 |
From Reich et al. (1998)
Bakery products (bread and rolls):
Mineral oils may occur not only as residues in flour and in vegetables from de-dusted grain but may also be added to breads and rolls directly as divider oils or as lubricants to prevent bread from adhering to the baking pan. Use of mineral oils in the bakery industry in the United Kingdom is now quite limited as compared with their wide use in the 1960s (Food Chemical Risk Analysis, 2001), as the British baking industry voluntarily withdrew the use of mineral oils as pan-release agents. Furthermore, use of white mineral oils in the baking industry in several countries of the European Union is now restricted by national legislation.
Few data were available on mineral oil residues in commercially produced breads and rolls. The total concentration of mineral oils in loaves in the United Kingdom in one study ranged from 220 to 490 mg/kg. A similar study on mineral oil residues in bread in Germany showed 330 mg/kg in the bottom crust and < 1 mg/kg in the top crust and centre of the bread (Food Chemical Risk Analysis, 2001).
The combined maximum permitted level for white mineral oils and petroleum jelly in baked products in the USA is 1500 mg/kg (21 CFR 172.878 and 172.880). Although vegetable oils and sometimes animal fats are also used as divider oils, pan-release oils and trough grease, the relative market shares in these uses are unknown. It has been reported that divider oils are not absorbed by dough in measurable amounts (Reich et al., 1998).
A survey conducted by the American Bakers Association in 1998 (Reich et al., 1998) showed that the overall average total concentration of mineral hydrocarbons in bread from pan-release oils and trough grease was 533 mg/kg. Some bread may contain only mineral oil, some only petrolatum jelly, some both kinds, and some may contain no mineral oils from the baking process.
Fruits and vegetables:
Mineral oil-based coatings are used on specific types of fruits and vegetables to prevent loss of moisture, act as a carrier for fungicides, protect the commodities from bruising and add various degrees of sheen. Fruits and vegetables are coated only when intended for fresh consumption. In computing the concentrations of mineral hydrocarbons in foods resulting from use of mineral oil-based coatings, it was conservatively assumed that all the relevant fruits and vegetables are coated with a standard composite of white mineral oil, petrolatum jelly, paraffin wax and microcrystalline wax. The residual concentrations of these mineral oils in standard coatings would then be 160, 44, 44 and 44 mg/kg, respectively. This assumption therefore resulted in a greatly exaggerated estimate of intake from fruit and vegetable coatings.
Grains and oil seeds:
Grain and oilseed products can contain mineral oil residues as a result of use in grain de-dusting, which is performed to reduce the free dust in stored grain and thus prevent explosions in grain elevators.
It was assumed that mineral oil is applied to all grains and soya beans at the maximum permitted level in the USA of 200 mg/kg (21 CFR 172.878), keeping in mind that the less expensive soya bean oil is also used in some grain elevators. Reich et al. (1998) showed that the concentrations of mineral oil on wheat grain stored for 6 months after spraying with low-viscosity white mineral oil at 230 mg/kg was 180–250 mg/kg. As low-viscosity mineral oils are quite volatile, losses are expected at the high temperatures used in food processing. In the study of Reich et al., no detectable mineral oil residues were found in finished bread products from milled and baked mineral oil-treated wheat grains. When mineral oil was added at 200 or 500 mg/kg to wheat flour, which was then made into dough and baked into bread, the concentration of mineral oil was reduced by approximately 80%. As a conservative measure, an 80% reduction factor was recommended for use in estimating the residual content of mineral oil in baked products and breakfast cereals when mineral oil-treated grains were the raw material.
Except where other estimates exist, such as for maize, soya beans and wheat meals, 20% of 200 mg/kg (i.e. 40 mg/kg) was used as the mineral oil concentration in other grain products, namely rice, barley, oats and rye.
Confectionery:
White mineral oils and petrolatum jelly are approved for use as release agents and as sealing and glazing agents in the manufacture of candy, chocolate and marshmallow-type items in the USA, with a maximum permitted level of 200 mg/kg (21 CFR 172.878 and 172.880).
Foods in contact with wax packaging materials:
Flexible wax packaging includes wax paper (e.g. paper cups for cold drinks), wax-coated cereal liners and wax-coated paperboard retail packaging. Reich et al. (1998) used the consumption factor proposed by the Food and Drug Administration in the USA (the proportion of consumers in the total population) to determine dietary intake of paraffin wax and microcrystalline wax from flexible wax packaging, and these are shown in Figure 1. The overall concentration of paraffin wax in crackers, cereals, meats, cheeses, milk and beverages (alcoholic, acidic and aqueous) due to contact with wax packagings was estimated to be 17 mg/kg, and that of microcrystalline wax was 9.7 mg/kg.
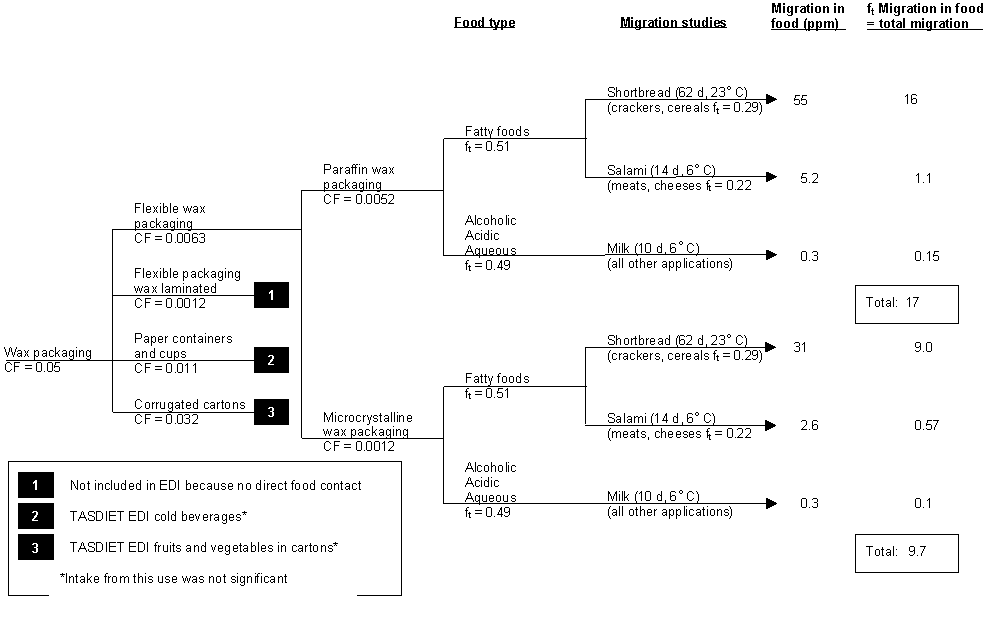
Figure 1. Estimated daily intake (EDI) derivation of paraffin and microcrystalline waxes from wax packaging
Hydrocarbons, or long-chain alkanes, are synthesized by a large number of edible plants and animals, and the background levels of these naturally occurring alkanes in food were investigated in order to estimate their contribution to overall mineral hydrocarbon intake. Like mineral hydrocarbons, biogenic alkanes have various structures, which can be divided into three categories: normal paraffins (straight-chain alkanes), isoparaffins (branched-chain alkanes) and cycloparaffins (cyclic alkanes). All these structures are saturated, i.e. without double bonds. In this monograph, the concentrations of the various naturally occurring hydrocarbons in foods were combined, although the range C18–C32 was used to limit the naturally occurring hydrocarbons to those of potential concern..
Reich et al. (1997) compiled a comprehensive list of the concentrations of naturally occurring hydrocarbons in about 180 varieties of plant and animal foods, and this data set was used to estimate the dietary intakes of naturally occurring hydrocarbons in the populations of the countries of the European Union, the United Kingdom and the USA. A simplified version of this data set, based on FAO Food Balance Sheet categories, is presented in Table 5.
Table 5. Concentrations of naturally occurring hydrocarbons in plant and animal foods
|
Food |
Concentration (mg/kg) |
|
Wheat |
15 |
|
Maize |
1.4 |
|
Beans |
3.1 |
|
Ground nuts |
13 |
|
Sunflower seed |
63 |
|
Rape, mustard, cotton seed |
44 |
|
Sesame seed |
24 |
|
Olives |
88 |
|
Oil crops, other |
44 |
|
Soya bean oil |
17 |
|
Groundnut oil |
34 |
|
Sunflower seed oil |
120 |
|
Vegetable oil, others |
59 |
|
Olive oil |
60 |
|
Maize germ oil |
30 |
|
Vegetables |
4.6 |
|
Oranges |
14 |
|
Lemons |
14 |
|
Grapefruit |
17 |
|
Citrus, others |
14 |
|
Apples |
140 |
|
Grapes |
3.1 |
|
Wine |
3.1 |
|
Beef |
0.15 |
|
Mutton |
0.15 |
|
Pork, fat |
40 |
|
Pork, lean |
6.8 |
|
Poultry meat |
0.03 |
|
Meat, other |
0.15 |
|
Animal fats |
27 |
|
Butter, ghee |
87 |
|
Cream |
87 |
|
Freshwater fish |
0.85 |
|
Marine fish |
46 |
|
Crustaceans |
0.17 |
|
Molluscs, other |
0.17 |
|
Milk, skim |
87 |
|
Milk fat, estimate in milk |
4.4 |
From Food Chemical Risk Analysis (2001)
Mineral oils of various viscosities were the major contributors to the total dietary intake of all mineral hydrocarbons in both Europe (90%) and the USA (50%). Paraffin wax and microcrystalline wax contributed very little to the total intake in both regions (Table 6).
Table 6. Average estimated intakes of mineral hydrocarbons in the populations of the United Kingdom (UK) and the USA
|
Food use |
Average intake (mg/kg bw per day) |
|||||||||
|
Paraffin wax |
Microcrystalline wax |
Mineral oil |
Total |
|||||||
|
< 65 mm2/sa |
> 65 mm2/sb |
|||||||||
|
UK |
USA |
UK |
USA |
UK |
USA |
UK |
USA |
UK |
USA |
|
|
Direct |
||||||||||
|
Fruit and vegetable coatings |
0.02 |
– |
0.02 |
– |
0.07 |
– |
– |
– |
0.11 |
– |
|
Confectionery |
– |
– |
– |
0.001 |
0.08 |
– |
– |
– |
0.08 |
0.001 |
|
Bakery |
– |
– |
– |
– |
– |
0.13 |
0.17 |
0.19 |
0.17 |
0.32 |
|
Grain and oil, de-dusting |
– |
– |
– |
– |
0.10 |
0.08 |
– |
– |
0.10 |
0.08 |
|
Others |
– |
0.001 |
– |
– |
– |
– |
– |
– |
– |
0.001 |
|
Indirect |
||||||||||
|
Wax packaging |
0.005 |
0.06 |
0.001 |
0.009 |
– |
– |
– |
– |
0.006 |
0.07 |
|
Plastic packaging |
– |
– |
– |
– |
– |
– |
0.001 |
0.001 |
0.001 |
0.001 |
|
Total mineral hydrocarbon intake |
0.03 |
0.06 |
0.02 |
0.01 |
0.25 |
0.21 |
0.17 |
0.19 |
0.47 |
0.47 |
From Reich et al. (1998); Food Chemical Risk Analysis (2001); UK, United Kingdom
a Viscosity at 40 °C; includes class III low-viscosity oils
b Viscosity at 40 °C; includes high viscosity and class I and II medium-viscosity oils
The average daily intakes of various categories of mineral hydrocarbons from different types of foods and various direct and indirect food uses in the United Kingdom and the USA are summarized in Table 6. In both countries, the average total intake of mineral carbons (excluding petroleum jelly) was estimated to be 0.47 mg/kg bw per day. Class III medium- and low-viscosity mineral oils accounted for 0.21 mg/kg bw per day in the United Kingdom and 0.25 mg/kg bw per day in the USA, whereas class I and II medium- and low-viscosity mineral oils and high-viscosity oil accounted for 0.18 and 0.19 mg/kg bw per day, respectively. The intake from paraffin wax was estimated to be 0.005 mg/kg bw per day in both countries; and the intake from microcrystalline wax was estimated to be 0.001 mg/kg bw/day in the USA (data for the United Kingdom were not available).
Data for intake in the United Kingdom were also given for consumers at a high percentile and for preschool children (Table 7). The mean, 90th and 97.5th percentile intakes, expressed per kilogram body weight, were significantly higher for children than for adults because of their lower body weight.
Table 7. Estimated intake of mineral oils by adults and preschool children in the United Kingdom
|
Mineral oil |
Intake (mg/kg bw per day) |
||
|
Mean |
90th percentile |
97.5th percentile |
|
|
White mineral oil |
|||
|
Adults |
0.39 |
0.68 |
0.91 |
|
Preschool children |
0.75 |
1.4 |
1.8 |
|
Paraffin and microcrystalline wax |
|||
|
Adults |
0.08 |
0.13 |
0.19 |
|
Preschool children |
0.23 |
0.44 |
0.64 |
|
Total |
|||
|
Adults |
0.47 |
0.16 |
0.22 |
|
Preschool children |
0.98 |
1.8 |
2.4 |
From Food Chemical Risk Analysis (2001)
(a) European Union
The average intakes of naturally occurring hydrocarbons by consumers in 15 countries of the European Union, estimated on the basis of food balance sheets, ranged from 0.33 mg/kg bw per day in the United Kingdom to 0.61 mg/kg bw per day in Germany. The European average was 0.47 mg/kg bw was day. The principal sources of intake were apples, wheat and dairy fat. Apples alone contributed about one-third of the total intake, and national intake levels were determined largely by consumption of this fruit (Food Chemical Risk Analysis, 2001).
(b) United Kingdom
The estimated average intake of mineral oils from naturally occurring hydrocarbons by adults in the United Kingdom (based on national food consumption data) was 0.25 mg/kg bw per day (Table 8), similar to the estimate of 0.33 mg/kg bw per day per capita calculated from FAO food balance sheets. Consumers at the 97.5th percentile could have an intake of up to 0.80 mg/kg bw per day. The mean intake of naturally occurring hydrocarbons by preschool children was 0.73 mg/kg bw per day, and that of consumers at the 97.5th percentile increased to 2.6 mg/kg bw per day (Table 8). The intake by children was higher than that by adults because of their lower body weight. The major source of intake was apples, followed by citrus fruits, cereals and dairy fat. The intake of naturally occurring hydrocarbons appeared to be determined largely by consumption of apples (Food Chemical Risk Analysis, 2001).
Table 8. Intakes of naturally occurring hydrocarbons by adults and preschool children in the United Kingdom
|
Food source |
Per capita intake (mg/kg bw per day) |
||
|
Average |
90th percentile |
97.5th percentile |
|
|
Adults |
|||
|
Brassicas |
0.004 |
0.007 |
0.011 |
|
Cereals |
0.034 |
0.056 |
0.071 |
|
Citrus |
0.015 |
0.046 |
0.087 |
|
Dairy fat |
0.031 |
0.060 |
0.085 |
|
Fish |
0.018 |
0.043 |
0.066 |
|
Grapes |
0.001 |
0.005 |
0.010 |
|
Meat |
0.002 |
0.004 |
0.006 |
|
Nuts |
0.002 |
0.007 |
0.018 |
|
Olives |
0.000 |
Few consumers |
Few consumers |
|
Apples |
0.13 |
0.37 |
0.64 |
|
Poultry |
0.000 |
0.000 |
0.001 |
|
Seeds/pulses |
0.002 |
0.004 |
0.006 |
|
Shellfish |
0.000 |
0.000 |
0.000 |
|
Vegetable oils |
0.008 |
0.018 |
0.031 |
|
Total for adults |
0.25 |
0.50 |
0.80 |
|
Preschool children |
|||
|
Brassicas |
0.004 |
0.012 |
0.019 |
|
Cereals |
0.043 |
0.076 |
0.10 |
|
Citrus |
0.060 |
0.18 |
0.37 |
|
Dairy fat |
0.10 |
0.20 |
0.31 |
|
Fish |
0.031 |
0.089 |
0.16 |
|
Grapes |
0.001 |
0.003 |
0.009 |
|
Meat |
0.003 |
0.006 |
0.011 |
|
Nuts |
0.005 |
0.017 |
0.050 |
|
Olives |
0.000 |
Few consumers |
Few consumers |
|
Apples |
0.43 |
1.2 |
2.2 |
|
Poultry |
0.000 |
0.000 |
0.002 |
|
Seeds/pulses |
0.004 |
0.009 |
0.014 |
|
Shellfish |
0.000 |
Few consumers |
Few consumers |
|
Vegetable oils |
0.014 |
0.031 |
0.048 |
|
Total for preschool children |
0.73 |
1.5 |
2.6 |
From Food Chemical Risk Analysis (2001)
(c) USA
The total estimated per capita intake of naturally occurring hydrocarbons from all foods was 0.19 mg/kg bw per day. The highest mean intakes of naturally occurring hydrocarbons for consumers of specific foods were from kale and rutabaga tops, but their contribution to per capita intake is small as these items are not consumed by a large proportion of the population. The next highest contributor to per capita intake was apples, which are widely consumed, so that apples contributed 34% of the total mean per capita daily intake (Reich et al., 1997).
In both the United Kingdom and the USA, the average total intake of the sum of white mineral oil, paraffin wax and microcrystalline wax from use in food (as direct and indirect food additives) was estimated to be 0.47 mg/kg bw per day, whereas the dietary intake of naturally occurring hydrocarbons from plant and animal foods was estimated to be 0.47 mg/kg bw per day for the population of the European Union, 0.25 mg/kg bw per day for that of the United Kingdom and 0.19 mg/kg bw per day for that of the USA. As naturally occurring hydrocarbons make a significant contribution to overall hydrocarbon intake in consumers in both the United Kingdom and the USA, the risk assessment of mineral oil residues from various food applications must include the dietary intake of naturally occurring hydrocarbons.
Estimates of gross intake of mineral hydrocarbons used in food tend to be overestimates because the proportion of the supply of a given food commodity that is affected is usually unknown and it is assumed that 100% of all foods contain mineral hydrocarbons. In one study in the United Kingdom (Food Chemical Risk Analysis, 2001), the gross intake level was adjusted by the actual proportion of mineral oil use derived from surveys. With this method, the gross mean intake of 0.47 mg/kg bw per day based on maximum use levels was reduced to 0.09 mg/kg bw per day (Table 9). In addition, some food uses of mineral oils are no longer common at the global level. For example, their use as pan-releasing agents is now restricted in Europe. Conversely, the intake of naturally occurring hydrocarbons may be underestimated because data on concentrations in all foods are not available.
Table 9. Intake of total added mineral hydrocarbons before and after adjustment for actual use in the United Kingdom
|
Intake (mg/kg bw per day) |
||||||
|
Adults |
Preschool children |
|||||
|
Mean |
90th |
97.5th |
Mean |
90th |
97.5th |
|
|
Gross total mineral hydrocarbons |
0.47 |
0.80 |
1.1 |
0.98 |
1.6 |
2.2 |
|
Use adjusted for total mineral hydrocarbons according to actual use |
0.09 |
0.15 |
0.21 |
0.19 |
0.31 |
0.41 |
From Food Chemical Risk Analysis (2001)
The intake of high-viscosity and classes I, II and III medium- and low-viscosity mineral oils that have migrated from coating and packaging materials was estimated to be 0.001 mg/kg bw per day in both the United Kingdom and the USA, and the intake of paraffin wax and microcrystalline wax was estimated to be 0.005 and 0.006 mg/kg bw per day, respectively (Table 6). These intakes clearly represent an insignificant proportion of the overall intake of mineral oils and waxes.
In the toxicological evaluation of mineral oils, different ADIs have been assigned to different groups of mineral oils and waxes on the basis of their physicochemical properties, because of differences in toxicity. The available data on intake could not, however, be matched to each group of mineral oils and waxes. Table 6 indicates that the intake of medium- and low-viscosity class III mineral oils by the populations of the United Kingdom and the USA was 0.21 and 0.25 mg/kg bw per day, respectively, which values are more than 20 times the temporary ADI for these oils, of 0–0.01 mg/kg bw. The intake of consumers at the 90th percentile in the United Kingdom, 0. 80 mg/kg bw per day, is 80 times the ADI. As separate intake estimates could not be made for high-viscosity and classes I and II medium- and low-viscosity mineral oils, these intake estimates could not be compared with the ADIs.
The intake of microcrystalline wax from food use was estimated to be 0.01 and 0.02 mg/kg bw per day in the United Kingdom and the USA, respectively, which values are well below the ADI of 0–20 mg/kg bw.
Although the intake of naturally occurring hydrocarbons was not taken into account in this evaluation, it is well known that they contribute a significant proportion of total dietary hydrocarbon intake. For example, in the United Kingdom (Food Chemical Risk Analysis, 2001), the intake of naturally occurring hydrocarbons in food (0.25 mg/kg bw per day) accounted for 35% of the total intake, 0.72 mg/kg bw per day, representing 0.47 mg/kg bw per day from total food use (including petroleum jelly) and 0.25 mg/kg bw per day from naturally occurring sources. In the USA (Reich et al., 1997, 1998), the estimated intake of naturally occurring hydrocarbons in food (0.19 mg/kg bw per day) accounted for 18% of the total intake, 1.1 mg/kg bw per day (0.88 plus 0.19 mg/kg bw per day). These estimated contributions of naturally occurring hydrocarbons are conservative, because the intake from food use was overestimated while the intake from natural sources was underestimated.
The estimated average total intake of mineral hydrocarbons (excluding petroleum jelly) resulting from use in food in both the United Kingdom and the USA was 0.47 mg/kg bw per day, white mineral oil accounting for the largest proportion (0.40 and 0.43 mg/kg bw per day, respectively). As limited data were available, the intake from various types of medium- and low-viscosity mineral oils could not be estimated separately. The best estimate is that the total intake of mineral hydrocarbons from low- and medium-viscosity mineral oils is more than 0.25 mg/kg bw per day. Bakery products, fruits and vegetables, grains and oil and confectionery are the major food sources of intake. This value is likely to be an overestimate, as it was assumed that 100% of each food category contained mineral oils at the maximum permitted level of use; furthermore, some food uses described above are not currently common at the global level.
Naturally occurring hydrocarbons are widely distributed in many edible plants and animals, and natural sources contribute significantly to the overall dietary intake of hydrocarbons. The dietary intake of naturally occurring hydrocarbons was estimated to be 0.47 mg/kg bw per day for the populations of the 15 countries of the European Union, 0.25 mg/kg bw per day for the population of the United Kingdom and 0.19 mg/kg bw per day for that of the USA. As data on the concentrations of naturally occurring hydrocarbons in food were limited, these values are likely to be underestimates.
The estimated intake of medium- and low-viscosity class III mineral oils (0.21 and 0.25 mg/kg bw per day in the United Kingdom and the USA) was more than 20 times the temporary ADI for this group (0–0.01 mg/kg bw). The estimated intake of microcrystalline wax in both countries were well below its ADI (0–20 mg/kg bw); however, microcrytalline wax contributes only a small proportion to overall intake of mineral hydrocarbons. The available data on intake of other classes of white mineral oils do not allow comparisons of intake with their ADIs, as separate data were not available for class I and II mineral oils and high-viscosity mineral oils. Data on the intake of paraffin wax were available, but no ADI has been established.
Intake data for each category of mineral oils should be made available to allow a complete evaluation of intake.
The Committee suggested that the Codex Committee on Food Additives and Contaminants consider reviewing the standards for mineral oil, particularly for class III medium- and low-viscosity mineral oils, because mineral oil is included in the draft General Standard for Food Additives in numerous food categories, with maximum use levels ranging from 200 to 3000 mg/kg (step 6). In some food items, its use level is required to be in accordance with good manufacturing practice; however, according to the principles of the Codex Committee, substances with numerical ADIs should not be included in the category of use according to good manufacturing practice.
Toxicological data
In the long-term studies of toxicity, Fischer 344 rats were given a diet containing P70(H) or P100(H) oil, at concentrations that varied to provide constant doses ranging from 60 to 1200 mg/kg bw per day, for up to 24 months. At 24 months, treatment with either test material was associated with mild treatment-related effects, which included: dose-related increases in the absolute and relative weights of the mesenteric lymph nodes; an increase in the severity of reticuloendothelial-cell hyperplasia (previously described as focal histiocytosis) of the mesenteric lymph nodes; increases in the incidence and/or severity of vacuolation of periportal hepatocytes; and an increased incidence of sinusoidal dilatation in the liver of male rats only, which was not dose-related. These effects were observed even at the lowest dose tested, 60 mg/kg bw per day. There were no treatment-related effects on survival with either material. Neither P70(H) nor P100(H)was carcinogenic in this study.
The effects observed at 12 months were marginal and included a slight increase in the severity of reticuloendothelial-cell hyperplasia of the mesenteric lymph nodes in male and female rats treated with P70(H) at the highest dose, the only dose for which histopathological data were available, as well as statistically significantly increased absolute and relative weights of mesenteric lymph nodes in males at this dose. Because of the marginal nature of these changes at 12 months, the reversibility of the lesions at the end of the recovery period could not be assessed; however, the results did indicate lack of progression of tissue alterations after dietary exposure to mineral hydrocarbons for 12 months.
No special indicators of immune function were measured in the long-term studies, although standard end-points of haematology, clinical chemistry, organ weight and histopathology, indicative of immune function, were evaluated. The only alteration in these end-points was exacerbation of mild cellular infiltration of the mesenteric lymph nodes, which had not been observed previously after exposure for 90 days.
Aside from the earlier onset and increased severity of reticuloendothelial-cell hyperplasia of the mesenteric lymph nodes and the greater increase in the weights of mesenteric lymph nodes in response to dietary administration of P70(H), there was little difference in response to the two oils. None of the effects at 24 months was reported in 90-day studies at concentrations up to 2% of the diet (equivalent or equal to 2000 mg/kg bw per day) of either material. The pigmented macrophages observed with P70(H) oil at 2% in the diet (equivalent to 2000 mg/kg bw per day) in the 90-day study reviewed at the forty-fourth meeting, on which the ADI was based, were not observed after 24 months’ intake at doses up to 1200 mg/kg bw per day.
Accumulated hydrocarbon was detected in the liver, mesenteric lymph nodes and spleen; the amounts of hydrocarbon could be quantified only in the liver. In the liver, a steady-state level of hydrocarbon accumulation appeared to have been reached in rats treated with the highest dose (1200 mg/kg bw per day) by 3 months, the only dose for which data on the complete time course were available. The concentration of P70(H) reached in the liver at any given time was about twice that of P100(H). The concentrations of mineral hydrocarbons in the liver returned to control levels within 12 months of cessation of treatment.
Medium-viscosity liquid petroleum (corresponding to class I medium- and low-viscosity mineral oils, including P70(H) oil) was not carcinogenic when fed to Fischer 344 rats at a concentration of 2.5% or 5% in the diet (equivalent to 1250 and 2500 mg/kg bw per day). Increased incidence or severity of reticuloendothelial-cell hyperplasia of the mesenteric lymph nodes was also observed in rats of each sex in this study at both dietary concentrations.
The pharmacokinetics of single doses of P15(H) oil given by gavage at a dose of 34 or 340 mg/kg bw over 96 h was studied in female Fischer 344 and Sprague-Dawley rats, with [14C]1-eicosanylcyclohexane as the tracer. In one of the studies, the animals received a 2-week dietary pretreatment with low-viscosity mineral oil.
The faeces were the major route of elimination, and urinary excretion was a minor route. Elimination of radiolabel by either route was more rapid in Sprague-Dawley than in Fischer 344 rats. Estimates of the area under the curve for serum concentrations of mineral hydrocarbons indicated that the bioavailability of mineral hydrocarbon was greater in Fischer 344 rats. Rats of this strain also accumulated and retained a larger proportion of the administered dose in the liver (2% vs 0.1–0.5%) and, to a much lesser extent (about 100-fold), the mesenteric lymph nodes. While a longer delay in faecal excretion of radiolabel was seen in Fischer 344 rats than in Sprague-Dawley rats in one study, suggesting a strain-related difference in intestinal absorption of mineral hydrocarbons, this difference was not observed in two similar studies. Rather, the results suggested that Sprague-Dawley rats metabolized and excreted radiolabelled mineral hydrocarbon tracer more efficiently than Fischer 344 rats, which exhibited deposition and retention in the liver. Dietary pretreatment with mineral oil did not affect the extent to which hydrocarbon accumulated in the liver of either strain of rats. In another study, the rate of metabolism of N-heptadecane by liver microsomes in vitro differed by rat strain, but not sex, in the order Wistar > Sprague-Dawley > Fischer 344. Metabolism of N-octadecane was negligible in all strains, consistent with the findings for hydrocarbons with more than 17 carbons in microsomal systems in vitro.
New short-term studies, in which the responses of female Fischer 344 and Sprague-Dawley rats to dietary administration of P15(H) oil (for 120 and 90 days, respectively) at concentrations of up to 2% were compared, indicated a more severe response in the Fischer 344 strain. The effects seen in these rats were: increased weights of the liver, mesenteric lymph nodes and spleen; granulomatous inflammation of the liver; and reticuloendothelial-cell hyperplasia of the mesenteric lymph nodes. In Sprague-Dawley rats, only statistically significant increases in liver weight and reticuloendothelial-cell hyperplasia of the lymph nodes, mostly of minimal severity, were observed. The increased liver weights in Fischer 344 rats receded during a 30-day recovery period. Reversibility was not assessed in Sprague-Dawley rats.
The results of the new 90-day study in female Fischer 344 rats with low-melting-point wax and with N15(H), N70(H) and P70(H) oils at a dietary concentration of 2% (equal to approximately 2000 mg/kg bw per day at 90 days) confirmed the previous findings in lymph nodes, liver and spleen, with an additional finding of inflammation of the cardiac mitral valve, which had not been seen previously with this material. This effect occurred at a lower incidence than with low-melting-point wax in this study.
The Committee reviewed the results of a study of T-dependent humoral immune function in response to 90 days’ administration of P15(H) oil in female Fischer 344 and Sprague-Dawley rats. No effects were observed in the Sprague-Dawley rats. Interpretation of the results in the Fischer 344 rats was complicated by the increase in spleen weight that resulted from treatment. No data were provided to determine whether the increased spleen weight was associated with a change in the proportion of spleen cell types, which could alter interpretation of the results.
The responses of female Fischer 344 and Sprague-Dawley rats to mineral hydrocarbons were also compared in a 90-day study with low-melting-point wax at a dietary concentration of 0.2% or 2.0%. This substance had induced the most severe effects in similar studies in Fischer 344 rats with a range of food-grade mineral hydrocarbons, in which, in general, female rats had a stronger response to these materials. The results for Fischer 344 rats in the new study confirmed the findings of the previously evaluated 90-day studies with low-melting-point wax: altered haematological and hepatic clinical chemical parameters, increased weights of the liver, mesenteric lymph nodes and spleen; hepatic vacuolation (affecting both hepatocytes and Kupffer cells) and granulomatous inflammation; reticuloendothelial-cell hyperplasia of mesenteric and cervical lymph nodes; and focal inflammation of the cardiac mitral valve. In addition, increased severity of extramedullary haematopoiesis of the spleen, differential effects on serum immunoglobin concentrations and lipid-like material in crystalline form in hepatocytes and Kupffer cells were observed in this strain. In contrast, the only effects observed in female Sprague-Dawley rats were increased liver weights and slightly increased reticuloendothelial-cell hyperplasia of the mesenteric lymph nodes and severity of hepatocellular and Kupffer-cell vacuolation. The crystalloid structures and inflammatory response in the liver were absent in this strain.
Tissue accumulation of mineral hydrocarbon after administration of low-melting-point wax also showed a clear strain difference in terms of degree, onset and the tissues affected. Quantifiable amounts of hydrocarbon were detected in the liver and mesenteric lymph nodes of Fischer 344 rats in a time- and dose-related manner, starting at 30 days in the livers of animals at the higher dose and occurring in both tissues in animals at both doses by 90 days. However, only the mesenteric lymph nodes of Sprague-Dawley rats had quantifiable amounts of hydrocarbon, and only at 90 days in animals at the higher dose. This observation was confirmed in a qualitative investigation of the accumulation of low-melting-point wax in the liver and Kupffer cells of female Fischer 344 and Sprague-Dawley rats after administration of a diet containing 2% wax for 14, 30 or 60 days. Detectable amounts were found in liver cells of Fischer 344 rats by 14 days, but none was found at any time in Sprague-Dawley rats.
In a separate study of low-melting-point wax, feeding a diet containing 2% resulted in alterations in Kupffer-cell function and morphology after 60 days in female Fischer 344, but not Sprague-Dawley, rats. Effects were also seen on selected parameters (i.e. leukocytes, serum liver enzyme activities and hepatic histopathology) that have been associated with administration of low-melting-point wax in this strain, and these were accompanied by accumulation of mineral hydrocarbon in the liver.
Intake
At its present meeting, the Committee also assessed the intake of mineral oils. The major food uses of mineral oils are in vegetable and fruit coatings and in packaging, bakery and confectionery manufacture and grain and seed dust control, and these uses are also the major sources of dietary intake of mineral oils in some countries. Dietary intake was assessed from data on the levels of use in foods and on migration from coatings and packaging materials into foods, combined with national data on food consumption in the 1990s. In both the United Kingdom and the USA, the average total intake of mineral hydrocarbons (excluding petroleum jelly) from food use was estimated to be 0.47 mg/kg bw per day; the intake at the 90th percentile of consumption by the population of the United Kingdom was 0.80 mg/kg bw per day. Class III medium- and low-viscosity mineral oils (including P15(H) oil) accounted for 0.21 mg/kg bw per day in the United Kingdom and 0.25 mg/kg bw per day in the USA; these values are 21 and 25 times the temporary ADI of 0–0.01 mg/kg bw, respectively, whereas class I and II medium- and low-viscosity mineral oils and high-viscosity mineral oil (including P70(H) and N70(H) oils, respectively) accounted for 0.18 and 0.19 mg/kg bw per day, respectively. As these different categories of mineral oil have different ADIs, but data on intake are not available for separate categories, the intake of each category cannot be compared with the corresponding ADI. Use of solid hydrocarbons (e.g. microcrystalline wax (high-melting-point wax) and paraffin wax (low-melting-point wax)) accounted for the remainder of the total intake.
The intake of high-viscosity and class I, II and III medium- and low-viscosity mineral oils that have migrated into food from coating and packaging materials was estimated to be 0.001 mg/kg bw per day in both the United Kingdom and the USA, while the combined intake of paraffin wax and microcrystalline wax from this source was estimated to be 0.005 and 0.006 mg/kg bw per day, respectively.
Naturally occurring hydrocarbons are widely distributed in many edible plants and animals, and they contribute significantly to the overall dietary intake of hydrocarbons. For example, the dietary intake of naturally occurring hydrocarbons was estimated to be 0.47, 0.25 and 0.19 mg/kg bw per day in the populations of the European Union, the United Kingdom and the USA, respectively. It is clear, therefore, that account should be taken of intakes from naturally occurring hydrocarbons when evaluating the safety of mineral oils.
Mineral oil is included in the Codex draft GSFA in numerous food categories, with maximum use levels of 200–3000 mg/kg and, in some categories, in accordance with good manufacturing practice. The Committee suggested that the Codex Committee on Food Additives and Contaminants consider reviewing the standards for mineral oil, particularly in view of the fact that the intake of class III medium- and low-viscosity mineral oil is more than 20 times higher than its temporary ADI of 0–0.01 mg/kg bw.
No new information was available on the compositional elements of mineral oils that affect their absorption or toxicity.
As the materials tested in the long-term studies reviewed at the present meeting, P70(H) and P100(H) oils, were not associated with induction of liver granulomas in Fischer 344 rats, the studies did not help the Committee to determine the long-term consequences or reversibility of the liver granulomas that had been seen in previous studies in response to consumption of low- and medium-viscosity mineral oils and low-melting-point waxes by Fischer 344 rats. In addition, the Committee was unable to interpret the effects in the study of humoral immune function in response to dietary administration of P15(H) oil.
The results of the studies on the effects of P15(H) oil and low-melting-point wax in Fischer 344 and Sprague-Dawley strains indicated that the more extensive response of Fischer 344 rats, in particular that of females, is associated with greatly enhanced retention of mineral hydrocarbons in the tissues, which is probably due to a reduced ability to metabolize absorbed hydrocarbons. The Committee concluded that additional studies are needed in order to determine whether the Fischer 344 rat is an appropriate model of human response to dietary intake of food-grade mineral hydrocarbons. In particular, elucidation of the metabolic differences between Fischer 344 rats and other strains and species, including humans, would be useful.
Neither P70(H) nor P100(H) oil was carcinogenic in the combined study of toxicity and carcinogenicity reviewed by the Committee at its present meeting. The effects observed even at the lowest dose, i.e. enhanced reticuloendothelial-cell hyperplasia, increased weights of mesenteric lymph nodes and increased incidence and grade of vacuolation of hepatocytes, were shown not to progress to more severe effects, and there was no indication that accumulated test material contributed to suppression or activation of an inflammatory response. Consequently, these effects were considered to be indicators of exposure to mineral hydrocarbon rather than adverse effects. At the present meeting, the NOEL for P70(H) oil was identified as the highest dose tested in the combined study of toxicity and carcinogenicity in rats, 1200 mg/kg bw per day, to which a safety factor of 100 was applied. An ADI of 0-10 mg/kg bw was allocated for class I medium- and low-viscosity mineral oils, which include P70(H) oil. An ADI of 0–20 mg/kg bw for P100(H) oil already exists (see Table 1).
No data were available that would permit allocation of a full ADI for medium- and low-viscosity mineral oils in classes II and III. The Committee noted that the new information reviewed at the present meeting indicated that the observed effects of these mineral oils, on which the temporary ADI is based, may be strain- and sex-specific. The Committee therefore extended the temporary group ADI of 0–0.01 mg/kg bw for class II and III medium- and low-viscosity mineral oils until 2006, pending information on the relevance to humans of the response of Fischer 344 and Sprague-Dawley rats to these materials. In order for the data to be applicable to as wide a range of mineral oils as possible, the Committee suggested that commercial mineral oils of the lowest viscosity be used in such studies. Further studies might be required, depending on the outcome of these studies.
Baldwin, M.K., Berry, P.H., Esdaile, D.J., Linnett, S.L., Martin, J.G., Peristianis, G.C., Priston, R.A.J., Simpson, B.J.E. & Smith, J.D. (1992) Feeding studies in rats with mineral hydrocarbon food grade white oils. Toxicol. Pathol., 20, 426–435.
Burleson, G.R. (2001) Unpublished study conducted by Burleson Research Technologies, Raleigh, North Carolina, USA for American Petroleum Institute, Washington DC, USA. Submitted to WHO by CONCAWE, Brussels, Belgium.
Cunninghame, M., Brantom, P., Scotter, M. & Castle, L. (2001) Identification of the causative agents(s) for mineral hydrocarbon toxicity. Unpublished report FD 00/51prepared by TNO-BIBRA International, Carshalton, Surrey, United Kingdom, and Central Science Laboratory, Sand Hutton, New York, USA. Submitted to WHO by CONCAWE, Brussels, Belgium.
Exxon Biomedical Sciences (2001) Combined chronic toxicity/carcinogenicity study of white oil in Fischer 344 rats. Unpublished report numbers 105970 and 106870. Submitted to WHO by CONCAWE, Brussels, Belgium.
Firriolo, J.M., Morris, C.F., Trimmer, G.W., Twitty, L.D., Smith, J.H. & Freeman, J.J. (1995) Comparative 90-day feeding study with low-viscosity white mineral oil in Fischer-344 and Sprague-Dawley-derived CRL:CD rats. Toxicol. Pathol., 23, 26–33.
Food Chemical Risk Analysis (2001) The usage, occurrence and dietary intake of white mineral oils and waxes and naturally occurring hydrocarbons in Europe. Prepared for CONCAWE and EWF.
Francke, S., Rojko, J.L. & Hall, W.C. (2001) Unpublished report IM392 prepared by Pathology Associates International, Frederick, Maryland, USA for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Hoglen, N.C., Regan, S.P., Hensel, J.L., Younis, H.S., Sauer, J.-M., Steup, D.R., Miller, M.J., Waterman, S.J., Twerdok, L.E. & Sipes, I.G. (1998) Alteration of Kupffer cell function and morphology by low melt point paraffin wax in female Fischer-344 but not Sprague-Dawley rats. Toxicol. Sci., 46, 176–184.
ImmunoTox (2001) Effects of dietary mineral hydrocarbons on T-cell dependent antibody production in two strains of rats. Unpublished study ITI 1298 prepared for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Institut National de la Recherche Agronomique (2000) Unpublished report EP/98/HYDROCARB/01 prepared for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Reich, A,G., Waylett, D.K., van der Reit, E., Doyl, E., Douglass, J.S., Eickhoff, J.C. & Heimbach, J.T. (1997) Intake of naturally occurring alkanes. Prepared for the American Petroleum Institute by TAS-ENVIRON.
Reich, A,G., Douglass, J.S., van der Reit, B., Eickhoff, J.C. & Heimbach, J.T. (1998) Dietary exposure assessment for direct and indirect food use applications for mineral hydrocarbons. Prepared for the American Petroleum Institute by TAS-ENVIRON.
Shoda, T., Toyoda, K., Uneyama, C., Takada, K. & Takahashi, M. (1997) Lack of carcinogenicity of medium-viscosity liquid paraffin given in the diet to Fischer 344 rats. Food Chem. Toxicol., 35, 1181–1190.
Sipes, I.G. & Halladay, J.S. (undated) Pilot pharmacokinetic studies on mineral hydrocarbons (MHC). Unpublished study prepared for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Sipes, I.G. & Halladay, J.S. (2001a) Final report: 10-fold lower dose study. Unpublished study prepared for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Sipes, I.G. & Halladay, J.S. (2001b) Final report: 2-week pretreatment study. Unpublished study prepared for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Sipes, I.G. & Hoglen, N.C. (1998) Analysis of low melt point paraffin wax in liver digests and Kupffer cells by gas chromatography and gas chromatography–mass spectrometry. Unpublished study prepared for American Petroleum Institute, Washington DC. Submitted to WHO by CONCAWE, Brussels, Belgium.
Smith, J.H., Mallett, A.K., Priston, R.A.J., Brantom, P.G., Worrell, N.R., Sexsmith, C. & Simpson, B.J. (1996) Ninety-day feeding study in Fischer-344 rats of highly refined petroleum-derived food-grade white oils and waxes. Toxicol. Pathol., 24, 214–230.
Thornton, S.R., Auletta, C.S., Bolte, H.F., Gosselin, S.J., Dadgar, D. & Cojocaru, L.V. (2000) A subchronic 90-day dietary toxicity study of a low melting point paraffin wax in two rat strains. Unpublished report from Huntingdon Life Sciences, East Millstone, New Jersey, USA. Submitted to WHO by CONCAWE, Brussels, Belgium.
See Also:
Toxicological Abbreviations