
INTERNATIONAL PROGRAMME ON CHEMICAL SAFETY
ENVIRONMENTAL HEALTH CRITERIA 122
n-HEXANE
This report contains the collective views of an international group of
experts and does not necessarily represent the decisions or the stated
policy of the United Nations Environment Programme, the International
Labour Organisation, or the World Health Organization.
Published under the joint sponsorship of
the United Nations Environment Programme,
the International Labour Organisation,
and the World Health Organization
First draft prepared by Dr. K. Chipman,
University of Birmingham, United Kingdom
World Health Orgnization
Geneva, 1991
The International Programme on Chemical Safety (IPCS) is a
joint venture of the United Nations Environment Programme, the
International Labour Organisation, and the World Health
Organization. The main objective of the IPCS is to carry out and
disseminate evaluations of the effects of chemicals on human health
and the quality of the environment. Supporting activities include
the development of epidemiological, experimental laboratory, and
risk-assessment methods that could produce internationally
comparable results, and the development of manpower in the field of
toxicology. Other activities carried out by the IPCS include the
development of know-how for coping with chemical accidents,
coordination of laboratory testing and epidemiological studies, and
promotion of research on the mechanisms of the biological action of
chemicals.
WHO Library Cataloguing in Publication Data
n-Hexane.
(Environmental health criteria ; 122)
1.Hexanes - adverse effects 2.Hexanes - toxicity
I.Series
ISBN 92 4 157122 5 (NLM Classification: QV 633)
ISSN 0250-863X
The World Health Organization welcomes requests for permission
to reproduce or translate its publications, in part or in full.
Applications and enquiries should be addressed to the Office of
Publications, World Health Organization, Geneva, Switzerland, which
will be glad to provide the latest information on any changes made
to the text, plans for new editions, and reprints and translations
already available.
(c) World Health Organization 1991
Publications of the World Health Organization enjoy copyright
protection in accordance with the provisions of Protocol 2 of the
Universal Copyright Convention. All rights reserved.
The designations employed and the presentation of the material
in this publication do not imply the expression of any opinion
whatsoever on the part of the Secretariat of the World Health
Organization concerning the legal status of any country, territory,
city or area or of its authorities, or concerning the delimitation
of its frontiers or boundaries.
The mention of specific companies or of certain manufacturers'
products does not imply that they are endorsed or recommended by the
World Health Organization in preference to others of a similar
nature that are not mentioned. Errors and omissions excepted, the
names of proprietary products are distinguished by initial capital
letters.
CONTENTS
ENVIRONMENTAL HEALTH CRITERIA FOR n-HEXANE
1. SUMMARY
2. IDENTITY, PHYSICAL AND CHEMICAL PROPERTIES, ANALYTICAL METHODS
2.1. Identity
2.2. Physical and chemical properties
2.3. Conversion factors
2.4. Analytical methods
3. SOURCES OF HUMAN AND ENVIRONMENTAL EXPOSURE
3.1. Natural occurrence
3.2. Production and uses
3.2.1. Production levels and processes
3.2.1.1 Production figures
3.2.1.2 Manufacturing processes
3.2.2. Uses
4. ENVIRONMENTAL TRANSPORT, DISTRIBUTION, AND TRANSFORMATION
4.1. Transport and distribution between media
4.2. Biotransformation and photochemical reactivity
5. ENVIRONMENTAL LEVELS AND HUMAN EXPOSURE
5.1. Environmental levels
5.1.1. Air
5.1.2. Water
5.1.3. Food
5.2. Occupational exposure during manufacture, formulation, or
use
6. KINETICS AND METABOLISM
6.1. Experimental animals
6.1.1. Absorption, distribution, metabolism, and
excretion
6.1.2. Kinetics of 2,5-hexanedione
6.1.3. In vitro studies
6.1.4. Effects of other chemicals on n-hexane metabolism
6.2. Human beings
7. EFFECTS ON ORGANISMS IN THE ENVIRONMENT
8. EFFECTS ON EXPERIMENTAL ANIMALS AND IN VITRO TEST SYSTEMS
8.1. Single exposures
8.1.1. Inhalation exposure
8.1.2. Oral administration
8.1.3. Dermal exposure
8.1.4. Parenteral administration
8.1.5. In vitro exposure
8.2. Short- and long-term exposures
8.2.1. Inhalation studies
8.2.1.1 Combined-exposure effects
8.2.1.2 Effects on the respiratory tract
8.2.1.3 Effects on the testes
8.2.1.4 Other effects
8.2.2. Oral studies
8.2.3. Dermal studies
8.2.4. In vitro studies
8.2.5. Parenteral studies
8.3. Reproduction, embryotoxicity, and teratogenicity
8.3.1. Teratogenicity studies
8.3.1.1 Inhalation studies
8.3.1.2 Oral
8.3.2. Fertility studies
8.4. Mutagenicity and related end-points
8.5. Carcinogenicity
8.5.1. Inhalation studies
8.5.2. Skin-painting studies
8.6. Neurotoxicity
8.6.1. Central nervous system effects
8.6.2. Peripheral nervous system effects
8.6.2.1 Clinical and morphological findings
8.6.2.2 Electrophysiological effects
8.6.2.3 Grip strength
8.7. Toxicity of n-hexane metabolites
9. EFFECTS ON MAN
9.1. Single exposures
9.2. Skin and eye irritation; sensitization
9.2.1. Skin irritation
9.2.2. Eye irritation
9.2.3. Skin sensitization
9.3. Short- and long-term exposures
9.3.1. Effects on the nervous system
9.3.1.1 Peripheral neuropathy
9.3.1.2 Effects on vision and optic nerves
9.3.2. Effects on the kidney
9.3.3. Effects on other organs
10. EVALUATION OF HUMAN HEALTH RISKS AND EFFECTS ON THE
ENVIRONMENT
10.1. Animal studies
10.1.1. Short- and long-term exposure
10.1.2. Genotoxicity
10.1.3. Carcinogenicity
10.1.4. Reproductive effects
10.2. Human studies
10.2.1. Effects on the nervous system
10.2.2. Other effects
10.3. Environmental effects
11. RECOMMENDATIONS
11.1. Human health protection
11.2. Environmental protection
12. FURTHER RESEARCH
REFERENCES
RESUME
EVALUATION DES RISQUES POUR LA SANTE HUMAINE ET DES EFFETS SUR
L'ENVIRONNEMENT
RECOMMANDATIONS
RECHERCHES A EFFECTUER
RESUMEN
EVALUACION DE LOS RIESGOS PARA LA SALUD HUMANA Y DE LOS EFECTOS EN
EL MEDIO AMBIENTE
RECOMENDACIONES
NUEVAS INVESTIGACIONES
WHO TASK GROUP ON ENVIRONMENTAL HEALTH CRITERIA FOR n-HEXANE
Members
Professor E.A. Bababunmi, Department of Tropical Paediatrics,
Liverpool School of Tropical Medicine, Liverpool, United
Kingdom (Rapporteur)
Dr M. Cikrt, Centre of Industrial Hygiene and Occupational
Diseases, Institute of Hygiene and Epidemiology, Prague,
Czechoslovakia (Vice-Chairman)
Dr S. Dobson, Pollution and Ecotoxicology Section, Institute of
Terrestrial Ecology, Monks Wood Experimental Station,
Huntingdon, United Kingdom
Professor C.L. Galli, Toxicology Laboratory, Institute of
Pharmacological Sciences, University of Milan, Milan, Italy
(Chairman)
Dr S.D. Gangolli, British Industrial Biological Research
Association, Carshalton, Surrey, United Kingdom
Dr C. Konantakieti, Technical Division, Food and Drug
Administration, Ministry of Public Health, Bangkok, Thailand
Dr O. Ladefoged, Laboratory of Pathology, Institute of
Toxicology, National Food Agency of Denmark, Ministry of Health,
Soborg, Denmark
Professor A. Massoud, Department of Community Environmental and
Occupational Medicine, Ainshams Faculty of Medicine, Cairo,
Egypt
Dr V. Riihimäki, Department of Industrial Hygiene and Toxicology,
Institute of Occupational Health, Helsinki, Finland
Observer
Dr H.P.A. Illing, Health and Safety Executive, Bootle, Merseyside,
United Kingdom
Secretariat
Dr P.G. Jenkins, International Programme on Chemical Safety, World
Health Organization, Geneva, Switzerland
Ms B. Labarthe, International Register of Potentially Toxic
Chemicals, United Nations Environment Programme, Geneva,
Switzerland
Dr E. Smith, International Programme on Chemical Safety, World
Health Organization, Geneva, Switzerland (Secretary)
NOTE TO READERS OF THE CRITERIA DOCUMENTS
Every effort has been made to present information in the
criteria monographs as accurately as possible without unduly
delaying their publication. In the interest of all users of the
environmental health criteria monographs, readers are kindly
requested to communicate any errors that may have occurred to the
Manager of the International Programme on Chemical Safety, World
Health Organization, Geneva, Switzerland, in order that they may be
included in corrigenda, which will appear in subsequent volumes.
* * *
A detailed data profile and a legal file can be obtained from
the International Register of Potentially Toxic Chemicals, Palais
des Nations, 1211 Geneva 10, Switzerland (Telephone No. 7988400 or
7985850).
ENVIRONMENTAL HEALTH CRITERIA FOR n-HEXANE
A WHO Task Group on Environmental Health Criteria for n-Hexane
met in Carshalton, United Kingdom, from 12 to 16 March 1990.
Dr E.M. Smith opened the meeting and welcomed the participants on
behalf of the heads of the three IPCS cooperating organizations
(UNEP/ILO/WHO). The Task Group reviewed and revised the draft
criteria document and made an evaluation of the health risks of
exposure to n-hexane.
The first draft of this document was prepared by Dr K. Chipman,
University of Birmingham, United Kingdom. The second draft was
also prepared by Dr Chipman, incorporating comments received
following circulation of the first draft to IPCS contact points for
Environmental Health Criteria monographs. Particularly valuable
comments on the draft were made by the National Food Agency,
Denmark, the National Institute of Public Health and Environmental
Protection, The Netherlands, the European Chemical Industry Ecology
and Toxicology Centre (ECETOC), and the US Environmental Protection
Agency, National Institute of Environmental Health Sciences, Food
and Drug Administration, and Centers for Disease Control.
Dr E.M. Smith and Dr P.G. Jenkins, both members of the IPCS
Central Unit, were responsible for the overall scientific content
and technical editing, respectively, of this monograph.
The efforts of all who helped in the preparation and
finalization of the document are gratefully acknowledged.
* * *
Financial support for the Task Group was provided by the United
Kingdom Department of Health as part of its contributions to IPCS.
Partial financial support for the publication of this monograph was
kindly provided by the United States Department of Health and Human
Services, through a contract from the National Institute of
Environmental Health Sciences, Research Triangle Park, North
Carolina, USA - a WHO Collaborating Centre for Environmental Health
Effects.
ABBREVIATIONS
ACGIH American Conference of Governmental Industrial Hygienists
ADI acceptable daily intake
BAER brainstem auditory-evoked response
EEC European Economic Community
EEG electroencephalogram
EMG electromyography
ip intraperitoneal
MEK methyl ethyl ketone
NOEL no-observed-effect level
TWA time-weighted average
1. SUMMARY
n-Hexane (normal hexane) is a colourless, volatile liquid.
Commercial hexane is mainly a mixture of hexane isomers and related
6-carbon compounds, and has an n-hexane content varying between 20
and 80%. Gas chromatography coupled with flame ionization
detection or mass spectroscopy is a suitable technique for the
measurement of n-hexane. Occupational exposure limits range from
100 - 1800 mg/m3 (time-weighted average, TWA) and 400 - 1500 mg/m3
(ceiling value, CLV) in various countries.
n-Hexane can be isolated from natural gas and crude oil. It is
used in food processing, including the extraction of vegetable oil,
and as a solvent in various products and processes.
Once emitted into the environment, n-hexane exists
predominantly in the vapour phase. In the atmosphere its half-life
is estimated to be approximately 2 days, based on its reactivity
with the OH radical alone. Reported LC50 values for aquatic
organisms are few and variable and have been conducted under
inappropriate conditions; an assessment of the toxic effects of
n-hexane in this environment is, therefore, not possible. Low
water solubility and high volatility make exposure of aquatic
organisms unlikely except from uncontrolled discharge into surface
waters.
In mammals, n-hexane is absorbed rapidly through the lungs and
is distributed widely in the adult body, as well as to fetal
tissue. Dermal absorption is limited. n-Hexane is metabolized
oxidatively to a number of compounds, including 2,5-hexanedione,
which is thought to be the ultimate neurotoxic agent. Particularly
high levels of n-hexane and 2,5-hexanedione can occur in the
sciatic nerve of rats. Most n-hexane is excreted unchanged in
exhaled air; some is excreted as metabolites in exhaled air and
urine.
n-Hexane is of low acute toxicity for adult rats by oral
administration or inhalation. Oral LD50 values of 15 - 30 g/kg
have been recorded, and an inhalation LC50 value of 271 040 mg/m3
(77 000 ppm) has been reported for a 1-h exposure. At high vapour
concentrations, animals show ataxia, seizures, and signs of central
nervous system depression.
Testicular lesions and neurotoxicity appear to be the principal
effects of repeated n-hexane exposure in rats. Severe testicular
lesions have resulted from inhalation exposure to n-hexane and oral
exposure to 2,5-hexanedione. Effects have been attributed to
disruption of the cytoskeleton of Sertoli cells. There are
secondary effects on post-spermatogonial germ cells, which
disappear from affected tubules. Testicular effects were
reversible after a single exposure for 24 h to 17 600 mg/m3 (5000
ppm) but irreversible after a 2-week exposure to the same
concentration for 16 h/day, 6 days/week. 2,5-Hexanedione at 1% in
drinking-water produced similar reversible testicular lesions after
2 to 3 weeks of dosing and irreversible effects (within 17 weeks)
after 5 weeks of dosing.
The neurotoxic effect is characterized clinically by hindlimb
weakness, which can progress to paralysis. Axonal swellings
develop in the central and peripheral nervous systems; more severe
lesions (axonal degeneration and loss) can occur, particularly in
the longest, largest-diameter nerves. In essentially continuous
6-month inhalation studies, peripheral and central nervous system
lesions were present at doses of 1760 mg/m3 (500 ppm) or more, but
no clinical or pathological effects were noted at 440 mg/m3 (125
ppm). Only limited recovery of amplitude of the fifth brainstem
auditory-evoked response (believed to reflect central nervous
system activity) and tail nerve action potential was recorded 15 -
22 weeks after cessation of continuous exposure to a vapour
concentration of 3520 mg/m3 (1000 ppm), 5 days/week, for 11 weeks.
Discontinuous exposure of rats to 3168 mg/m3 (900 ppm) for 72 weeks
did not cause any apparent peripheral or central nervous system
lesions, but there was some evidence of electrophysiological
effects on peripheral nerves.
n-Hexane-induced neurotoxicity can be enhanced by combined
exposure to methyl ethyl ketone, methyl isobutyl ketone, and lead
acetate, and decreased by co-exposure to toluene. Toluene and
n-hexane also have a synergistic effect in the disturbance of
dopamine levels.
Severe microscopic lesions were noted in skin when n-hexane
was applied dermally under occlusive conditions for short periods.
Prolonged exposure to an n-hexane vapour concentration of 10 560
mg/m3 (3000 ppm) can cause conjunctival irritation in rats and
marked ocular irritation in rabbits. No skin sensitization data
are available from animal studies.
Chromosomal damage (polyploidy in one study, structural
aberrations in a second study) has been reported in both in vitro
and in vivo studies. No increase in point mutation frequency or
effects in tests for DNA damage has been noted.
There has been one carcinogenicity study with n-hexane (skin
painting on mice), which provided no evidence of carcinogenicity.
The reproductive toxicity of n-hexane has not been studied
adequately. There was no substantial evidence of embryotoxicity or
teratogenicity in rats following inhalation, though concentrations
were relatively low, or in mice after oral dosing. Postnatal
development of rats was transiently delayed when dams were exposed
to an n-hexane vapour concentration of 3520 mg/m3 (1000 ppm).
Very little information is available on the acute toxicity of
n-hexane to humans. Most studies have involved occupational
exposure to solvent mixtures. The available data suggest that
n-hexane has low acute toxicity. Signs of central nervous system
depression, such as drowsiness, vertigo, and giddiness, have been
reported after exposure to a commercial hexane level of 3520 to
17 600 mg/m3 (1000 - 5000 ppm) for 10 - 60 min.
n-Hexane is a mild irritant causing transient erythema when in
contact with human skin for short periods. More severe effects
(erythema and blistering) were documented after occlusive skin
contact for 5 h with commercial grade hexane. There have been no
case reports of sensitization of skin in exposed workers, and no
skin sensitization was noted in a maximization test with n-hexane.
On repeated exposure, n-hexane is neurotoxic, inducing a type
of sensorimotor peripheral neuropathy. Many studies on the
prevalence of n-hexane-induced neurotoxicity have been published;
however, adequate exposure data are often lacking. Exposure to
n-hexane concentrations in air varying from 106 - 8800 mg/m3
(30 - 2500 ppm) has been associated with neuropathy. Cases of
marked peripheral neuropathy were reported among Japanese sandal
workers and Taiwanese press proofers exposed to n-hexane levels of
approximately 176 and 352 mg/m3 (50 and 100 ppm), respectively, for
periods exceeding 8 h per day. In many cases exposure measurements
were recent and probably did not accurately reflect previous
exposures causing neuropathy.
Several cross-sectional studies have independently reported
mild subclinical effects (for example, electrophysiological
changes in peripheral nerves) in workers exposed to 70 - 352 mg/m3
(20 - 100 ppm). However, no clear cases of clinically overt
peripheral neuropathy were identified in any of these studies at
exposure levels of less than 352 mg/m3 (100 ppm).
The effects of n-hexane on the central nervous system have been
investigated only in a few studies. Changes in somatosensory
evoked potentials recorded from workers exposed to n-hexane were
suggested to result from a central nerve conduction block. Altered
visual evoked potentials and EEG traces have also been noted.
These results suggest that n-hexane may produce central nervous
system dysfunction, but the available data provide no information
on related exposure levels.
2. IDENTITY, PHYSICAL AND CHEMICAL PROPERTIES, ANALYTICAL METHODS
2.1. Identity
Common synonyms: Hexyl hydride, hexane, Skellysolve B
Chemical structure:
H H H H H H
| | | | | |
| | | | | |
H---C---C---C---C---C---C---H
| | | | | |
| | | | | |
H H H H H H
Chemical formula: C6H14
CAS registry number: 110-54-3
Relative molecular mass: 86.177
2.2. Physical and chemical properties
Some physical and chemical properties of various grades of
n-hexane are given in Table 1.
Table 1. Physical and chemical properties of n-hexanea
------------------------------------------------------------
Boiling point (°C) 68.74b
Melting point (°C) -95.35b
Relative density (20 °C/4 °C) 0.66
Vapour pressure (25 °C) 20 kPa (150 mmHg)
Vapour density 2.97
Autoignition temperature (°C) 225
Explosive limit in air (% by volume) 1.1-7.5
Flash point (°C) -21.7
Closed-cup flash point (°C)c -30.56
Solubility in water (mg/litre at 25 °C) 9.5
Log n-octanol/water partition coefficient
(log Pow at 25 °C) 3.6
Refractive index (20 °C) 1.37
Colour, Saybolt +39
------------------------------------------------------------
a From: Mellan (1977)and IRPTC (1990).
b From: Clayton & Clayton (1981).
c From: ACGIH (1986).
n-Hexane is colourless, highly volatile (NIOSH, 1977a), and
flammable (Dale & Drehman, 1980; ACGIH, 1986). It is poorly
soluble in water but is soluble in most organic solvents including
ethanol and ether (McAuliffe, 1963; NIOSH, 1977a; ACGIH, 1986).
Purified n-hexane contains 95 - 99.5% n-hexane, together with
small amounts of other hexane isomers as impurities (Mellan, 1977;
Baker & Rickert, 1981; Sandmeyer, 1981). Traces of benzene (0.05%)
have been detected (Baker & Rickert, 1981). Commercial hexane is a
mixture of hexane isomers ( n-hexane, 2-methylpentane,
3-methylpentane, 2,3-dimethylbutane), cyclohexane, methyl
cyclopentane and small amounts of pentane and heptane isomers,
acetone, methyl ethyl ketone, dichloromethane, and
trichloroethylene (Perbellini et al., 1981a,b,c; ACGIH, 1986). The
n-hexane content of commercial hexane (Table 2) can vary between
20% and 80% (ACGIH, 1986).
Table 2. Composition (% by weight) of different
grades of n-hexanea
------------------------------------------------
Research Pure Technical
grade grade grade
------------------------------------------------
n-Hexane 99.98 99.5 95-97.7
2-Methylpentane trace trace trace
3-Methylpentane 0.02 0.1 0.2
Methylcyclopentane trace 0.4 2.1
------------------------------------------------
a From: Mellan (1977).
Pure n-hexane contains approximately 0.0005% of non-volatile
material (Mellan, 1977), whereas commercial hexane may contain up
to 0.04% (Patty & Yant, 1929; Vicedo et al., 1985). In commercial
hexane, a number of phthalate esters (including dimethyl, diethyl,
di- n-butyl, di-isobutyl, dihexyl, and diethylhexyl), adipate
esters (dibutyl and dioctyl), and organophosphorus compounds (e.g.,
triphenyl phosphate) have been identified and total about 0.3% of
the distillation residue (Vicedo et al., 1985).
Commercial hexane (containing about 30% n-hexane) has a
slightly disagreeable odour, perceptible at 282 mg/m3 (80 ppm)
(intermittent exposure) or 528 mg/m3 (150 ppm) (continuous
exposure) (Patty & Yant, 1929). An odour threshold of 211 mg/m3
(60 ppm) for hexane (purity not stated) has also been reported
(Laffort & Dravnieks, 1973).
2.3. Conversion Factors
1 ppm n-hexane in air = 3.52 mg/m3
1 mg/m3 = 0.284 ppm n-hexane in air
2.4. Analytical Methods
n-Hexane may be analysed by gas chromatography with flame
ionization detection or mass spectroscopy. A summary of the
techniques employed is given in Table 3.
Carbon cloth can be used for diffusive sampling. It has been
shown that when it is wrapped in aluminium foil there is no
significant loss of n-hexane during storage for at least four days
(Kasahara & Ikeda, 1987).
Table 3. Techniques for the analysis of n-hexane
---------------------------------------------------------------------------------------
Medium Separation Detection Limit of Reference
method system sensitivity
---------------------------------------------------------------------------------------
Air trap with charcoal; flame validated over the NIOSH (1977b)
desorb with CS2; ionization range 877-3679
steel column; gas mg/m3 at 17 °C,
chromatography 764 mmHg; probable
(FFAP) useful range:
180-5400 mg/m3
Air trap with charcoal; flame measurement range: NIOSH (1984)
desorb with CS2; gas ionization 3.56-14.5 mg for
chromatography 5-µl injection
Air diffusive sampling; stain tube detection range: Gentry & Walsh
thermal desorption (Kitagawa 18-500 µg (1987)
133 5B)
Liquid glass column; gas
chromatography; 5%
carbowax (partition) flame not given Franke et al.
or 0.3% carbowax ionization (1988)
(adsorption)
Liquid capillary glass flame 0.05 µg Nomeir &
column; gas ionization Abou-Donia
chromatography (1985)
(OV101)
Biological glass column; gas mass 80 ng/g (biological Tsuruta (1980)
samples chromatography spectrometer sample)
(Porapak Q) (multiple ion
monitoring
m/z 85)
---------------------------------------------------------------------------------------
A high performance liquid chromatographic method using a silica
cartridge has been developed for the analysis of n-hexane
metabolites (2-hexanone, 2,5-dimethylfuran, gamma-valqerolactone,
5-hydroxy-2-hexanone, and 2,5-hexanedione) in chicken plasma
(Nomeir & Abou-Donia 1985). Metabolites of n-hexane have also been
analysed by gas chromatography coupled with mass spectroscopy
(DiVincenzo et al., 1976).
3. SOURCES OF HUMAN AND ENVIRONMENTAL EXPOSURE
3.1. Natural Occurrence
n-Hexane is present in natural gas and crude oil (Guthrie,
1960).
3.2. Production and Uses
3.2.1. Production levels and processes
3.2.1.1 Production figures
It has been estimated that 306 000 tonnes of n-hexane was
recovered from petroleum and natural gas in 1977 in the USA (Dale &
Montgomery, 1983). Babich et al. (1982) cited approximate
estimates of 240 000 to 465 000 tonnes (80 - 155 million gallons)
for the annual production of naphtha (of which hexane is the
principal component) in the USA in 1978/1979, and the total
production in the USA in 1987 was 386 500 tonnes (USITC, 1988).
3.2.1.2 Manufacturing processes
Two-tower distillation of a suitable hydrocarbon feedstock is
used for the manufacture of commercial hexanes. The feedstock may
be straight-run gasolines distilled from crude oil or natural gas
liquids stripped of natural gas. Hexanes can also be obtained from
the remains of catalytic reformates after the removal of aromatics.
Very pure n-hexane can be produced from hexane mixtures by
adsorption on molecular sieves (Dale & Drehman, 1983).
3.2.2. Uses
In most cases, n-hexane is used as a mixture with other hexane
isomers and various solvents. The following uses of n-hexane have
been reported (Dale & Drehman, 1983; CCOHS, 1985):
* in food processing, including the extraction of vegetable oil
from soybeans, flaxseed, peanuts, safflower seed, corngerm, and
cottonseed;
* as a polyolefins solvent and as a cleaning agent;
* as a rubber polymerization solvent;
* as a laboratory chemical;
* in low-temperature thermometers;
* in the manufacture of pharmaceuticals;
* in other products (e.g., adhesives, lacquers).
The consumption of hexane (not specifically n-hexane) in the
USA and Canada during 1975 for the above purposes was estimated to
be 450 000 to 490 000 m3 (120 - 130 million gallons) (Dale &
Drehman, 1983). The consumption of n-hexane in the EEC in 1979 was
10 000 tonnes.
In general, petroleum distillate solvents of a relatively low
boiling point contain a relatively high proportion of n-hexane
(Kasahara et al., 1987). Rubber surface softener samples and
approximately one half of the analysed samples of thinner-cleaner
used for printing and painting were in this category (Kasahara et
al., 1987). Ikeda & Kasahara (1986) found the n-hexane content to
be 0.4 - 9% in four samples of gasoline with boiling points in the
range 30 - 160 °C.
4. ENVIRONMENTAL TRANSPORT, DISTRIBUTION, AND TRANSFORMATION
4.1. Transport and Distribution Between Media
There is little information on the transport and mobility of
n-hexane in the environment. It is very insoluble in water (9.5
mg/litre at 25 °C) (McAuliffe, 1963), and transport occurs
predominantly in the vapour phase once it is emitted into the
environment. The solubility of n-hexane in water is increased by
the presence of methanol (Groves, 1988).
Chiou et al. (1988) determined that the sorptive capacity of
soil for hexane is 11 mg/g (0.0167 ml/g).
4.2. Biotransformation and Photochemical Reactivity
Information is only available on the abiotic degradation of
n-hexane. Atmospheric n-hexane is not expected to have a
pronounced effect on the physical properties of the atmosphere, to
participate in the depletion of the ozone layer, or to alter
precipitation patterns (CIIT, 1977). The rate constant for the
reaction of hydroxyl (OH) radicals with n-hexane was determined
(using a smog chamber) to be 6.2 (± 0.6) x 10-12 cm3 sec-1 at 39 °C
(Nolting et al., 1988). Other reported values are 5.63 (± 0.09) x
10-12 cm3 sec-1 at 26 °C (Atkinson et al., 1982) and 5.68 (± 0.04)
x 10-12 cm3 sec-1 (Behnke et al., 1988). In the latter study,
there was additional degradation in the presence of titanium
dioxide aerosol. These rate coefficients are in good agreement
with the value of Klopffer et al. (1988), who measured a KOH of 6.8
x 10-12 cm3 sec-1 in a smog chamber at 27 °C. The half-life of
n-hexane in the troposphere is estimated to be 2 to 2.4 days, based
on degradation by OH radicals alone.
Similar hydrocarbons ( n-pentane and methyl pentane) undergo
photochemical conversion to a "smog" containing
peroxyacetylnitrate and ozone, but n-hexane is one of the least
photochemically reactive hydrocarbons (Katagiri & Ohashi, 1975).
5. ENVIRONMENTAL LEVELS AND HUMAN EXPOSURE
5.1. Environmental Levels
5.1.1. Air
Fugitive losses of n-hexane may occur in industries in which it
is used as a solvent or cleaning agent and in the rubber industry.
In 1970, prior to the mandatory use of catalytic converters,
n-hexane was also estimated to represent 1.2% (by volume) of total
emitted hydrocarbons from vehicular exhausts in the USA (equivalent
to approximately 170 000 tonnes of hexane) (CIIT, 1977). Lonneman
et al. (1974) detected n-hexane in air samples collected in the
Lincoln Tunnel, New York, over a 0.5-h period. n-Hexane has been
detected at a level of 0.11 mg/m3 (0.03 ppm) in the ambient
atmosphere of Los Angeles (Nelligan, 1962). Hodgson et al. (1986)
reported that the concentrations of n-hexane in a single open-
office space were 1.97 µg/m3 (0.56 ppb) and 4 µg/m3 (1.14 ppb)
(with 100% ventilation exhaust on and off, respectively), compared
with an outdoor concentration that was below the limit of
quantification.
In the Federal Republic of Germany, n-hexane belongs to class
III of chemical substances, the total emission of which (as the sum
of all compounds in this class) must not exceed 150 mg/m3 at a mass
flow of 3 kg/h or more (IRPTC, 1990). In the USSR, the ambient
vapour concentration of n-hexane is regulated at a maximum of 300
mg/m3 (IRPTC, 1990).
5.1.2. Water
n-Hexane has been detected in the USA in trace quantities in
chlorinated tap water derived from a lake (CEC, 1979).
5.1.3. Food
The Joint FAO/WHO Expert Committee on Food Additives (JECFA)
has not set an ADI for n-hexane but stresses that the solvent
should be used only in accordance with good manufacturing practice
to ensure minimal residues in food (WHO, 1971). In the USA,
cottonseed products and hop extract, modified for human
consumption, may contain no more than 60 mg/kg and 25 mg/kg,
respectively (CFR, 1987a). The latter limit also applies to
certain food colouring agents (CFR, 1987b). The EEC Directive on
Extraction Solvents (June 1988) set maximum residue levels in food
of between 1 and 30 mg/kg depending on the particular use (IRPTC,
1990).
5.2. Occupational Exposure During Manufacture, Formulation, or Use
In addition to the presence of n-hexane in the air during its
manufacture, a study has indicated detectable levels in air samples
from a variety of industrial uses of products containing n-hexane.
The relative abundance (percentage of samples in which it was
detected) was 15% (printing), 7% (painting), 10% (car repair), and
6% (various other operations) (Veulemans et al., 1987). Levels of
hexane exposure in six olive extraction plants in Granada, Spain,
were found to vary from 3 to 341 mg per m3 (0.9 - 97 ppm)
(Medinilla & Espigares, 1989). An Italian worker employed at home
manufacturing shoes was found to have a urinary 2,5-hexanedione
level of 5.7 mg/litre, which exceeded the ACGIH biological exposure
index for hexane of 5 mg/litre (Discalzi et al., 1988).
Concentrations of n-hexane have also been measured in the air to
which petroleum service attendants, transport drivers, and outside
operators were exposed. For outside operators, 54 out of 56
samples contained measurable concentrations of n-hexane, the mean
concentration being 0.473 ± 0.594 mg/m3. All 49 samples of air for
transport drivers contained n-hexane (mean concentration, 1.019 ±
1.953 mg/m3). Service attendants were exposed to a mean
concentration of 1.175 ± 0.894 mg/m3 (detected in 48/49 samples).
These values represented 2.5, 2.2, and 1.7% of the total
hydrocarbon concentration for the three job categories,
respectively (Rappaport et al., 1987). Maximum time-weighted
average (8 h) concentrations of hexane at a hexane extraction
facility were found to be 92 mg/m3 (26 ppm) (NIOSH, 1981a). NIOSH
(1983) reported hexane concentrations in air samples from six
breathing zones at a soybean extraction facility that ranged from
15.5 to 46.5 mg/m3 (4.4 - 13.2 ppm). In area spot samples, the
concentrations were 46.5 - 94.7 mg/m3 (13.2 - 26.9 ppm).
In one metropolitan sewer in Cincinnati, USA, in 1981, the
airspace was found to contain hexane at a concentration greater
than the lower explosive limit (1.2% by volume in air). Two sewers
contained hexane levels of at least 39 600 mg/m3 (11 250 ppm)
(NIOSH, 1981b).
Some limits for occupational exposure in various countries are
shown in Table 4. Some countries (e.g., USA, Belgium, and the
United Kingdom) also recommend an exposure limit (TWA) of 360 mg/m3
for all hexane isomers other than n-hexane.
Table 4. Some national occupational air exposure limit valuesa
-----------------------------------------------------------------------------------
Country/ Exposure limit descriptionb Value Effective
organization datec
-----------------------------------------------------------------------------------
Australia Threshold limit value (TLV)
- Time-weighted average (TWA) 360 mg/m3 1985(r)
Belgium Threshold limit value (TLV)
- Time-weighted average (TWA) 180 mg/m3 1989(r)
Canada Threshold limit value (TLV)
- Time-weighted average (TWA) 180 mg/m3 1989
Finland Maximum permissible concentration (MPC)
- Time-weighted average (TWA) 180 mg/m3 1989(r)
- Short-term exposure limit (STEL) 530 mg/m3
-----------------------------------------------------------------------------------
Table 4. (contd.)
-----------------------------------------------------------------------------------
Country/ Exposure limit descriptionb Value Effective
organization datec
-----------------------------------------------------------------------------------
Germany, Maximum acceptable concentration (MAK)
Federal - Time-weighted average (TWA) 180 mg/m3
Republic of - Short-term exposure limit (STEL) 360 mg/m3 1989(r)
- Biological tolerance value (BAT) urine: 9 mg/litre
hexane-2,5-dione plus 4,5-dihydroxy-2-
haxanone at end of exposure or end of
shift
Germany, Maximum acceptable concentration (MAC)
Democratic - Time-weighted average (TWA) 100 mg/m3
Republic of - Short-term exposure limit (STEL) 400 mg/m3 1988(r)
Italy Threshold limit value (TLV)
- Time-weighted average (TWA) 360 mg/m3 1985(r)
Japan Maximum acceptable concentration (MAC)
- Time-weighted average (TWA) 141 mg/m3 1985
(skin absorption must also be considered)
Poland Maximum permissible concentration (MPC)
- ceiling value (CLV) 400 mg/m3 1986(r)
Romania Maximum permissible concentration (MPC)
- Time-weighted average (TWA) 1200 mg/m3
- Ceiling value (CLV) 1500 mg/m3 1975(r)
Switzerland Maximum acceptable concentration (MAK)
- Time-weighted averge (TWA) 180 mg/m3 1987
Sweden Hygienic limit value (HLV)
- Time-weighted average (TWA) 90 mg/m3
- Short-term exposure limit (STEL) 180 mg/m3 1990(n)
United Guidance limit (under review)
Kingdom - Time-weighted average (TWA) 360 mg/m3
- Short-term exposure limit (STEL) (10 min) 450 mg/m3 1990(n)
USA (ACGIH) Threshold limit value (TLV)
- Time-weighted average (TWA) 176 mg/m3 1990(r)
- Biological exposure index (BEI) 5 mg/litre
2,5-hexanedione in urine (end of shift)
n-hexane in end-exhaled air (during 144 mg/m3
shift)
-----------------------------------------------------------------------------------
Table 4. (contd.)
-----------------------------------------------------------------------------------
Country/ Exposure limit descriptionb Value Effective
organization datec
-----------------------------------------------------------------------------------
USA (OSHA) Permissible exposure limit (PEL) 180 mg/m3 1990(r)
Yugoslavia Maximum permissible concentration (MAC)
- Time-weighted average (TWA) 1800 mg/m3 1971(r)
-----------------------------------------------------------------------------------
a From: IRPTC (1990).
b TWA = a maximum mean exposure limit based generally over the period of a working
day (usually 8 or 12 h)
STEL = a maximum concentration of exposure for a specified time duration
(generally 15 or 30 min).
c When no effective date appears in the IRPTC legal file, the year of the reference
from which the data are taken is shown, indicated by (r); n = notified direct by
country.
6. KINETICS AND METABOLISM
6.1. Experimental Animals
6.1.1. Absorption, distribution, metabolism, and excretion
Radiolabelled 14C- n-hexane was absorbed by F-344 rats
following inhalation of concentrations of 1760, 3520, 10 560, and
35 200 mg/m3 (500, 1000, 3000, and 10 000 ppm) for 6 h daily for 3
days (Bus et al., 1982). The proportion of 14C- n-hexane recovered
in expired air was dose dependent and increased from 12% at 1760
mg/m3 (500 ppm) to 62% at 35 200 mg/m3 (10 000 ppm). There was a
corresponding decrease in the proportion recovered in urine (from
35% at 1760 mg/m3 (500 ppm) to 18% at 35 200 mg/m3 (10 000 ppm), in
faeces (from 4.9% at 1760 mg/m3 (500 ppm) to 0.7% at 35 200 mg/m3
(10 000 ppm), and in the carcass (from 10% at 1760 mg/m3 (500 ppm)
to 1.5% at 35 200 mg/m3 (10 000 ppm). The levels of 14C- n-hexane
in expired air decreased biphasically, with half-lives of 1 and
4.5 h. A biphasic elimination profile was reported for labelled
carbon dioxide in exhaled air, the initial half-life being 4.5 -
7.5 h and 85 - 96% of the exhaled labelled carbon dioxide being
eliminated within 24 h after exposure. Urinary excretion of
metabolites also occurred in two phases, most being excreted in the
initial phase with a half-life of 7 - 8 h. The greater part of the
radioactivity absorbed during inhalation of n-hexane was excreted
within 24 h. The rate of metabolism was non-linear above 1056 mg/m3
(300 ppm) but not below this concentration: it rose from 47 µmol/h
per kg body weight at 1056 mg/m3 (300 ppm) to 245 µmol/h per kg
body weight at 10 560 mg/m3 (3000 ppm). The amount of n-hexane in
the rat increased in a dose-dependent manner to a limiting value of
9.6 relative to the atmospheric concentration (Filser et al.,
1987).
The rate of uptake following inhalation of hexane in male
F-344/N rats was found to be 5 - 7 nmol/kg per min (Dahl et al.,
1988). Absorption and distribution into tissues and organs
(including blood) was rapid following inhalation exposure of F-344
rats to n-hexane (Baker & Rickert, 1981; Howd et al., 1982).
Exposure to concentrations between 3520 mg/m3 and 35 200 mg/m3
(1000 ppm and 10 000 ppm) resulted in plateau levels of n-hexane
being reached within 30 min in blood and within 2 h in the other
tissues examined (liver, kidney, lungs, testis, brain, and sciatic
nerve) (Baker & Rickert, 1981). Acute inhalation exposure to
302 720 mg/m3 (86 000 ppm) resulted in saturated blood levels
within 10 min (Raje et al., 1984).
Dermal absorption of n-hexane by guinea-pigs is poor (Jakobson
et al., 1982). Tsuruta (1982) measured the penetration of
n-hexane through excised rat skin and came to a similar conclusion.
However, skin absorption of n-hexane may be enhanced by other
solvents in the mixture. The highest tissue concentrations of
n-hexane were found in peripheral nerves (sciatic) (Baker &
Rickert, 1981; Bus et al., 1981). Steady-state concentrations in
blood, sciatic nerve, liver, and lung were directly proportional to
the exposure level, but there was some evidence of saturation in
the kidney, brain, and testis.
In female albino rats exposed to a hexane level of 17 000 mg/m3
(50 000 ppm) for up to 10 h, the hexane concentration in the liver
increased with time and steady-state levels were not achieved
(Böhlen et al., 1973). However, kidney, adrenal, blood, brain, and
spleen levels of hexane reached a steady state after approximately
5 h of exposure. The high level of hexane exposure resulted in
lipid accumulation in the liver, and, as hexane is highly lipid
soluble, this could account for the non-saturability of liver for
hexane.
n-Hexane has been detected in the fat, muscle, and bone
(sternum) of F-344 rats exposed to an n-hexane level of 21 123
mg/m3 (6800 ppm) for 4 h and in rat fetuses following inhalation
exposure of pregnant females (Bus et al., 1979). The concentration
in total fetal tissue was similar to that in maternal blood
(Babanov & Babanov, 1981).
Elimination of n-hexane from rat blood and brain was found to
be rapid and multiphasic, with initial half-lives of 2 - 4 and 7
min and a subsequent half-life of 1 - 2 h (Bus et al., 1979; White
et al., 1979; Baker & Rickert, 1981; Howd et al., 1982). In the
rat, elimination from other adult tissues, including sciatic nerve
and liver, and from fetal tissue was also rapid but was slower from
the kidneys (half-life, 6 h). In the guinea-pig, biphasic
elimination from blood (half-lives of 0.5 h and 4 h) has been
reported (Couri et al., 1978).
n-Hexane is oxidized to 1-, 2-, or 3-hexanol. 2-Hexanol may be
further metabolized to 2-hexanone (methyl- n-butyl ketone).
2,5-Hexanediol, 5-hydroxy-2-hexanone, 2,5-hexanedione, gamma-
valerolactone, 2,5-dimethylfuran, 2,5-dimethyl-2,3-dihydrofuran,
2,5-dimethyltetrahydrofuran, and carbon dioxide have been
identified following exposure to n-hexane or 2-hexanone. However,
furan derivatives and gamma-valerolactone are questionable
metabolites of n-hexane because they may be artefacts (DiVincenzo
et al., 1976; Perbellini et al., 1982a; Fedtke & Bolt, 1986b).
2,5-Dimethyl-2,3-dihydrofuran and small amounts of labelled
2-aminohexanoic acid (norleucine), another unidentified amino acid,
and urea have been detected following oral dosing of rats with
1-14C-2-hexanone. Pentanone (isomeric form not specified) has been
detected in rat fetuses from dams exposed to 2-hexanone vapour
(DiVincenzo et al., 1977; Peters et al., 1981). Radiolabelled
2,5-hexanedione is metabolized to carbon dioxide in the rat (Angelo
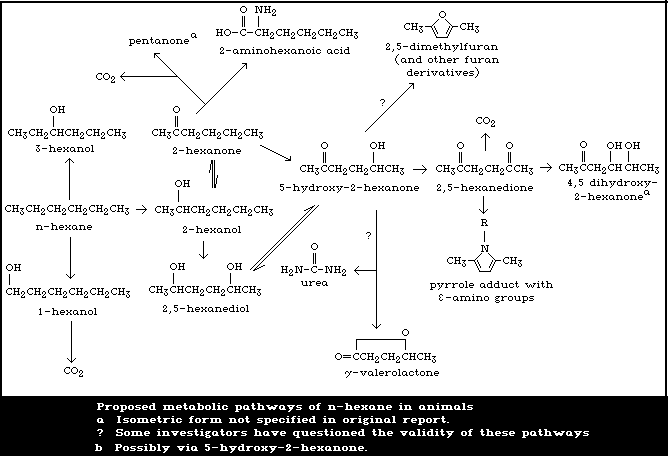
& Bischoff, 1982). Proposed metabolic pathways for n-hexane in
mammals are shown in Fig. 1.
2-Hexanol was the major urinary metabolite excreted by rats
during n-hexane inhalation exposure for up to 24 h (Perbellini et
al., 1982a; Iwata et al., 1983b). After exposure, the main urinary
metabolites were 2,5-hexanedione, 2,5-dimethylfuran, and gamma-
valerolactone (Baker & Rickert, 1981; Perbellini et al., 1982a;
Iwata et al., 1983b). All the gamma-valerolactone and a
significant proportion of the other metabolites were conjugated;
glucuronides of 2-hexanol and 2,5-dimethylfuran were identified.
Only small amounts of 2-hexanone, 5-hydroxy-2-hexanone, and
3-hexanol were present. 1-Hexanol, mainly as the glucuronide, has
been detected following intraperitoneal injection of n-hexane
(Dolara et al., 1978). With prolonged exposure to 1760 mg/m3 (500
ppm) (3 h/day, for 33 weeks), 80 - 90% of all urinary metabolites
were conjugated, the predominant metabolites being 2-hexanol and
2,5-hexanedione (Iwata et al., 1984). In urine collected from rats
during and up to 24 h after exposure to 3520 mg per m3 (1000 ppm),
the predominant urinary metabolites were 2-hexanol and
2,5-hexanedione, of which 99 and 90%, respectively, were found
(following acid hydrolysis) to be conjugated. In addition, all the
1-hexanol and 3-hexanol and 30% of the 2-hexanol were conjugated.
The hexanol conjugates were considered to be glucuronides and
sulfate esters, but the identity of the other conjugates was
unclear (Fedtke & Bolt, 1986b). Subsequently, Fedtke & Bolt
(1987a) identified 4,5-dihydroxy-2-hexanone as an additional
glucuronic acid conjugate in the urine of male Wistar rats treated
with n-hexane at 7040 mg/m3 (2000 ppm) for 8 h or given a single
ip dose of 2,5-hexanedione (200 mg/kg). This metabolite was
considered to be produced either by hydroxylation of 5-hydroxy-2-
hexanone (which may be produced by reduction of 2,5-hexanedione) or
alternatively by hydroxylation and subsequent reduction of
2,5-hexanedione. Thus, 4,5-dihydroxy-2-hexanone may be a
detoxified product of 2,5-hexanedione. This metabolite was the
second most abundant metabolite in the urine of male Wistar rats
following inhalation of n-hexane (176 - 10 820 mg/m3, 50 - 3074
ppm) for 8 h (Fedtke & Bolt, 1987b).
 In studies on the rabbit and monkey (Macaca mulatta), the
urinary excretion of n-hexane metabolites was found to differ from
that of rats (Perbellini et al., 1982b). 2-Hexanol was the main
urinary metabolite during and after inhalation exposure. The other
metabolites present were 3-hexanol and 2,5-hexanedione; gamma-
valerolactone and 2,5-dimethylfuran were not detected. Following
intraperitoneal injection of n-hexane in guinea-pigs, 2-hexanol,
mainly as the glucuronide, and n-hexane itself were identified in
the urine (Couri et al., 1978).
Elimination half-lives in rat urine for individual metabolites
following inhalation of n-hexane have been determined for
2-hexanone (4 h), 3-hexanol and 2,5-hexanedione (7 h),
2,5-dimethylfuran and gamma-valerolactone (11 - 14 h), and
2-hexanol (18 h) (Perbellini et al., 1982a). The half-life for
2-hexanol in the rabbit and monkey was found to be shorter (8 h)
than in the rat.
5-Hydroxy-2-hexanone and 2,5-hexanedione were detected in
guinea-pig serum after intraperitoneal administration of n-hexane
(DiVincenzo et al., 1976). Following administration of 2-hexanol,
a major urinary metabolite of n-hexane in some species, two
additional compounds, 2-hexanone and 2,5-hexanediol, were found. A
similar metabolic profile was found following the administration of
2-hexanone, with 5-hydroxy-2-hexanone, 2,5-hexanedione, and
2-hexanol being identified as metabolites. 2,5-Hexanedione, which
is considered to be the active neurotoxic agent, can thus be formed
from n-hexane, 2-hexanol, or 2-hexanone.
6.1.2. Kinetics of 2,5-hexanedione
A marked increase in blood levels of 2,5-hexanedione on
inhalation exposure to n-hexane at 3168 or 17 600 mg/m3 (900 or
5000 ppm) for 6 - 24 h has been observed in separate studies
(Perbellini et al., 1982a; Kulig, 1983; Kulig et al., 1984). No
evidence of 2,5-hexanedione accumulation in blood was found in
Wistar rats after repeated exposures to n-hexane at 3168 mg/m3 (900
ppm), 8 h/day, for 9 days (Kulig, 1983). 2,5-Hexanedione was
completely eliminated, within 6.25 h of the end of exposure, from
the blood of Wistar rats exposed to an n-hexane level of 3168 mg/m3
(900 ppm) 8 h/day, for 72 weeks (De Groot & Kepner, 1984). There
was no significant difference in 2,5-hexanedione blood levels in
Fischer-344 rats exposed for 1 or 5 days to 3520 mg/m3 (1000 ppm)
for 6 h/day (Bus et al., 1981). In contrast, there were
significantly increased 2,5-hexanedione concentrations in the blood
of Fischer-344 rats after 3 exposures to 14 080 mg/m3 (4000 ppm)
for 8 h/day compared with a single 8-h exposure (Howd et al., 1982).
The elimination half-life for 2,5-hexanedione from blood has
been reported to vary between different strains of rat. Values for
Sprague-Dawley and Fischer-344 rats were 2.3 h and 3.9 - 6 h,
respectively, and an initial half-life of 1 h has been reported for
Wistar rats (Bus et al., 1979; Angelo & Bischoff, 1982; Howd et
al., 1982; Kulig, 1983). It has been suggested that differences in
the rates of elimination of 2,5-hexanedione from the blood may
explain the reported greater susceptibility to n-hexane-induced
neurotoxicity of Fischer-344 rats compared with Wistar rats (Kulig,
1983; Kulig et al., 1984).
In a study by Ladefoged & Perbellini (1986), 2,5-hexanedione
(50 mg/kg) was administered to five male rabbits by intravenous
injection. The pharmacokinetic data fitted a two-compartment model
in which the half-life was 42 (± 11) min (body clearance was 0.0117
(± 0.0026) ml/min per kg).
Blood concentrations of 2,5-hexanedione have been estimated
following continuous exposure of rats to n-hexane (Kulig, 1983;
Kulig et al., 1984). Marked reductions in blood 2,5-hexanedione
levels were reported following exposure to 3168 mg/m3 (900 ppm) for
3 days compared with one day. There were no further decreases
after 9 or 20 days of exposure. Smaller reductions were observed
in short-term studies after 8 or 12 weeks of exposure to 3168 mg/m3
(900 ppm) for either 8 or 24 h/day, 5 days per week, compared with
the blood levels of 2,5-hexanedione recorded after 4 weeks of
exposure to 3168 mg/m3 (900 ppm). No effects were reported in
similar short-term studies at 1056 mg/m3 (300 ppm) (Kulig, 1983).
This may be due to the enhancement of glucuronidation (a major
elimination pathway) by hexane, a phenomenon noted in both in vitro
and in vivo studies in the guinea-pig (Notten & Henderson,
1975a,b). Similar levels of 2,5-hexanedione in blood were found in
weanling and young adult rats following one week of continuous
exposure to 3520 mg/m3 (1000 ppm) (Pryor et al., 1982).
2,5-Hexanedione was detected in the blood, sciatic nerve,
brain, kidneys, liver, and lungs, but not in the testes, following
exposure of Fischer-344 rats to levels of n-hexane between 1760 and
35 200 mg/m3 (500 to 10 000 ppm) for 6 h (Baker & Rickert, 1981).
The relationship between peak tissue concentrations of
2,5-hexanedione and n-hexane exposure levels was complex. In the
kidneys, sciatic nerve, and brain, the highest concentrations were
obtained after exposure to 3520 mg/m3 (1000 ppm). In a further
study, lower tissue levels and greater elimination of
2,5-hexanedione were found in mice than in rats after a single
exposure to 3520 mg/m3 (1000 ppm) for 6 h (Baker et al., 1980).
The elimination of 2,5-hexanedione from the rat liver, kidney,
brain, and sciatic nerve was determined after exposure to 3520
mg/m3 (1000 ppm) 6 h/day, for 1 or 5 days (Bus et al., 1981).
2,5-Hexanedione was selectively retained in sciatic nerve but not
in the brain. However, comparable rates of elimination of
2,5-hexanedione from the brain, sciatic nerve, and blood were
reported in Sprague-Dawley rats given a single oral dose of
2,5-hexanedione (Iwasaki & Tsuruta, 1984). Terminal half-lives of
32 - 33 days were reported in a review of some earlier studies
(O'Donoghue, 1985).
2,5-Hexanedione has also been detected in fetal tissue, at a
level similar to that in maternal blood, following exposure of
F-344 rats to n-hexane (Bus et al., 1979). A fetal half-life of
3 h was calculated.
Less than 10% of 2,5-hexanedione in blood is bound to plasma
components (Angelo & Bischoff, 1982). A study on hens dosed orally
with 2,5-hexanedione showed that it can form a pyrrole adduct with
serum protein (DeCaprio et al., 1982).
6.1.3. In vitro studies
n-Hexane penetrates excised rat skin slowly (Tsuruta, 1977,
1982).
In rat liver and lung preparations, n-hexane has been shown to
be hydroxylated to 1-, 2-, and 3-hexanol (Frommer et al., 1974;
Walseth et al., 1982; Toftgard et al., 1984). 2-Hexanol was found
to be the predominant metabolite, and 2,5-hexanedione was also
formed in studies with rat liver microsomes (Frommer et al., 1974;
Walseth et al., 1982). More than one form of cytochrome P-450 is
involved in n-hexane oxidation (Frommer et al., 1974). In the rat
lung the formation of 2- and 3-hexanol is catalysed by a different
microsomal cytochrome P-450 isoenzyme (cytochrome P-450-PB-B) from
that responsible for the formation of 1-hexanol (Toftgard et al.,
1984). Toftgard et al. (1984, 1986) found that rat liver
microsomes, but not lung microsomes, contained a cytochrome P-450
isoenzyme that converted 2-hexanol to 2,5-hexanediol, and that the
oxidation of hexanols and 2,5-hexanediol by alcohol dehydrogenase
was restricted to the liver. These results suggest that, during
inhalation of n-hexane, the metabolite 2,5-hexanedione is likely to
be formed in the liver but not in the lung.
Preparations of purified rabbit liver cytochrome P-450
hydroxylate n-hexane, and mouse liver microsomes hydroxylate hexane
(Ichihara et al., 1969; Nilsen et al., 1981). Studies using liver
fractions from guinea-pigs demonstrated that n-hexane can be
metabolized to 2-hexanol, 2-hexanone, and 2,5-hexanedione (Couri et
al., 1978). 2-Hexanone was reduced to 2-hexanol by the cytosolic
fraction and oxidized to 2,5-hexanedione by the microsomal
fraction.
In mouse nerve and muscle tissue cultures, n-hexane was
metabolized to 2-hexanediol and 2,5-hexanedione, and 5-hydroxy-2-
hexanone was detected following incubation with 2-hexanol or
2-hexanone (Veronesi et al., 1978, 1980; Spencer et al., 1980).
2,5-Hexanedione has been found to react with primary amino
groups in proteins, such as the epsilon-amino group of lysine, to
yield 2,5-dimethylpyrrole adducts (DeCaprio et al., 1982; Graham et
al., 1982a,b). Pyrrole formation is an obligatory step in the
pathogenesis of neuropathy caused by n-hexane (Sayre et al., 1986).
2,5-Hexanedione appears to be the active neurotoxic metabolite of
n-hexane.
6.1.4. Effects of other chemicals on n-hexane metabolism
Urinary excretion of n-hexane metabolites by rats has been
shown to be increased following pre-treatment with phenobarbital,
an inducer of the microsomal oxidation of foreign compounds by
cytochrome P-450 isoenzymes (Perbellini et al., 1979, 1982a).
In in vitro studies, pre-treatment of rats with phenobarbital
increased the extent of liver microsomal oxidation of n-hexane to
2- and 3-hexanol, and pre-treatment with another inducer,
3,4-benzo[ a]pyrene, enhanced 3-hexanol formation (Frommer et al.,
1974; Näslund & Halpert, 1984). However, phenobarbital pre-
treatment of rats had no effect on n-hexane metabolism by lung
microsomes (Näslund & Halpert, 1984). Phenobarbital pre-treatment
of guinea-pigs increased the metabolism of n-hexane to 2-hexanol
and 2,5-hexanedione by hepatic post-mitochondrial supernatant, and
increased the metabolism of 2-hexanone to 2,5-hexanedione by the
hepatic microsomal fraction (Couri et al., 1978).
Intraperitoneal injection of phthalate esters to rats affects
the metabolism of n-hexane in vitro, presumably by induction of
cytochrome P-450 (Walseth et al., 1982). Treatment with
dimethylphthalate or dibutylphthalate increased significantly the
rate of formation of 2- and 3-hexanol by liver microsomes but
decreased slightly their formation by lung microsomes. A large
increase in 2,5-hexanedione formation by both types of microsomes
was noted following exposure to di(2-ethylhexyl)phthalate. Similar
results have been reported with inhalation exposures to
dibutylphthalate (Walseth & Nilsen, 1984). Male Sprague-Dawley
rats were exposed continuously for 5 days to atmospheric
dibutylphthalate concentrations of 5.7, 28.5, and 79.7 mg/m3 (0.5,
2.5, and 7 ppm), and liver and lung microsomal fractions were
prepared. The in vitro formation of n-hexane metabolites was
assayed by incubating 50 ml of 8% (v/v) n-hexane in sodium
phosphate buffer with 1 mg lung or liver microsomal protein for 10
min. The formation of 1-, 2-, and 3-hexanol was markedly increased
with liver microsomes but not with lung microsomes. At the
intermediate and high exposure concentrations of 28.5 and 79.7
mg/m3 (2.5 and 7 ppm), there was a decrease in in vitro n-hexane
metabolism particularly in lung microsomes.
The formation of 1-, 2-, and 3-hexanol from n-hexane was
significantly increased with kidney and liver microsomes obtained
from rats previously exposed to isopropanol (Zahlsen et al., 1985).
Enhanced formation of 2- and 3-hexanol (greater in male than in
female rats), but not of 1-hexanol, occurred when n-hexane was
incubated with liver microsomes from rats previously exposed to
xylene (mixed isomers) (Toftgard et al., 1983).
Treatment of rats with chloramphenicol (100 mg/kg
intraperitoneal or intravenous) inhibited 2- and 3-hexanol
formation in vitro by lung and liver microsomes derived from these
animals (Näslund & Halpert, 1984).
Toluene has been shown to be a non-competitive inhibitor of
n-hexane metabolism in in vitro studies with rat liver preparations
(Perbellini et al., 1982b). Exposure of rats to mixtures of
n-hexane and toluene resulted in reduced urinary excretion of
n-hexane metabolites (Perbellini et al., 1982b; Iwata et al.,
1983b, 1984). However, no effect on blood n-hexane levels was
reported when rats were given an intraperitoneal n-hexane injection
of 0.91 g/kg with or without 1.18 g toluene/kg (Suzuki et al.,
1974). In a study on the kinetics of 2,5-hexanedione (section
6.1.2), Ladefoged & Perbellini (1986) dosed six male rabbits
intravenously with 50 mg 2,5-hexanedione/kg with or without acetone
(150 mg/kg). A significant reduction of 2,5-hexanedione clearance
was caused by the acetone co-treatment.
Blood levels of 2,5-hexanedione were reduced after a single 6-h
co-exposure to n-hexane (3520 mg/m3, 1000 ppm) and methyl ethyl
ketone (2950 mg/m3, 1000 ppm), but not in animals pre-treated
orally with methyl ethyl ketone. However, pre-treatment resulted
in increases in liver cytochrome P-450 levels and in reactions
mediated by mixed-function oxidase (Robertson et al., 1982;
Robertson et al., 1989). Lower levels of 2,5-hexanedione have been
observed in the rat sciatic nerve after exposure to mixtures of
n-hexane and methyl ethyl ketone, but full data were not presented
(White & Bus, 1980).
In studies by Ralston et al. (1985), the blood and tissue
clearance of 2,5-hexanedione was measured after single or repeated
oral administration of 2,5-hexanedione or of a mixture of equimolar
doses of 2,5-hexanedione and methyl ethyl ketone. There was
reduced blood elimination of 2,5-hexanedione after administration
of the mixture only. The increased blood bioavailability
correlated with neurophysiological findings in co-exposed animals.
The results suggest that methyl ethyl ketone increases the
persistence of 2,5-hexanedione in the blood. Levels of
radiolabelled 2,5-hexanedione in neurofilament-enriched
preparations from the sciatic nerve and spinal cord were generally
unaffected by concurrent exposure to methyl ethyl ketone during the
first two weeks of treatment, and a trend towards decreased tissue
levels was evident after 3 weeks.
Reduced urinary excretion of n-hexane metabolites has been
reported in rats following exposure to mixtures of n-hexane and
methyl ethyl ketone (Perbellini et al., 1982b; Iwata et al., 1983b,
1984). The effect appeared to be transitory and no significant
effects remained after 33 weeks of co-exposure. In contrast,
increased urinary excretion of 2,5-hexanedione was observed in
guinea-pigs exposed to mixtures of methyl ethyl ketone and
2-hexanone (Couri et al., 1978).
6.2. Human Beings
The disposition and metabolism of n-hexane have been studied in
human beings following exposure by inhalation and skin contact.
Most studies were conducted on workers occupationally exposed to
commercial hexane and thus exposed to varying levels of
cyclohexane, 2-methylpentane, and 3-methylpentane, as well as
n-hexane; significant levels of methyl ethyl ketone and toluene
have also been recorded. Three experimental studies on human
volunteers presumably used pure n-hexane (Nomiyama & Nomiyama,
1974a,b; Ralston et al., 1985; Filser et al., 1987). n-Hexane is
absorbed following inhalation (Nomiyama & Nomiyama, 1974a,b;
Brugnone et al., 1978, 1980; Veulemans et al., 1982; Mutti et al.,
1984; Perbellini et al., 1985a). Steady-state pulmonary retention
(calculated by measuring the percentage of hexane in inhaled and
expired air) was in the region of 15 - 30%, and there was no
evidence of saturation at concentrations of up to 704 mg/m3 (200
ppm). Pulmonary retention was greater in more obese individuals,
and, although the alveolar uptake rate decreased during physical
exercise, the total uptake of n-hexane increased slightly as a
result of the higher lung ventilation rate. A net lung uptake of
112 mg in 8 h was reported in workers exposed to an n-hexane level
of 180 mg/m3 (51 ppm) (Perbellini et al., 1985a). Alveolar air
concentrations of n-hexane correlated with blood concentrations in
industrial workers exposed to commercial hexane (Brugnone et al.,
1984).
Some n-hexane is exhaled following cessation of exposure
(Nomiyama & Nomiyama, 1974b), and it has been suggested that this
amounts to about 10% of the net amount absorbed (Mutti et al.,
1984). Elimination was rapid and biphasic, with half-lives of 0.2
and 1.7 h. Steady-state levels of n-hexane in blood were linearly
dose dependent following inhalation of up to 704 mg/m3 (200 ppm)
(Veulemans et al., 1982). Near-plateau levels were obtained within
15 min, both in resting volunteers and in those undergoing physical
exercise. Following the end of exposure, elimination of n-hexane
from blood was rapid and biphasic; two half-lives were obtained,
one of approximately 0.2 h and the other of 1.5 - 2 h.
The percutaneous absorption of n-hexane in humans has not been
well studied, although this route of exposure has been implicated
in case reports of peripheral neuropathy (Nomiyama et al., 1973;
Takahashi et al., 1977). In a limited study, no hexane was
detected in the blood or exhaled air of a volunteer who immersed
one hand in n-hexane for 1 min (Nomiyama & Nomiyama, 1975).
Filser et al. (1987) measured an n-hexane metabolic clearance
of 2.2 litres/min at a concentration of approximately 3.52 mg/m3
(1 ppm) (steady-state concentration), and n-hexane accumulated to a
factor of 2.3. At low concentrations, the clearance of n-hexane
was not limited by saturation of metabolism but rather by the rate
of transport by the blood to the metabolic system. n-Hexane
tissue/gas partition coefficients were determined in vitro using
tissue samples obtained from autopsy cases (Perbellini et al.,
1985b). Values for heart, muscle, brain, kidney, and liver ranged
from 2.8 to 5.2, whereas the fat/air partition coefficient was 104.
The blood/air partition coefficient was 0.8. A half-life of 64 h
for n-hexane in fat has been calculated from a mathematical model
of hexane distribution (Perbellini et al., 1986).
The metabolism of n-hexane in humans is qualitatively similar
to that in the rat. 2,5-Hexanedione, 2,5-dimethylfuran, gamma-
valerolactone, and small amounts of 2-hexanol have been identified
in urine samples from workers exposed to n-hexane (Perbellini &
Brugnone, 1980; Perbellini et al., 1981a,b,c,d; Iwata et al.,
1983b; Mutti et al., 1984). All these compounds were present as
conjugates, together with some free 2,5-hexanedione and
2,5-dimethylfuran (Perbellini & Brugnone, 1980; Perbellini et al.,
1981c). The total amounts of these compounds in urine accounted
for only 15% of the estimated uptake of n-hexane (Mutti et al.,
1984). Fedtke & Bolt (1987b) have also identified 4,5-dihydroxy-2-
hexanone as a major metabolite in the urine of a male volunteer
exposed to n-hexane at a level of 764 mg/m3 (217 ppm) for 4 h.
These authors pointed out that the acid hydrolysis commonly used in
urine analysis may lead to the actual production of 2,5-hexanedione
and 2,5-dimethylfuran from 4,5-dihydroxy-2-hexanone glucuronide.
This finding needs to be taken into account for the measurement of
urinary 2,5-hexanedione in biological monitoring.
Radiolabelled carbon dioxide has been found in the exhaled air
of volunteers after an oral dose of 1-14C-2-hexanone (DiVincenzo et
al., 1978). As 2-hexanone is a metabolite of n-hexane, the latter
may also be converted to carbon dioxide and exhaled.
The end-of-shift urine concentration of n-hexane correlates
strongly with the time-weighted average (TWA) n-hexane air
concentration. Imbriani et al. (1984a,b) reported that 2 h after
exposure had ended, urinary concentrations of n-hexane were reduced
to trace levels. 2,5-Hexanedione was detected in the urine
following exposure to a time-weighted average concentration of
n-hexane in air of more than 53 mg/m3 (15 ppm) (Iwata et al.,
1983b). The end-of-shift urine concentration of 2,5-hexanedione
showed a positive correlation with both the n-hexane time-weighted
average concentration and end-of-shift blood levels of n-hexane
(Perbellini et al., 1981b, 1985a; Iwata et al., 1983b; Mutti et
al., 1984; Ahonen & Schimberg, 1988). A mean half-life of 13 to
14 h for urinary excretion of 2,5-hexanedione has been reported
(Mutti et al., 1980; Perbellini et al., 1985a). Urinary excretion
of 2,5-hexanedione was greatest 3 - 5 h after a shift. The end-of-
shift level was similar to that of the next morning and was highest
at the end of the working week. This finding, and the high
partition coefficient and calculated long half-life of n-hexane in
fat, led to the conclusion that n-hexane may accumulate in the
human body. The level of urinary 2,5-hexanedione excretion was
also positively correlated with the airborne concentration of
methyl ethyl ketone. However, it is likely that the individuals
with the highest exposure to methyl ethyl ketone also had the
highest n-hexane exposure, so that the increased urinary
2,5-hexanedione may not be the result of an effect of methyl ethyl
ketone on n-hexane metabolism. De Rosa et al. (1988) examined 20
workers exposed to n-hexane and other solvents during glueing
operations in shoe factories. The end-of-shift concentrations of
2,5-hexanedione in the urine correlated (r = 0.87) with the 8-h
time-weighted average n-hexane exposure (measured as four
sequential 2-h samples). A biological exposure index of 4.21
mg/litre was obtained for urine collection on a Thursday, but it
was indicated that the index may vary depending on the day of
analysis. Ahonen & Schimberg (1988) estimated a reference value
for 2,5-hexanedione of 10 µmol/litre (approximately 0.7 mmol/mol
creatinine), corresponding to an 8-h time-weighted average
n-hexane concentration of 176 mg/m3 (50 ppm) on the sampling day.
This value of 2,5-hexanedione represents the difference between the
post-shift and pre-shift samples.
Governa et al. (1987) detected 2,5-hexanedione and gamma-
valerolactone in the urine of all of 40 shoe factory workers
exposed to n-hexane. 2,5-Hexanedione was the major metabolite in
39 of the 40 cases. 2-Hexanol was found in 11 cases and in one
case a low level of 2-methyl-2-pentanol was detected. The level of
2,5-hexanedione and gamma-valerolactone gave a statistically
significant correlation with concomitant electroneuromyographic
changes characteristic of neuropathy and the former metabolite was
considered a suitable predictive measurement.
Ghittori et al. (1987) found that the urinary concentration
(Cu) of n-hexane in exposed workers could be expressed by the
following equation:
Cu = (0.05 x Ca) + 3.97
where Ca is the time-weighted average environmental air
concentration. The 4-h exposure urinary concentration value in
workers exposed to a time-weighted average concentration of 180
mg/m3 was 13 µg/litre, whereas the ACGIH biological equivalent
exposure limit is 9 µg/litre.
2,5-Hexanedione has also be detected in the urine (0.45
(± 0.20) mg/litre) of people apparently not exposed to n-hexane.
It was speculated that n-hexane may be produced in the body via
lipid peroxidation (Fedtke & Bolt, 1986a).
7. EFFECTS ON ORGANISMS IN THE ENVIRONMENT
There is a lack of information on the effects of n-hexane on
organisms in the environment. Sax (1984) reported that > 1000
mg/litre is required to kill 50% of exposed "aquatic organisms",
but gave no details of species or exposure. Bringmann & Kühn
(1982) gave an LC50 of > 1000 mg/litre for the waterflea Daphnia
magna. This was a static test with no measurement of actual
exposure concentration. Bobra et al. (1983) reported an LC50 for
Daphnia magna of 3.88 mg/litre, but the test was conducted over
48 h in a vessel closed with an airtight cap. However, it is
unlikely that organisms in the natural environment would ever be
exposed to n-hexane continuously over this period, because the
highly volatile n-hexane with its low water solubility would
rapidly be lost from water. Juhnke & Ludemann (1978) reported LC50
values for golden orfe (Leuciscus melanotus) of 150 - 4480
mg/litre. It appears that this was a static test without
measurement of actual exposure; the test was reported to have
lasted 48 h with "continuous aeration", and results, therefore,
should be treated with caution. Stratton & Smith (1988)
demonstrated a 50% reduction in the growth of a culture of the
green alga Chlorella pyrenoidosa in the presence of hexane at 2.66%
(v/v) (confidence limits 1.97 - 3.35%). Hexane had a knock-down
effect on the house fly (Musca domestica L.) of 7, 20, 47, and 87%
2 h after a topical application of 0.25, 0.5, 1.9, and 2 µl/insect,
respectively (Singh & Jain, 1987). The respective mortality rates
after 5 h were 0, 20, 60, and 80% and at 24 h were 0, 13, 53, and
87%.
8. EFFECTS ON EXPERIMENTAL ANIMALS AND IN VITRO TEST SYSTEMS
8.1. Single Exposures
8.1.1. Inhalation exposure
A 1-h LC50 value of 271 040 mg/m3 (77 000 ppm) has been
reported for adult Fischer male rats (Pryor et al., 1982). A 4-h
LC50 value for hexane of 260 480 mg/m3 (74 000 ppm) was reported in
Long-Evans male rats (Hine & Zuidema, 1970), but it was later
estimated that the concentration of n-hexane in the test material
was probably only about 45% (Delbrück & Kluge, 1982). In mice
(strain unspecified) exposed to pure n-hexane for 2 h, 100 mg per
litre was reported to be the minimum lethal concentration (Lazarev,
1929). Anaesthesia occurred in Swiss mice within 1 min of exposure
to an n-hexane (99% pure) concentration of 225 280 mg/m3 (64 000
ppm) and respiratory arrest occurred within 4.5 min (Swann et al.,
1974). In mice (strain unspecified) exposed to hexane, deaths
occurred at concentrations above 133 760 mg/m3 (38 000 ppm),
normally preceded by loss of postural reflexes. The toxicity of
inhaled hexane, as measured by its lethality, increased with
elevation of temperature from 20 °C to 35 °C (Babanov et al.,
1988).
Male Fischer rats exposed by inhalation to n-hexane showed
myoclonic seizures and ataxia at concentrations above 168 960 mg/m3
(48 000 ppm) (Pryor et al., 1982). Sprague-Dawley rats showed
ataxia and decreased motor activity after 25 - 30 min exposure to
concentrations between 302 720 and 316 800 mg/m3 (86 000 and 90 000
ppm) (Honma, 1983). Sedation, hypothermia, and ptosis followed
exposure of male Sprague-Dawley rats to 7040, 14 080, and 28 160 mg
n-hexane/m3 (2000, 4000, and 8000 ppm) for 8 h (Raje et al.,
1984). No acute behavioural effects were noted in rats exposed to
84 480 mg/m3 (24 000 ppm) for 10 min (Pryor et al., 1982). In
Swiss mice, light anaesthesia followed exposure to 56 320 mg/m3
(16 000 ppm) for 5 min, and deep anaesthesia, with periods of
apnoea, occurred during exposure to 112 640 mg/m3 (32 000 ppm) for
the same length of time (Swann et al., 1974).
Histopathological effects on the lung (lamellar inclusions in
type II pneumocytes) were observed in Wistar rats exposed to an
n-hexane (96 - 99% pure) level of 35 200 mg/m3 (10 000 ppm) for 4
or 8 h or to 24 640 mg/m3 (7000 ppm) for 8 h (Schnoy et al., 1982;
Schmidt et al., 1984). Light and electron microscopy revealed no
effects on intrapulmonary nerves.
Testicular lesions, characterized by degeneration of primary
spermatocytes and mild exfoliation of spermatids, were reported in
a study on Sprague-Dawley rats exposed to 17 600 mg/m3 (5000 ppm)
(99% pure) for 24 h (DeMartino et al., 1987). There were no deaths
or other manifestations of toxicity. Complete recovery had taken
place within 30 days after exposure.
8.1.2. Oral administration
Using a one-week observation period, the LD50 values for
analytical grade n-hexane were estimated to be 28.7 g/kg for older
adult (300 - 470 g) male Sprague-Dawley rats, 32.4 g/kg for young
adult males (80 - 160 g), 15.8 g/kg for 14-day-old rats, and less
than 0.7 g/kg for newborn rats (Kimura et al., 1971).
8.1.3. Dermal exposure
Signs of discomfort and incoordination, but no deaths, were
noted in New Zealand male rabbits dermally exposed under an
occlusive dressing for 4 h to 3 g hexane (45% n-hexane) per kg
(Hine & Zuidema, 1970).
8.1.4. Parenteral administration
Following intraperitoneal injection of 3.6 g n-hexane
(analytical grade) per kg, 8 out of 10 guinea-pigs died within 24 h
(Wahlberg & Boman, 1979). The approximate lethal intraperitoneal
doses of hexane were reported to be 530 mg/kg at an environmental
temperature of 36 °C, 4000 mg/kg at 8 °C, and 9100 mg/kg at 26 °C
in male and female albino Sprague-Dawley rats (Keplinger et al.,
1959). All of 10 male NMRI mice died following slow intravenous
infusion of n-hexane (purity not stated, 830 mg/kg) in a lipid
emulsion. A dose of 750 mg/kg was estimated to be the ED50 value
for loss of righting reflex (Jeppsson, 1975).
8.1.5. In vitro exposure
Gillies et al. (1980) investigated whether or not the
inhibition of sterologenesis induced by chronic ingestion of
2,5-hexanedione could also be induced in vitro. Sterologenesis
was not inhibited in sciatic nerves of rats incubated with
2,5-hexanedione (1 mmol/litre for 3 h). Although it has been shown
that 2,5-hexanedione and other compounds that cause distal
axonopathy inhibit cholesterol biosynthesis, Gillies et al. (1980)
proposed that the neurotoxicity of 2,5-hexanedione is related to
the inhibition of the biosynthesis of ubiquinone rather than that
of cholesterol. Perfusion of isolated New Zealand rabbit hearts
with an n-hexane (laboratory grade) solution of 9.6 mg/litre for
1 h reduced the force of cardiac contraction, but no effects on
heart rate or coronary blood flow were reported. Histopathological
examination showed shrinkage of myocardial fibres (Raje, 1983). The
dose for 50% inhibition (IC50) of cytochrome-P-450-dependent
benzyloxydealkylase activity by n-hexane in lung microsomes from
beta-naphthoflavone-treated rats was found to be 8.8 ± 3.2
µmol/litre (Rabovsky & Judy, 1989).
Decreased oxygen consumption at n-hexane (purity not specified)
plate concentrations of 40 µg/mg or more was reported following
incubation (15 - 38 °C) of rabbit cardiac mitochondria with 0 - 160
µg n-hexane/mg mitochondrial protein (Borgatti et al., 1981).
Adding 1.7 to 20 µl of n-hexane (pure) to a 500-µl suspension of
isolated human erythrocytes resulted in a dose-dependent increase
of specific binding to insulin. In the extracellular medium a
simultaneous increase in the amount of degraded insulin, dependent
on the concentration of n-hexane, was recorded. n-Hexane may
affect the availability of membrane receptors for hormones (Svabova
et al., 1987).
In studies by Notten & Henderson (1975a), male guinea-pigs were
given daily ip injections of n-hexane (purity not specified) of 3
or 60 mg/kg per day in sesame oil for 3 days and killed on day 4.
The livers were removed for homogenization, and the mitochondrial
supernatant was incubated with 0.1 or 0.5% (v/v) n-hexane (purity
not specified). With preparations from animals treated with 60
mg/kg per day, there was a dose-dependent increase in uridine 5'-
diphosphate (UDP) glucuronidation of p-nitrophenol, and a slight
decrease in p-hydroxylation of aniline, but no effect on
N-demethylation of aminopyrine. There was no effect on UDP
glucuronidation of p-nitrophenol when a guinea-pig microsomal
preparation, pre-activated with Triton X-100, was used (Notten &
Henderson, 1975a). In hepatic microsomal preparations (from
untreated guinea-pigs) incubated with up to 7% (v/v) n-hexane
(purity not specified) for 20 min, glucuronidation of
p-nitrophenol and o-aminophenol was increased. Aniline
p-hydroxylation was increased at n-hexane concentrations below
0.1%. N-demethylation of aminopyrine was reduced in a dose-related
manner at n-hexane concentrations above 0.5% (Notten & Henderson,
1975b).
In male Wistar rat hepatic microsome preparations incubated
with n-hexane (analytical grade) at concentrations of 0.44, 1.11,
or 2.21 mol/litre for 15 min, there was an increase in
UDP-glucuronosyl-transferase activity only at 2.21 mol/litre
(Vainio, 1974). Rabovsky et al. (1986) investigated the effect of
n-hexane (2 mmol per litre) on the activities of benzo[ a]pyrene
hydroxylase and 7-ethoxycoumarin de-ethylase in rat liver and lung
microsomes. The approximate extent of enzyme inhibition was 55%
and 30% in the case of benzopyrene hydroxylase and 50% and 35% in
the case of 7-ethoxycoumarin de-ethylase for liver and lung,
respectively. In rat hepatocytes there was a slight increase (14%)
in oxygen consumption and minimal leakage of aspartate
aminotransferase following incubation with n-hexane (20 mmol/litre)
for 1 h (Berger & Sozeri, 1987). At concentrations approaching the
solubility limit (about 0.08 mmol/litre), n-hexane selectively
stimulated the slow phase of cholesterol metabolism in isolated rat
adrenal mitochondria (McNamara & Jefcoate, 1988). This effect was
dependent on mitochondrial integrity and is apparently due to
facilitation of the transfer of cholesterol from the outer membrane
to the inner membrane where the responsible enzymes reside.
8.2. Short- and Long-Term Exposures
8.2.1. Inhalation studies
Combined groups of 10 weanling and 10 young adult male Fischer-
344 rats were exposed to an n-hexane (95% pure) concentration of
3520 mg/m3 (1000 ppm) for 24 h/day, 7 days/week, for 4 weeks and
then for 24 h/day, 6 days per week, for 7 weeks. There was reduced
weight gain, and 5 out of 10 adults died by week 14 (Howd et al.,
1983). When the same strain of rats was exposed for 24 h/day, 5
days per week, to an n-hexane (95% pure) level of 3520 mg/m3, there
was inhibition of body weight gain (Pryor et al., 1982). In both
studies, some recovery of body weight occurred after the end of
exposure. Body weight gain was reduced in Sprague-Dawley rats,
exposed for up to 6 months, 22 h/day, to an n-hexane (95% pure)
concentration of 1760 mg/m3 (500 ppm) (API, 1983a,b).
In Wistar rats exposed to 35 200 mg/m3 (10 000 ppm) n-hexane
(96 - 99% pure), 8 h/day, 7 days/week, for 15 or 19 weeks, there
was loss of body weight and some deaths occurred (Altenkirch et
al., 1978). The same results were reported for male Wistar rats
exposed to 10 563 mg/m3 (3000 ppm) n-hexane (99% pure) for 12
h/day, 7 days/week, for 16 weeks (Takeuchi et al., 1980). Reduced
body weight gain was noted in Wistar rats exposed 12 h/day for 16
weeks to 3520 mg/m3 (1000 ppm) n-hexane (99% pure) (Takeuchi et
al., 1981) and in Fischer-344 male weanling rats exposed 14 h/day,
7 days/week, for 14 weeks to n-hexane (95% pure) (Pryor et al.,
1983). A dose-related reduced weight gain was also reported in
male Wistar rats exposed to 704 or 1760 mg/m3 (200 or 500 ppm)
n-hexane (99% pure), 12 h/day, for 24 weeks (Ono et al., 1982). No
effect on body weight was noted in Sprague-Dawley rats exposed to
n-hexane (analytical grade) (443 mg/m3, 126 ppm), 21 h/day, 7
days/week, for up to 34 weeks (API, 1978), or in male Wistar rats
exposed to 373 mg/m3 (106 ppm) n-hexane, for 12 h/day, 7 days/week,
for 24 weeks (Takeuchi et al., 1983). No statistically significant
effects on body weight were reported in Wistar rats exposed to
concentrations of 352, 1056, or 3168 mg/m3 (100, 300, or 900 ppm)
n-hexane (99% pure) for 8 h/day, 5 days/week, for 72 weeks (De
Groot & Kepner, 1984).
8.2.1.1 Combined-exposure effects
The neuropathic effects of n-hexane on rats are enhanced by co-
exposure to methyl ethyl ketone. Clinical signs and histological
evidence of neuropathy developed earlier and were more severe in
male Wistar rats exposed to n-hexane (31 328 mg/m3, 8900 ppm) with
methyl ethyl ketone (MEK) (3250 mg/m3, 1100 ppm) for 8 h/day, 7
days per week, for 15 weeks, or 31 680 mg n-hexane/m3 (9000 ppm)
with 2950 mg MEK/m3 (1000 ppm) for 19 weeks than in those exposed
to 35 200 mg n-hexane/m3 (10 000 ppm) alone (Altenkirch et al.,
1978). In another study, male Wistar rats were exposed to 1760 or
2646 mg/m3 (500 or 700 ppm) n-hexane (purity not specified) or
n-hexane-MEK mixtures of 1056 mg/m3 (300 ppm) plus 590 mg/m3 (200
ppm), 1408 mg/m3 (400 ppm) plus 295 mg/m3 (100 ppm), or 1760 mg/m3
(500 ppm) plus 590 mg/m3 (Altenkirch et al., 1982). There was a
reduction in the time to onset of hindlimb paralysis in rats
exposed continuously to 1408 mg n-hexane/m3 plus 295 mg MEK/m3
compared with those exposed only to 1760 mg n-hexane/m3.
Hypersalivation occurred in rats exposed to the mixtures in both
studies. Reduced tail motor and mixed nerve conduction velocities
were reported in male Wistar rats after 20 weeks of exposure
(12 h/day) to 352 mg n-hexane/m3 (100 ppm) plus 590 mg MEK/m3 (200
ppm), but there were no toxicologically significant effects with
the same concentration of either chemical alone (Takeuchi et al.,
1983).
Methyl isobutyl ketone (MIBK) enhances n-hexane-induced
neurotoxicity in hens. Abou-Donia et al. (1985a) treated groups of
5 hens for 90 days with 3520 mg n-hexane/m3 (1000 ppm)
simultaneously with 0, 410, 1025, 2050, or 4100 mg MIBK/m3 (0, 100,
250, 500, or 1000 ppm). An untreated control group and a group
treated with MIBK (4100 mg/m3) alone were also included. The
treatments were followed by a 30-day observation period. n-Hexane
alone produced mild ataxia, while hens exposed to mixtures of
n-hexane and MIBK exhibited clinical signs of neurotoxicity
accompanied by large swollen axons and degeneration of the axon and
myelin of the spinal cord and peripheral nerves. The severity of
neurotoxicity depended on the dose of the non-neurotoxic agent
MIBK. The synergistic effect of MIBK may be related to its ability
to induce the cytochrome P-450 system responsible for the
metabolism of n-hexane to neurotoxic metabolites.
A marked increase in the clinical signs of impaired limb
function in male Wistar rats exposed to 2464 mg/m3 (700 ppm)
n-hexane (purity not specified), 23 h/day for 7 - 9 weeks, followed
pre-treatment with, and subsequent co-exposure to, lead acetate.
These effects did not occur with lead acetate alone (Wagner et al.,
1984).
n-Hexane-induced neurotoxicity can be reduced by co-exposure to
toluene. Effects on nerve conduction were less marked when rats
were exposed 12 h/day for 16 weeks to a mixture of 3696 mg/m3 (1050
ppm) n-hexane (99% pure) and 3940 mg toluene/m3 (1050 ppm) than
when they were exposed to 3696 mg n-hexane/m3 alone (Takeuchi et
al., 1981). On the other hand, Ikeda et al. (1986) showed that
co-administration of n-hexane and toluene may potentiate central
nervous system toxicity. Male Wistar rats were exposed to a
mixture of 704 mg n-hexane/m3 and 750 mg toluene/m3 (200 ppm each)
continuously for 30 days. The treatment produced a decrease in the
dopamine levels in the midbrain and hypothalamus and an increase in
the hippocampus. These changes were not found following exposure
to either solvent alone.
The effects of longer exposure (22 h/day, 7 days/week, for 6
months) to mixtures of n-hexane (1760 mg/m3, 500 ppm) and an equal
concentration of other hexane "isomers" (true isomers and related
6-carbon compounds) have been studied (Spencer, 1982; API,
1983a,b). The other hexane isomers appeared to have no effect on
the neuropathic activity of n-hexane in rats. A similar pattern of
neuropathy was seen as a result of exposure to 1760 mg n-hexane/m3
both with (at an equal concentration) or without other hexane
"isomers". There were no neuropathic changes following exposure to
440 mg n-hexane/m3 (125 ppm) either with (at 3 or 11 times the
n-hexane concentration) or without other hexane "isomers". Pellin
et al. (1988) investigated the possibility of an interactive effect
of n-hexane and organophosphorus compounds on neurotoxicity. Hens
that were pretreated with n-hexane (300 mg/kg per day ip for
7 - 15 days) showed a synergistic effect for the inhibition of
neuropathy target esterase (NTE) and acetyl- and butyryl-
cholinesterase induced by a single oral dose of tri- o-cresyl
phosphate (TOCP). At a lower dose of TOCP (20 mg/kg), which was
not neurotoxic, there was still an increase in NTE inhibition to
levels close to the minimum threshold associated with neuropathy.
8.2.1.2 Effects on the respiratory tract
In studies by Dunnick (1989) and Dunnick et al. (1989), B6C3F1
mice were exposed to 0, 1760, 3520, 14 080, and 35 200 mg
n-hexane/m3 (0, 500, 1000, 4000, and 10 000 ppm) for 6 h/day, 5
days/week, for 13 weeks, and also to 3520 mg/m3 for 22 h/day, 5
days/week, for 13 weeks. Body weight gain was reduced at 35 200
mg/m3 (6 h/day) and at 3520 mg/m3 (22 h/day). The final mean body
weights were 17% and 10% lower, respectively, for males and 6% and
0% lower, respectively, for females. Sneezing was also observed
at 35 200 mg/m3. Histopathological changes included mild
inflammatory, erosive, and regenerative lesions in the olfactory
and respiratory epithelium of the nasal cavity at 14 080 and 35 000
mg per 3 (6 h/day) and at 3520 mg/m3 (22 h/day).
In male Wistar rats exposed to n-hexane (96 - 99% pure) 8
h/day, at concentrations of 1760 mg/m3 (500 ppm) for 48 - 70 days,
2464 mg/m3 (700 ppm) for 7 days, or 35 200 mg/m3 (10 000 ppm) for
2 - 4 days, there were ultrastructural changes in the lungs. Light
microscopy did not reveal any confirmed pathological changes in the
lung tissue of animals that had been exposed over 4 - 24 h to
n-hexane or to mixtures of n-hexane and methyl ethyl ketone (MEK).
Cytoplasmic swelling and focal desquamation of alveolar epithelium
followed exposure over 2 - 7 days to 2464 - 35 200 mg/m3 (Schnoy et
al., 1982). Numerous fat-laden alveolar macrophages, degenerate
type 1 pneumocytes and increased numbers of alveolar brush cells
occurred after 7 days of exposure to 1760 mg/m3 (500 ppm). The
basic structures of alveoli and alveolar receptors were preserved.
The authors hypothesized that n-hexane alone or in combination with
methyl ethyl ketone caused fatty degeneration of the alveolar
epithelium due to interference with cellular metabolism. Under
identical test conditions axonal changes in pulmonary nerves were
reported (Schmidt et al., 1984). More pronounced effects,
including degenerative changes in the ciliary cells of small air
passages, followed exposure to mixtures of 1056 - 1760 mg
n-hexane/m3 (300 - 500 ppm) plus 295 - 590 mg MEK/m3 (100 - 200
ppm) for 24 - 89 days or 31 680 mg n-hexane/m3 (9000 ppm) plus 2950
mg MEK/m3 (1000 ppm) for 2 - 14 days. However, no treatment-related
lesions were observed in the lungs of rats (Wistar, Fischer-344,
CD) exposed to n-hexane by a variety of exposure patterns. These
included exposures to up to 35 200 mg/m3 (10 000 ppm) over 13 weeks
and 3168 mg/m3 (900 ppm) over 72 weeks (Kurita, 1974; Toxigenics,
1982; API, 1983a,b; De Groot & Kepner, 1984).
In New Zealand rabbits, severe respiratory tract lesions
followed exposures to an n-hexane (research grade) concentration of
10 560 mg/m3 (3000 ppm), 8 h/day, for 8 days (Lungarella et al.,
1980, 1984; Barni-Comparini et al., 1982). Centriacinar emphysema
and scattered microhaemorrhages were observed, lung damage being
most marked at the transition zone between the terminal bronchioles
and alveolar ducts. Focal subpleural atelectasis, and alveolar and
interstitial oedema were also observed. In another study
(Lungarella et al., 1984), 24 New Zealand male rabbits were exposed
to 10 560 mg/m3, 8 h/day, 5 days/week, for 24 weeks. Clinical
signs of ocular and upper respiratory tract irritation and
difficulty in respiration were reported, but there was no evidence
of neurotoxicity. Animals sacrificed 1 day after the last exposure
showed inflammation of the nasal turbinates and necrotic erosion of
the nasal mucosa. The lungs showed centriacinar emphysema,
scattered foci of pulmonary fibrosis, and papillary proliferations
of non-ciliated bronchiolar cells (described by the authors as
papillary tumours). In a group of rabbits retained following the
end of exposure for a further 120 days, irregular foci of cellular
proliferation, papillary proliferations of non-ciliated bronchiolar
epithelium, scattered pulmonary fibrosis, and centriacinar
emphysema were still present.
Dose-related biochemical changes, indicative of increased
pulmonary secretion and cell damage in the lungs, were reported in
lung lavage fluid of male Sprague-Dawley rats exposed to n-hexane
(purity not stated) concentrations of 1690, 4048, or 5833 mg/m3
(480, 1150, or 1657 ppm) for 6 h/day, 5 days/week, for 4 weeks
(Sahu et al., 1982). There were dose-related increases in protein,
lipid, and sialic content and in acid and alkaline phosphatase,
lactate dehydrogenase, glucose-6-phosphate dehydrogenase, and
angiotensin-converting enzyme activities. Altered enzyme activity
was reported in the lungs of New Zealand rabbits exposed to 10 560
mg n-hexane/m3 (3000 ppm) 8 h/day for 8 days (Barni-Comparini et
al., 1982). The activities of acid phosphatase, beta-
glucuronidase, lactate dehydrogenase, and glucose 6-phosphate
dehydrogenase were significantly increased.
In a study by Hadjiivanova et al. (1987), male Wistar rats were
treated with n-hexane by inhalation at a concentration of 14 995
mg/m3 (4260 ppm, 5 h/day) and pulmonary surfactant was examined at
1 and 15 days. There was a treatment-dependent moderate increase
in the phospholipids of the bronchoalveolar lavage, and an altered
relative concentration of individual phospholipids in lung tissue
homogenate. In combination with irradiation, n-hexane treatment
depleted lung tissue phospholipids due to their release in the
alveoli.
8.2.1.3 Effects on the testes
When Sprague-Dawley rats were exposed to 17 600 mg n-hexane/m3
(5000 ppm) for 16 h/day, 6 days/week, up to the development of
clear symptoms of polyneuropathy, damage to the germinal epithelium
increased with increasing exposure. Early signs were abnormalities
in primary spermatocytes including vacuolation of the cytoplasm and
nuclear pycnosis. Maturing spermatids were also affected and
showed swollen cytoplasm and multinucleated heads with vacuolation.
Sertoli cells revealed vacuolation of the cytoplasm, primarily in
the basal region, and retraction of the apical cytoplasm. Shedding
of damaged spermatocytes and spermatids into the lumen of the
tubule and their appearance in the epididymis were also reported.
Progressive damage continued after cessation of dosing, leading to
tubules devoid of all germinal cells, with the exception of some
spermatogonia, and containing only damaged Sertoli cells. The
authors could not determine whether the primary effect was on the
Sertoli cell, the germinal cells, or both. Signs of testicular
damage were clear before neuropathic symptoms developed in the rats
(DeMartino et al., 1987).
These results are comparable to testicular effects reported
after dosing orally with 2,5-hexanedione, a metabolite of
n-hexane, which is the probable causative agent for these effects
(see section 8.7). A decrease in relative testis weight was
reported in adult and weanling Fischer-344 rats 5 weeks after the
end of exposure to 3520 mg/m3 (1000 ppm) n-hexane (95% pure),
24 h/day, 7 days per week, for 4 weeks followed by the same
concentration, 24 h/day, 6 days/week, for a further 7 weeks; the
testes were not examined histologically (Howd et al., 1983). No
testicular lesions were reported in Wistar or CD rats exposed for
up to 6 months to 1760 mg/m3 (500 ppm) or 18 months to 3168 mg/m3
(900 ppm) n-hexane (99% pure) (API, 1983a,b; De Groot & Kepner,
1984). There was slight congestion in the testes of Wistar rats
following inhalation of 2992 mg/m3 (850 ppm) n-hexane (purity not
specified), 6 days/week, for 20 weeks (Kurita, 1974). There were
no testicular lesions in Fischer-344 rats exposed to 10 560,
22 880, or 35 200 mg/m3 (3000, 6500, or 10 000 ppm) n-hexane (99.5%
pure), 6 h/day, 5 days/week, for 13 weeks (Toxigenics, 1982;
Cavender et al., 1984). However, adult male Sprague-Dawley rats
continuously exposed to n-hexane for 61 days at 3520 mg/m3 (1000
ppm) showed lesions of the tubule. All germinal cells were
progressively lost leaving Sertoli cells (damaged) as the only
component of the tubule. There was no impairment of androgen
synthesis, and circulating androgen levels were not different from
controls. Simultaneous administration of 3520 mg n-hexane/m3 with
4340 mg xylene/m3 (1000 ppm) caused no adverse effects on the
testis (Nylen et al., 1989).
8.2.1.4 Other effects
Panlobular necrosis occurred in the livers of some male CD rats
exposed to n-hexane (99% pure) concentrations of 440 or 1760 mg/m3
(125 or 500 ppm) 22 h/day, 7 days/week, for up to 6 months, and
relative liver weight was increased at 1760 mg/m3 (API, 1983a).
However, no hepatic effects were reported at 1760 mg/m3 in a
further study (API, 1983b). Increased relative liver weight and
increases in hepatic microsomal protein, cytochrome P-450 and
cytochrome b5 levels have been reported in NMRI mice exposed to
88 000 to 105 600 mg/m3 (25 000 to 30 000 ppm) n-hexane (analytical
grade), 23 h/day, for up to 4 days; no histological examination was
performed (Krämer et al., 1974). Howd et al. (1983) found an
increase in relative liver weight in Fischer-344 rats exposed to
3520 mg/m3 (1000 ppm) n-hexane (95% pure), 24 h/day for 4 weeks
followed by 24 h/day (6 days/week) for 7 weeks.
Increased relative kidney weights were reported in two studies
on male CD rats exposed to 1760 mg/m3 (500 ppm) n-hexane (99% pure)
continuously for up to 6 months (API, 1983a,b). In one study,
there were traces of degenerative and regenerative changes in the
kidneys of 4/34 exposed rats (API, 1983a), but in the other no
renal lesions were reported (API, 1983b). In male Fischer-344 rats
exposed to 10 560, 22 880, or 35 200 mg/m3 (3000, 6500, or 10 000
ppm) n-hexane (99.5% pure), 6 h/day, 5 days/week, for 13 weeks,
relative kidney weights were increased in rats exposed to 22 880
and 35 200 mg/m3 and there was a reduction in urinary pH at 35 200
mg/m3, but no treatment-related renal lesions were observed
(Cavender et al., 1984).
A slight increase in giant cell numbers and haemosiderin
precipitation was reported in the spleens of male Wistar rats
exposed to 2992 mg/m3 (850 ppm) n-hexane (purity not stated)
continuously, 6 days/week, for 20 weeks (Kurita, 1974).
No significant haematological changes followed exposure to
n-hexane in studies on Fischer-344, Wistar, and CD rats and New
Zealand rabbits (Kurita, 1974; API, 1978; Rebert et al., 1982;
Cavender et al., 1984; Lungarella et al., 1984). Slight reductions
in blood haemoglobin and in red cell and total white cell counts,
and an increase in immature cells in the bone marrow were reported
in guinea-pigs exposed by inhalation to n-hexane of unstated purity
(Spagna et al., 1967). Exposures were to 4928 mg/m3 (1400 ppm), 2
h/day for 120 days, 29 920 mg/m3 (8500 ppm), 2 h/day for 60 days,
or to 149 952 mg/m3 (42 600 ppm), 2 h/day for 30 days. The
haematological effects occurred at 149 952 mg/m3 and, to a lesser
extent, at 29 920 mg/m3 but not at 4928 mg/m3.
In other studies on Fischer-344, Wistar, and CD rats, no
treatment-related lesions were reported in the spleen (or other
lymphoid organs) or salivary glands following exposures to 1760
mg/m3 (500 ppm) for 6 months, 3168 mg/m3 (900 ppm) for 72 weeks, or
35 200 mg/m3 (10 000 ppm) for 13 weeks (Toxigenics, 1982; API,
1983a,b; Cavender et al., 1984; De Groot & Kepner, 1984).
In Wistar rats exposed to n-hexane (purity not stated)
concentrations of 1760 - 2464 mg/m3 (500 - 700 ppm), 22 h/day, 7
days/week, for up to 9 weeks, there was fatty degeneration and
glandular duct widening of the parotid and salivary glands
(Altenkirch et al., 1982). The effects were more noticeable in the
rats exposed to mixtures of n-hexane and methyl ethyl ketone, and
were associated with hypersalivation.
8.2.2. Oral studies
Body weight gain was reduced in male CD rats given (by gavage)
n-hexane (99% pure) doses of 570 or 1140 mg/kg per day, 5
days/week, for 13 weeks, 4000 mg/kg per day for 17 weeks, or 4000
mg/kg per day technical grade hexane (40% n-hexane), 5 days/week,
for 13 weeks or until hindlimb paralysis was observed. Severe
hindlimb weakness or paralysis (from about day 100), tibial nerve
lesions, and atrophy of testicular germinal epithethelium were
reported in the rats given 4000 mg n-hexane/kg per day (Krasavage
et al., 1980). The tibial nerve lesions (also found in one rat
exposed to the technical grade hexane) included axonal swellings,
adaxonal myelin infolding, and paranodal myelin retraction. A
reduction in tail nerve conduction velocity was reported in male
Wistar rats given daily doses of n-hexane (purity not stated) for 4
weeks (Ono et al., 1979). Doses of 0.5 or 1 g/kg per day in olive
oil were given daily by gavage for 4 weeks and the rats were
observed for a further 4 weeks. At 8 weeks both groups showed
reduced motor and mixed nerve conduction velocities.
A slight reduction in body weight gain (but no clinical or
pathological signs of neurotoxicity or other gross pathological
effects) was reported in male CD rats given a solution (less than
1%) of commercial hexane containing 40% n-hexane as drinking-water
for 10 months (O'Donaghue et al., 1978a; Krasavage et al., 1979).
In a toxicity study in rats, five groups of 30 male and 30
female Wistar rats each received daily doses of 0, 0.04, 0.2, 1, or
5 g commercial hexane/kg body weight by gavage for 13 weeks.
Increased relative kidney weights were observed in the rats that
received 0.2, 1, or 5 g/kg, and histopathological changes occurred
in the kidneys of both male and female rats that received 5 g/kg.
Rats given 0.2, 1, or 5 g/kg also showed increased liver weights.
Changes in plasma enzymes, indicative of liver damage and elevated
cholesterol and triglyceride levels, were detected in the highest-
dose group. Also in this group, histopathological changes were
detected in the adrenals, liver, kidneys, peripheral nerves,
spleen, testes, and thymus. It was concluded that the no-observed-
effect level of technical hexane is 0.04 g/kg body weight per day
but treatment-related effects at the adjacent dose level (0.2 g/kg
per day) were slight and occurred in male rats only (Til et al.,
1989).
8.2.3. Dermal studies
No deaths and no effects on weight gain occurred in guinea-pigs
for up to one month following dermal application (sealed chamber)
of 3.5 g n-hexane (analytical grade) per kg for one week (Wahlberg
& Boman, 1979).
Abou-Donia et al. (1985b) investigated the neurotoxicity of
n-hexane, 2,5-hexanediol, and 2,5-hexanedione in hens with and
without 0-ethyl- 0-4-nitrophenyl phenylphosphonothioate (EPN).
Following a daily dermal application of n-hexane (1 mmol/kg) for 90
days followed by a 30-day observation period, leg weakness was
observed. The other two aliphatic hexacarbons were more toxic at
this dose level causing gross ataxia. Concurrent dermal
application of EPN with n-hexane or 2,5-hexanediol at the same or
different sites produced an additive neurotoxic action with
histopathological changes characteristic of EPN neurotoxicity. The
additive effect of n-hexane and 2,5-hexanediol and a potentiating
effect of 2,5-hexanedione were considered to result from the
enhancing effect of the hexacarbons on EPN absorption and/or
metabolism.
8.2.4. In vitro studies
In mouse spinal cord/dorsal root ganglion/thigh-muscle explants
incubated with n-hexane (97% pure), axonal swellings, degeneration
of central and peripheral nerve fibre, and necrosis of muscle
fibres were reported (Veronesi et al., 1983, 1984). In explants
exposed to 25, 50, 80, 100, or 250 µg n-hexane/ml for up to 8
weeks, giant axonal swellings developed at 100 µg/ml or more and
appeared first in distal, paranodal regions of large diameter
fibres (Veronesi et al., 1984). Explants exposed to 50 - 650 µg/ml
for 3 - 8 weeks showed marked changes in nerve fibres at 400 - 650
µg/ml after 4 - 5 days and progressive axonal changes at 245 - 325
µg/ml after 2 - 6 weeks (Veronesi et al., 1983). In this study,
explants removed from exposure and maintained in plain nutrient
solution for 12 - 15 weeks showed some remyelination of viable
axons. Exposure of explants to n-hexane with non-cytotoxic levels
of methyl ethyl ketone potentiated the axonal effects. Cytoplasmic
bubbling and lysosome proliferation were reported in murine
neuroblastoma cells incubated with up to 1.5% n-hexane (purity not
stated) for up to 10 days (Selkoe et al., 1978).
8.2.5. Parenteral studies
n-Hexane administered subcutaneously to mice at doses of up to
10 ml/kg was found to decrease metallothionein concentrations in
the pancreas. The maximum effect was observed 24 h after
administration. Levels had returned to normal 48 h after
administration (Onosaka et al., 1988).
In male Donryu rats given subcutaneous n-hexane (97% pure)
injections of 330 mg/kg per day, 5 days/week, for 21 weeks, there
was reduced growth rate, reduced movement, disturbed gait, and
decreased amplitude of the tail nerve action potential, but no
effect on nerve conduction velocity (Misumi & Nagano, 1984). In
male Donryu rats injected subcutaneously with n-hexane (purity not
stated; 330 mg/kg per day, 5 days/week, for 5 months), gait
disturbances and decreased sweating response to beta-methylcholine
were reported in all animals (Abe et al., 1980). Peripheral nerve
lesions were reported in Sprague-Dawley rats following subcutaneous
injections of n-hexane (99% pure) (650 - 2000 mg/kg per day, 5
days/week) for up to 35 weeks (Schaumburg & Spencer, 1976). In
male Sprague-Dawley rats given daily intraperitoneal injections of
n-hexane (purity not stated; 540 mg/kg for 5 weeks), reductions in
peak conduction velocity and duration of action potentials in the
sciatic and sural nerves, in the absence of any behavioural or
histological effects, were reported (Anderson & Dunham, 1984).
The potential for n-hexane to affect the haemopoietic system
was indicated by the significant inhibition of uptake of iron by
the bone marrow in rats that had received 1 ml n-hexane/kg (ip) on
two successive days (Goel et al., 1987). The effects of the
metabolite 2,5-hexanediol on the spleen are discussed in section
8.7.
Hepatic necrosis occurred in male rats (strain not specified)
following intraperitoneal n-hexane (analytical grade) injections of
660 mg/kg per day daily for 2 or 7 days or twice weekly for 45 days
(Goel et al., 1982). The activities of serum acetylcholinesterase
and carboxyesterase and serum levels of protein, albumin, and
cholesterol were reduced in all groups.
When female albino rats were treated with n-hexane (1 ml/kg by
intraperitoneal injection) for 1, 2, 7, and 45 days, hepatotoxicity
was evident from a loss of total hepatic sulfhydryl content and
there was a significant increase in lipid peroxidation at 1 and 2
days. There was also a decrease in microsomal drug-metabolizing
activity and microsomal glucose-6-phosphatase activity (Goel et
al., 1988).
Sclerodermatous skin changes were observed in ddy mice after 17
daily intraperitoneal injections of 3.3 mg n-hexane in 1 ml of 0.9%
saline (Yamakage & Ishikawa, 1982). The purity of the n-hexane was
not stated and sclerotic skin changes also occurred in positive
control mice.
Severe lung lesions, including oedema, cellular infiltration,
abcesses, necrosis, fibrosis, and haemorrhage, were reported in
male rabbits given daily intramuscular n-hexane (purity not stated)
injections of 660 mg/kg undiluted for 5 days or 66 mg/kg in olive
oil for 21 - 77 days (Taira, 1975).
Klimes et al. (1987) investigated the effect of intraperitoneal
injections of n-hexane (1 ml/kg daily for 7 days) on the enzymic
degradation of insulin. Insulin degradation in the liver of
treated male Wistar rats was greater than in control rat liver.
However, there was inhibition of insulin degradation in the
erythrocytes of treated rats. There was, therefore, evidence for
some potential disturbance by n-hexane of glucose tolerance. Five
daily intraperitoneal doses of n-hexane (1 g/kg) per week for 2
weeks to female Sprague-Dawley rats (a dose equivalent to one tenth
of the intraperitoneal LD50) had no effect on kidney tubular
function as shown by measurements of urinary N-acetyl-beta-D-
glucosaminidase (NAG), beta2-microglobulin and albumin (Bernard et
al., 1989).
8.3. Reproduction, Embryotoxicity, and Teratogenicity
8.3.1. Teratogenicity studies
8.3.1.1 Inhalation studies
The exposure of groups of 7 - 9 pregnant Fischer-344 rats to
3520 mg/m3 (1000 ppm) n-hexane (purity unspecified) for 6 h/day on
days 8 - 12, 12 - 16, or 8 - 16 of gestation did not result in a
significant increase in resorption rate or in the incidence of
visceral or skeletal malformations (Bus & Tyl, 1979; Bus et al.,
1979). There were slight, but not statistically significant,
increases in minor anomalies such as dilated renal pelvis and
misaligned fourth sternebra in the offspring of all the exposure
groups. No signs of maternal toxicity were reported. Groups of
17 - 20 pregnant CD rats were exposed to 327 or 1436 mg/m3 (93 or
408 ppm) n-hexane (purity not stated) for 6 h/day on days 6 - 15 of
gestation (Litton Bionetics, 1979). There was no embryotoxicity or
increase in the incidence of malformations, but small, not
statistically significant, increases in the incidence of
subcutaneous haematomas and retarded bone ossification were
observed at both exposure levels.
A group of 14 pregnant Fischer-344 rats was exposed to 3520
mg/m3 (1000 ppm) n-hexane (99% pure) for 6 h/day on days 8 - 16 of
gestation and allowed to deliver naturally on day 23 (Bus & Tyl,
1979; Bus et al., 1979). Litters were culled to 6 pups/litter and
postnatal development was followed for 7 weeks with weaning at 4
weeks. There was no statistically significant difference between
the mean litter weights of the exposed and control groups.
However, a significant transient depression in mean litter weight
occurred over the first 6 weeks and was most marked at week 3.
There were no signs of neuropathy in the pups during the 7 weeks of
observation.
In a study on the offspring of Sprague-Dawley rats exposed to
352, 7040, or 35 200 mg n-hexane/m3 (100, 2000, or 10 000 ppm), 7
h/day, from 15 days prior to conception to day 18 of gestation, no
physical malformations or effects on postnatal growth rate or age
at eye opening were reported (Howell, 1979; Howell & Cooper, 1981).
Electrophysiological measurements of visual evoked response (VER)
and inter-hemisphere evoked response (IHR) were performed on
neonatal Sprague-Dawley rats from dams exposed to 0, 352, 7040, or
35 200 mg/m3 (0, 100, 2000, or 10 000 ppm) n-hexane (purity
unknown), 7 h/day, from 15 days prior to conception to day 18 of
gestation (Howell, 1979; Howell & Cooper, 1981). There were no
abnormalities in the VER and IHR at 11, 20, and 60 days. A
significant increase in the amplitude of the early VER peaks was
observed in unanaesthetized pups aged 45 days that had been exposed
in utero to 35 200 mg/m3. However, neither of the studies was
reported in sufficient detail for an assessment of the significance
of these observations to be made.
Exposure of pregnant rats (number and strain not specified) to
1760, 2815, or 5280 mg/m3 (500, 800, or 1500 ppm) n-hexane (purity
not stated) during gestation and lactation resulted in reduced
maternal weight gain, increased resorption rates, reduced pup
weight gain, and retardation of cerebellar histogenesis and
neuronal maturation (Stoltenburg-Didinger et al., 1984).
Peripheral neuropathy (motor weakness) was reported in the dams but
not in the pups. Similar results were obtained following exposure
to a mixture of 5280 mg n-hexane/m3 and 4425 mg MEK/m3 (1500 ppm)
or to MEK alone at concentrations of 2360 (800 ppm) and 4425 mg/m3.
8.3.1.2 Oral
In a study on CD-1 mice, dams were dosed with n-hexane (99%
pure) at 0 (37 mice), 260 (13 mice), 660 (6 mice), 1320 (6 mice),
or 2200 (14 mice) mg/kg per day (in three divided doses) on days
6 - 15 of gestation and killed on day 18. There were no
embryotoxic, fetotoxic, or teratogenic effects. At the highest
dose, one dam died, and decreased weight gain was noted in
surviving dams. In a follow-up study, dams were dosed with 0 (24
mice), 2170 (24 mice), 2830 (25 mice), 7920 (34 mice), or 9900 (33
mice) mg/kg per day on days 6 - 15 of gestation and killed on day
18. A dose-related increase in maternal mortality was noted at
daily doses of 2830 mg/kg or more. A statistically significant
reduction in fetal weight was reported at daily doses of 7920 mg/kg
or more. There were no increases in the number of resorptions or
statistically significant dose-related increases in malformations
(Marks et al., 1980).
8.3.2. Fertility studies
There was no reduction in male or female fertility in a
dominant lethal study in which groups of 12 male CD-1 mice were
exposed by inhalation to 352 or 1408 mg/m3 (100 or 400 ppm)
n-hexane (purity unstated), 6 h/day, 5 days per week, for 8 weeks
and mated during the 2 weeks following exposure (Litton Bionetics,
1980).
8.4. Mutagenicity and Related End-Points
n-Hexane (> 99% pure) at concentrations of up to 100 µg/plate
was negative, both with and without metabolic activation (S-9 from
the liver of rats and hamsters pretreated with Aroclor(R)) in a
pre-incubation method using 95% ethanol as solvent and Salmonella
typhimurium strains TA1535, TA1537, TA100, and TA98 (Mortelmans et
al., 1986). An n-hexane preparation in dimethyl sulfoxide was not
mutagenic in S. typhimurium strains TA92, TA94, TA98, TA100,
TA1535, and TA1537 either in the presence or absence of liver S-9
fractions prepared from rats pre-treated with Kanechlor KC-400(R)
(Ishidate & Sofuni, 1984). No evidence of mutagenic activity was
reported in a microsuspension fluctuation assay with S. typhimurium
(McCarroll et al., 1980).
There was no evidence of mutagenic activity using n-hexane
(ultraviolet grade) in dimethyl sulfoxide in a TK +/- mouse
lymphoma (L 5178Y) forward mutation assay with plate concentrations
ranging from 80 - 200 µg/ml without S-9 fraction or 10 - 240 µg/ml
in the presence or absence of metabolic activation with rat liver
(Arochlor(R) pre-treated) S-9 fraction (Hazelton Laboratories,
1981). There were problems of cytotoxicity and possible losses of
n-hexane by evaporation. A preliminary study using ethyl acetate
as the solvent and plate concentrations of n-hexane (purity not
specified) of 400 - 3200 µg per ml (without S-9 fraction) or 200 -
1000 µg/ml (with S-9 fraction) was inconclusive. In an automated
adaptation of the S. typhimurium assay using strains TA98 and
TA100, n-hexane was found not to be mutagenic either with or
without the addition of liver S-9 fraction (Houk et al., 1989).
Lankas et al. (1978) conducted a limited investigation of the
effect of n-hexane (certified grade) on forward mutation to ouabain
resistance by Chinese hamster V79 cells at a plate concentration of
10.34 mg/litre in acetone for 2 weeks. Although a negative result
was reported in this study, only one concentration was tested
without exogenous metabolic activation.
Evidence of chromosomal damage following incubation with
undiluted n-hexane (95% pure) at plate concentrations of 66, 198,
or 330 µg/ml for 24 or 48 h was reported in in vitro studies using
a Chinese hamster fibroblast cell line (CHL) without exogenous
metabolic activation (Ishidate & Sofuni, 1984). An increase in
polyploidy, but with no increase in structural aberrations, was
noted after incubation of the cells with undiluted n-hexane (95%
pure) at a concentration of 330 µg/ml for 48 h.
In CD rats exposed to 528, 1056, or 1805 mg/m3 (150, 300, or
600 ppm) n-hexane (ultraviolet grade) vapour (6 h/day for 5
consecutive days), there was an increase in the incidence of bone
marrow cells with chromatid breaks at 528 and 1056 mg/m3 (Hazelton
Laboratories, 1981). Severe chromosomal damage with fragments and
dicentrics was reported at 2112 mg/m3 (600 ppm). In a preliminary
study using a different batch of n-hexane, an increase in the
number of aberrations per cell was reported following exposure to
352 and 1408 mg/m3 (100 and 400 ppm).
In a study on the frequency of chromosomal alterations induced
in Vicia faba root tip cells exposed to n-hexane solutions in water
at concentrations of 0.10, 0.30, 0.50, 0.75, and 1%, an increase in
chromosomal aberrations occurred at the concentration of 0.75% but
not at the other concentrations tested (Gomez-Arroyo et al., 1986).
In male Sprague-Dawley rats exposed to 17 600 mg/m3 (5000 ppm)
n-hexane vapour (99% pure) (16 h/day, 6 days per week), chromosomal
changes were observed in the germ cells (DeMartino et al., 1987).
After 2 - 4 weeks of exposure, spermatocytes showed an abnormal
arrangement of chromosomes at metaphase and chromosomal bridges at
anaphase. Spermatids had giant nuclei, probably polyploid. There
was no evidence of dominant lethality when male CD-1 mice were
exposed by inhalation to 352 or 1408 mg/m3 (100 or 400 ppm)
n-hexane (purity not stated) (6 h/day, 5 days per week for 8 weeks)
and mated during the 2 weeks following exposure (Litton Bionetics,
1980). There were no effects on the number of implantations or on
the dead implant/live implant ratio.
In studies by McCarroll et al. (1981a,b), microsuspension
(sealed microwells for 8 h) differential toxicity assays were
carried out using two bacterial species with exposure to n-hexane
(high technical grade) in the presence and absence of Arochlor(R)-
induced rat liver S-9. The findings were negative in at least
three separate trials per strain, using Escherichia coli WP2, its
DNA repair-deficient derivatives (WP2 uvr A, CM611, WP67, WP100,
and WP110), its DNA repair-deficient derivative p3478, Bacillus
subtilis H17, and its DNA repair-deficient derivative M45 (rec
assay). The B. subtilis rec assay was negative for n-hexane both
in the presence and absence of S-9 exogenous metabolic activation
(Kawachi et al., 1982). There was no increase in unscheduled DNA
synthesis in human lymphocytes exposed to n-hexane (99% pure) in 1%
dimethylsulfoxide (0.008, 0.08, and 0.8 mg/ml culture) in vitro
either with or without metabolic activation with rat liver S-9 mix.
Cytotoxicity was reported at 0.08 and 0.8 mg/ml without S-9 mix
(Perocco et al., 1983).
8.5. Carcinogenicity
8.5.1. Inhalation studies
In a 24-week study in which male New Zealand rabbits were
exposed to an n-hexane (research grade) concentration of 10 560
mg/m3 (3000 ppm) for 8 h/day, 6 days per week, papillary
proliferations of non-ciliated bronchiolar cells were reported
(Lungarella et al., 1984) (see section 8.2.1.2). The short
duration, single exposure concentration, and the use of rabbits of
only one sex present problems in interpreting this as a
carcinogenicity study.
8.5.2. Skin-painting studies
No tumours were found in 15 C17 inbred mice following daily
skin painting with n-hexane (solvent grade) and once-weekly
application (presumably for the life-time of each animal) of croton
oil (3% in liquid paraffin) as co-carcinogen (Ranadive et al., 1972).
In a tumour promotion study, 30 female mice were given a non-
carcinogenic dermal dose of 7,12-dimethylbenz[ a]-anthracene
followed by a dermal application of about 20 µl n-hexane (99.8%
pure) in cyclohexane (20 g in 100 ml) three times a week for 60
weeks (Sice, 1966). No skin tumours were reported but there was
dermal irritation. A similar study using 1-hexanol (97% pure), a
metabolite of n-hexane, and a group of 50 mice yielded similar
results. However, Bock & Chortyk (1970) found that n-hexane
induced tumours in mice previously primed with 7,12-dimethyl-
benz[ a]anthracene.
8.6. Neurotoxicity
Although effects on the central nervous system have been noted,
the principal toxic effect of n-hexane is a peripheral neuropathy.
8.6.1. Central nervous system effects
Altered latency and amplitude values for the fifth component of
the brainstem auditory-evoked response (BAER), which reflects
central nervous system activity, were reported from week 3 - 4 of
exposure, and the visual-evoked cortical response was affected
(Rebert & Sorenson, 1983). At 1760 mg/m3 (500 ppm) (the lowest
concentration investigated in this series of studies), reduced
forelimb and hindlimb grip strengths and increased latency of the
fifth BAER component were noted after 4 - 6 weeks of exposure.
There was slight recovery of the amplitude of the fifth BAER
component at the time of last measurement (15 weeks post exposure).
Neuropathological changes were seen in the spinal cord, medulla
oblongata, and cerebellar vermis of Sprague-Dawley rats exposed for
1 - 23 weeks to 1408 - 2112 mg/m3 (400 - 600 ppm) n-hexane (95%
pure) (Spencer & Schaumburg, 1977a,b).
In Fischer-344 rats exposed to 10 560, 22 880, or 35 200 mg/m3
(3000, 6500, or 10 000 ppm) n-hexane (99.5% pure) 6 h/day, 5
days/week, for 13 weeks, axonal swelling was noted in the brain
(medulla) in one out of 10 animals exposed to 35 200 mg/m3
(Cavender et al., 1984). There were no clinical signs of
neurotoxicity.
8.6.2. Peripheral nervous system effects
8.6.2.1 Clinical and morphological findings
The initial clinical signs of n-hexane-induced neuropathy in
rats are effects on the hind feet (flat foot, foot eversion),
followed by a waddling gait, an inability to extend the hindlimbs,
and symmetrical, distal hindlimb weakness. In the studies of
Dunnick (1989) and Dunnick et al. (1989) reported in section
8.2.1.2, B6C3F mice were exposed to 0, 1760, 3520, 14 080, and 35
200 mg/m3 (0, 500, 1000, 4000, and 10 000 ppm) n-hexane for 6
h/day, 5 days/week, for 13 weeks. Mice were also exposed to 3520
mg/m3 for 22 h/day, 5 days/week for 13 weeks. A decrease in
locomotor activity in female mice exposed to 3520 mg/m3 for 22
h/day or to 35 200 mg/m3 for 6 h/day was the only neurobehavioural
effect observed. Paranodal axonal swellings in tibial nerves were
observed in 3 out of 4 males and 3 out of 4 females at these
concentrations, but in none of the 4 controls. The severity of the
peripheral nerve lesion was mild and neither segmental demyelination
nor distal axonal degeneration was observed (Dunnick, 1989; Dunnick
et al., 1989). The relative insensitivity of the mouse to
alpha-diketone neurotoxicity has been related to the relatively
short length and smaller diameter of the myelinated axons in this
species compared to rats or human beings (Graham & Gottfreid,
1984). There can be progression to flaccid paralysis of the
hindlimbs and eventual quadriplegia (Spencer & Schaumburg, 1977b;
Altenkirch et al., 1978, 1982; Howd et al., 1983).
Inhalation exposure of Sprague-Dawley rats for 1 - 23 weeks to
n-hexane (98% pure) at 1408 - 2112 mg/m3 (400 - 600 ppm) produced
axonal swellings, containing numerous neurofilaments, on the
peripheral and central nervous systems (Schaumburg & Spencer, 1976;
Spencer & Schaumburg, 1977a,b; Spencer, 1982). In limb nerves,
these lesions occurred principally in long, large-diameter
myelinated fibres, but were also found in smaller myelinated and
unmyelinated fibres. The swellings in myelinated fibres were
accompanied by paranodal myelin retraction and finally distal axon
disintegration and axon loss. Remyelination and, rarely, axonal
regeneration have been observed in the limb nerves of rats
continuously exposed to 1408 - 2112 mg n-hexane/m3, but the extent
to which regeneration may occur post exposure in vivo has not been
reported (Spencer & Schaumburg, 1977a). In male Sprague-Dawley
rats exposed to n-hexane (99% pure), axonal degeneration in the
tibial nerve was observed following exposure to 8800 mg/m3 (2500
ppm) for 30 weeks or 17 600 mg/m3 (5000 ppm) for 14 weeks (Frontali
et al., 1981).
Neuropathological lesions have been reported in Sprague-Dawley
rats following exposure to n-hexane concentrations ranging between
1408 - 3520 mg/m3 (400 - 1000 ppm) (Schaumburg & Spencer, 1976;
Spencer & Schaumburg, 1977a,b; Altenkirch et al., 1982; Pryor et
al., 1982; Spencer, 1982; API, 1983a,b). Axonal swellings were
observed in hindlimb nerves (tibial nerve branches to the calf and
then plantar nerve branches) and in the central nervous system
(spinal cord, medulla oblongata, and cerebellar vermis) of Sprague-
Dawley rats exposed for 1 - 23 weeks to 1408 - 2112 mg/m3 (400 -
600 ppm) n-hexane (98.98% pure) (Spencer & Schaumberg, 1977a,b).
These occurred before the appearance of hindlimb weakness. In male
CD rats exposed to 440 or 1760 mg/m3 (125 or 500 ppm) n-hexane (99%
pure) 22 h/day, 7 days/week, axonal swelling was seen in the tibial
nerve and brain (medulla oblongata) after 2 months of exposure to
1760 mg/m3 (Spencer, 1982; API, 1983a,b). Pronounced degeneration
and loss of axons in tibial (most severe) and sciatic nerves,
lumbar spinal cord, and medulla oblongata were seen after 6 months.
There was complete recovery from the effects seen at 1760 mg/m3
(500 ppm) by 6 weeks after the end of the exposure period.
Clinical signs of hindlimb paralysis had disappeared by 22 weeks
after exposure to 3520 mg/m3 (1000 ppm), but there was only partial
recovery of hindlimb grip strength and slight recovery of latency
of tail nerve action potential.
In male Fischer-344 rats exposed by inhalation to 3520 mg/m3
(1000 ppm) n-hexane (95% pure) for 24 h/day, 5 days/week, for 11
weeks, spontaneous motor activity was inhibited (Pryor et al.,
1982). Hindlimb grip strength was diminished but there was less
effect on forelimb grip strength. Multisensory conditioned
avoidance response and pole-climbing ability decreased after 8
weeks of exposure but recovered between weeks 18 and 36. The fifth
component of the BAER showed increased latency and decreased
amplitude after 3 weeks. Latency returned to normal 15 weeks after
the end of exposure, although there was only a slight recovery of
amplitude.
Clinical signs of chronic neurotoxicity were observed after 10
weeks in male Wistar rats exposed to 10 560 mg per m3 (3000 ppm)
n-hexane (99% pure) for 12 h/day, 7 days/week, for 16 weeks
(Takeuchi et al., 1980). Following daily inhalation of 3696 mg/m3
(1050 ppm) n-hexane (99% pure) for 16 weeks (12 h/day), tail nerve
conduction velocity in male Wistar rats was reduced, but without
definite clinical signs of neurotoxicity (Takeuchi et al., 1981).
In another study at n-hexane concentrations of 722 or 1725 mg/m3
(205 or 490 ppm) for 24 weeks there were similar findings (Ono et
al., 1982). The effect was still marked 4 weeks after the end of
the 16-week exposure (Takeuchi et al., 1981). Axonal swelling and
demyelination were observed in the tail nerves after 24 weeks of
exposure to 722 mg/m3 (Ono et al., 1982). There were no
significant effects on tail nerve conduction time in rats exposed
to 373 mg/m3 (106 ppm), 12 h/day, 7 days/week, for 24 weeks; the
tail nerve of the one rat examined histologically was normal
(Takeuchi et al., 1983).
Higher no-observed-effect levels have been reported for male
Wistar rats in studies using daily 8-h exposures to n-hexane. No
effects on tail nerve structure or function were observed after 33
weeks of exposure to 1795 mg/m3 (510 ppm) n-hexane (97% pure)
(Iwata et al., 1984). There were no clinical signs of
neurotoxicity or treatment-related lesions in regions of the
nervous system considered to be vulnerable to hexacarbon
intoxication (including areas of the lumbar spinal cord and tibial
nerve) following inhalation of 1760 or 2464 mg/m3 (500 or 700 ppm)
n-hexane (purity not stated) for 9 or 40 weeks (Altenkirch et al.,
1982).
No clinical or histological evidence of n-hexane-induced
peripheral or central neurotoxicity was noted in two separate
studies following exposure of rats to 440 mg/m3 (125 ppm) for about
22 h/day, 7 days/week, for 26 - 34 weeks (API, 1978, 1983a,b;
Spencer, 1981, 1982).
8.6.2.2 Electrophysiological effects
The behavioural and neurophysiological effects of n-hexane
have been studied in rats exposed for 24 h/day, 5 - 7 days/week,
for 7 - 11 weeks (Pryor et al., 1982; Rebert et al., 1982; Howd et
al., 1983; Rebert & Sorenson, 1983). In male Fischer-344 rats,
exposed by inhalation for 24 h/day, 5 days/week, for 10 or 11 weeks
to 1760, 3520, or 5280 mg/m3 (500, 1000, or 1500 ppm) n-hexane (95%
pure), clinical signs of neuropathy, impairment of a conditioned
avoidance response, and increased latency in the action potential
of the caudal nerve were noted at the two highest exposure levels.
Grip strength was reduced in the hindlimbs and, to a lesser extent,
in the forelimbs of all exposed groups (Rebert & Sorenson, 1983).
Weanling male Fischer-344 rats exposed 14 h/day, 7 days/week,
for 14 weeks to 7040 mg/m3 (2000 ppm) n-hexane (95% pure) showed
behavioural and neurophysiological effects, including reduction of
motor activity, startle response, pole-climbing ability, avoidance
response, and grip strength, from week 2 of treatment (Pryor et
al., 1983). Tail nerve latency was prolonged from week 8, the
visual evoked cortical response was increased at 6 weeks, and there
was a reduced amplitude of the fifth BAER component from week 10.
There was no recovery of the tail nerve latency or the fifth BAER
component six weeks after exposure. The visual-evoked cortical
response had fully recovered 1 week after the end of exposure.
In a study of effects on the peripheral and central nervous
systems, no significant behavioural or neuropathological effects
were observed in Wistar rats exposed to 352, 1056, or 3168 mg/m3
(100, 300, or 900 ppm) n-hexane (99% pure) for 8 h/day, 5
days/week, for 72 weeks (De Groot & Kepner, 1984). Adult male rats
were exposed to 17 600 mg/m3 (5000 ppm) for 16 h/day, 6 days/week,
for 4 weeks. At all weekly intervals motor conduction velocity was
reduced in tail nerves and did not approach control values in a
4-week post-treatment period. Raised plasma acetylcholinesterase
levels (25 - 40%) were measured at all weekly intervals during
treatment, but these returned to control values 2 weeks after the
treatment (Bastone et al., 1987).
Exposure to commercial grade hexane (65 - 70% n-hexane) at
concentrations of 348, 950, 1936, 3626, and 6688 mg per/m3 (99,
270, 550, 1030, and 1900 ppm), for 24 h/day, 6 days/week, for one
year, caused electromyographic effects on the hindlimb muscles of
male SM-A mice (Miyagaki, 1967). The effects were indicative of
damage to the peripheral nervous system and included insertion-type
fibrillation and polyphasic discharges at 950 mg/m3 (270 ppm) or
more, spontaneous fibrillation at 3626 and 6688 mg per/m3, and
reduced interference patterns at 1936 mg per m3 or more. The minor
effect at 348 mg/m3 (mild polyphasic discharges) was not considered
to be toxicologically significant, and 348 mg/m3 was taken as the
no-observed-adverse-effect level in this study.
8.6.2.3 Grip strength
In male Sprague-Dawley rats exposed to n-hexane (99% pure) for
6 - 9 h/day, 5 days/week, at concentrations of 1740, 5280, 8800, or
17 600 mg/m3 (500, 1500, 2500, or 5000 ppm) for 14 - 30 weeks,
lesions were observed in the hindlimb nerves following inhalation
of 8800 or 17 600 mg/m3 (Frontali et al., 1981). In a study using
repeated 10-min exposures (6 - 24 exposures/day, 5 days/week),
reduced grip strength was noted in male Fischer-344 rats after
5 - 6 weeks of exposure at n-hexane (95% pure) levels of 84 480 to
168 960 mg/m3 (24 000 to 48 000 ppm) (Pryor et al., 1982).
Amplitude and latency of the fifth BAER component were affected by
exposure to 168 960 mg/m3. The amplitude decreased from week 11,
and there was no recovery after the end of exposure. There were
significant effects on grip strength and nerve conduction velocity
in Wistar rats after exposure for 8 h/day, 5 days/week, for 8 weeks
to 3168 mg hexane/m3 (900 ppm) (De Groot & Kepner, 1984).
8.7. Toxicity of n-Hexane Metabolites
The production of testicular lesions in rats following exposure
to 2,5-hexanedione and other metabolites of n-hexane has been
described by Katz et al. (1980), Krasavage et al. (1980), Cavanagh
& Bennetts (1981), Chapin et al. (1982), Anthony et al. (1983), and
Sayre et al. (1986). Studies in Fischer-344 rats given a 1%
solution of 2,5-hexanedione in drinking-water, in which severe
seminiferous epithelial degeneration and azoospermia were reported,
indicated that the Sertoli cell was probably the initial target for
2,5-hexanedione action (Chapin et al., 1982, 1983). Testicular
atrophy, induced by 2,5-hexanedione at levels of 0.5% and 1% in the
drinking-water or 200 mg/kg per day intraperitoneally, was observed
in Sprague-Dawley rats before clinical signs of neuropathy or
histological axonal swelling (O'Donoghue et al., 1978b).
In Charles River CD rats treated with 1% 2,5-hexanedione in the
drinking-water for 4 weeks, testicular toxicity, as shown by
decreased testis weight compared to controls, and enlarged and more
numerous membrane-bound vacuoles basally located in Sertoli cells,
was seen, together with occasional giant cells and chromatin
margination in spermatids. Brain and testis tubulin from treated
animals was found to be cross-linked and had a shortened nucleation
phase of assembly and a more rapid rate of elongation. Tubulin
cross-linking was postulated to play an important role in the
testicular toxicity of the gamma-diketone (Boekelheide, 1987).
When Boekelheide (1988a) treated Charles River CD rats
intraperitoneally with 4 mmol 2,5-hexanedione/kg per day, 5
days/week, for 2 weeks, no evidence of neurotoxicity was seen.
Testis weight and histology were normal 2 weeks after the start of
dosing, but large basally located Sertoli cell vacuoles were seen
in the testis at 4 weeks. At 7 weeks most of the seminiferous
tubules were normal and vacuolation of Sertoli cells was no longer
prominent. In the same study, rats were treated with 1%
2,5-hexanedione in the drinking-water for 2 or 3 weeks. Only after
3 weeks was neurotoxicity evident from an unsteady gait. Decreased
testicular weights were observed 7 weeks from the start of dosing
in both groups, and loss of germ cells in seminiferous tubules was
seen in both groups. At 27 weeks the testis weights were variable
with partial recovery to normal values, and there was variable
repopulation of seminiferous tubules. Rats were also dosed with 1%
2,5-hexanedione in the drinking-water for 5 weeks. This produced
moderate to severe clinical neurotoxicity, and basally located
vacuoles first appeared in Sertoli cells at 4 weeks. There was
progressive necrosis and generalized sloughing of germ cells. A
17-week recovery period resulted in an absence of all
postspermatogonial germ cells from the seminiferous epithelium in 3
out of 5 treated rats (Boekelheide 1988a). Under identical
conditions of treatment and recovery, pyrrole reactivity of testis
proteins increased to maximal levels within 2 weeks of treatment
and nucleation time for microtubule assembly from purified testis
tubulin was decreased at 2 weeks. This was associated with the
presence of cross-linked tubulin, which was suggested to lead to
germ cell loss (Boekelheide, 1988b).
In a study by Boekelheide & Eveleth (1988), Charles River CD
rats were treated with a total dose of 131 (± 2) mmol
2,5-hexanedione/kg at dose rates ranging from 1.9 to 6.1 mmol/kg
per day, and testicular damage was assessed 4 weeks after the
cessation of treatment. Reduced testis weight and germ cell
depletion were dependent on the rate of intoxication rather than
the total dose. The rate of intoxication also determined the
extent of testis pyrrole formation.
The metabolite 2,5-hexanediol was found to have an effect on
the spleens of female albino rats (Goel et al., 1987). Following
oral dosing with 2,5-hexanediol (960 mg/kg) daily for 7 days, the
haematocrit value, haemoglobin content, and level of neutrophils
were raised, while the level of lymphocytes was reduced relative to
controls. The same treatment led to a significantly greater uptake
of iron into the spleen and a significant inhibition of spleen
delta-aminolevulinic acid dehydratase activity.
Atrophy of the spleen, thymus, and lymph nodes has been
reported in albino rats given oral doses of 0.25, 0.5, 0.75, or 1
ml 2,5-hexanedione/kg body weight (Singh et al., 1983, 1984; Kannan
et al., 1985). Atrophy of the thymus, due to loss of cortical
cells, was severe but reversible. Immunocompetence (including
resistance to endotoxin shock) was impaired in mice. Lymphoid
lesions have been reported in rats dosed orally with
2,5-hexanedione (0.428 ml/kg) (Upreti et al., 1986).
2,5-Hexanedione is thought to be the ultimate neurotoxic
metabolite of n-hexane (Graham et al., 1982a). An intermediate
metabolite, 2,5-hexanediol, has been found to produce widespread
axonal swellings throughout the nervous system including severe
swelling in the distal optic tract (Cavanagh & Bennetts, 1981).
Axonal swellings did not progress and were reversible (Cavanagh,
1982; Jones & Cavanagh, 1982). However, there was progression to
axonal degeneration during and/or after exposure in long, large-
diameter fibres of hindlimb nerves and many spinal tracts. No
significant regeneration of peripheral nerves was seen. In a study
on cats exposed to 2-hexanone, one animal, retained for 4´ months
after exposure ceased, still showed a moderate loss of myelinated
nerve fibres and axonal degeneration in the sciatic nerve (Saida et
al., 1976). Using a sensitive automated technique for detecting
altered coordination of hindlimb movement in male Wistar rats,
Kulig et al. (1985) detected impairment of coordination 1 week
after oral treatment with 600 mg 2,5-hexanedione/kg and 2 weeks
after treatment with 250 mg/kg. The performance of the rats
treated with 600 mg/kg was still impaired relative to controls 5
weeks after treatment. In the optic tract of cats, 2,5-hexanedione
(given as a 0.5% solution in drinking-water up to a cumulative
maximum of 5.8 ml/kg; maximum of 62 days of dosing) caused axonal
swellings that were not selective for fibre size, impaired axonal
transport, and neuronal degeneration in large cells. There was
also a loss of flicker resolution (Pasternak et al., 1985).
Reduced glucose utilization and altered blood flow and glucose
uptake in various areas of the brain have been reported in rats
exposed to 2,5-hexanedione (Griffiths et al., 1981; Cooper et al.,
1985). The inferior colliculus was one of the regions most
affected in terms of reduced glucose utilization. This region is
believed to be the site where the fifth component of the brainstem
auditory evoked response (BAER) is generated and this component has
been shown to be affected in rats by n-hexane exposure (Rebert et
al., 1982; Pryor et al., 1982, 1983; Howd et al., 1983; Rebert &
Sorenson, 1983). Oral treatment of male Sprague-Dawley rats with
2,5-hexanedione (1% in drinking-water) led to increased plasma
concentrations of acetylcholinesterase at 2 and 4 weeks (p < 0.05
and < 0.01, respectively). This agrees with the observed increase
of plasma acetylcholinesterase following n-hexane inhalation
(section 8.6).
Graham et al. (1982a) suggested that the accumulation of
neurofilaments is not secondary to a defect in axonal transport
that may be brought about by impairment of energy production by
2-hexanone or 2,5-hexanedione (Sabri et al., 1979; Sabri, 1984),
but rather that the aggregation may be a primary event leading to
altered axonal transport. 2,5-Hexanedione-induced neurofilament
cross-linking was proposed as a mechanism leading to occlusion of
flow at the nodes of Ranvier, with subsequent enlargement of the
axon proximal and degenerative changes distal to this point (Graham
et al., 1982b). Polymer formation by cross-linking of nerve
protein (see section 6.1.3) was demonstrated using an analogue of
2,5-hexanedione (3,5-dimethyl-2,5-hexanedione) which underwent
pyrrole formation more rapidly (Graham et al., 1985).
DeCaprio & O'Neill (1985) treated male Wistar rats with 0.5%
2,5-hexanedione in the drinking-water for 8 weeks and found
clinical signs of neuropathy after 5 weeks of exposure.
2,5-Dimethylpyrrole adduct was detected in both serum and axonal
cytoskeletal proteins in the exposed animals. After 2 weeks of
exposure, < 1% of epsilon-amine groups were converted into pyrrole
adducts in axonal protein. The formation of proteins of high
relative molecular mass in the brain stem and spinal cord was
indicative of protein cross-linking. This was associated with a
loss of native neurofilament subunit proteins in the brain stem.
In vitro incubation of axonal protein with 2,5-hexanedione also
resulted in the formation of proteins of high relative molecular
mass. In support of the theory that pyrrole formation is a
requirement for gamma-diketone neurotoxicity, DeCaprio et al.
(1988) demonstrated that perdeuterio-2,5-hexanedione pyrrolylated
protein at only one-third of the initial rate observed with 2,5-
hexanedione, and that the deuterium analogue also caused less body
weight loss than 2,5-hexanedione treatment of male Wistar rates (5
days/week, 3.5 mmol/kg per day for 17 days or 2.5 mmol/kg per day
for 38 days). Hindlimb paralysis and neuropathological changes
were seen only in the rats treated with 2,5-hexanedione. Pyrrole
adduct concentrations were 2- to 3-fold higher in the serum and
axonal cytoskeletal proteins of 2,5-hexanedione-treated rats
compared with those treated with perdeuterio-2,5-hexanedione. The
primary isotope effect was considered to be due to the requirement
for C-H bond cleavage in pyrrole formation.
The 2,5-hexanedione analogue, 3-acetyl-2,5-hexanedione,
pyrrolated protein in vitro at an equal rate to the potent analogue
3,4-dimethyl-2,3-hexanedione but did not form protein cross-links.
Sprague-Dawley rats receiving 0.1 or 0.25 mmol 3-acetyl-2,5-
hexanedione/kg per day by intraperitoneal injection and sacrificed
at 20 and 7 days, respectively, did not show hindlimb paralysis or
neurofilament-filled axonal swellings characteristic of
gamma-diketone neurotoxicity. Thus protein cross-linking appears
to be a critical step in the neurotoxicity of pyrrolating agents
such as 2,5-hexanedione (Genter St Clair et al., 1988). In a study
of the relative effects of a range of analogues of 2,5-hexanedione,
Genter et al., (1987) found a strong correlation between the rate
of pyrrole formation, the rate of protein cross-linking in vitro
and the production of neurofilament-filled axonal swellings.
Rosenberg et al. (1987b) studied male Sprague-Dawley rats given
either the dl- or meso-diastereoisomers of 3,4-dimethyl-2,5-
hexanedione (0.2 mmol/kg per day by intraperitoneal injection).
The dl-stereoisomer produced hindlimb paralysis in one-third of the
time of the meso-stereoisomer, paralleling the relative rates of
in vitro amine pyrrolation. The more distal location of
neurofilament-filled swellings after meso-3,4-dimethyl-2,5-
hexanedione treatment was suggested to be due to the relatively
slow rate of neurofilament cross-linking. Hyperbaric oxygen
treatment made the onset of hindlimb paralysis more rapid in male
Sprague-Dawley rats after daily treatment with 2,5-hexanedione (4
mmol/kg by intraperitoneal injection, 7 days/week). Rats treated
with 2,5-hexanedione plus hyperbaric oxygen reached the end-point
of hindlimb paralysis at 19.8 (± 1.2) days compared to 23.5 (± 0.7)
days in rats treated with 2,5-hexanedione alone (p < 0.02). The
results support the role of pyrrole auto-oxidation and resultant
protein cross-linking in the neurotoxic process (Rosenberg et al.,
1987a).
A summary of a proposed molecular mechanism linking
neurofilament pyrrolylation with nerve degeneration in 2,5-
hexanedione neuropathy is shown in Fig. 2.
In studies on the rabbit and monkey (Macaca mulatta), the
urinary excretion of n-hexane metabolites was found to differ from
that of rats (Perbellini et al., 1982b). 2-Hexanol was the main
urinary metabolite during and after inhalation exposure. The other
metabolites present were 3-hexanol and 2,5-hexanedione; gamma-
valerolactone and 2,5-dimethylfuran were not detected. Following
intraperitoneal injection of n-hexane in guinea-pigs, 2-hexanol,
mainly as the glucuronide, and n-hexane itself were identified in
the urine (Couri et al., 1978).
Elimination half-lives in rat urine for individual metabolites
following inhalation of n-hexane have been determined for
2-hexanone (4 h), 3-hexanol and 2,5-hexanedione (7 h),
2,5-dimethylfuran and gamma-valerolactone (11 - 14 h), and
2-hexanol (18 h) (Perbellini et al., 1982a). The half-life for
2-hexanol in the rabbit and monkey was found to be shorter (8 h)
than in the rat.
5-Hydroxy-2-hexanone and 2,5-hexanedione were detected in
guinea-pig serum after intraperitoneal administration of n-hexane
(DiVincenzo et al., 1976). Following administration of 2-hexanol,
a major urinary metabolite of n-hexane in some species, two
additional compounds, 2-hexanone and 2,5-hexanediol, were found. A
similar metabolic profile was found following the administration of
2-hexanone, with 5-hydroxy-2-hexanone, 2,5-hexanedione, and
2-hexanol being identified as metabolites. 2,5-Hexanedione, which
is considered to be the active neurotoxic agent, can thus be formed
from n-hexane, 2-hexanol, or 2-hexanone.
6.1.2. Kinetics of 2,5-hexanedione
A marked increase in blood levels of 2,5-hexanedione on
inhalation exposure to n-hexane at 3168 or 17 600 mg/m3 (900 or
5000 ppm) for 6 - 24 h has been observed in separate studies
(Perbellini et al., 1982a; Kulig, 1983; Kulig et al., 1984). No
evidence of 2,5-hexanedione accumulation in blood was found in
Wistar rats after repeated exposures to n-hexane at 3168 mg/m3 (900
ppm), 8 h/day, for 9 days (Kulig, 1983). 2,5-Hexanedione was
completely eliminated, within 6.25 h of the end of exposure, from
the blood of Wistar rats exposed to an n-hexane level of 3168 mg/m3
(900 ppm) 8 h/day, for 72 weeks (De Groot & Kepner, 1984). There
was no significant difference in 2,5-hexanedione blood levels in
Fischer-344 rats exposed for 1 or 5 days to 3520 mg/m3 (1000 ppm)
for 6 h/day (Bus et al., 1981). In contrast, there were
significantly increased 2,5-hexanedione concentrations in the blood
of Fischer-344 rats after 3 exposures to 14 080 mg/m3 (4000 ppm)
for 8 h/day compared with a single 8-h exposure (Howd et al., 1982).
The elimination half-life for 2,5-hexanedione from blood has
been reported to vary between different strains of rat. Values for
Sprague-Dawley and Fischer-344 rats were 2.3 h and 3.9 - 6 h,
respectively, and an initial half-life of 1 h has been reported for
Wistar rats (Bus et al., 1979; Angelo & Bischoff, 1982; Howd et
al., 1982; Kulig, 1983). It has been suggested that differences in
the rates of elimination of 2,5-hexanedione from the blood may
explain the reported greater susceptibility to n-hexane-induced
neurotoxicity of Fischer-344 rats compared with Wistar rats (Kulig,
1983; Kulig et al., 1984).
In a study by Ladefoged & Perbellini (1986), 2,5-hexanedione
(50 mg/kg) was administered to five male rabbits by intravenous
injection. The pharmacokinetic data fitted a two-compartment model
in which the half-life was 42 (± 11) min (body clearance was 0.0117
(± 0.0026) ml/min per kg).
Blood concentrations of 2,5-hexanedione have been estimated
following continuous exposure of rats to n-hexane (Kulig, 1983;
Kulig et al., 1984). Marked reductions in blood 2,5-hexanedione
levels were reported following exposure to 3168 mg/m3 (900 ppm) for
3 days compared with one day. There were no further decreases
after 9 or 20 days of exposure. Smaller reductions were observed
in short-term studies after 8 or 12 weeks of exposure to 3168 mg/m3
(900 ppm) for either 8 or 24 h/day, 5 days per week, compared with
the blood levels of 2,5-hexanedione recorded after 4 weeks of
exposure to 3168 mg/m3 (900 ppm). No effects were reported in
similar short-term studies at 1056 mg/m3 (300 ppm) (Kulig, 1983).
This may be due to the enhancement of glucuronidation (a major
elimination pathway) by hexane, a phenomenon noted in both in vitro
and in vivo studies in the guinea-pig (Notten & Henderson,
1975a,b). Similar levels of 2,5-hexanedione in blood were found in
weanling and young adult rats following one week of continuous
exposure to 3520 mg/m3 (1000 ppm) (Pryor et al., 1982).
2,5-Hexanedione was detected in the blood, sciatic nerve,
brain, kidneys, liver, and lungs, but not in the testes, following
exposure of Fischer-344 rats to levels of n-hexane between 1760 and
35 200 mg/m3 (500 to 10 000 ppm) for 6 h (Baker & Rickert, 1981).
The relationship between peak tissue concentrations of
2,5-hexanedione and n-hexane exposure levels was complex. In the
kidneys, sciatic nerve, and brain, the highest concentrations were
obtained after exposure to 3520 mg/m3 (1000 ppm). In a further
study, lower tissue levels and greater elimination of
2,5-hexanedione were found in mice than in rats after a single
exposure to 3520 mg/m3 (1000 ppm) for 6 h (Baker et al., 1980).
The elimination of 2,5-hexanedione from the rat liver, kidney,
brain, and sciatic nerve was determined after exposure to 3520
mg/m3 (1000 ppm) 6 h/day, for 1 or 5 days (Bus et al., 1981).
2,5-Hexanedione was selectively retained in sciatic nerve but not
in the brain. However, comparable rates of elimination of
2,5-hexanedione from the brain, sciatic nerve, and blood were
reported in Sprague-Dawley rats given a single oral dose of
2,5-hexanedione (Iwasaki & Tsuruta, 1984). Terminal half-lives of
32 - 33 days were reported in a review of some earlier studies
(O'Donoghue, 1985).
2,5-Hexanedione has also been detected in fetal tissue, at a
level similar to that in maternal blood, following exposure of
F-344 rats to n-hexane (Bus et al., 1979). A fetal half-life of
3 h was calculated.
Less than 10% of 2,5-hexanedione in blood is bound to plasma
components (Angelo & Bischoff, 1982). A study on hens dosed orally
with 2,5-hexanedione showed that it can form a pyrrole adduct with
serum protein (DeCaprio et al., 1982).
6.1.3. In vitro studies
n-Hexane penetrates excised rat skin slowly (Tsuruta, 1977,
1982).
In rat liver and lung preparations, n-hexane has been shown to
be hydroxylated to 1-, 2-, and 3-hexanol (Frommer et al., 1974;
Walseth et al., 1982; Toftgard et al., 1984). 2-Hexanol was found
to be the predominant metabolite, and 2,5-hexanedione was also
formed in studies with rat liver microsomes (Frommer et al., 1974;
Walseth et al., 1982). More than one form of cytochrome P-450 is
involved in n-hexane oxidation (Frommer et al., 1974). In the rat
lung the formation of 2- and 3-hexanol is catalysed by a different
microsomal cytochrome P-450 isoenzyme (cytochrome P-450-PB-B) from
that responsible for the formation of 1-hexanol (Toftgard et al.,
1984). Toftgard et al. (1984, 1986) found that rat liver
microsomes, but not lung microsomes, contained a cytochrome P-450
isoenzyme that converted 2-hexanol to 2,5-hexanediol, and that the
oxidation of hexanols and 2,5-hexanediol by alcohol dehydrogenase
was restricted to the liver. These results suggest that, during
inhalation of n-hexane, the metabolite 2,5-hexanedione is likely to
be formed in the liver but not in the lung.
Preparations of purified rabbit liver cytochrome P-450
hydroxylate n-hexane, and mouse liver microsomes hydroxylate hexane
(Ichihara et al., 1969; Nilsen et al., 1981). Studies using liver
fractions from guinea-pigs demonstrated that n-hexane can be
metabolized to 2-hexanol, 2-hexanone, and 2,5-hexanedione (Couri et
al., 1978). 2-Hexanone was reduced to 2-hexanol by the cytosolic
fraction and oxidized to 2,5-hexanedione by the microsomal
fraction.
In mouse nerve and muscle tissue cultures, n-hexane was
metabolized to 2-hexanediol and 2,5-hexanedione, and 5-hydroxy-2-
hexanone was detected following incubation with 2-hexanol or
2-hexanone (Veronesi et al., 1978, 1980; Spencer et al., 1980).
2,5-Hexanedione has been found to react with primary amino
groups in proteins, such as the epsilon-amino group of lysine, to
yield 2,5-dimethylpyrrole adducts (DeCaprio et al., 1982; Graham et
al., 1982a,b). Pyrrole formation is an obligatory step in the
pathogenesis of neuropathy caused by n-hexane (Sayre et al., 1986).
2,5-Hexanedione appears to be the active neurotoxic metabolite of
n-hexane.
6.1.4. Effects of other chemicals on n-hexane metabolism
Urinary excretion of n-hexane metabolites by rats has been
shown to be increased following pre-treatment with phenobarbital,
an inducer of the microsomal oxidation of foreign compounds by
cytochrome P-450 isoenzymes (Perbellini et al., 1979, 1982a).
In in vitro studies, pre-treatment of rats with phenobarbital
increased the extent of liver microsomal oxidation of n-hexane to
2- and 3-hexanol, and pre-treatment with another inducer,
3,4-benzo[ a]pyrene, enhanced 3-hexanol formation (Frommer et al.,
1974; Näslund & Halpert, 1984). However, phenobarbital pre-
treatment of rats had no effect on n-hexane metabolism by lung
microsomes (Näslund & Halpert, 1984). Phenobarbital pre-treatment
of guinea-pigs increased the metabolism of n-hexane to 2-hexanol
and 2,5-hexanedione by hepatic post-mitochondrial supernatant, and
increased the metabolism of 2-hexanone to 2,5-hexanedione by the
hepatic microsomal fraction (Couri et al., 1978).
Intraperitoneal injection of phthalate esters to rats affects
the metabolism of n-hexane in vitro, presumably by induction of
cytochrome P-450 (Walseth et al., 1982). Treatment with
dimethylphthalate or dibutylphthalate increased significantly the
rate of formation of 2- and 3-hexanol by liver microsomes but
decreased slightly their formation by lung microsomes. A large
increase in 2,5-hexanedione formation by both types of microsomes
was noted following exposure to di(2-ethylhexyl)phthalate. Similar
results have been reported with inhalation exposures to
dibutylphthalate (Walseth & Nilsen, 1984). Male Sprague-Dawley
rats were exposed continuously for 5 days to atmospheric
dibutylphthalate concentrations of 5.7, 28.5, and 79.7 mg/m3 (0.5,
2.5, and 7 ppm), and liver and lung microsomal fractions were
prepared. The in vitro formation of n-hexane metabolites was
assayed by incubating 50 ml of 8% (v/v) n-hexane in sodium
phosphate buffer with 1 mg lung or liver microsomal protein for 10
min. The formation of 1-, 2-, and 3-hexanol was markedly increased
with liver microsomes but not with lung microsomes. At the
intermediate and high exposure concentrations of 28.5 and 79.7
mg/m3 (2.5 and 7 ppm), there was a decrease in in vitro n-hexane
metabolism particularly in lung microsomes.
The formation of 1-, 2-, and 3-hexanol from n-hexane was
significantly increased with kidney and liver microsomes obtained
from rats previously exposed to isopropanol (Zahlsen et al., 1985).
Enhanced formation of 2- and 3-hexanol (greater in male than in
female rats), but not of 1-hexanol, occurred when n-hexane was
incubated with liver microsomes from rats previously exposed to
xylene (mixed isomers) (Toftgard et al., 1983).
Treatment of rats with chloramphenicol (100 mg/kg
intraperitoneal or intravenous) inhibited 2- and 3-hexanol
formation in vitro by lung and liver microsomes derived from these
animals (Näslund & Halpert, 1984).
Toluene has been shown to be a non-competitive inhibitor of
n-hexane metabolism in in vitro studies with rat liver preparations
(Perbellini et al., 1982b). Exposure of rats to mixtures of
n-hexane and toluene resulted in reduced urinary excretion of
n-hexane metabolites (Perbellini et al., 1982b; Iwata et al.,
1983b, 1984). However, no effect on blood n-hexane levels was
reported when rats were given an intraperitoneal n-hexane injection
of 0.91 g/kg with or without 1.18 g toluene/kg (Suzuki et al.,
1974). In a study on the kinetics of 2,5-hexanedione (section
6.1.2), Ladefoged & Perbellini (1986) dosed six male rabbits
intravenously with 50 mg 2,5-hexanedione/kg with or without acetone
(150 mg/kg). A significant reduction of 2,5-hexanedione clearance
was caused by the acetone co-treatment.
Blood levels of 2,5-hexanedione were reduced after a single 6-h
co-exposure to n-hexane (3520 mg/m3, 1000 ppm) and methyl ethyl
ketone (2950 mg/m3, 1000 ppm), but not in animals pre-treated
orally with methyl ethyl ketone. However, pre-treatment resulted
in increases in liver cytochrome P-450 levels and in reactions
mediated by mixed-function oxidase (Robertson et al., 1982;
Robertson et al., 1989). Lower levels of 2,5-hexanedione have been
observed in the rat sciatic nerve after exposure to mixtures of
n-hexane and methyl ethyl ketone, but full data were not presented
(White & Bus, 1980).
In studies by Ralston et al. (1985), the blood and tissue
clearance of 2,5-hexanedione was measured after single or repeated
oral administration of 2,5-hexanedione or of a mixture of equimolar
doses of 2,5-hexanedione and methyl ethyl ketone. There was
reduced blood elimination of 2,5-hexanedione after administration
of the mixture only. The increased blood bioavailability
correlated with neurophysiological findings in co-exposed animals.
The results suggest that methyl ethyl ketone increases the
persistence of 2,5-hexanedione in the blood. Levels of
radiolabelled 2,5-hexanedione in neurofilament-enriched
preparations from the sciatic nerve and spinal cord were generally
unaffected by concurrent exposure to methyl ethyl ketone during the
first two weeks of treatment, and a trend towards decreased tissue
levels was evident after 3 weeks.
Reduced urinary excretion of n-hexane metabolites has been
reported in rats following exposure to mixtures of n-hexane and
methyl ethyl ketone (Perbellini et al., 1982b; Iwata et al., 1983b,
1984). The effect appeared to be transitory and no significant
effects remained after 33 weeks of co-exposure. In contrast,
increased urinary excretion of 2,5-hexanedione was observed in
guinea-pigs exposed to mixtures of methyl ethyl ketone and
2-hexanone (Couri et al., 1978).
6.2. Human Beings
The disposition and metabolism of n-hexane have been studied in
human beings following exposure by inhalation and skin contact.
Most studies were conducted on workers occupationally exposed to
commercial hexane and thus exposed to varying levels of
cyclohexane, 2-methylpentane, and 3-methylpentane, as well as
n-hexane; significant levels of methyl ethyl ketone and toluene
have also been recorded. Three experimental studies on human
volunteers presumably used pure n-hexane (Nomiyama & Nomiyama,
1974a,b; Ralston et al., 1985; Filser et al., 1987). n-Hexane is
absorbed following inhalation (Nomiyama & Nomiyama, 1974a,b;
Brugnone et al., 1978, 1980; Veulemans et al., 1982; Mutti et al.,
1984; Perbellini et al., 1985a). Steady-state pulmonary retention
(calculated by measuring the percentage of hexane in inhaled and
expired air) was in the region of 15 - 30%, and there was no
evidence of saturation at concentrations of up to 704 mg/m3 (200
ppm). Pulmonary retention was greater in more obese individuals,
and, although the alveolar uptake rate decreased during physical
exercise, the total uptake of n-hexane increased slightly as a
result of the higher lung ventilation rate. A net lung uptake of
112 mg in 8 h was reported in workers exposed to an n-hexane level
of 180 mg/m3 (51 ppm) (Perbellini et al., 1985a). Alveolar air
concentrations of n-hexane correlated with blood concentrations in
industrial workers exposed to commercial hexane (Brugnone et al.,
1984).
Some n-hexane is exhaled following cessation of exposure
(Nomiyama & Nomiyama, 1974b), and it has been suggested that this
amounts to about 10% of the net amount absorbed (Mutti et al.,
1984). Elimination was rapid and biphasic, with half-lives of 0.2
and 1.7 h. Steady-state levels of n-hexane in blood were linearly
dose dependent following inhalation of up to 704 mg/m3 (200 ppm)
(Veulemans et al., 1982). Near-plateau levels were obtained within
15 min, both in resting volunteers and in those undergoing physical
exercise. Following the end of exposure, elimination of n-hexane
from blood was rapid and biphasic; two half-lives were obtained,
one of approximately 0.2 h and the other of 1.5 - 2 h.
The percutaneous absorption of n-hexane in humans has not been
well studied, although this route of exposure has been implicated
in case reports of peripheral neuropathy (Nomiyama et al., 1973;
Takahashi et al., 1977). In a limited study, no hexane was
detected in the blood or exhaled air of a volunteer who immersed
one hand in n-hexane for 1 min (Nomiyama & Nomiyama, 1975).
Filser et al. (1987) measured an n-hexane metabolic clearance
of 2.2 litres/min at a concentration of approximately 3.52 mg/m3
(1 ppm) (steady-state concentration), and n-hexane accumulated to a
factor of 2.3. At low concentrations, the clearance of n-hexane
was not limited by saturation of metabolism but rather by the rate
of transport by the blood to the metabolic system. n-Hexane
tissue/gas partition coefficients were determined in vitro using
tissue samples obtained from autopsy cases (Perbellini et al.,
1985b). Values for heart, muscle, brain, kidney, and liver ranged
from 2.8 to 5.2, whereas the fat/air partition coefficient was 104.
The blood/air partition coefficient was 0.8. A half-life of 64 h
for n-hexane in fat has been calculated from a mathematical model
of hexane distribution (Perbellini et al., 1986).
The metabolism of n-hexane in humans is qualitatively similar
to that in the rat. 2,5-Hexanedione, 2,5-dimethylfuran, gamma-
valerolactone, and small amounts of 2-hexanol have been identified
in urine samples from workers exposed to n-hexane (Perbellini &
Brugnone, 1980; Perbellini et al., 1981a,b,c,d; Iwata et al.,
1983b; Mutti et al., 1984). All these compounds were present as
conjugates, together with some free 2,5-hexanedione and
2,5-dimethylfuran (Perbellini & Brugnone, 1980; Perbellini et al.,
1981c). The total amounts of these compounds in urine accounted
for only 15% of the estimated uptake of n-hexane (Mutti et al.,
1984). Fedtke & Bolt (1987b) have also identified 4,5-dihydroxy-2-
hexanone as a major metabolite in the urine of a male volunteer
exposed to n-hexane at a level of 764 mg/m3 (217 ppm) for 4 h.
These authors pointed out that the acid hydrolysis commonly used in
urine analysis may lead to the actual production of 2,5-hexanedione
and 2,5-dimethylfuran from 4,5-dihydroxy-2-hexanone glucuronide.
This finding needs to be taken into account for the measurement of
urinary 2,5-hexanedione in biological monitoring.
Radiolabelled carbon dioxide has been found in the exhaled air
of volunteers after an oral dose of 1-14C-2-hexanone (DiVincenzo et
al., 1978). As 2-hexanone is a metabolite of n-hexane, the latter
may also be converted to carbon dioxide and exhaled.
The end-of-shift urine concentration of n-hexane correlates
strongly with the time-weighted average (TWA) n-hexane air
concentration. Imbriani et al. (1984a,b) reported that 2 h after
exposure had ended, urinary concentrations of n-hexane were reduced
to trace levels. 2,5-Hexanedione was detected in the urine
following exposure to a time-weighted average concentration of
n-hexane in air of more than 53 mg/m3 (15 ppm) (Iwata et al.,
1983b). The end-of-shift urine concentration of 2,5-hexanedione
showed a positive correlation with both the n-hexane time-weighted
average concentration and end-of-shift blood levels of n-hexane
(Perbellini et al., 1981b, 1985a; Iwata et al., 1983b; Mutti et
al., 1984; Ahonen & Schimberg, 1988). A mean half-life of 13 to
14 h for urinary excretion of 2,5-hexanedione has been reported
(Mutti et al., 1980; Perbellini et al., 1985a). Urinary excretion
of 2,5-hexanedione was greatest 3 - 5 h after a shift. The end-of-
shift level was similar to that of the next morning and was highest
at the end of the working week. This finding, and the high
partition coefficient and calculated long half-life of n-hexane in
fat, led to the conclusion that n-hexane may accumulate in the
human body. The level of urinary 2,5-hexanedione excretion was
also positively correlated with the airborne concentration of
methyl ethyl ketone. However, it is likely that the individuals
with the highest exposure to methyl ethyl ketone also had the
highest n-hexane exposure, so that the increased urinary
2,5-hexanedione may not be the result of an effect of methyl ethyl
ketone on n-hexane metabolism. De Rosa et al. (1988) examined 20
workers exposed to n-hexane and other solvents during glueing
operations in shoe factories. The end-of-shift concentrations of
2,5-hexanedione in the urine correlated (r = 0.87) with the 8-h
time-weighted average n-hexane exposure (measured as four
sequential 2-h samples). A biological exposure index of 4.21
mg/litre was obtained for urine collection on a Thursday, but it
was indicated that the index may vary depending on the day of
analysis. Ahonen & Schimberg (1988) estimated a reference value
for 2,5-hexanedione of 10 µmol/litre (approximately 0.7 mmol/mol
creatinine), corresponding to an 8-h time-weighted average
n-hexane concentration of 176 mg/m3 (50 ppm) on the sampling day.
This value of 2,5-hexanedione represents the difference between the
post-shift and pre-shift samples.
Governa et al. (1987) detected 2,5-hexanedione and gamma-
valerolactone in the urine of all of 40 shoe factory workers
exposed to n-hexane. 2,5-Hexanedione was the major metabolite in
39 of the 40 cases. 2-Hexanol was found in 11 cases and in one
case a low level of 2-methyl-2-pentanol was detected. The level of
2,5-hexanedione and gamma-valerolactone gave a statistically
significant correlation with concomitant electroneuromyographic
changes characteristic of neuropathy and the former metabolite was
considered a suitable predictive measurement.
Ghittori et al. (1987) found that the urinary concentration
(Cu) of n-hexane in exposed workers could be expressed by the
following equation:
Cu = (0.05 x Ca) + 3.97
where Ca is the time-weighted average environmental air
concentration. The 4-h exposure urinary concentration value in
workers exposed to a time-weighted average concentration of 180
mg/m3 was 13 µg/litre, whereas the ACGIH biological equivalent
exposure limit is 9 µg/litre.
2,5-Hexanedione has also be detected in the urine (0.45
(± 0.20) mg/litre) of people apparently not exposed to n-hexane.
It was speculated that n-hexane may be produced in the body via
lipid peroxidation (Fedtke & Bolt, 1986a).
7. EFFECTS ON ORGANISMS IN THE ENVIRONMENT
There is a lack of information on the effects of n-hexane on
organisms in the environment. Sax (1984) reported that > 1000
mg/litre is required to kill 50% of exposed "aquatic organisms",
but gave no details of species or exposure. Bringmann & Kühn
(1982) gave an LC50 of > 1000 mg/litre for the waterflea Daphnia
magna. This was a static test with no measurement of actual
exposure concentration. Bobra et al. (1983) reported an LC50 for
Daphnia magna of 3.88 mg/litre, but the test was conducted over
48 h in a vessel closed with an airtight cap. However, it is
unlikely that organisms in the natural environment would ever be
exposed to n-hexane continuously over this period, because the
highly volatile n-hexane with its low water solubility would
rapidly be lost from water. Juhnke & Ludemann (1978) reported LC50
values for golden orfe (Leuciscus melanotus) of 150 - 4480
mg/litre. It appears that this was a static test without
measurement of actual exposure; the test was reported to have
lasted 48 h with "continuous aeration", and results, therefore,
should be treated with caution. Stratton & Smith (1988)
demonstrated a 50% reduction in the growth of a culture of the
green alga Chlorella pyrenoidosa in the presence of hexane at 2.66%
(v/v) (confidence limits 1.97 - 3.35%). Hexane had a knock-down
effect on the house fly (Musca domestica L.) of 7, 20, 47, and 87%
2 h after a topical application of 0.25, 0.5, 1.9, and 2 µl/insect,
respectively (Singh & Jain, 1987). The respective mortality rates
after 5 h were 0, 20, 60, and 80% and at 24 h were 0, 13, 53, and
87%.
8. EFFECTS ON EXPERIMENTAL ANIMALS AND IN VITRO TEST SYSTEMS
8.1. Single Exposures
8.1.1. Inhalation exposure
A 1-h LC50 value of 271 040 mg/m3 (77 000 ppm) has been
reported for adult Fischer male rats (Pryor et al., 1982). A 4-h
LC50 value for hexane of 260 480 mg/m3 (74 000 ppm) was reported in
Long-Evans male rats (Hine & Zuidema, 1970), but it was later
estimated that the concentration of n-hexane in the test material
was probably only about 45% (Delbrück & Kluge, 1982). In mice
(strain unspecified) exposed to pure n-hexane for 2 h, 100 mg per
litre was reported to be the minimum lethal concentration (Lazarev,
1929). Anaesthesia occurred in Swiss mice within 1 min of exposure
to an n-hexane (99% pure) concentration of 225 280 mg/m3 (64 000
ppm) and respiratory arrest occurred within 4.5 min (Swann et al.,
1974). In mice (strain unspecified) exposed to hexane, deaths
occurred at concentrations above 133 760 mg/m3 (38 000 ppm),
normally preceded by loss of postural reflexes. The toxicity of
inhaled hexane, as measured by its lethality, increased with
elevation of temperature from 20 °C to 35 °C (Babanov et al.,
1988).
Male Fischer rats exposed by inhalation to n-hexane showed
myoclonic seizures and ataxia at concentrations above 168 960 mg/m3
(48 000 ppm) (Pryor et al., 1982). Sprague-Dawley rats showed
ataxia and decreased motor activity after 25 - 30 min exposure to
concentrations between 302 720 and 316 800 mg/m3 (86 000 and 90 000
ppm) (Honma, 1983). Sedation, hypothermia, and ptosis followed
exposure of male Sprague-Dawley rats to 7040, 14 080, and 28 160 mg
n-hexane/m3 (2000, 4000, and 8000 ppm) for 8 h (Raje et al.,
1984). No acute behavioural effects were noted in rats exposed to
84 480 mg/m3 (24 000 ppm) for 10 min (Pryor et al., 1982). In
Swiss mice, light anaesthesia followed exposure to 56 320 mg/m3
(16 000 ppm) for 5 min, and deep anaesthesia, with periods of
apnoea, occurred during exposure to 112 640 mg/m3 (32 000 ppm) for
the same length of time (Swann et al., 1974).
Histopathological effects on the lung (lamellar inclusions in
type II pneumocytes) were observed in Wistar rats exposed to an
n-hexane (96 - 99% pure) level of 35 200 mg/m3 (10 000 ppm) for 4
or 8 h or to 24 640 mg/m3 (7000 ppm) for 8 h (Schnoy et al., 1982;
Schmidt et al., 1984). Light and electron microscopy revealed no
effects on intrapulmonary nerves.
Testicular lesions, characterized by degeneration of primary
spermatocytes and mild exfoliation of spermatids, were reported in
a study on Sprague-Dawley rats exposed to 17 600 mg/m3 (5000 ppm)
(99% pure) for 24 h (DeMartino et al., 1987). There were no deaths
or other manifestations of toxicity. Complete recovery had taken
place within 30 days after exposure.
8.1.2. Oral administration
Using a one-week observation period, the LD50 values for
analytical grade n-hexane were estimated to be 28.7 g/kg for older
adult (300 - 470 g) male Sprague-Dawley rats, 32.4 g/kg for young
adult males (80 - 160 g), 15.8 g/kg for 14-day-old rats, and less
than 0.7 g/kg for newborn rats (Kimura et al., 1971).
8.1.3. Dermal exposure
Signs of discomfort and incoordination, but no deaths, were
noted in New Zealand male rabbits dermally exposed under an
occlusive dressing for 4 h to 3 g hexane (45% n-hexane) per kg
(Hine & Zuidema, 1970).
8.1.4. Parenteral administration
Following intraperitoneal injection of 3.6 g n-hexane
(analytical grade) per kg, 8 out of 10 guinea-pigs died within 24 h
(Wahlberg & Boman, 1979). The approximate lethal intraperitoneal
doses of hexane were reported to be 530 mg/kg at an environmental
temperature of 36 °C, 4000 mg/kg at 8 °C, and 9100 mg/kg at 26 °C
in male and female albino Sprague-Dawley rats (Keplinger et al.,
1959). All of 10 male NMRI mice died following slow intravenous
infusion of n-hexane (purity not stated, 830 mg/kg) in a lipid
emulsion. A dose of 750 mg/kg was estimated to be the ED50 value
for loss of righting reflex (Jeppsson, 1975).
8.1.5. In vitro exposure
Gillies et al. (1980) investigated whether or not the
inhibition of sterologenesis induced by chronic ingestion of
2,5-hexanedione could also be induced in vitro. Sterologenesis
was not inhibited in sciatic nerves of rats incubated with
2,5-hexanedione (1 mmol/litre for 3 h). Although it has been shown
that 2,5-hexanedione and other compounds that cause distal
axonopathy inhibit cholesterol biosynthesis, Gillies et al. (1980)
proposed that the neurotoxicity of 2,5-hexanedione is related to
the inhibition of the biosynthesis of ubiquinone rather than that
of cholesterol. Perfusion of isolated New Zealand rabbit hearts
with an n-hexane (laboratory grade) solution of 9.6 mg/litre for
1 h reduced the force of cardiac contraction, but no effects on
heart rate or coronary blood flow were reported. Histopathological
examination showed shrinkage of myocardial fibres (Raje, 1983). The
dose for 50% inhibition (IC50) of cytochrome-P-450-dependent
benzyloxydealkylase activity by n-hexane in lung microsomes from
beta-naphthoflavone-treated rats was found to be 8.8 ± 3.2
µmol/litre (Rabovsky & Judy, 1989).
Decreased oxygen consumption at n-hexane (purity not specified)
plate concentrations of 40 µg/mg or more was reported following
incubation (15 - 38 °C) of rabbit cardiac mitochondria with 0 - 160
µg n-hexane/mg mitochondrial protein (Borgatti et al., 1981).
Adding 1.7 to 20 µl of n-hexane (pure) to a 500-µl suspension of
isolated human erythrocytes resulted in a dose-dependent increase
of specific binding to insulin. In the extracellular medium a
simultaneous increase in the amount of degraded insulin, dependent
on the concentration of n-hexane, was recorded. n-Hexane may
affect the availability of membrane receptors for hormones (Svabova
et al., 1987).
In studies by Notten & Henderson (1975a), male guinea-pigs were
given daily ip injections of n-hexane (purity not specified) of 3
or 60 mg/kg per day in sesame oil for 3 days and killed on day 4.
The livers were removed for homogenization, and the mitochondrial
supernatant was incubated with 0.1 or 0.5% (v/v) n-hexane (purity
not specified). With preparations from animals treated with 60
mg/kg per day, there was a dose-dependent increase in uridine 5'-
diphosphate (UDP) glucuronidation of p-nitrophenol, and a slight
decrease in p-hydroxylation of aniline, but no effect on
N-demethylation of aminopyrine. There was no effect on UDP
glucuronidation of p-nitrophenol when a guinea-pig microsomal
preparation, pre-activated with Triton X-100, was used (Notten &
Henderson, 1975a). In hepatic microsomal preparations (from
untreated guinea-pigs) incubated with up to 7% (v/v) n-hexane
(purity not specified) for 20 min, glucuronidation of
p-nitrophenol and o-aminophenol was increased. Aniline
p-hydroxylation was increased at n-hexane concentrations below
0.1%. N-demethylation of aminopyrine was reduced in a dose-related
manner at n-hexane concentrations above 0.5% (Notten & Henderson,
1975b).
In male Wistar rat hepatic microsome preparations incubated
with n-hexane (analytical grade) at concentrations of 0.44, 1.11,
or 2.21 mol/litre for 15 min, there was an increase in
UDP-glucuronosyl-transferase activity only at 2.21 mol/litre
(Vainio, 1974). Rabovsky et al. (1986) investigated the effect of
n-hexane (2 mmol per litre) on the activities of benzo[ a]pyrene
hydroxylase and 7-ethoxycoumarin de-ethylase in rat liver and lung
microsomes. The approximate extent of enzyme inhibition was 55%
and 30% in the case of benzopyrene hydroxylase and 50% and 35% in
the case of 7-ethoxycoumarin de-ethylase for liver and lung,
respectively. In rat hepatocytes there was a slight increase (14%)
in oxygen consumption and minimal leakage of aspartate
aminotransferase following incubation with n-hexane (20 mmol/litre)
for 1 h (Berger & Sozeri, 1987). At concentrations approaching the
solubility limit (about 0.08 mmol/litre), n-hexane selectively
stimulated the slow phase of cholesterol metabolism in isolated rat
adrenal mitochondria (McNamara & Jefcoate, 1988). This effect was
dependent on mitochondrial integrity and is apparently due to
facilitation of the transfer of cholesterol from the outer membrane
to the inner membrane where the responsible enzymes reside.
8.2. Short- and Long-Term Exposures
8.2.1. Inhalation studies
Combined groups of 10 weanling and 10 young adult male Fischer-
344 rats were exposed to an n-hexane (95% pure) concentration of
3520 mg/m3 (1000 ppm) for 24 h/day, 7 days/week, for 4 weeks and
then for 24 h/day, 6 days per week, for 7 weeks. There was reduced
weight gain, and 5 out of 10 adults died by week 14 (Howd et al.,
1983). When the same strain of rats was exposed for 24 h/day, 5
days per week, to an n-hexane (95% pure) level of 3520 mg/m3, there
was inhibition of body weight gain (Pryor et al., 1982). In both
studies, some recovery of body weight occurred after the end of
exposure. Body weight gain was reduced in Sprague-Dawley rats,
exposed for up to 6 months, 22 h/day, to an n-hexane (95% pure)
concentration of 1760 mg/m3 (500 ppm) (API, 1983a,b).
In Wistar rats exposed to 35 200 mg/m3 (10 000 ppm) n-hexane
(96 - 99% pure), 8 h/day, 7 days/week, for 15 or 19 weeks, there
was loss of body weight and some deaths occurred (Altenkirch et
al., 1978). The same results were reported for male Wistar rats
exposed to 10 563 mg/m3 (3000 ppm) n-hexane (99% pure) for 12
h/day, 7 days/week, for 16 weeks (Takeuchi et al., 1980). Reduced
body weight gain was noted in Wistar rats exposed 12 h/day for 16
weeks to 3520 mg/m3 (1000 ppm) n-hexane (99% pure) (Takeuchi et
al., 1981) and in Fischer-344 male weanling rats exposed 14 h/day,
7 days/week, for 14 weeks to n-hexane (95% pure) (Pryor et al.,
1983). A dose-related reduced weight gain was also reported in
male Wistar rats exposed to 704 or 1760 mg/m3 (200 or 500 ppm)
n-hexane (99% pure), 12 h/day, for 24 weeks (Ono et al., 1982). No
effect on body weight was noted in Sprague-Dawley rats exposed to
n-hexane (analytical grade) (443 mg/m3, 126 ppm), 21 h/day, 7
days/week, for up to 34 weeks (API, 1978), or in male Wistar rats
exposed to 373 mg/m3 (106 ppm) n-hexane, for 12 h/day, 7 days/week,
for 24 weeks (Takeuchi et al., 1983). No statistically significant
effects on body weight were reported in Wistar rats exposed to
concentrations of 352, 1056, or 3168 mg/m3 (100, 300, or 900 ppm)
n-hexane (99% pure) for 8 h/day, 5 days/week, for 72 weeks (De
Groot & Kepner, 1984).
8.2.1.1 Combined-exposure effects
The neuropathic effects of n-hexane on rats are enhanced by co-
exposure to methyl ethyl ketone. Clinical signs and histological
evidence of neuropathy developed earlier and were more severe in
male Wistar rats exposed to n-hexane (31 328 mg/m3, 8900 ppm) with
methyl ethyl ketone (MEK) (3250 mg/m3, 1100 ppm) for 8 h/day, 7
days per week, for 15 weeks, or 31 680 mg n-hexane/m3 (9000 ppm)
with 2950 mg MEK/m3 (1000 ppm) for 19 weeks than in those exposed
to 35 200 mg n-hexane/m3 (10 000 ppm) alone (Altenkirch et al.,
1978). In another study, male Wistar rats were exposed to 1760 or
2646 mg/m3 (500 or 700 ppm) n-hexane (purity not specified) or
n-hexane-MEK mixtures of 1056 mg/m3 (300 ppm) plus 590 mg/m3 (200
ppm), 1408 mg/m3 (400 ppm) plus 295 mg/m3 (100 ppm), or 1760 mg/m3
(500 ppm) plus 590 mg/m3 (Altenkirch et al., 1982). There was a
reduction in the time to onset of hindlimb paralysis in rats
exposed continuously to 1408 mg n-hexane/m3 plus 295 mg MEK/m3
compared with those exposed only to 1760 mg n-hexane/m3.
Hypersalivation occurred in rats exposed to the mixtures in both
studies. Reduced tail motor and mixed nerve conduction velocities
were reported in male Wistar rats after 20 weeks of exposure
(12 h/day) to 352 mg n-hexane/m3 (100 ppm) plus 590 mg MEK/m3 (200
ppm), but there were no toxicologically significant effects with
the same concentration of either chemical alone (Takeuchi et al.,
1983).
Methyl isobutyl ketone (MIBK) enhances n-hexane-induced
neurotoxicity in hens. Abou-Donia et al. (1985a) treated groups of
5 hens for 90 days with 3520 mg n-hexane/m3 (1000 ppm)
simultaneously with 0, 410, 1025, 2050, or 4100 mg MIBK/m3 (0, 100,
250, 500, or 1000 ppm). An untreated control group and a group
treated with MIBK (4100 mg/m3) alone were also included. The
treatments were followed by a 30-day observation period. n-Hexane
alone produced mild ataxia, while hens exposed to mixtures of
n-hexane and MIBK exhibited clinical signs of neurotoxicity
accompanied by large swollen axons and degeneration of the axon and
myelin of the spinal cord and peripheral nerves. The severity of
neurotoxicity depended on the dose of the non-neurotoxic agent
MIBK. The synergistic effect of MIBK may be related to its ability
to induce the cytochrome P-450 system responsible for the
metabolism of n-hexane to neurotoxic metabolites.
A marked increase in the clinical signs of impaired limb
function in male Wistar rats exposed to 2464 mg/m3 (700 ppm)
n-hexane (purity not specified), 23 h/day for 7 - 9 weeks, followed
pre-treatment with, and subsequent co-exposure to, lead acetate.
These effects did not occur with lead acetate alone (Wagner et al.,
1984).
n-Hexane-induced neurotoxicity can be reduced by co-exposure to
toluene. Effects on nerve conduction were less marked when rats
were exposed 12 h/day for 16 weeks to a mixture of 3696 mg/m3 (1050
ppm) n-hexane (99% pure) and 3940 mg toluene/m3 (1050 ppm) than
when they were exposed to 3696 mg n-hexane/m3 alone (Takeuchi et
al., 1981). On the other hand, Ikeda et al. (1986) showed that
co-administration of n-hexane and toluene may potentiate central
nervous system toxicity. Male Wistar rats were exposed to a
mixture of 704 mg n-hexane/m3 and 750 mg toluene/m3 (200 ppm each)
continuously for 30 days. The treatment produced a decrease in the
dopamine levels in the midbrain and hypothalamus and an increase in
the hippocampus. These changes were not found following exposure
to either solvent alone.
The effects of longer exposure (22 h/day, 7 days/week, for 6
months) to mixtures of n-hexane (1760 mg/m3, 500 ppm) and an equal
concentration of other hexane "isomers" (true isomers and related
6-carbon compounds) have been studied (Spencer, 1982; API,
1983a,b). The other hexane isomers appeared to have no effect on
the neuropathic activity of n-hexane in rats. A similar pattern of
neuropathy was seen as a result of exposure to 1760 mg n-hexane/m3
both with (at an equal concentration) or without other hexane
"isomers". There were no neuropathic changes following exposure to
440 mg n-hexane/m3 (125 ppm) either with (at 3 or 11 times the
n-hexane concentration) or without other hexane "isomers". Pellin
et al. (1988) investigated the possibility of an interactive effect
of n-hexane and organophosphorus compounds on neurotoxicity. Hens
that were pretreated with n-hexane (300 mg/kg per day ip for
7 - 15 days) showed a synergistic effect for the inhibition of
neuropathy target esterase (NTE) and acetyl- and butyryl-
cholinesterase induced by a single oral dose of tri- o-cresyl
phosphate (TOCP). At a lower dose of TOCP (20 mg/kg), which was
not neurotoxic, there was still an increase in NTE inhibition to
levels close to the minimum threshold associated with neuropathy.
8.2.1.2 Effects on the respiratory tract
In studies by Dunnick (1989) and Dunnick et al. (1989), B6C3F1
mice were exposed to 0, 1760, 3520, 14 080, and 35 200 mg
n-hexane/m3 (0, 500, 1000, 4000, and 10 000 ppm) for 6 h/day, 5
days/week, for 13 weeks, and also to 3520 mg/m3 for 22 h/day, 5
days/week, for 13 weeks. Body weight gain was reduced at 35 200
mg/m3 (6 h/day) and at 3520 mg/m3 (22 h/day). The final mean body
weights were 17% and 10% lower, respectively, for males and 6% and
0% lower, respectively, for females. Sneezing was also observed
at 35 200 mg/m3. Histopathological changes included mild
inflammatory, erosive, and regenerative lesions in the olfactory
and respiratory epithelium of the nasal cavity at 14 080 and 35 000
mg per 3 (6 h/day) and at 3520 mg/m3 (22 h/day).
In male Wistar rats exposed to n-hexane (96 - 99% pure) 8
h/day, at concentrations of 1760 mg/m3 (500 ppm) for 48 - 70 days,
2464 mg/m3 (700 ppm) for 7 days, or 35 200 mg/m3 (10 000 ppm) for
2 - 4 days, there were ultrastructural changes in the lungs. Light
microscopy did not reveal any confirmed pathological changes in the
lung tissue of animals that had been exposed over 4 - 24 h to
n-hexane or to mixtures of n-hexane and methyl ethyl ketone (MEK).
Cytoplasmic swelling and focal desquamation of alveolar epithelium
followed exposure over 2 - 7 days to 2464 - 35 200 mg/m3 (Schnoy et
al., 1982). Numerous fat-laden alveolar macrophages, degenerate
type 1 pneumocytes and increased numbers of alveolar brush cells
occurred after 7 days of exposure to 1760 mg/m3 (500 ppm). The
basic structures of alveoli and alveolar receptors were preserved.
The authors hypothesized that n-hexane alone or in combination with
methyl ethyl ketone caused fatty degeneration of the alveolar
epithelium due to interference with cellular metabolism. Under
identical test conditions axonal changes in pulmonary nerves were
reported (Schmidt et al., 1984). More pronounced effects,
including degenerative changes in the ciliary cells of small air
passages, followed exposure to mixtures of 1056 - 1760 mg
n-hexane/m3 (300 - 500 ppm) plus 295 - 590 mg MEK/m3 (100 - 200
ppm) for 24 - 89 days or 31 680 mg n-hexane/m3 (9000 ppm) plus 2950
mg MEK/m3 (1000 ppm) for 2 - 14 days. However, no treatment-related
lesions were observed in the lungs of rats (Wistar, Fischer-344,
CD) exposed to n-hexane by a variety of exposure patterns. These
included exposures to up to 35 200 mg/m3 (10 000 ppm) over 13 weeks
and 3168 mg/m3 (900 ppm) over 72 weeks (Kurita, 1974; Toxigenics,
1982; API, 1983a,b; De Groot & Kepner, 1984).
In New Zealand rabbits, severe respiratory tract lesions
followed exposures to an n-hexane (research grade) concentration of
10 560 mg/m3 (3000 ppm), 8 h/day, for 8 days (Lungarella et al.,
1980, 1984; Barni-Comparini et al., 1982). Centriacinar emphysema
and scattered microhaemorrhages were observed, lung damage being
most marked at the transition zone between the terminal bronchioles
and alveolar ducts. Focal subpleural atelectasis, and alveolar and
interstitial oedema were also observed. In another study
(Lungarella et al., 1984), 24 New Zealand male rabbits were exposed
to 10 560 mg/m3, 8 h/day, 5 days/week, for 24 weeks. Clinical
signs of ocular and upper respiratory tract irritation and
difficulty in respiration were reported, but there was no evidence
of neurotoxicity. Animals sacrificed 1 day after the last exposure
showed inflammation of the nasal turbinates and necrotic erosion of
the nasal mucosa. The lungs showed centriacinar emphysema,
scattered foci of pulmonary fibrosis, and papillary proliferations
of non-ciliated bronchiolar cells (described by the authors as
papillary tumours). In a group of rabbits retained following the
end of exposure for a further 120 days, irregular foci of cellular
proliferation, papillary proliferations of non-ciliated bronchiolar
epithelium, scattered pulmonary fibrosis, and centriacinar
emphysema were still present.
Dose-related biochemical changes, indicative of increased
pulmonary secretion and cell damage in the lungs, were reported in
lung lavage fluid of male Sprague-Dawley rats exposed to n-hexane
(purity not stated) concentrations of 1690, 4048, or 5833 mg/m3
(480, 1150, or 1657 ppm) for 6 h/day, 5 days/week, for 4 weeks
(Sahu et al., 1982). There were dose-related increases in protein,
lipid, and sialic content and in acid and alkaline phosphatase,
lactate dehydrogenase, glucose-6-phosphate dehydrogenase, and
angiotensin-converting enzyme activities. Altered enzyme activity
was reported in the lungs of New Zealand rabbits exposed to 10 560
mg n-hexane/m3 (3000 ppm) 8 h/day for 8 days (Barni-Comparini et
al., 1982). The activities of acid phosphatase, beta-
glucuronidase, lactate dehydrogenase, and glucose 6-phosphate
dehydrogenase were significantly increased.
In a study by Hadjiivanova et al. (1987), male Wistar rats were
treated with n-hexane by inhalation at a concentration of 14 995
mg/m3 (4260 ppm, 5 h/day) and pulmonary surfactant was examined at
1 and 15 days. There was a treatment-dependent moderate increase
in the phospholipids of the bronchoalveolar lavage, and an altered
relative concentration of individual phospholipids in lung tissue
homogenate. In combination with irradiation, n-hexane treatment
depleted lung tissue phospholipids due to their release in the
alveoli.
8.2.1.3 Effects on the testes
When Sprague-Dawley rats were exposed to 17 600 mg n-hexane/m3
(5000 ppm) for 16 h/day, 6 days/week, up to the development of
clear symptoms of polyneuropathy, damage to the germinal epithelium
increased with increasing exposure. Early signs were abnormalities
in primary spermatocytes including vacuolation of the cytoplasm and
nuclear pycnosis. Maturing spermatids were also affected and
showed swollen cytoplasm and multinucleated heads with vacuolation.
Sertoli cells revealed vacuolation of the cytoplasm, primarily in
the basal region, and retraction of the apical cytoplasm. Shedding
of damaged spermatocytes and spermatids into the lumen of the
tubule and their appearance in the epididymis were also reported.
Progressive damage continued after cessation of dosing, leading to
tubules devoid of all germinal cells, with the exception of some
spermatogonia, and containing only damaged Sertoli cells. The
authors could not determine whether the primary effect was on the
Sertoli cell, the germinal cells, or both. Signs of testicular
damage were clear before neuropathic symptoms developed in the rats
(DeMartino et al., 1987).
These results are comparable to testicular effects reported
after dosing orally with 2,5-hexanedione, a metabolite of
n-hexane, which is the probable causative agent for these effects
(see section 8.7). A decrease in relative testis weight was
reported in adult and weanling Fischer-344 rats 5 weeks after the
end of exposure to 3520 mg/m3 (1000 ppm) n-hexane (95% pure),
24 h/day, 7 days per week, for 4 weeks followed by the same
concentration, 24 h/day, 6 days/week, for a further 7 weeks; the
testes were not examined histologically (Howd et al., 1983). No
testicular lesions were reported in Wistar or CD rats exposed for
up to 6 months to 1760 mg/m3 (500 ppm) or 18 months to 3168 mg/m3
(900 ppm) n-hexane (99% pure) (API, 1983a,b; De Groot & Kepner,
1984). There was slight congestion in the testes of Wistar rats
following inhalation of 2992 mg/m3 (850 ppm) n-hexane (purity not
specified), 6 days/week, for 20 weeks (Kurita, 1974). There were
no testicular lesions in Fischer-344 rats exposed to 10 560,
22 880, or 35 200 mg/m3 (3000, 6500, or 10 000 ppm) n-hexane (99.5%
pure), 6 h/day, 5 days/week, for 13 weeks (Toxigenics, 1982;
Cavender et al., 1984). However, adult male Sprague-Dawley rats
continuously exposed to n-hexane for 61 days at 3520 mg/m3 (1000
ppm) showed lesions of the tubule. All germinal cells were
progressively lost leaving Sertoli cells (damaged) as the only
component of the tubule. There was no impairment of androgen
synthesis, and circulating androgen levels were not different from
controls. Simultaneous administration of 3520 mg n-hexane/m3 with
4340 mg xylene/m3 (1000 ppm) caused no adverse effects on the
testis (Nylen et al., 1989).
8.2.1.4 Other effects
Panlobular necrosis occurred in the livers of some male CD rats
exposed to n-hexane (99% pure) concentrations of 440 or 1760 mg/m3
(125 or 500 ppm) 22 h/day, 7 days/week, for up to 6 months, and
relative liver weight was increased at 1760 mg/m3 (API, 1983a).
However, no hepatic effects were reported at 1760 mg/m3 in a
further study (API, 1983b). Increased relative liver weight and
increases in hepatic microsomal protein, cytochrome P-450 and
cytochrome b5 levels have been reported in NMRI mice exposed to
88 000 to 105 600 mg/m3 (25 000 to 30 000 ppm) n-hexane (analytical
grade), 23 h/day, for up to 4 days; no histological examination was
performed (Krämer et al., 1974). Howd et al. (1983) found an
increase in relative liver weight in Fischer-344 rats exposed to
3520 mg/m3 (1000 ppm) n-hexane (95% pure), 24 h/day for 4 weeks
followed by 24 h/day (6 days/week) for 7 weeks.
Increased relative kidney weights were reported in two studies
on male CD rats exposed to 1760 mg/m3 (500 ppm) n-hexane (99% pure)
continuously for up to 6 months (API, 1983a,b). In one study,
there were traces of degenerative and regenerative changes in the
kidneys of 4/34 exposed rats (API, 1983a), but in the other no
renal lesions were reported (API, 1983b). In male Fischer-344 rats
exposed to 10 560, 22 880, or 35 200 mg/m3 (3000, 6500, or 10 000
ppm) n-hexane (99.5% pure), 6 h/day, 5 days/week, for 13 weeks,
relative kidney weights were increased in rats exposed to 22 880
and 35 200 mg/m3 and there was a reduction in urinary pH at 35 200
mg/m3, but no treatment-related renal lesions were observed
(Cavender et al., 1984).
A slight increase in giant cell numbers and haemosiderin
precipitation was reported in the spleens of male Wistar rats
exposed to 2992 mg/m3 (850 ppm) n-hexane (purity not stated)
continuously, 6 days/week, for 20 weeks (Kurita, 1974).
No significant haematological changes followed exposure to
n-hexane in studies on Fischer-344, Wistar, and CD rats and New
Zealand rabbits (Kurita, 1974; API, 1978; Rebert et al., 1982;
Cavender et al., 1984; Lungarella et al., 1984). Slight reductions
in blood haemoglobin and in red cell and total white cell counts,
and an increase in immature cells in the bone marrow were reported
in guinea-pigs exposed by inhalation to n-hexane of unstated purity
(Spagna et al., 1967). Exposures were to 4928 mg/m3 (1400 ppm), 2
h/day for 120 days, 29 920 mg/m3 (8500 ppm), 2 h/day for 60 days,
or to 149 952 mg/m3 (42 600 ppm), 2 h/day for 30 days. The
haematological effects occurred at 149 952 mg/m3 and, to a lesser
extent, at 29 920 mg/m3 but not at 4928 mg/m3.
In other studies on Fischer-344, Wistar, and CD rats, no
treatment-related lesions were reported in the spleen (or other
lymphoid organs) or salivary glands following exposures to 1760
mg/m3 (500 ppm) for 6 months, 3168 mg/m3 (900 ppm) for 72 weeks, or
35 200 mg/m3 (10 000 ppm) for 13 weeks (Toxigenics, 1982; API,
1983a,b; Cavender et al., 1984; De Groot & Kepner, 1984).
In Wistar rats exposed to n-hexane (purity not stated)
concentrations of 1760 - 2464 mg/m3 (500 - 700 ppm), 22 h/day, 7
days/week, for up to 9 weeks, there was fatty degeneration and
glandular duct widening of the parotid and salivary glands
(Altenkirch et al., 1982). The effects were more noticeable in the
rats exposed to mixtures of n-hexane and methyl ethyl ketone, and
were associated with hypersalivation.
8.2.2. Oral studies
Body weight gain was reduced in male CD rats given (by gavage)
n-hexane (99% pure) doses of 570 or 1140 mg/kg per day, 5
days/week, for 13 weeks, 4000 mg/kg per day for 17 weeks, or 4000
mg/kg per day technical grade hexane (40% n-hexane), 5 days/week,
for 13 weeks or until hindlimb paralysis was observed. Severe
hindlimb weakness or paralysis (from about day 100), tibial nerve
lesions, and atrophy of testicular germinal epithethelium were
reported in the rats given 4000 mg n-hexane/kg per day (Krasavage
et al., 1980). The tibial nerve lesions (also found in one rat
exposed to the technical grade hexane) included axonal swellings,
adaxonal myelin infolding, and paranodal myelin retraction. A
reduction in tail nerve conduction velocity was reported in male
Wistar rats given daily doses of n-hexane (purity not stated) for 4
weeks (Ono et al., 1979). Doses of 0.5 or 1 g/kg per day in olive
oil were given daily by gavage for 4 weeks and the rats were
observed for a further 4 weeks. At 8 weeks both groups showed
reduced motor and mixed nerve conduction velocities.
A slight reduction in body weight gain (but no clinical or
pathological signs of neurotoxicity or other gross pathological
effects) was reported in male CD rats given a solution (less than
1%) of commercial hexane containing 40% n-hexane as drinking-water
for 10 months (O'Donaghue et al., 1978a; Krasavage et al., 1979).
In a toxicity study in rats, five groups of 30 male and 30
female Wistar rats each received daily doses of 0, 0.04, 0.2, 1, or
5 g commercial hexane/kg body weight by gavage for 13 weeks.
Increased relative kidney weights were observed in the rats that
received 0.2, 1, or 5 g/kg, and histopathological changes occurred
in the kidneys of both male and female rats that received 5 g/kg.
Rats given 0.2, 1, or 5 g/kg also showed increased liver weights.
Changes in plasma enzymes, indicative of liver damage and elevated
cholesterol and triglyceride levels, were detected in the highest-
dose group. Also in this group, histopathological changes were
detected in the adrenals, liver, kidneys, peripheral nerves,
spleen, testes, and thymus. It was concluded that the no-observed-
effect level of technical hexane is 0.04 g/kg body weight per day
but treatment-related effects at the adjacent dose level (0.2 g/kg
per day) were slight and occurred in male rats only (Til et al.,
1989).
8.2.3. Dermal studies
No deaths and no effects on weight gain occurred in guinea-pigs
for up to one month following dermal application (sealed chamber)
of 3.5 g n-hexane (analytical grade) per kg for one week (Wahlberg
& Boman, 1979).
Abou-Donia et al. (1985b) investigated the neurotoxicity of
n-hexane, 2,5-hexanediol, and 2,5-hexanedione in hens with and
without 0-ethyl- 0-4-nitrophenyl phenylphosphonothioate (EPN).
Following a daily dermal application of n-hexane (1 mmol/kg) for 90
days followed by a 30-day observation period, leg weakness was
observed. The other two aliphatic hexacarbons were more toxic at
this dose level causing gross ataxia. Concurrent dermal
application of EPN with n-hexane or 2,5-hexanediol at the same or
different sites produced an additive neurotoxic action with
histopathological changes characteristic of EPN neurotoxicity. The
additive effect of n-hexane and 2,5-hexanediol and a potentiating
effect of 2,5-hexanedione were considered to result from the
enhancing effect of the hexacarbons on EPN absorption and/or
metabolism.
8.2.4. In vitro studies
In mouse spinal cord/dorsal root ganglion/thigh-muscle explants
incubated with n-hexane (97% pure), axonal swellings, degeneration
of central and peripheral nerve fibre, and necrosis of muscle
fibres were reported (Veronesi et al., 1983, 1984). In explants
exposed to 25, 50, 80, 100, or 250 µg n-hexane/ml for up to 8
weeks, giant axonal swellings developed at 100 µg/ml or more and
appeared first in distal, paranodal regions of large diameter
fibres (Veronesi et al., 1984). Explants exposed to 50 - 650 µg/ml
for 3 - 8 weeks showed marked changes in nerve fibres at 400 - 650
µg/ml after 4 - 5 days and progressive axonal changes at 245 - 325
µg/ml after 2 - 6 weeks (Veronesi et al., 1983). In this study,
explants removed from exposure and maintained in plain nutrient
solution for 12 - 15 weeks showed some remyelination of viable
axons. Exposure of explants to n-hexane with non-cytotoxic levels
of methyl ethyl ketone potentiated the axonal effects. Cytoplasmic
bubbling and lysosome proliferation were reported in murine
neuroblastoma cells incubated with up to 1.5% n-hexane (purity not
stated) for up to 10 days (Selkoe et al., 1978).
8.2.5. Parenteral studies
n-Hexane administered subcutaneously to mice at doses of up to
10 ml/kg was found to decrease metallothionein concentrations in
the pancreas. The maximum effect was observed 24 h after
administration. Levels had returned to normal 48 h after
administration (Onosaka et al., 1988).
In male Donryu rats given subcutaneous n-hexane (97% pure)
injections of 330 mg/kg per day, 5 days/week, for 21 weeks, there
was reduced growth rate, reduced movement, disturbed gait, and
decreased amplitude of the tail nerve action potential, but no
effect on nerve conduction velocity (Misumi & Nagano, 1984). In
male Donryu rats injected subcutaneously with n-hexane (purity not
stated; 330 mg/kg per day, 5 days/week, for 5 months), gait
disturbances and decreased sweating response to beta-methylcholine
were reported in all animals (Abe et al., 1980). Peripheral nerve
lesions were reported in Sprague-Dawley rats following subcutaneous
injections of n-hexane (99% pure) (650 - 2000 mg/kg per day, 5
days/week) for up to 35 weeks (Schaumburg & Spencer, 1976). In
male Sprague-Dawley rats given daily intraperitoneal injections of
n-hexane (purity not stated; 540 mg/kg for 5 weeks), reductions in
peak conduction velocity and duration of action potentials in the
sciatic and sural nerves, in the absence of any behavioural or
histological effects, were reported (Anderson & Dunham, 1984).
The potential for n-hexane to affect the haemopoietic system
was indicated by the significant inhibition of uptake of iron by
the bone marrow in rats that had received 1 ml n-hexane/kg (ip) on
two successive days (Goel et al., 1987). The effects of the
metabolite 2,5-hexanediol on the spleen are discussed in section
8.7.
Hepatic necrosis occurred in male rats (strain not specified)
following intraperitoneal n-hexane (analytical grade) injections of
660 mg/kg per day daily for 2 or 7 days or twice weekly for 45 days
(Goel et al., 1982). The activities of serum acetylcholinesterase
and carboxyesterase and serum levels of protein, albumin, and
cholesterol were reduced in all groups.
When female albino rats were treated with n-hexane (1 ml/kg by
intraperitoneal injection) for 1, 2, 7, and 45 days, hepatotoxicity
was evident from a loss of total hepatic sulfhydryl content and
there was a significant increase in lipid peroxidation at 1 and 2
days. There was also a decrease in microsomal drug-metabolizing
activity and microsomal glucose-6-phosphatase activity (Goel et
al., 1988).
Sclerodermatous skin changes were observed in ddy mice after 17
daily intraperitoneal injections of 3.3 mg n-hexane in 1 ml of 0.9%
saline (Yamakage & Ishikawa, 1982). The purity of the n-hexane was
not stated and sclerotic skin changes also occurred in positive
control mice.
Severe lung lesions, including oedema, cellular infiltration,
abcesses, necrosis, fibrosis, and haemorrhage, were reported in
male rabbits given daily intramuscular n-hexane (purity not stated)
injections of 660 mg/kg undiluted for 5 days or 66 mg/kg in olive
oil for 21 - 77 days (Taira, 1975).
Klimes et al. (1987) investigated the effect of intraperitoneal
injections of n-hexane (1 ml/kg daily for 7 days) on the enzymic
degradation of insulin. Insulin degradation in the liver of
treated male Wistar rats was greater than in control rat liver.
However, there was inhibition of insulin degradation in the
erythrocytes of treated rats. There was, therefore, evidence for
some potential disturbance by n-hexane of glucose tolerance. Five
daily intraperitoneal doses of n-hexane (1 g/kg) per week for 2
weeks to female Sprague-Dawley rats (a dose equivalent to one tenth
of the intraperitoneal LD50) had no effect on kidney tubular
function as shown by measurements of urinary N-acetyl-beta-D-
glucosaminidase (NAG), beta2-microglobulin and albumin (Bernard et
al., 1989).
8.3. Reproduction, Embryotoxicity, and Teratogenicity
8.3.1. Teratogenicity studies
8.3.1.1 Inhalation studies
The exposure of groups of 7 - 9 pregnant Fischer-344 rats to
3520 mg/m3 (1000 ppm) n-hexane (purity unspecified) for 6 h/day on
days 8 - 12, 12 - 16, or 8 - 16 of gestation did not result in a
significant increase in resorption rate or in the incidence of
visceral or skeletal malformations (Bus & Tyl, 1979; Bus et al.,
1979). There were slight, but not statistically significant,
increases in minor anomalies such as dilated renal pelvis and
misaligned fourth sternebra in the offspring of all the exposure
groups. No signs of maternal toxicity were reported. Groups of
17 - 20 pregnant CD rats were exposed to 327 or 1436 mg/m3 (93 or
408 ppm) n-hexane (purity not stated) for 6 h/day on days 6 - 15 of
gestation (Litton Bionetics, 1979). There was no embryotoxicity or
increase in the incidence of malformations, but small, not
statistically significant, increases in the incidence of
subcutaneous haematomas and retarded bone ossification were
observed at both exposure levels.
A group of 14 pregnant Fischer-344 rats was exposed to 3520
mg/m3 (1000 ppm) n-hexane (99% pure) for 6 h/day on days 8 - 16 of
gestation and allowed to deliver naturally on day 23 (Bus & Tyl,
1979; Bus et al., 1979). Litters were culled to 6 pups/litter and
postnatal development was followed for 7 weeks with weaning at 4
weeks. There was no statistically significant difference between
the mean litter weights of the exposed and control groups.
However, a significant transient depression in mean litter weight
occurred over the first 6 weeks and was most marked at week 3.
There were no signs of neuropathy in the pups during the 7 weeks of
observation.
In a study on the offspring of Sprague-Dawley rats exposed to
352, 7040, or 35 200 mg n-hexane/m3 (100, 2000, or 10 000 ppm), 7
h/day, from 15 days prior to conception to day 18 of gestation, no
physical malformations or effects on postnatal growth rate or age
at eye opening were reported (Howell, 1979; Howell & Cooper, 1981).
Electrophysiological measurements of visual evoked response (VER)
and inter-hemisphere evoked response (IHR) were performed on
neonatal Sprague-Dawley rats from dams exposed to 0, 352, 7040, or
35 200 mg/m3 (0, 100, 2000, or 10 000 ppm) n-hexane (purity
unknown), 7 h/day, from 15 days prior to conception to day 18 of
gestation (Howell, 1979; Howell & Cooper, 1981). There were no
abnormalities in the VER and IHR at 11, 20, and 60 days. A
significant increase in the amplitude of the early VER peaks was
observed in unanaesthetized pups aged 45 days that had been exposed
in utero to 35 200 mg/m3. However, neither of the studies was
reported in sufficient detail for an assessment of the significance
of these observations to be made.
Exposure of pregnant rats (number and strain not specified) to
1760, 2815, or 5280 mg/m3 (500, 800, or 1500 ppm) n-hexane (purity
not stated) during gestation and lactation resulted in reduced
maternal weight gain, increased resorption rates, reduced pup
weight gain, and retardation of cerebellar histogenesis and
neuronal maturation (Stoltenburg-Didinger et al., 1984).
Peripheral neuropathy (motor weakness) was reported in the dams but
not in the pups. Similar results were obtained following exposure
to a mixture of 5280 mg n-hexane/m3 and 4425 mg MEK/m3 (1500 ppm)
or to MEK alone at concentrations of 2360 (800 ppm) and 4425 mg/m3.
8.3.1.2 Oral
In a study on CD-1 mice, dams were dosed with n-hexane (99%
pure) at 0 (37 mice), 260 (13 mice), 660 (6 mice), 1320 (6 mice),
or 2200 (14 mice) mg/kg per day (in three divided doses) on days
6 - 15 of gestation and killed on day 18. There were no
embryotoxic, fetotoxic, or teratogenic effects. At the highest
dose, one dam died, and decreased weight gain was noted in
surviving dams. In a follow-up study, dams were dosed with 0 (24
mice), 2170 (24 mice), 2830 (25 mice), 7920 (34 mice), or 9900 (33
mice) mg/kg per day on days 6 - 15 of gestation and killed on day
18. A dose-related increase in maternal mortality was noted at
daily doses of 2830 mg/kg or more. A statistically significant
reduction in fetal weight was reported at daily doses of 7920 mg/kg
or more. There were no increases in the number of resorptions or
statistically significant dose-related increases in malformations
(Marks et al., 1980).
8.3.2. Fertility studies
There was no reduction in male or female fertility in a
dominant lethal study in which groups of 12 male CD-1 mice were
exposed by inhalation to 352 or 1408 mg/m3 (100 or 400 ppm)
n-hexane (purity unstated), 6 h/day, 5 days per week, for 8 weeks
and mated during the 2 weeks following exposure (Litton Bionetics,
1980).
8.4. Mutagenicity and Related End-Points
n-Hexane (> 99% pure) at concentrations of up to 100 µg/plate
was negative, both with and without metabolic activation (S-9 from
the liver of rats and hamsters pretreated with Aroclor(R)) in a
pre-incubation method using 95% ethanol as solvent and Salmonella
typhimurium strains TA1535, TA1537, TA100, and TA98 (Mortelmans et
al., 1986). An n-hexane preparation in dimethyl sulfoxide was not
mutagenic in S. typhimurium strains TA92, TA94, TA98, TA100,
TA1535, and TA1537 either in the presence or absence of liver S-9
fractions prepared from rats pre-treated with Kanechlor KC-400(R)
(Ishidate & Sofuni, 1984). No evidence of mutagenic activity was
reported in a microsuspension fluctuation assay with S. typhimurium
(McCarroll et al., 1980).
There was no evidence of mutagenic activity using n-hexane
(ultraviolet grade) in dimethyl sulfoxide in a TK +/- mouse
lymphoma (L 5178Y) forward mutation assay with plate concentrations
ranging from 80 - 200 µg/ml without S-9 fraction or 10 - 240 µg/ml
in the presence or absence of metabolic activation with rat liver
(Arochlor(R) pre-treated) S-9 fraction (Hazelton Laboratories,
1981). There were problems of cytotoxicity and possible losses of
n-hexane by evaporation. A preliminary study using ethyl acetate
as the solvent and plate concentrations of n-hexane (purity not
specified) of 400 - 3200 µg per ml (without S-9 fraction) or 200 -
1000 µg/ml (with S-9 fraction) was inconclusive. In an automated
adaptation of the S. typhimurium assay using strains TA98 and
TA100, n-hexane was found not to be mutagenic either with or
without the addition of liver S-9 fraction (Houk et al., 1989).
Lankas et al. (1978) conducted a limited investigation of the
effect of n-hexane (certified grade) on forward mutation to ouabain
resistance by Chinese hamster V79 cells at a plate concentration of
10.34 mg/litre in acetone for 2 weeks. Although a negative result
was reported in this study, only one concentration was tested
without exogenous metabolic activation.
Evidence of chromosomal damage following incubation with
undiluted n-hexane (95% pure) at plate concentrations of 66, 198,
or 330 µg/ml for 24 or 48 h was reported in in vitro studies using
a Chinese hamster fibroblast cell line (CHL) without exogenous
metabolic activation (Ishidate & Sofuni, 1984). An increase in
polyploidy, but with no increase in structural aberrations, was
noted after incubation of the cells with undiluted n-hexane (95%
pure) at a concentration of 330 µg/ml for 48 h.
In CD rats exposed to 528, 1056, or 1805 mg/m3 (150, 300, or
600 ppm) n-hexane (ultraviolet grade) vapour (6 h/day for 5
consecutive days), there was an increase in the incidence of bone
marrow cells with chromatid breaks at 528 and 1056 mg/m3 (Hazelton
Laboratories, 1981). Severe chromosomal damage with fragments and
dicentrics was reported at 2112 mg/m3 (600 ppm). In a preliminary
study using a different batch of n-hexane, an increase in the
number of aberrations per cell was reported following exposure to
352 and 1408 mg/m3 (100 and 400 ppm).
In a study on the frequency of chromosomal alterations induced
in Vicia faba root tip cells exposed to n-hexane solutions in water
at concentrations of 0.10, 0.30, 0.50, 0.75, and 1%, an increase in
chromosomal aberrations occurred at the concentration of 0.75% but
not at the other concentrations tested (Gomez-Arroyo et al., 1986).
In male Sprague-Dawley rats exposed to 17 600 mg/m3 (5000 ppm)
n-hexane vapour (99% pure) (16 h/day, 6 days per week), chromosomal
changes were observed in the germ cells (DeMartino et al., 1987).
After 2 - 4 weeks of exposure, spermatocytes showed an abnormal
arrangement of chromosomes at metaphase and chromosomal bridges at
anaphase. Spermatids had giant nuclei, probably polyploid. There
was no evidence of dominant lethality when male CD-1 mice were
exposed by inhalation to 352 or 1408 mg/m3 (100 or 400 ppm)
n-hexane (purity not stated) (6 h/day, 5 days per week for 8 weeks)
and mated during the 2 weeks following exposure (Litton Bionetics,
1980). There were no effects on the number of implantations or on
the dead implant/live implant ratio.
In studies by McCarroll et al. (1981a,b), microsuspension
(sealed microwells for 8 h) differential toxicity assays were
carried out using two bacterial species with exposure to n-hexane
(high technical grade) in the presence and absence of Arochlor(R)-
induced rat liver S-9. The findings were negative in at least
three separate trials per strain, using Escherichia coli WP2, its
DNA repair-deficient derivatives (WP2 uvr A, CM611, WP67, WP100,
and WP110), its DNA repair-deficient derivative p3478, Bacillus
subtilis H17, and its DNA repair-deficient derivative M45 (rec
assay). The B. subtilis rec assay was negative for n-hexane both
in the presence and absence of S-9 exogenous metabolic activation
(Kawachi et al., 1982). There was no increase in unscheduled DNA
synthesis in human lymphocytes exposed to n-hexane (99% pure) in 1%
dimethylsulfoxide (0.008, 0.08, and 0.8 mg/ml culture) in vitro
either with or without metabolic activation with rat liver S-9 mix.
Cytotoxicity was reported at 0.08 and 0.8 mg/ml without S-9 mix
(Perocco et al., 1983).
8.5. Carcinogenicity
8.5.1. Inhalation studies
In a 24-week study in which male New Zealand rabbits were
exposed to an n-hexane (research grade) concentration of 10 560
mg/m3 (3000 ppm) for 8 h/day, 6 days per week, papillary
proliferations of non-ciliated bronchiolar cells were reported
(Lungarella et al., 1984) (see section 8.2.1.2). The short
duration, single exposure concentration, and the use of rabbits of
only one sex present problems in interpreting this as a
carcinogenicity study.
8.5.2. Skin-painting studies
No tumours were found in 15 C17 inbred mice following daily
skin painting with n-hexane (solvent grade) and once-weekly
application (presumably for the life-time of each animal) of croton
oil (3% in liquid paraffin) as co-carcinogen (Ranadive et al., 1972).
In a tumour promotion study, 30 female mice were given a non-
carcinogenic dermal dose of 7,12-dimethylbenz[ a]-anthracene
followed by a dermal application of about 20 µl n-hexane (99.8%
pure) in cyclohexane (20 g in 100 ml) three times a week for 60
weeks (Sice, 1966). No skin tumours were reported but there was
dermal irritation. A similar study using 1-hexanol (97% pure), a
metabolite of n-hexane, and a group of 50 mice yielded similar
results. However, Bock & Chortyk (1970) found that n-hexane
induced tumours in mice previously primed with 7,12-dimethyl-
benz[ a]anthracene.
8.6. Neurotoxicity
Although effects on the central nervous system have been noted,
the principal toxic effect of n-hexane is a peripheral neuropathy.
8.6.1. Central nervous system effects
Altered latency and amplitude values for the fifth component of
the brainstem auditory-evoked response (BAER), which reflects
central nervous system activity, were reported from week 3 - 4 of
exposure, and the visual-evoked cortical response was affected
(Rebert & Sorenson, 1983). At 1760 mg/m3 (500 ppm) (the lowest
concentration investigated in this series of studies), reduced
forelimb and hindlimb grip strengths and increased latency of the
fifth BAER component were noted after 4 - 6 weeks of exposure.
There was slight recovery of the amplitude of the fifth BAER
component at the time of last measurement (15 weeks post exposure).
Neuropathological changes were seen in the spinal cord, medulla
oblongata, and cerebellar vermis of Sprague-Dawley rats exposed for
1 - 23 weeks to 1408 - 2112 mg/m3 (400 - 600 ppm) n-hexane (95%
pure) (Spencer & Schaumburg, 1977a,b).
In Fischer-344 rats exposed to 10 560, 22 880, or 35 200 mg/m3
(3000, 6500, or 10 000 ppm) n-hexane (99.5% pure) 6 h/day, 5
days/week, for 13 weeks, axonal swelling was noted in the brain
(medulla) in one out of 10 animals exposed to 35 200 mg/m3
(Cavender et al., 1984). There were no clinical signs of
neurotoxicity.
8.6.2. Peripheral nervous system effects
8.6.2.1 Clinical and morphological findings
The initial clinical signs of n-hexane-induced neuropathy in
rats are effects on the hind feet (flat foot, foot eversion),
followed by a waddling gait, an inability to extend the hindlimbs,
and symmetrical, distal hindlimb weakness. In the studies of
Dunnick (1989) and Dunnick et al. (1989) reported in section
8.2.1.2, B6C3F mice were exposed to 0, 1760, 3520, 14 080, and 35
200 mg/m3 (0, 500, 1000, 4000, and 10 000 ppm) n-hexane for 6
h/day, 5 days/week, for 13 weeks. Mice were also exposed to 3520
mg/m3 for 22 h/day, 5 days/week for 13 weeks. A decrease in
locomotor activity in female mice exposed to 3520 mg/m3 for 22
h/day or to 35 200 mg/m3 for 6 h/day was the only neurobehavioural
effect observed. Paranodal axonal swellings in tibial nerves were
observed in 3 out of 4 males and 3 out of 4 females at these
concentrations, but in none of the 4 controls. The severity of the
peripheral nerve lesion was mild and neither segmental demyelination
nor distal axonal degeneration was observed (Dunnick, 1989; Dunnick
et al., 1989). The relative insensitivity of the mouse to
alpha-diketone neurotoxicity has been related to the relatively
short length and smaller diameter of the myelinated axons in this
species compared to rats or human beings (Graham & Gottfreid,
1984). There can be progression to flaccid paralysis of the
hindlimbs and eventual quadriplegia (Spencer & Schaumburg, 1977b;
Altenkirch et al., 1978, 1982; Howd et al., 1983).
Inhalation exposure of Sprague-Dawley rats for 1 - 23 weeks to
n-hexane (98% pure) at 1408 - 2112 mg/m3 (400 - 600 ppm) produced
axonal swellings, containing numerous neurofilaments, on the
peripheral and central nervous systems (Schaumburg & Spencer, 1976;
Spencer & Schaumburg, 1977a,b; Spencer, 1982). In limb nerves,
these lesions occurred principally in long, large-diameter
myelinated fibres, but were also found in smaller myelinated and
unmyelinated fibres. The swellings in myelinated fibres were
accompanied by paranodal myelin retraction and finally distal axon
disintegration and axon loss. Remyelination and, rarely, axonal
regeneration have been observed in the limb nerves of rats
continuously exposed to 1408 - 2112 mg n-hexane/m3, but the extent
to which regeneration may occur post exposure in vivo has not been
reported (Spencer & Schaumburg, 1977a). In male Sprague-Dawley
rats exposed to n-hexane (99% pure), axonal degeneration in the
tibial nerve was observed following exposure to 8800 mg/m3 (2500
ppm) for 30 weeks or 17 600 mg/m3 (5000 ppm) for 14 weeks (Frontali
et al., 1981).
Neuropathological lesions have been reported in Sprague-Dawley
rats following exposure to n-hexane concentrations ranging between
1408 - 3520 mg/m3 (400 - 1000 ppm) (Schaumburg & Spencer, 1976;
Spencer & Schaumburg, 1977a,b; Altenkirch et al., 1982; Pryor et
al., 1982; Spencer, 1982; API, 1983a,b). Axonal swellings were
observed in hindlimb nerves (tibial nerve branches to the calf and
then plantar nerve branches) and in the central nervous system
(spinal cord, medulla oblongata, and cerebellar vermis) of Sprague-
Dawley rats exposed for 1 - 23 weeks to 1408 - 2112 mg/m3 (400 -
600 ppm) n-hexane (98.98% pure) (Spencer & Schaumberg, 1977a,b).
These occurred before the appearance of hindlimb weakness. In male
CD rats exposed to 440 or 1760 mg/m3 (125 or 500 ppm) n-hexane (99%
pure) 22 h/day, 7 days/week, axonal swelling was seen in the tibial
nerve and brain (medulla oblongata) after 2 months of exposure to
1760 mg/m3 (Spencer, 1982; API, 1983a,b). Pronounced degeneration
and loss of axons in tibial (most severe) and sciatic nerves,
lumbar spinal cord, and medulla oblongata were seen after 6 months.
There was complete recovery from the effects seen at 1760 mg/m3
(500 ppm) by 6 weeks after the end of the exposure period.
Clinical signs of hindlimb paralysis had disappeared by 22 weeks
after exposure to 3520 mg/m3 (1000 ppm), but there was only partial
recovery of hindlimb grip strength and slight recovery of latency
of tail nerve action potential.
In male Fischer-344 rats exposed by inhalation to 3520 mg/m3
(1000 ppm) n-hexane (95% pure) for 24 h/day, 5 days/week, for 11
weeks, spontaneous motor activity was inhibited (Pryor et al.,
1982). Hindlimb grip strength was diminished but there was less
effect on forelimb grip strength. Multisensory conditioned
avoidance response and pole-climbing ability decreased after 8
weeks of exposure but recovered between weeks 18 and 36. The fifth
component of the BAER showed increased latency and decreased
amplitude after 3 weeks. Latency returned to normal 15 weeks after
the end of exposure, although there was only a slight recovery of
amplitude.
Clinical signs of chronic neurotoxicity were observed after 10
weeks in male Wistar rats exposed to 10 560 mg per m3 (3000 ppm)
n-hexane (99% pure) for 12 h/day, 7 days/week, for 16 weeks
(Takeuchi et al., 1980). Following daily inhalation of 3696 mg/m3
(1050 ppm) n-hexane (99% pure) for 16 weeks (12 h/day), tail nerve
conduction velocity in male Wistar rats was reduced, but without
definite clinical signs of neurotoxicity (Takeuchi et al., 1981).
In another study at n-hexane concentrations of 722 or 1725 mg/m3
(205 or 490 ppm) for 24 weeks there were similar findings (Ono et
al., 1982). The effect was still marked 4 weeks after the end of
the 16-week exposure (Takeuchi et al., 1981). Axonal swelling and
demyelination were observed in the tail nerves after 24 weeks of
exposure to 722 mg/m3 (Ono et al., 1982). There were no
significant effects on tail nerve conduction time in rats exposed
to 373 mg/m3 (106 ppm), 12 h/day, 7 days/week, for 24 weeks; the
tail nerve of the one rat examined histologically was normal
(Takeuchi et al., 1983).
Higher no-observed-effect levels have been reported for male
Wistar rats in studies using daily 8-h exposures to n-hexane. No
effects on tail nerve structure or function were observed after 33
weeks of exposure to 1795 mg/m3 (510 ppm) n-hexane (97% pure)
(Iwata et al., 1984). There were no clinical signs of
neurotoxicity or treatment-related lesions in regions of the
nervous system considered to be vulnerable to hexacarbon
intoxication (including areas of the lumbar spinal cord and tibial
nerve) following inhalation of 1760 or 2464 mg/m3 (500 or 700 ppm)
n-hexane (purity not stated) for 9 or 40 weeks (Altenkirch et al.,
1982).
No clinical or histological evidence of n-hexane-induced
peripheral or central neurotoxicity was noted in two separate
studies following exposure of rats to 440 mg/m3 (125 ppm) for about
22 h/day, 7 days/week, for 26 - 34 weeks (API, 1978, 1983a,b;
Spencer, 1981, 1982).
8.6.2.2 Electrophysiological effects
The behavioural and neurophysiological effects of n-hexane
have been studied in rats exposed for 24 h/day, 5 - 7 days/week,
for 7 - 11 weeks (Pryor et al., 1982; Rebert et al., 1982; Howd et
al., 1983; Rebert & Sorenson, 1983). In male Fischer-344 rats,
exposed by inhalation for 24 h/day, 5 days/week, for 10 or 11 weeks
to 1760, 3520, or 5280 mg/m3 (500, 1000, or 1500 ppm) n-hexane (95%
pure), clinical signs of neuropathy, impairment of a conditioned
avoidance response, and increased latency in the action potential
of the caudal nerve were noted at the two highest exposure levels.
Grip strength was reduced in the hindlimbs and, to a lesser extent,
in the forelimbs of all exposed groups (Rebert & Sorenson, 1983).
Weanling male Fischer-344 rats exposed 14 h/day, 7 days/week,
for 14 weeks to 7040 mg/m3 (2000 ppm) n-hexane (95% pure) showed
behavioural and neurophysiological effects, including reduction of
motor activity, startle response, pole-climbing ability, avoidance
response, and grip strength, from week 2 of treatment (Pryor et
al., 1983). Tail nerve latency was prolonged from week 8, the
visual evoked cortical response was increased at 6 weeks, and there
was a reduced amplitude of the fifth BAER component from week 10.
There was no recovery of the tail nerve latency or the fifth BAER
component six weeks after exposure. The visual-evoked cortical
response had fully recovered 1 week after the end of exposure.
In a study of effects on the peripheral and central nervous
systems, no significant behavioural or neuropathological effects
were observed in Wistar rats exposed to 352, 1056, or 3168 mg/m3
(100, 300, or 900 ppm) n-hexane (99% pure) for 8 h/day, 5
days/week, for 72 weeks (De Groot & Kepner, 1984). Adult male rats
were exposed to 17 600 mg/m3 (5000 ppm) for 16 h/day, 6 days/week,
for 4 weeks. At all weekly intervals motor conduction velocity was
reduced in tail nerves and did not approach control values in a
4-week post-treatment period. Raised plasma acetylcholinesterase
levels (25 - 40%) were measured at all weekly intervals during
treatment, but these returned to control values 2 weeks after the
treatment (Bastone et al., 1987).
Exposure to commercial grade hexane (65 - 70% n-hexane) at
concentrations of 348, 950, 1936, 3626, and 6688 mg per/m3 (99,
270, 550, 1030, and 1900 ppm), for 24 h/day, 6 days/week, for one
year, caused electromyographic effects on the hindlimb muscles of
male SM-A mice (Miyagaki, 1967). The effects were indicative of
damage to the peripheral nervous system and included insertion-type
fibrillation and polyphasic discharges at 950 mg/m3 (270 ppm) or
more, spontaneous fibrillation at 3626 and 6688 mg per/m3, and
reduced interference patterns at 1936 mg per m3 or more. The minor
effect at 348 mg/m3 (mild polyphasic discharges) was not considered
to be toxicologically significant, and 348 mg/m3 was taken as the
no-observed-adverse-effect level in this study.
8.6.2.3 Grip strength
In male Sprague-Dawley rats exposed to n-hexane (99% pure) for
6 - 9 h/day, 5 days/week, at concentrations of 1740, 5280, 8800, or
17 600 mg/m3 (500, 1500, 2500, or 5000 ppm) for 14 - 30 weeks,
lesions were observed in the hindlimb nerves following inhalation
of 8800 or 17 600 mg/m3 (Frontali et al., 1981). In a study using
repeated 10-min exposures (6 - 24 exposures/day, 5 days/week),
reduced grip strength was noted in male Fischer-344 rats after
5 - 6 weeks of exposure at n-hexane (95% pure) levels of 84 480 to
168 960 mg/m3 (24 000 to 48 000 ppm) (Pryor et al., 1982).
Amplitude and latency of the fifth BAER component were affected by
exposure to 168 960 mg/m3. The amplitude decreased from week 11,
and there was no recovery after the end of exposure. There were
significant effects on grip strength and nerve conduction velocity
in Wistar rats after exposure for 8 h/day, 5 days/week, for 8 weeks
to 3168 mg hexane/m3 (900 ppm) (De Groot & Kepner, 1984).
8.7. Toxicity of n-Hexane Metabolites
The production of testicular lesions in rats following exposure
to 2,5-hexanedione and other metabolites of n-hexane has been
described by Katz et al. (1980), Krasavage et al. (1980), Cavanagh
& Bennetts (1981), Chapin et al. (1982), Anthony et al. (1983), and
Sayre et al. (1986). Studies in Fischer-344 rats given a 1%
solution of 2,5-hexanedione in drinking-water, in which severe
seminiferous epithelial degeneration and azoospermia were reported,
indicated that the Sertoli cell was probably the initial target for
2,5-hexanedione action (Chapin et al., 1982, 1983). Testicular
atrophy, induced by 2,5-hexanedione at levels of 0.5% and 1% in the
drinking-water or 200 mg/kg per day intraperitoneally, was observed
in Sprague-Dawley rats before clinical signs of neuropathy or
histological axonal swelling (O'Donoghue et al., 1978b).
In Charles River CD rats treated with 1% 2,5-hexanedione in the
drinking-water for 4 weeks, testicular toxicity, as shown by
decreased testis weight compared to controls, and enlarged and more
numerous membrane-bound vacuoles basally located in Sertoli cells,
was seen, together with occasional giant cells and chromatin
margination in spermatids. Brain and testis tubulin from treated
animals was found to be cross-linked and had a shortened nucleation
phase of assembly and a more rapid rate of elongation. Tubulin
cross-linking was postulated to play an important role in the
testicular toxicity of the gamma-diketone (Boekelheide, 1987).
When Boekelheide (1988a) treated Charles River CD rats
intraperitoneally with 4 mmol 2,5-hexanedione/kg per day, 5
days/week, for 2 weeks, no evidence of neurotoxicity was seen.
Testis weight and histology were normal 2 weeks after the start of
dosing, but large basally located Sertoli cell vacuoles were seen
in the testis at 4 weeks. At 7 weeks most of the seminiferous
tubules were normal and vacuolation of Sertoli cells was no longer
prominent. In the same study, rats were treated with 1%
2,5-hexanedione in the drinking-water for 2 or 3 weeks. Only after
3 weeks was neurotoxicity evident from an unsteady gait. Decreased
testicular weights were observed 7 weeks from the start of dosing
in both groups, and loss of germ cells in seminiferous tubules was
seen in both groups. At 27 weeks the testis weights were variable
with partial recovery to normal values, and there was variable
repopulation of seminiferous tubules. Rats were also dosed with 1%
2,5-hexanedione in the drinking-water for 5 weeks. This produced
moderate to severe clinical neurotoxicity, and basally located
vacuoles first appeared in Sertoli cells at 4 weeks. There was
progressive necrosis and generalized sloughing of germ cells. A
17-week recovery period resulted in an absence of all
postspermatogonial germ cells from the seminiferous epithelium in 3
out of 5 treated rats (Boekelheide 1988a). Under identical
conditions of treatment and recovery, pyrrole reactivity of testis
proteins increased to maximal levels within 2 weeks of treatment
and nucleation time for microtubule assembly from purified testis
tubulin was decreased at 2 weeks. This was associated with the
presence of cross-linked tubulin, which was suggested to lead to
germ cell loss (Boekelheide, 1988b).
In a study by Boekelheide & Eveleth (1988), Charles River CD
rats were treated with a total dose of 131 (± 2) mmol
2,5-hexanedione/kg at dose rates ranging from 1.9 to 6.1 mmol/kg
per day, and testicular damage was assessed 4 weeks after the
cessation of treatment. Reduced testis weight and germ cell
depletion were dependent on the rate of intoxication rather than
the total dose. The rate of intoxication also determined the
extent of testis pyrrole formation.
The metabolite 2,5-hexanediol was found to have an effect on
the spleens of female albino rats (Goel et al., 1987). Following
oral dosing with 2,5-hexanediol (960 mg/kg) daily for 7 days, the
haematocrit value, haemoglobin content, and level of neutrophils
were raised, while the level of lymphocytes was reduced relative to
controls. The same treatment led to a significantly greater uptake
of iron into the spleen and a significant inhibition of spleen
delta-aminolevulinic acid dehydratase activity.
Atrophy of the spleen, thymus, and lymph nodes has been
reported in albino rats given oral doses of 0.25, 0.5, 0.75, or 1
ml 2,5-hexanedione/kg body weight (Singh et al., 1983, 1984; Kannan
et al., 1985). Atrophy of the thymus, due to loss of cortical
cells, was severe but reversible. Immunocompetence (including
resistance to endotoxin shock) was impaired in mice. Lymphoid
lesions have been reported in rats dosed orally with
2,5-hexanedione (0.428 ml/kg) (Upreti et al., 1986).
2,5-Hexanedione is thought to be the ultimate neurotoxic
metabolite of n-hexane (Graham et al., 1982a). An intermediate
metabolite, 2,5-hexanediol, has been found to produce widespread
axonal swellings throughout the nervous system including severe
swelling in the distal optic tract (Cavanagh & Bennetts, 1981).
Axonal swellings did not progress and were reversible (Cavanagh,
1982; Jones & Cavanagh, 1982). However, there was progression to
axonal degeneration during and/or after exposure in long, large-
diameter fibres of hindlimb nerves and many spinal tracts. No
significant regeneration of peripheral nerves was seen. In a study
on cats exposed to 2-hexanone, one animal, retained for 4´ months
after exposure ceased, still showed a moderate loss of myelinated
nerve fibres and axonal degeneration in the sciatic nerve (Saida et
al., 1976). Using a sensitive automated technique for detecting
altered coordination of hindlimb movement in male Wistar rats,
Kulig et al. (1985) detected impairment of coordination 1 week
after oral treatment with 600 mg 2,5-hexanedione/kg and 2 weeks
after treatment with 250 mg/kg. The performance of the rats
treated with 600 mg/kg was still impaired relative to controls 5
weeks after treatment. In the optic tract of cats, 2,5-hexanedione
(given as a 0.5% solution in drinking-water up to a cumulative
maximum of 5.8 ml/kg; maximum of 62 days of dosing) caused axonal
swellings that were not selective for fibre size, impaired axonal
transport, and neuronal degeneration in large cells. There was
also a loss of flicker resolution (Pasternak et al., 1985).
Reduced glucose utilization and altered blood flow and glucose
uptake in various areas of the brain have been reported in rats
exposed to 2,5-hexanedione (Griffiths et al., 1981; Cooper et al.,
1985). The inferior colliculus was one of the regions most
affected in terms of reduced glucose utilization. This region is
believed to be the site where the fifth component of the brainstem
auditory evoked response (BAER) is generated and this component has
been shown to be affected in rats by n-hexane exposure (Rebert et
al., 1982; Pryor et al., 1982, 1983; Howd et al., 1983; Rebert &
Sorenson, 1983). Oral treatment of male Sprague-Dawley rats with
2,5-hexanedione (1% in drinking-water) led to increased plasma
concentrations of acetylcholinesterase at 2 and 4 weeks (p < 0.05
and < 0.01, respectively). This agrees with the observed increase
of plasma acetylcholinesterase following n-hexane inhalation
(section 8.6).
Graham et al. (1982a) suggested that the accumulation of
neurofilaments is not secondary to a defect in axonal transport
that may be brought about by impairment of energy production by
2-hexanone or 2,5-hexanedione (Sabri et al., 1979; Sabri, 1984),
but rather that the aggregation may be a primary event leading to
altered axonal transport. 2,5-Hexanedione-induced neurofilament
cross-linking was proposed as a mechanism leading to occlusion of
flow at the nodes of Ranvier, with subsequent enlargement of the
axon proximal and degenerative changes distal to this point (Graham
et al., 1982b). Polymer formation by cross-linking of nerve
protein (see section 6.1.3) was demonstrated using an analogue of
2,5-hexanedione (3,5-dimethyl-2,5-hexanedione) which underwent
pyrrole formation more rapidly (Graham et al., 1985).
DeCaprio & O'Neill (1985) treated male Wistar rats with 0.5%
2,5-hexanedione in the drinking-water for 8 weeks and found
clinical signs of neuropathy after 5 weeks of exposure.
2,5-Dimethylpyrrole adduct was detected in both serum and axonal
cytoskeletal proteins in the exposed animals. After 2 weeks of
exposure, < 1% of epsilon-amine groups were converted into pyrrole
adducts in axonal protein. The formation of proteins of high
relative molecular mass in the brain stem and spinal cord was
indicative of protein cross-linking. This was associated with a
loss of native neurofilament subunit proteins in the brain stem.
In vitro incubation of axonal protein with 2,5-hexanedione also
resulted in the formation of proteins of high relative molecular
mass. In support of the theory that pyrrole formation is a
requirement for gamma-diketone neurotoxicity, DeCaprio et al.
(1988) demonstrated that perdeuterio-2,5-hexanedione pyrrolylated
protein at only one-third of the initial rate observed with 2,5-
hexanedione, and that the deuterium analogue also caused less body
weight loss than 2,5-hexanedione treatment of male Wistar rates (5
days/week, 3.5 mmol/kg per day for 17 days or 2.5 mmol/kg per day
for 38 days). Hindlimb paralysis and neuropathological changes
were seen only in the rats treated with 2,5-hexanedione. Pyrrole
adduct concentrations were 2- to 3-fold higher in the serum and
axonal cytoskeletal proteins of 2,5-hexanedione-treated rats
compared with those treated with perdeuterio-2,5-hexanedione. The
primary isotope effect was considered to be due to the requirement
for C-H bond cleavage in pyrrole formation.
The 2,5-hexanedione analogue, 3-acetyl-2,5-hexanedione,
pyrrolated protein in vitro at an equal rate to the potent analogue
3,4-dimethyl-2,3-hexanedione but did not form protein cross-links.
Sprague-Dawley rats receiving 0.1 or 0.25 mmol 3-acetyl-2,5-
hexanedione/kg per day by intraperitoneal injection and sacrificed
at 20 and 7 days, respectively, did not show hindlimb paralysis or
neurofilament-filled axonal swellings characteristic of
gamma-diketone neurotoxicity. Thus protein cross-linking appears
to be a critical step in the neurotoxicity of pyrrolating agents
such as 2,5-hexanedione (Genter St Clair et al., 1988). In a study
of the relative effects of a range of analogues of 2,5-hexanedione,
Genter et al., (1987) found a strong correlation between the rate
of pyrrole formation, the rate of protein cross-linking in vitro
and the production of neurofilament-filled axonal swellings.
Rosenberg et al. (1987b) studied male Sprague-Dawley rats given
either the dl- or meso-diastereoisomers of 3,4-dimethyl-2,5-
hexanedione (0.2 mmol/kg per day by intraperitoneal injection).
The dl-stereoisomer produced hindlimb paralysis in one-third of the
time of the meso-stereoisomer, paralleling the relative rates of
in vitro amine pyrrolation. The more distal location of
neurofilament-filled swellings after meso-3,4-dimethyl-2,5-
hexanedione treatment was suggested to be due to the relatively
slow rate of neurofilament cross-linking. Hyperbaric oxygen
treatment made the onset of hindlimb paralysis more rapid in male
Sprague-Dawley rats after daily treatment with 2,5-hexanedione (4
mmol/kg by intraperitoneal injection, 7 days/week). Rats treated
with 2,5-hexanedione plus hyperbaric oxygen reached the end-point
of hindlimb paralysis at 19.8 (± 1.2) days compared to 23.5 (± 0.7)
days in rats treated with 2,5-hexanedione alone (p < 0.02). The
results support the role of pyrrole auto-oxidation and resultant
protein cross-linking in the neurotoxic process (Rosenberg et al.,
1987a).
A summary of a proposed molecular mechanism linking
neurofilament pyrrolylation with nerve degeneration in 2,5-
hexanedione neuropathy is shown in Fig. 2.
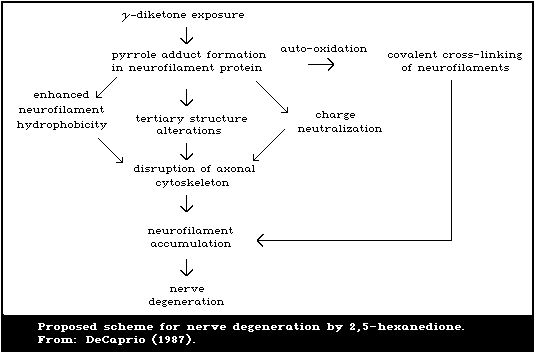 Monoclonal antibodies to neurofilament proteins have been used
to demonstrate protein cross-linking in the spinal cords of rats
treated with 2,5-hexanedione (Lapadula et al., 1986). In male
Sprague-Dawley rats given 2,5-hexanedione in drinking-water at
concentrations of up to 1% (v/v) for 70 days, there was dose-
dependent protein cross-linking, with decreased protein
phosphorylation of neurofilament proteins and microtubulin-
associated protein-2 (MAP-2) (Abou-Donia et al., 1988). The use of
monoclonal antibodies to neurofilament proteins has permitted the
demonstration of protein cross-linking in the spinal cords of rats
treated with 2,5-hexanedione (Lapadula et al., 1986). It was also
found that protein phosphorylation was decreased in association
with protein cross-linking in 2,5-hexanedione-treated rats (Abou-
Donia et al., 1988). It is not known if the decrease in protein
phosphorylation is a primary event or a consequence of cross-
linking. Exposure of three cell lines derived from human tumours
or fetal mouse brain to 2,5-hexanedione in vitro (0.04 - 0.16%)
resulted in reduced proliferation and an increase in cell
protrusions or dendritic-like processes (Malorni et al., 1989),
indicating that in vitro toxicity is not restricted to the neuronal
system.
2,5-Hexanedione causes aggregation of intermediate filaments of
the vimentin type in cultured human skin fibroblasts. Exposure to
2.8 mmol 2,5-hexanedione/litre for 1 or 2 weeks induced aggregation
in 50% and 80% of the cells, respectively; a concentration of 2
mmol/litre required 2 and 3 weeks to produce corresponding effects
(Durham et al., 1983). The non-neurotoxic analogues,
1,6-hexanedione and 2,4-hexanedione, had no effect on filament
distribution in fibroblasts, although in the latter case
cytotoxicity prevented the study of effects at concentrations
equivalent to those required for 2,5-hexanedione-induced
aggregation (Durham et al., 1988).
The neurotoxicity of 2,5-hexanedione was increased by
co-exposure to methyl ethyl ketone (Ralston et al., 1985). Exposure
to other apparently non-neurotoxic aliphatic ketones (2-pentanone,
2-heptanone, and 2-octanone) increased the neurotoxicity of the
n-hexane metabolite 2-hexanone; 2-octanone had a greater effect
than methyl ethyl ketone (Misumi & Nagano, 1985). Ladefoged et al.
(1989) dosed rats with 2,5-hexanedione (0.5% in the drinking-water)
for six weeks and compared the effects obtained with or without
administration of 0.5% acetone or 5.0% ethanol. Nerve conduction
velocity and behavioural rotarod performance were reduced after 3
weeks of dosing with 2,5-hexanedione. Acetone, but not ethanol,
together with 2,5-hexanedione, had an additional significant effect
on these parameters from the fourth week of dosing.
9. EFFECTS ON MAN
9.1. Single Exposures
In studies on human volunteers, inhalation of 17 600 mg
hexane/m3 (5000 ppm) for 10 min resulted in vertigo and giddiness
but there were no symptoms with exposure to 7040 mg/m3 (2000 ppm)
for the same duration (Patty & Yant, 1929). Occupational exposures
to hexane concentrations of 3520 to 89 760 mg/m3 (1000 to 25 500
ppm) for periods of 30 - 60 min caused drowsiness (Yamada, 1967).
9.2. Skin and Eye Irritation; Sensitization
9.2.1. Skin irritation
Prolonged occlusive skin contact for 1 - 5 h with liquid hexane
caused erythema and, after 5 h, blistering (Oettel, 1936).
Application of 1.5 ml n-hexane (analytical grade) to 3.1 cm2 of
forearm skin for 5 min resulted in a stinging and burning sensation
and transient erythema (Wahlberg, 1984a). 0.1 ml n-hexane rubbed
gently into the forearm skin for 18 days did not produce erythema
or oedema (Wahlberg, 1984b).
Operators at a soybean hexane-extraction facility had a higher
incidence of dry or irritated skin than maintenance workers (65%
versus 20%) (NIOSH, 1983).
9.2.2. Eye irritation
Eye irritation did not occur in volunteers exposed to hexane
vapour at a level of 1760 mg/m3 (500 ppm) for 3 - 5 min (Nelson et
al., 1943).
9.2.3. Skin sensitization
A maximization test on 25 subjects with n-hexane applied
undiluted for induction and as a 25% solution for challenge did not
induce skin sensitization (Kligman, 1966).
9.3. Short- and Long-Term Exposures
9.3.1. Effects on the nervous system
9.3.1.1 Peripheral neuropathy
Peripheral neuropathy has been reported in humans exposed
industrially to n-hexane or through solvent abuse of glues or
solvents containing n-hexane. The majority of occupational cases
have occurred in small industrial concerns such as shoe manufacture
in Italy and vinyl sandal production in Japan (Sobue & Yamamura,
1968; Yamamura, 1969; Inoue et al., 1970; Abbritti et al., 1976;
Carapella, 1977; Palao & Lajo, 1981; Nakajima & Murayama, 1985).
Workers were exposed to many volatile compounds, not only
n-hexane, in badly ventilated rooms, sometimes for periods in
excess of 8 h/day under poor hygienic conditions (Yamamura, 1969;
Inoue et al., 1970, 1971; Battistini et al., 1974; Abbritti et al.,
1976; Carlomagno et al., 1983).
The first signs of n-hexane neuropathy are symmetrical
paraesthesia and weakness. The lower extremities are normally
affected first. A number of other symptoms such as headache,
anorexia, and dizziness may precede or coincide with the
neuropathy. A "glove and stocking" sensory impairment to touch,
pain, vibration, and temperature develops, with weakness and
atrophy affecting proximal muscles of the extremities. On clinical
examination, most patients show reduced body weight and diminished
or absent reflexes. There is a marked reduction in conduction
velocity in sensory and motor nerves. Electromyography (EMG) shows
a neuropathic pattern including fibrillation potentials, positive
sharp waves, polyphasic potentials, and reduced interference
patterns. Axonal lesions of the large myelinated fibres (identical
to those produced in experimental animals) with axonal swelling
followed by myelin retraction at the node of Ranvier are found on
nerve biopsy. There is only a slight to moderate reduction in the
number of fibres, mainly of large myelinated fibres (Goto et al.,
1974; Shirbae et al., 1974; Okamoto et al., 1977; Rizzuto et al.,
1977, 1980; Scelsi et al., 1980; Chauplannaz et al., 1982; Lagueny
et al., 1982). There are masses of 9- to 10-nm long neurofilaments
in the axons. Lesions are rarely found in the small myelinated and
unmyelinated fibres (Shirbae et al., 1974; Altenkirch et al., 1977;
Rizzuto et al., 1977, 1980; Scelsi et al., 1980; Ruff et al., 1981;
Chauplannaz et al., 1982). Muscle biopsies show signs of
myofibrillar atrophy (Yamamura, 1969; Gonzalez & Downey, 1972;
Shirbae et al., 1974; Altenkirch et al., 1977). Recovery is always
gradual and slow and signs of residual neuropathy may persist in
severe cases for 3 - 4 years (Yamamura, 1969; Iida & Yamamoto,
1973; Altenkirch et al., 1977; Sobue et al., 1978; Rizzuto et al.,
1980; Carlomagno et al., 1983). Signs of pyramidal tract defects
such as residual hyper-reflexia and spasiticity and autonomic
defects have been reported in a few severe cases (Korobkin et al.,
1975; Altenkirch et al., 1977; Altenkirch & Schultz, 1979;
Schaumburg & Spencer, 1979). Electrophysiological evidence of
central nervous system dysfunction (abnormal evoked potentials) has
occasionally been reported in individual cases of peripheral
neuropathy (Yoshida et al., 1974; Lagueny et al., 1982).
Neuropathy has occurred following exposure to a wide range of
n-hexane levels in air from 97 to 8125 mg/m3 (30 - 2500 ppm) (Oishi
& Mineno, 1964; Yamada, 1964, 1967; Wada & Okamoto, 1965;
Herskowitz et al., 1971; Nomiyama et al., 1973; Yoshida et al.,
1974; Takeuchi et al., 1975; Paulson & Waylonis, 1976; Takahashi et
al., 1977; Brugnone & Perbellini, 1980; Ono & Takeuchi, 1980; Ruff
et al., 1981; Carlomagno et al., 1983). Exposure durations ranging
from 2 months to 5 years may precede the onset of overt neuropathy,
and a mild recurrent neuropathy with n-hexane exposure may go
undetected (Wada & Okamoto, 1965; Herskowitz et al., 1971; Nomiyama
et al., 1973; Takeuchi et al., 1975; Takahashi et al., 1977; Ruff
et al., 1981).
In the Italian shoe industry, n-hexane (a component of the
glues used) has been implicated as the cause of peripheral
neuropathies (Di-Bosco & Fonzi, 1974; Abbritti et al., 1976;
Cianchetti et al., 1976; Carapella, 1977; Buitatti et al., 1978;
Aiello et al., 1980; Sanagi et al., 1980; Palao & Lajo, 1981; Mutti
et al., 1982a,b,c).
Towfighi et al. (1976) reported the development of neuropathy
in two patients who sniffed glue containing n-hexane. Complete
clinical recovery occurred with one patient who changed to a glue
not containing n-hexane. Sural nerve biopsies of the affected
individuals indicated loss of axons. Some remaining axons had
accumulation of filaments, widened nodes of Ranvier, and focal
enlargements. Muscle biopsy revealed neurogenic atrophy. Three
individuals in a laboratory developed neuropathy that was
attributed to n-hexane in glue (80.4% n-hexane). In addition to
axonal changes, the main findings were breakdown of nerve fibres
and swelling and degeneration of myelin. In addition, a
denervation muscle atrophy, combined with focal myopathic changes,
occurred (Scelsi et al., 1981).
a) Japanese sandal workers
Health surveillance studies were carried out in Japan following
a mass outbreak of polyneuropathy in 1967 (Sobue & Yamamura, 1968;
Iida et al., 1969, Yamamura, 1969; Inoue et al., 1970, 1971; Iida &
Yamamoto, 1973). Through the use of questionnaires and medical
examinations, 93 cases of polyneuropathy were identified out of
1662 vinyl sandal workers who were using glues based on solvents
containing more than 70% commercial grade hexane. Their work was
carried out for periods of up to 14 h/day in small, badly
ventilated rooms with estimated hexane air concentrations of
between 1625 and 8125 mg/m3 (500 and 2500 ppm). Air concentrations
of the other volatile solvents in the glue were not described.
The workrooms were often used also as dwelling places so that
continuous exposure for up to 24 h/day was possible. Clinically,
the cases were classified into three groups, according to the
severity of the disease:
Group I: sensory neuropathy (53 cases);
Group II: sensorimotor neuropathy (32 cases); and
Group III: motor neuropathy with amyotrophy (8 cases).
Single exposure estimations at the homes/workrooms of some of
the patients indicated a positive relationship between the air
concentration of hexane, the duration of the working day and the
number of sandals made, and the severity of the polyneuropathy.
n-Hexane concentrations of 1625 - 3250 mg/m3 (500 - 1000 ppm)
during an 8-h day were sufficient to cause severe peripheral
neuropathy (Inoue et al., 1971).
In 89% of the cases, the onset was insidious. Initial symptoms
were sensory impairment (88%), muscular weakness (14%), blurred
vision (14%), coldness of extremities (13%), headache (9%), loss of
body weight (9%), lassitude (3%), skin eruptions (2%), anorexia
(2%), and dizziness (1%). On examination, the most common finding
was a symmetrical sensorimotor disorder of the extremities.
Hypoaesthesia was found in all cases and dysaesthesia was found in
the early stages of the disease in 23% of patients. Other finding
were muscular weakness of the limbs (43%), diminished tendon
reflexes (39%), and exaggerated tendon reflexes (11%). Numbness of
the face was described in five cases. Of 39 cases examined
ophthalmologically, eight had abnormal findings, seven had
restriction of the visual field, two had optic nerve atrophy, and
one had optic neuritis. There was no correlation between the
severity of neuropathy and the severity of optic effects.
Electromyography (EMG) was performed on 44 patients (11 in
Group I, 25 in Group II, and 8 in Group III) (Iida et al., 1969;
Yamamura, 1969). Signs of denervation, including fibrillation and
positive sharp waves, were found in 70% of cases in Groups I and II
and in all cases in Group III. A reduction in the number of motor
units under voluntary control (reduced interference pattern) was
found in 50% of cases in Group I and 70% of cases in Group III.
High amplitude and polyphasic action potentials were more common in
Group III than in Groups I and II. The severity of the neuropathy
correlated with the type and magnitude of electromyographic
abnormalities.
Motor nerve conduction velocities and evoked potentials (mixed
nerve conduction velocity) were measured in the ulnar, median,
peroneal, and tibial nerves in the same 44 cases. Sensory nerve
conduction velocities were measured in the ulnar and median nerves.
The number of cases with reduced motor nerve and sensory nerve
conduction velocities correlated with the severity of the
neuropathy. Reduced mixed nerve conduction velocity correlated
only with the severity of neuropathy in measurements obtained from
peroneal and tibial nerves.
Muscle and nerve biopsies were taken from a small number of
cases. Sections of muscle showed neurogenic atrophy. Sections of
nerve showed axonal and myelin lesions, and demyelination was
considered to be the primary lesion (data obtained from animal
studies indicate that the primary lesion in n-hexane neuropathy is
axonal).
Polyneuropathy progressed in all severe cases (Group III) for
up to 4 months after cessation of exposure. A gradual, slow
recovery then ensued. Of 36 cases that were followed-up, 50%,
including six severe cases (Group III), had completely or nearly
completely recovered within 3 - 18 months. Two cases in Group III
displayed residual atrophy and weakness. There were no deaths
(Sobue & Yamamura, 1968; Yamamura, 1969; Inoue et al., 1970).
The clinical recovery of 90 cases was followed for
approximately 5 years. Approximately 3 years after the outbreak,
51 cases had recovered completely. The remaining 39 cases were
reclassified into Groups I and II (34 and 5 cases, respectively).
After 5 years, 82 cases had recovered completely, while the
remainder had residual sensory symptoms (Iida & Yamamoto, 1973;
Sobue et al., 1978). A further study reported a significant
improvement during the 5-year follow-up period in the EMG and nerve
conduction velocities of some patients initially placed in Groups I
and II (Iida, 1982). Patients in Group III also showed
improvements in the EMG. Denervation activity and polyphasic
potentials disappeared and interference patterns and high-voltage
action potentials reappeared. In some patients, nerve conduction
velocities were depressed for over a year after the onset of the
neuropathy.
In 1981, 21 new cases of mild neuropathy were identified in the
same district (Iida, 1982). Air measurements at the workplaces of
the sandal makers indicated that n-hexane air levels were below 176
mg/m3 (50 ppm). These cases were divided by clinical examination
into 20 cases in Group I and one case in Group II. Signs of
neuropathy included numbness (twelve cases), dysaesthesia (six
cases), slight muscle weakness (six cases), diminished tendon
reflexes (ten cases), and exaggerated tendon reflexes (three
cases). Skin changes (coldness, redness, and roughness) were
observed in four cases. A slight floating sensation on change of
posture was reported by five patients.
EMG did not reveal any denervation in resting muscles. EMG
abnormalities, more severe in the lower extremities, including
reduced interference pattern, large amplitude, and polyphasic
potentials, were recorded during voluntary muscle contraction in
approximately 75% of cases. Significant reductions in motor or
sensory nerve conduction velocities were reported in nearly half
the patients.
b) Italian shoe industry
A study of exposure to organic solvents was carried out in an
Italian shoe factory (Mutti et al., 1982a). The exposed group
consisted of 95 workers (24 males, 71 females) with an age range of
16 - 62 years (mean, 30.9 ± 11.7 years), and the exposure duration
ranged from 1 to 25 years (mean, 9.1 ± 8 years). The approximate
mean air concentrations in the breathing zone, over a 2-year
period, for a number of solvents were: n-hexane, 317 mg/m3 (90
ppm); cyclohexane, 315 mg/m3 (92 ppm); methyl ethyl ketone, 115
mg/m3 (39 ppm); and ethyl acetate, 205 mg/m3 (57 ppm). A
concurrent reference group from the same factory of 52 non-exposed
workers (12 males, 40 females) with an age range of 16 - 58 years
(mean, 29.6 ± 12.3 years) and an employment duration ranging from 4
months to 29 years (mean, 10.2 months ± 9.7 years) was selected.
Both exposed and reference workers underwent detailed physical
examination and electrophysiological studies of the median, ulnar,
and peroneal nerves. An exposure score (work time in years
multiplied by an estimate of total solvent exposure) was calculated
for each of the exposed workers.
Acute and chronic neurological signs and symptoms that were
more frequently reported by, or found in, the exposed workers
included sleepiness, dizziness, weakness, paraesthesia, and
hypoaesthesia. Other neurological symptoms, such as headache,
muscular cramps, neurasthenic syndrome, and sleep disturbances,
were found more often in exposed workers, but the differences in
incidence between the exposed and reference group were not
statistically significant. Some parameters of electrophysiological
measurements indicated subclinical neuropathy among exposed
workers. The mean motor nerve conduction velocity was
significantly reduced in the median and peroneal nerves but not in
the ulnar nerve. The amplitude of the motor action potential (MAP)
was significantly reduced in all nerves and its duration was
increased in the ulnar nerve. There were no significant effects on
the distal latency. A significantly higher number of abnormal
action potentials was observed in the median and peroneal nerves of
exposed workers. There was a correlation between the reduction in
motor conduction velocity and the exposure score, but not with age
of the individual. Comparison of the electrophysiological
parameters between two exposed sub-groups and the reference group
indicated an exposure-related trend for reduced motor conduction
velocity in the median and peroneal nerves, and an increased motor
action potential amplitude duration.
When the 95 exposed workers were divided into two sub-groups
based on n-hexane exposure, 65 workers having low exposure (243
mg/m3, 69 ppm) and 30 having high exposure (475 mg/m3, 135 ppm),
there was an exposure-related trend in the reduction of median
nerve motor conduction velocity and increase in the duration of the
ulnar nerve motor action potential (Mutti et al., 1982b).
In a follow-up study, electrophysiological measurements
including somatosensory evoked potentials (SEPs) were recorded from
a group of 15 women shoe factory workers aged 19 - 53 years (mean
age, 26.6 ± 11.4 years) with an exposure duration to solvents of
2 - 8 years (mean, 4.5 ± 2.3 years) (Mutti et al., 1982c). The
mean air concentrations for various solvents in the breathing zone
of the workers were: n-hexane, 690 mg/m3 (196 ppm); cyclohexane,
585 mg/m3 (170 ppm); methyl ethyl ketone, 177 mg/m3 (60 ppm); and
ethyl acetate, 360 mg/m3 (100 ppm). However, total solvent levels
occasionally exceeded 5000 - 7000 mg/m3 when the ventilation system
failed. The study started 3 months after hygiene improvements
successfully controlled air levels of solvents. A control group of
15 age- and health-matched women was used.
Electrophysiological measurements in peripheral nerves showed
significant reductions in maximal motor and distal sensory nerve
conduction velocities in the median and ulnar nerves and reduced
maximal motor nerve conduction velocity in the peroneal nerve. The
latency of the sensory peak action potential was significantly
increased in the median and ulnar nerves. The amplitude of all
peripheral nerve action potentials was slightly reduced but this
was not statistically significant.
There were changes in the somatosensory evoked potentials
(SEPs) of exposed workers with significant increases in the latency
of some early component peaks. The amplitude of some of the early
peaks was also increased, but the amplitude of the later peaks was
significantly reduced. Comparison of SEPs from exposed and non-
exposed workers showed that later components in exposed workers
were flatter.
An electrophysiological study was carried out on 25 workers (13
women, 12 men), placed into two groups according to job
specification, out of 31 workers at an Italian shoe factory (Aiello
et al., 1980). Group A contained 11 subjects (seven women and four
men), aged 21 - 50 years with 0.3 to 20 years of exposure, who
worked in direct contact with solvents. Group B contained 14
subjects (six women and eight men), aged 20 - 49 years and exposed
for 0.25 - 20 years, who had no direct contact with solvents but
were exposed by inhalation. Peak values for air concentrations of
various solvents in the factory were: n-hexane, 70 mg/m3 (20 ppm);
ethyl acetate, 1.8 mg/m3 (0.5 ppm); cyclohexane, 620 mg/m3 (40
ppm); and trichloroethylene, 2.7 mg/m3 (0.5 ppm).
Motor nerve conduction velocity was measured in the median and
peroneal nerves, and sensory nerve conduction velocity in the
median and ulnar nerves. The distal latency and amplitude of
action potentials were measured in motor and in sensory fibres.
Statistically significant electrophysiological changes were
found in seven workers from Group A. An increased distal latency
in sensory or motor fibres was found in six of these workers and
reduced motor nerve conduction velocity in the peroneal nerve in
one worker. In three workers from Group B, there was a significant
increase in the distal latency in sensory nerves of the upper limb.
In one of these, there was reduced motor nerve conduction velocity.
EMG abnormalities were detected in two workers, one from each
group. Seven out of 14 workers exposed for more than 5 years
exhibited one or more electrophysiological abnormalities. In three
out of eight workers exposed for between 1 and 5 years, there were
abnormal findings. There were no abnormalities in the three
workers with less than one year's exposure.
c) Taiwan press proofing workers
In a cross-sectional survey among Taiwanese press proofers,
using solvents containing n-hexane to clean machinery, workers from
16 factories who had been employed in the industry for more than 2
months underwent detailed neurological examination (Wang et al.,
1986). A total of 59 workers (57 males, and 2 females) with a mean
age of 25.8 (± 10.2) years and a mean employment duration of 5.8
years (range, 2 months to 25 years) was examined. Nerve conduction
velocity studies were undertaken in 54 workers, and a diagnosis of
peripheral neuropathy was made on the basis of objective clinical
signs compatible with peripheral neuropathy and at least two
abnormally slow conduction velocities (45 m/sec in the upper
extremities and 40 m/sec in the lower extremities). Mean nerve
conduction velocities for the exposed group were compared with a
control group of 150 individuals (50 in each of the age ranges
10 -35, 36 - 50, and 51 - 80 years). The sex ratio of the control
group was not provided and exposed and control groups were not
matched for age.
n-Hexane was present in samples of the commercial solvents used
to clean the machines from all the factories at concentrations of
between 3 and 65%. Post-event measurements of n-hexane air
concentrations (14 factories) varied from 0 to 669 mg/m3 (190
ppm). Prior to the discovery of neuropathy, standards of hygiene
were described as poor. Groups of 3 - 9 workers cleaned press
proof machines in small poorly ventilated rooms for more than 8
h/day.
Fifteen cases of overt peripheral neuropathy were diagnosed
among exposed workers (14 males, and 1 female; employment range, 6
months to 5 years; median, 1.5 years). Sural nerve biopsies from
three cases were consistent with n-hexane peripheral neuropathy.
The reported n-hexane air concentrations for two factories where
eight of these people worked were 669 mg/m3 (190 ppm) (six cases)
and 74 mg/m3 (21 ppm) (two cases). No exposure data were available
for the remaining seven cases employed in one factory. Twelve of
the 15 cases regularly slept in the factory and were exposed to
n-hexane for daily periods in excess of 8 h.
Exposure to n-hexane air concentrations of less than 352 mg/m3
(100 ppm) was not associated with clinical peripheral neuropathy.
There were significant reductions in the median, ulnar, and
peroneal nerve mean motor nerve conduction velocities in the
n-hexane-exposed workers without diagnosed peripheral neuropathy,
compared with the control group, but an exposure-related trend was
not found.
In a subsequent neurological investigation of workers at these
factories, there was evidence of central nervous system
malfunction, indicated by somatosensory and visual evoked
responses, in individuals with peripheral neuropathy and also in
other workers (Chang, 1985). Twenty-two of the polyneuropathy
cases, five subclinical cases and seven unaffected workers were
investigated. The absolute and interpeak latencies of patterned
visual evoked potential in affected cases were longer than in
normal controls, and the evoked potential interpeak amplitude was
decreased in individuals with polyneuropathy (Chang, 1987).
Effects on the brainstem auditory evoked response (BAER) were also
reported (Chang, 1985). Wave I latency between factory workers and
normal controls did not differ, but prolongation of the wave I-V
interpeak latencies was noted in workers and corresponded to the
severity of polyneuropathy. Five factory workers exposed to
n-hexane for 5 - 30 months had a reduction in nerve conduction
velocity that was correlated with clinical signs (Huang & Chu,
1989). Neurological symptoms consisted of muscle cramp and
weakness, distal numbness, and, in one case, blurred vision and
tinnitus. The tibial-central conduction in the somatosensory evoked
potentials was delayed and the central conduction time of the BAER
was also prolonged, indicating primary defects in the spinal cord
and brainstem.
d) Tungsten carbide milling
The effect of exposure to n-hexane was investigated in a
factory producing tungsten carbide alloys where powders of tungsten
carbide and other metals and carbides were mixed with n-hexane or
acetone by ball mills (Sanagi et al., 1980). Fourteen workers
(concurrently exposed), aged less than 50 with exposure durations
of 1 - 12 years (mean, 6.2 years), who were engaged in the ball
milling and drying of tungsten carbide alloys were studied. A
further group (previously exposed) of five workers, aged less than
50 with exposure durations of 1 - 16 years (mean, 5.2 years), who
had worked in ball milling in the past were also investigated.
Workers exhibiting any evidence of neurological disorders were
excluded from this study. A concurrent control group of 14 workers
not exposed to n-hexane was selected from the sintering workshop.
The mean 8-h time-weighted average concentration of n-hexane over a
2-year period was 204 mg/m3 (58 ppm). The average concentrations
for acetone were in the range 52 - 164 mg per m3 (22 - 69 ppm).
The medical history of each worker was assessed by interview and
questionnaire. Neurological status was determined by detailed
clinical examination and electrophysiological studies.
Headaches, hyperaesthesia in the limbs, and muscle weakness
tended to be reported more often by workers in the exposed group.
Neurological examination did not reveal any abnormalities of
cranial nerves, the pyramidal tract, or peripheral motor nerves.
Paraesthesia was noted in three exposed subjects and one control
subject. There was a slightly higher prevalence of diminished
bicipital and patellar tendon reflexes in the exposed group. There
were significant effects on muscle strength (jump test on one foot)
and vibration sensation (tuning fork) when mean values for the
exposed groups were compared with the control group.
Electrophysiological measurements showed no signs of neuropathy in
any worker, but in the concurrently exposed group mean maximal
motor conduction velocities in the posterior tibial nerve were
significantly reduced compared with those of the control group. In
addition, the mean distal latency in the posterior tibial nerve was
significantly increased in both the concurrently and previously
exposed groups.
e) Extraction facilities
Two workers at a hexane extraction facility reported transient
paraesthesia following excessive acute exposure to hexane. The
predominant complaint at the facility was of temporary episodes of
light-headedness and dizziness. The maximum time-weighted average
(8 h) hexane concentration at this site was 92 mg/m3 (26 ppm)
(NIOSH, 1981a). The most frequently reported symptoms in workers
associated with a soybean extraction facility were dizziness,
giddiness, and light-headedness (each reported by 74% of
participants). Headache and weakness were also reported by 56 and
52%, respectively, of cases. Operators had a higher incidence of
sleepiness (41%) than did maintenance workers (0%). Exposure
concentrations in personal breathing zones ranged from 15.5 to 46.5
mg/m3 (4.4 - 13.2 ppm), but leaks from some process equipment may
have led to higher levels of acute exposure (NIOSH, 1983).
f) Maizlish study
Workers with long-term exposure to mixtures of organic solvents
have been reported to experience mild, clinically detectable,
sensory or sensorimotor polyneuropathies (Husman & Karli, 1980).
In a study by Maizlish et al. (1987), there was no association
between impaired neurological function and solvent concentration in
mixed exposure to solvents (isopropanol, naphtha, hexane) in
workers occupationally exposed for an average of 6 years. Of 240
workers exposed to mixtures of these solvents, with hexane at
breathing-zone concentrations of 3.5 - 137 mg/m3 (1 - 39 ppm), 16%
had presumed, but not clinically significant, signs of peripheral
neuropathy (diminished pain response, vibratory sensation, and
ankle reflex). The signs were not related to solvent exposure
concentration. Overall, the relative risk (RR = 1.4) of the
presumed signs of peripheral neuropathy was not statistically
significant.
9.3.1.2 Effects on vision and optic nerves
Blurred vision has been reported in some cases of peripheral
neuropathy attributed to n-hexane exposure (Wada & Okamoto, 1965;
Sobue & Yamamura, 1968; Yamamura, 1969). No effects on visual
acuity or on the optic fundus have been reported in case histories
of peripheral neuropathy where information on ophthalmic
examinations was provided (Wada et al., 1965; Yamamura, 1969;
Shirbae et al., 1974). Constriction of the visual field, optic
neuritis, and optic atrophy were reported in one study on Japanese
vinyl sandal makers with peripheral neuropathy (Rizzuto et al.,
1980).
In a study of the ocular effects of n-hexane, 15 workers (11
males, 4 females) making adhesive bandages or extracting vegetable
oils, aged 30 - 65 (mean age, 46 years) with exposure durations of
5 - 21 years (mean exposure, 12 years), were examined
ophthalmologically and given colour discrimination tests and
fluorescein angiography (Raitta et al., 1978). Eight workers were
from adhesive bandage factories where peak n-hexane air
concentrations were 1489, 2126, and 3992 mg/m3 (423, 604, and 1134
ppm) around a glue-spreading machine, 1866 and 4506 mg/m3 (530 and
1280 ppm) at an opened mixing machine, and 11 440 mg/m3 (3250 ppm)
at floor level near a tape machine. Air levels of n-hexane in the
breathing zone of workers were lower than these values and
n-hexane air levels in the factories were generally below 1760
mg/m3 (500 ppm). Seven individuals worked in the extraction of
vegetable oils at a factory where the n-hexane air concentrations
in the processing room were between 35 and 176 mg/m3 (10 and 50
ppm), although air levels as high as 7040 - 10 560 mg/m3 (2000 -
3000 ppm) could be reached when processing was interrupted. There
was no control group.
In three individuals (two of whom had pre-existing defects)
there were disturbances in visual acuity or field of vision.
Colour vision defects, mainly blue-yellow, were found in 12
workers. There were no cases of optic atrophy. In nine workers
the maculae showed slight pigment dystrophy, in three there were
yellowish dots, and in one the foveal reflex was absent.
Fluorescein angiography showed patchy choroidal underfilling,
capillary widening of the perifoveal arcade, and loss of retinal
pigment around the macula. Colour discrimination defects and
maculopathy appeared to be related.
In a subsequent study, visual evoked potentials and
electroretinograms (ERGs) were recorded in the same 15 workers and
compared with control measurements obtained from 10 healthy
subjects aged 21 - 57 (mean age, 34.9 years) who were not
occupationally exposed to solvents (Seppalainen et al., 1979).
There were significant reductions in peak-to-peak amplitudes for a
number of visual evoked potential components, possibly due to
conduction block in intracerebral axons. The latency of several
components was significantly increased. This was attributed to
partial axonal degeneration in the visual pathways. A significant
decrease in the amplitude of the electroretinogram (ERG) was
reported, with a reduced latency in the "b" wave. The effects on
the ERG in the exposed workers could not be correlated directly
with the effects on retinal pigments and colour discrimination
reported by Raitta et al. (1978). Seppalainen & Raitta (1981)
found electroneuromyographic signs of peripheral neuropathy,
including reduced nerve conduction velocity and/or fibrillation or
long duration motor unit potentials, in 11 of the 15 workers.
Symptoms included "drunkenness" (8), headache (7), tiredness (5),
and paraesthesia in the limbs (5). All workers with paraesthesia
had at least one electrophysiological sign of neuropathy. Abnormal
EEG findings (slow wave abnormalities) were found in 9 of the
workers.
Signs of maculopathy and colour discrimination defects were
noted in 10 out of the 11 workers with abnormal electrophysiological
signs or peripheral neuropathy. Colour discrimination defects were
present in 8 of the 9 workers with EEG abnormalities. There were
altered EMG and EEG traces in 6 of the 12 workers with colour
discrimination defects. The authors suggested that there was a
correlation between maculopathy and peripheral neuropathy.
Abnormal visual evoked potentials were reported in a study of
glue sniffers exposed to n-hexane (Cooper et al., 1985). Abnormal
electrophysiological signs persisted in two individuals who were
retested 6 months after they had stopped inhaling solvents.
However, these individuals had been exposed to a wide range of
organic solvents and exposure information was inadequate.
In somatosensory evoked potentials, both the absolute latencies
and central conduction time were prolonged in affected workers
(Chang, 1987). In a study of 25 patients, 5 subclinical cases, and
45 apparently healthy workers, Chang & Yip (1987) reported various
electrophysiological abnormalities that reflected, in
polyneuropathy patients, pathological changes of primary giant
axonal degeneration with secondary myelinopathy. This was based on
the observation of a marked fall in nerve conduction velocities,
profound amplitude reduction of muscle action potentials and
sensory action potentials and, prolongation of latencies. Early
electroneurographic changes in the apparently healthy individuals
exposed to n-hexane were reduction in the nerve conduction
velocities, prolongation of motor distal latency, and attenuation
of sensory action potential amplitude.
9.3.2. Effects on the kidney
An investigation of kidney function was conducted on a group of
64 workers (age and sex not reported) exposed for 1 - 24 years to
hydrocarbon mixtures at a shoe factory and an undefined control
group of 80 subjects (Nakajima & Murayama, 1985). Mean air
concentrations for solvents in the factory over a period of one
year were: n-hexane, 250 mg/m3 (71 ppm); cyclohexane, 203 mg/m3
(59 ppm); methyl ethyl ketone, 44 mg/m3 (15 ppm); and ethyl
acetate, 180 mg/m3 (50 ppm). No effects on mean total urinary
protein, albumin, beta-glucuronidase, or muramidase levels were
reported.
A study of kidney function was also undertaken on a group of
182 Italian shoe workers selected from 185 workers employed at four
factories (59 males, 123 females; mean age, 39.4 years; mean
exposure duration, 10.2 years) (Mutti et al., 1981; Franchini et
al., 1983). Breathing-zone concentrations were estimated for a
number of solvents (hexane, ethyl acetate, cyclohexane, methyl
ethyl ketone, and acetone), and concentrations of hexane ( n-hexane
and 2- and 3-methylpentane) were reported to be significantly
higher than 100 ppm (352 mg n-hexane per m3) on a number of
occasions. Eighty healthy subjects (30 males, 50 females; mean
age, 37.9 years) without a history of renal disease or exposure to
organic solvents were used as a control group. A further control
group of 30 unexposed workers (10 males, 20 females; mean age, 41.6
years) was randomly selected from 124 workers who had left the
factories during the previous 5 years (mean exposure duration, 8.8
years).
Spot samples of urine were analysed for total protein, albumin,
and creatinine levels and for beta-glucuronidase activity. Urinary
lysozyme activity was estimated in a few workers. Blood samples
were analysed for creatinine levels. The mean total urinary
protein level was significantly higher in the exposed workers than
in either control or historically exposed workers. Seven workers
(4 males, 3 females) had abnormally high levels (greater than two
standard deviations of the mean control value). Four of these
workers also had abnormally high urinary lysozyme activity, and two
had increased beta-glucuronidase activity. There were no effects
on urinary albumin or serum creatinine levels.
9.3.3. Effects on other organs
Health surveillance studies were carried out in Japan following
a mass outbreak of polyneuropathy in 1967 (Sobue & Yamamura, 1968,
Iida et al., 1969, Yamamura, 1969; Inoue et al., 1970, 1971; Iida &
Yamamoto, 1973).
No effects on the respiratory system, kidney, liver, or
haematopoietic organs were reported.
Laboratory examinations were inconsistent. Urinalysis showed
positive urobilinogen in 15 cases and raised coproporphyrin levels
in four cases. The cephalin cholesterol flocculation test was
positive in 9 cases and a slight to moderately raised serum lactate
dehydrogenase level was found in 17 cases. Depressed cholinesterase
levels were reported in 26 out of 42 cases examined. There was no
correlation between the depressed cholinesterase levels and severity
of neuropathy. The cerebrospinal fluid was examined in 7 cases and
was normal, with no significant effect on pressure or protein
levels. Plethysmography was performed in 11 cases and weak
vasomotor responses were found in two cases.
10. EVALUATION OF HUMAN HEALTH RISKS AND EFFECTS ON THE ENVIRONMENT
10.1. Animal Studies
10.1.1. Short- and long-term exposure
n-Hexane is of low acute toxicity, LD50 values in the range
15 - 30 g/kg and an LC50 (1 h) of 271 040 mg per m3 having been
reported.
In essentially continuous inhalation studies in rats, deaths
occurred following n-hexane exposure to 3520 mg per m3 (1000 ppm)
for up to 11 weeks, and reduced body weight gain after 3 weeks of
exposure to 1760 mg/m3 (500 ppm). In studies involving daily 12-h
exposures to n-hexane, deaths were recorded at 10 560 mg/m3 (3000
ppm) after 15 weeks and slightly reduced body weight gain at 740 mg
per m3 (200 ppm) after 24 weeks. No effects on body weight were
noted in rats exposed to 443 mg/m3 (126 ppm) virtually continuously
for 34 weeks.
Testicular toxicity and peripheral and central neurotoxicity
appear to be the principal non-lethal effects of repeated n-hexane
exposure in rats. Severe testicular lesions have resulted from
inhalation exposure to n-hexane and oral exposure to
2,5-hexanedione. Effects have been attributed to disruption of the
cytoskeleton of Sertoli cells. There are secondary effects on
post-spermatogonial germ cells, which disappear from affected
tubules. Testicular effects were reversible after a single
exposure for 24 h to 17 600 mg/m3 (5000 ppm) but irreversible after
a 2-weeks exposure to the same concentration for 16 h/day, 6
days/week. 2,5-Hexanedione at 1% in drinking water produced
reversible testicular lesions after 2 to 3 weeks of dosing and
irreversible effects (within 17 weeks) after 5 weeks of dosing.
Clinically, neurotoxicity is characterized by hindlimb
weakness, which can progress to paralysis and eventually to
quadriplegia. Clinical signs have been seen after 10 weeks of
discontinuous exposure to 10 560 mg/m3 (3000 ppm) (12 h/day, 7
days/week) and after 6 - 10 weeks of continuous exposure to 1408 to
2112 mg/m3 (400 to 600 ppm).
Lesions (axonal swellings) in hindlimb nerves, brain, and
spinal cord have been observed prior to the appearance of hindlimb
weakness. More severe effects (axonal degeneration and loss) were
noted, particularly in the longest nerves with the largest
diameter, after 6 months of exposure to 1760 mg/m3 (500 ppm).
Overall, no-observed-adverse-effect levels for n-hexane-induced
neurotoxicity in rats are difficult to establish because of the
range of vapour concentrations and exposure regimes tested, and the
variety of effects investigated. In essentially continuous 6-month
inhalation studies, nerve lesions were seen at n-hexane
concentrations of 1760 mg per m3 (500 ppm) or more, but no clinical
or pathological effects were noted at 440 mg/m3 (125 ppm).
Discontinuous exposure of rats to 3168 mg/m3 (900 ppm) for 72 weeks
did not cause any apparent nerve lesions, but there was some
preliminary evidence for an effect on grip strength.
Resolution (partial or complete) of clinical signs of toxicity,
reduced grip strength, and electrophysiological effects can occur
following cessation of exposure. Only limited recovery of
amplitude of the fifth component of the brainstem auditory-evoked
response component (believed to reflect central nervous system
activity) and tail nerve action potential was recorded 15 to 22
weeks after the end of repeated exposure to 3520 mg/m3 (1000 ppm)
for 11 weeks. Resolution of pathological lesions has not been
systematically investigated following inhalation exposure, but
remyelination and limited axonal regeneration have been observed
following cessation of exposure to 1408 to 2112 mg/m3 (400 to 600
ppm).
Inhalation of 1760 mg/m3 (500 ppm) discontinuously for 7 weeks
can cause minor damage to the lungs of rats. In rabbits, severe
respiratory tract lesions were seen following exposure to 10 560
mg/m3 (3000 ppm), 8 h/day for 8 days, and papillary proliferations
of non-ciliated bronchiolar cells were noted after 24 weeks of
exposure.
Effects on the kidney and liver have been reported in a few
inhalation studies on rats but only at higher exposure levels.
Most lesions and other effects were considered to be of minor
toxicological significance and/or were not reproducible in repeated
studies. However, severe liver lesions have been noted in rats
following repeated intraperitoneal administration of n-hexane.
Combined exposure studies have shown that the neurotoxic
effects of n-hexane vapour can be enhanced in hens by methyl
isobutyl ketone (MIBK) and in rats by both methyl ethyl ketone and
lead acetate, but decreased by toluene. Pulmonary lesions in rats
have also been reported to be enhanced by co-exposure to methyl
ethyl ketone.
There have been few studies on the effects of oral or dermal
exposure to n-hexane. Daily oral administration of 500 mg/kg per
day for 4 weeks resulted in reduced tail nerve conduction velocity
in rats.
10.1.2. Genotoxicity
Only limited mutagenicity testing has been conducted.
Chromosomal damage (polyploidy in one study, structural aberrations
in a second study) was reported in in vitro assays in which a
Chinese hamster cell line was exposed to undiluted liquid n-hexane
in the absence of a metabolizing fraction. No significant increase
in point mutation rate occurred in cultured mammalian cells or
bacteria following exposure to n-hexane. DNA damage, as measured
in differential bacterial toxicity assays, was not increased by
exposure to n-hexane. Similarly, no increase in unscheduled DNA
synthesis was noted in human lymphocytes in vitro. Although
n-hexane has given negative results in assays for point mutations
there is some evidence that it can produce chromosomal aberrations
in vitro. Chromosomal damage (chromatid breaks) in rat bone marrow
cells was reported in an in vivo study.
10.1.3. Carcinogenicity
The carcinogenic potential of n-hexane has not been adequately
studied. In a limited study, dermally applied n-hexane did not
appear to enhance the formation of skin tumours in mice following
exposure to a tumour initiator (7,12-dimethylbenzanthracene) or
co-exposure with a promoting agent (croton oil).
10.1.4. Reproductive effects
The reproductive toxicity of n-hexane has not been adequately
investigated. There is no substantial evidence of embryotoxicity
or teratogenicity in rats following inhalation or in mice after
oral dosing. Slight increases in the incidence of some anomalies
were noted in rats exposed to 10 560 mg/m3 (3000 ppm), but
inadequacies in the conduct of the study (e.g., the concentration
tested was probably not high enough) limit the conclusions that can
be reached. Signs of fetotoxicity (reduced pup weight) were noted
in mice, but only at doses that resulted in significant maternal
mortality. A transient delay in postnatal development was reported
in one study in the pups of rats exposed to 10 560 mg/m3 (3000
ppm). Although no signs of neuropathy were reported in these
animals, it is not possible to assess from this study whether in
utero exposure could induce neuropathy, as the animals were only
exposed to n-hexane during a small part of the gestation period.
Other reports have claimed that in utero exposure to n-hexane
retards cerebellar histogenesis and neuronal maturation and induces
electrophysiological abnormalities of the central nervous system in
neonatal rats. However, this work has not been reported in
sufficient detail for a full assessment to be made.
There is evidence that n-hexane causes irreversible sterility
in the rat. No effects on male fertility were reported in a
limited dominant lethal study on the mouse. No investigations of
fertility in female animals or multi-generation studies have been
reported.
10.2. Human Studies
10.2.1. Effects on the nervous system
Many studies have linked occupational n-hexane exposure to the
incidence of peripheral neuropathy in humans. However, few of
these studies reported air concentrations, those reported may be
inaccurate, and it is often not clear whether these concentrations
are of hexane or n-hexane. In addition, industrial exposure to
n-hexane usually occurs during the use of solvent-based products,
so workers are exposed to a mixture of volatile compounds (e.g.,
ethyl acetate, methyl ethyl ketone, toluene, acetone, and other
aliphatic hydrocarbons). Air concentrations for the other solvents
present have rarely been documented. Only those studies that
present some air concentration data have been included in this
review.
The results of peripheral nerve electrophysiology may vary
considerably depending on the type of technique used, the ambient
temperature at which measurements are taken, the segment of the
nerve studied, and the age of the individual. Thus, the assessment
of studies that rely on peripheral nerve electrophysiology is
difficult in the absence of information on these factors.
Studies on 93 cases of neuropathy characterized the clinical
features of n-hexane neuropathy and showed that electro-
physiological measurements correlated well with the severity of the
condition. An association between increasing severity of the
disease and increasing exposure was found. However, from the
limited details of exposure reported, it was not possible to
establish any definite relationship.
Mild subclinical signs of neuropathy were found in shoe workers
exposed to 70 mg/m3 (20 ppm). The degree of electrophysiological
abnormalities increased with duration of exposure. However, no
cases of overt neuropathy occurred, even in workers with an
exposure duration longer than 5 years.
Subjective and electrophysiological signs of subclinical
peripheral neuropathy were found in a group of workers exposed to
approximately 317 mg/m3 (90 ppm). However, these effects cannot be
wholly ascribed to exposure to this level of n-hexane, since
previous exposure to solvents was probably underestimated and
worker self-selection may have introduced bias. In addition, there
may have been co-exposure to methyl ethyl ketone, which has been
demonstrated to enhance the neurotoxic effects of n-hexane.
Exposure to a solvent mixture containing an n-hexane level of
approximately 690 mg/m3 (196 ppm) resulted in reduced nerve
conduction velocity along peripheral nerves, which persisted after
exposure had been significantly reduced. In addition, an effect of
n-hexane on the central nervous system has been suggested to be the
cause of some of the somatosensory evoked potential changes noted.
Subjective and electrophysiological signs of mild subclinical
neuropathy occurred in a small group of workers exposed to mean
n-hexane concentrations in air of 208 mg/m3 (59 ppm). Clinical
overt neuropathy was not detected. In this study exposure to other
solvents was limited and the nerve conduction studies were
performed under defined conditions. However, the small number of
subjects studied, lack of historical exposure data, and the poor
matching of the exposed and control groups limit the value of the
conclusions reached. Nevertheless, the results are in general
agreement with the data obtained in the earlier studies reported
above. Exposure to 669 mg/m3 (190 ppm) for periods in excess of 8
h/day was associated with the onset of clinically overt peripheral
neuropathy. Partial recovery of nerve function can occur slowly
over several years. Exposure to less than 352 mg/m3 (100 ppm) did
not result in neuropathy but showed electrophysiological evidence
of subclinical effects.
Colour discrimination defects and signs of maculopathy have
been described in a small number of workers occupationally exposed
to a wide range of n-hexane levels in air. Visual-evoked potential
and electroretinogram measurements of these individuals showed
abnormalities compared with a control group of non-exposed workers.
Abnormal visual-evoked potential traces have also been reported in
some glue sniffers exposed to n-hexane. It has been suggested that
axonal degeneration may occur in the visual pathway and that
n-hexane may induce a central nerve conduction block. There is
also some evidence that signs of maculopathy may be correlated with
electrophysiological signs of peripheral neuropathy. Abnormal EEG
traces were also found in some of the individuals studied.
10.2.2. Other effects
No studies have been reported on fertility or possible
testicular effects in males. Very little information is available
on the acute toxicity of n-hexane to humans. Most studies have
involved exposure to solvent mixtures. The available data suggest
that n-hexane is of low acute toxicity.
Very few studies of skin or eye irritancy involving volunteers
have been published. n-Hexane is a mild irritant causing transient
erythema when in contact with human skin for short periods. More
severe effects (erythema and blistering) were documented after
occlusive skin contact for 5 h to commercial grade hexane.
Insufficient data are available to assess whether the liquid can
cause eye irritation in human beings. No signs of eye irritation
were noted in a study in which volunteers were exposed to a hexane
vapour concentration of 1760 mg/m3 (500 ppm) for 3 - 5 min. There
have been no reports of skin sensitization in exposed workers and
no skin sensitization was noted in a maximization test with
n-hexane.
10.3. Environmental Effects
n-Hexane has low solubility in water and is highly volatile.
Exposure of aquatic organisms is, therefore, likely to be low and
transient except for uncontrolled discharges into surface waters.
Data on the toxicity of n-hexane to aquatic organisms are few,
variable, conducted under inappropriate conditions and inadequate
to make an assessment of this environmental risk.
11. RECOMMENDATIONS
11.1. Human Health Protection
At the levels of n-hexane to which the general population is
exposed, there is unlikely to be any hazard. In the occupational
health context, atmospheric levels should be kept below the
recommended occupational exposure limits by suitably designed work
processes and engineering controls including ventilation. Suitable
protective clothing and respiratory protection should be readily
available for use in enclosed spaces, in emergencies, and for
certain maintenance operations. The use of a supplied-air
respirator or a self-contained breathing apparatus in continuous
flow mode for n-hexane concentrations up to 8800 mg/m3 (2500 ppm)
is recommended. It should be fitted with a full facepiece for
concentrations between 8800 mg/m3 (2500 ppm) and 17 600 mg/m3 (5000
ppm). Although protective gloves may resist penetration by
n-hexane, mixtures of n-hexane with other solvents, such as methyl
ethyl ketone, may permit the n-hexane to penetrate gloves and other
protective clothing. This factor should be considered when using
n-hexane mixtures.
11.2. Environmental Protection
n-Hexane is unlikely to present a hazard to the environment,
except in the case of major spills or discharges where there could
be transient local effects.
12. FURTHER RESEARCH
a) A study of indices of testicular function should be made in an
occupationally exposed group(s) to establish whether any or all
of the effects reported to occur in the testes of animals are
also manifested in man.
b) The genotoxic effects of n-hexane should be further
investigated in vivo in order to supplement the limited in
vitro information on genotoxicity.
c) A repeat dose study is required to establish the toxicity of
n-hexane given by the oral route. This is needed to provide a
NOEL in order to establish an ADI.
d) The mechanism of neurotoxicity at the molecular level is not
clear. Understanding the mechanism will contribute directly to
the validity of risk assessment. In vivo and in vitro studies
are required to investigate further and clarify these mechanisms.
e) Available teratogenicity studies are inadequate for an
evaluation of risk. Studies should be undertaken to provide
usable data.
f) Dose-response relationships concerning neurotoxic effects of
n-hexane in humans are unclear because of incomplete or
unreliable exposure data. Moreover, although peripheral
neurotoxicity by n-hexane has been amply demonstrated, effects
on the central nervous system have been explored only to a
limited extent. Further epidemiological investigations such as
prospective follow-up studies should be undertaken in order to
overcome the shortcomings of previous investigations.
REFERENCES
ABBRITTI, G., SIRACUSA, A., CIANCHETTI, C., COLI, C.A., CURRADI,
F., PERTICONI, G.F., & DE ROSA, F. (1976) Shoe makers'
polyneuropathy in Italy: the aetiological problem. Br. J. ind.
Med., 33: 92-99.
ABDEL-RAHMAN, M.S., HETLAND, L.B., & COURI, D. (1976) Toxicity and
metabolism of methyl n-butyl ketone. Am. Ind. Hyg. Assoc. J., 37:
95-102.
ABE, K., MISUMI, J., KAWAKAMI, M., & NOMURA, S. (1980) Effects of
n-hexane, methyl n-butyl ketone, and 2,5-hexanedione on the
excitability of sweat glands in rats to mecholyl. Jpn. J. ind.
Health, 22: 380-381.
ABOU-DONIA, M.B., LAPADULA, D.M., CAMPBELL, G., & TIMMONS, P.R.
(1985a) The synergism of n-hexane-induced neurotoxicity by methyl
isobutyl ketone following subchronic (90 days) inhalation in hens:
Induction of hepatic microsomal cytochrome P-450. Toxicol. appl.
Pharmacol., 81: 1-16.
ABOU-DONIA, M.B., MAKKAWY, H.M., & CAMPBELL, G.M. (1985b) Pattern
of neurotoxicity of n-hexane, methyl n-butyl ketone,
2,5-hexanediol, and 2,5-hexanedione alone and in combination with
0-ethyl 0-4-nitrophenyl phenylphosphonothioate in hens. J. Toxicol.
environ. Health, 16: 85-100.
ABOU-DONIA, M.B., LAPADULA, D.M., & SUWITA, E. (1988) Cytoskeletal
proteins as targets for organophosphorus compound and aliphatic
hexacarbon-induced neurotoxicity. Toxicology, 49: 469-477.
ACGIH (1986) Documentation of the threshold limit values, 5th ed.,
Cincinnati, Ohio, American Conference of Governmental Industrial
Hygienists.
AHONEN, I. & SCHIMBERG, R.W. (1988) 2,5-hexanedione excretion after
occupational exposure to n-hexane. Br. J. ind. Med., 45: 133-136.
AIELLO, I., ROSATI, G., SERRA, G., & MANCA, M. (1980) Subclinical
neuropathic disorders and precautionary measures in the shoe
industry. An electrophysiological investigation. Acta neurol.
(Napoli), 35: 285-292.
ALTENKIRCH, H. & SCHULTZ, H. (1979) [Glue sniffing and neuropathy.
Neurological findings and social data from 40 cases.] Nervenarzt,
50: 21-27 (in German).
ALTENKIRCH, H., MAGER, J., STOLTENBURG, G., & HELMBRECHT, J. (1977)
Toxic polyneuropathies after sniffing a glue thinner. J. Neurol.,
214: 137-152.
ALTENKIRCH, H., STOLTENBURG-DIDINGER, G., & WAGNER, H.M. (1978)
Experimental studies on hydrocarbon neuropathies induced by methyl
ethyl ketone (MEK). J. Neurol., 219: 159-170.
ALTENKIRCH, H., STOLTENBURG-DIDINGER, G., & WAGNER, H.M. (1979)
Experimental data on the neurotoxicity of methyl ethyl ketone
(MEK). Experientia (Basel), 35: 503-504.
ALTENKIRCH, H., WAGNER, H.M., STOLTENBURG, G., & SPENCER, P.S.
(1982) Nervous system responses of rats to subchronic inhalation of
n-hexane and n-hexane plus methyl ethyl ketone mixtures. J. neurol.
Sci., 57: 209-219.
ANDERSON, R.J. & DUNHAM, C.B. (1984) Electrophysiologic deficits in
peripheral nerve as a discriminator of early hexacarbon
neurotoxicity. J. Toxicol. environ. Health, 13: 835-843.
ANGELO, M.J. (1977) The pharmacokinetics of the neurotoxin
2,5-hexanedione: distribution, elimination and model simulations,
Newark, University of Delaware (Ph.D Thesis).
ANGELO, M.J. & BISCHOFF, K.B. (1982) A physiologically based
pharmacokinetic model for 2,5-hexanedione. In: Proceedings of the
12th Conference on Environmental Toxicology, 1981, Dayton, Ohio,
Aerospace Medical Research Laboratory, pp. 250-286 (Paper No. 16)
(AFAMRL-TR-81-149).
ANTHONY, D.C., BOEKELHEIDE, K., & GRAHAM, D.G. (1983) The effect of
3,4-dimethyl substitution on the neurotoxicity of 2,5-hexanedione.
I. Accelerated clinical neuropathy is accompanied by more proximal
axonal swellings. Toxicol. appl. Pharmacol., 71: 362-371.
API (1978) 26 week inhalation toxicity study of n-hexane in the
rat, Washington, DC, American Petroleum Institute (API Medical
Research Report No. 28-30077).
API (1983a) Six month continuous inhalation exposures of rats to
hexane mixtures - phase I, Washington, DC, American Petroleum
Institute (API Medical Research Publication No. 30-32858).
API (1983b) Six month continuous inhalation exposures of rats to
hexane mixtures - phase II, Washington, DC, American Petroleum
Institute (API Medical Research Publication No. 30-32846).
ATKINSON, R., ASCHMANN, S.M., CARTER, W.P.L., WINER, A.M., & PITTS,
J.N., Jr (1982) Kinetics of the reactions of OH radicals with
n-alkanes at 299 ± 2 K. Int. J. chem. Kinet., 14: 781-788.
BABANOV, G.P. & BABANOV, A.G. (1981) [Permeability of tissues to
alkanes and their relationship with molecular weight, parity of
carbon atoms and branching of hydrocarbon chains.] Gig. Tr. prof.
Zabol., 12: 48-51 (in Russian).
BABANOV, G.P., ABRAMYAN, G.G., ARTEMYEVA, G.N., VERKHOVSKI, L.G., &
SHAINA, V.K. (1988) [Elevated temperature effect on animals'
sensitivity to n-pentane, n-hexane, n-heptane, n-octane and toluene
and their distribution in the tissues.] Gig. Tr. prof. Zabol., 10:
23-25 (in Russian).
BABICH, H., DAVIS, D.L., & ADLER, R. (1982) Updating federal
standards for toxicants: n-hexane as the model. Environ. Monit.
Assess., 2: 287-299.
BAKER, T.S. & RICKERT, D.E. (1981) Dose-dependent uptake,
distribution and elimination of inhaled n-hexane in the Fischer-344
rat. Toxicol. appl. Pharmacol., 61: 414-422.
BAKER, T.S., WHITE, E.L., & RICKERT, D.E. (1980) Differences in the
metabolism of n-hexane in Fischer 344 rats and C57B1 mice.
Pharmacologist, 22: 279.
BARLOTTA, F., MALFITANO, D., GORGONE, G., & INSERRA, A. (1972)
[Electroretinographic study in experimental hexane intoxication.]
Boll. Soc. Ital. Biol. Sper., 48: 115-117 (in Italian).
BARNI-COMPARINI, I., LUNGARELLA, G., & FONZI, L. (1982) Responses
of lung enzyme activities in rabbits following short-term exposure
to n-hexane: correlation between morphological and biochemical
changes. Agents Actions, 12: 737-742.
BARTOLUCCI, G.B., MAZZOTTA, M., MANNO, M., & DEROSA, E. (1977)
[Industrial exposure to TOCP and n-hexane: behaviour of serum
cholinesterase.] Med. Lav., 68: 222-231 (in Italian).
BASTONE, A., FRONTALI, N., MALLOZZI, C., SBRACCIA, M., & SETTIMI,
L. (1987) Cholinesterases in blood plasma and tissues of rats
treated with n-hexane or with its neurotoxic metabolite
2,4-hexanedione. Arch. Toxicol., 61: 138-144.
BATTISTINI, N., LENZI, G.L., ZANETTE, E., FIESCHI, C., BATTISTA,
F., FRANZINELLI, A., & SARTORELLI, E. (1974) [Observations of a
group of cases of glue polyneuropathy (Clinical electromyographic
and experimental contribution).] Riv. Patol. nerv. ment., 95:
871-885 (in Italian).
BATTISTINI, N., CIONI, R., LENZI, G.L., & ZANETTE, E., (1975)
[Study of the H-reflex in glue neuropathy.] Riv. Neurol., 45: 74-79
(in Italian).
BEHNKE, W., HOLLANDER, W., KOCH, W., NOLTING, F., & ZETZSCH, C.
(1988) A smog chamber for studies of the photochemical degradation
of chemicals in the presence of aerosols. Atmos. Environ., 22:
1113-1120.
BERGER, M.L. & SOZERI, T. (1987) Rapid halogenated hydrocarbon
toxicity in isolated hepatocytes is mediated by direct solvent
effects. Toxicology, 45: 319-330.
BERNARD, A.M., DE RUSSIS, R., NORMAND, J.-C., & LAUWERYS, R.R.
(1989) Evaluation of the subacute nephrotoxicity of cyclohexane and
other industrial solvents in the female Sprague-Dawley rat.
Toxicol. Lett., 45: 271-280.
BOBRA, A.M., WAN, Y.S., & MACKAY, D. (1983) A predictive
correlation for the acute toxicity of hydrocarbons and chlorinated
hydrocarbons to the water flea (Daphnia magna). Chemosphere, 12:
1121-1129.
BOCK, F.G. & CHORTYK, O.T. (1970) Tumor promoting activity of
aliphatic hydrocarbons. Proc. Am. Assoc. Cancer Res., 11: 9.
BOEKELHEIDE, K. (1987) 2,5-Hexanedione alters microtubule assembly.
1. Testicular atrophy, not nervous system toxicity, correlates with
enhanced tubulin polymerization. Toxicol. appl. Pharmacol., 88:
379-382.
BOEKELHEIDE, K. (1988a) Rat testis during 2,5-hexanedione
intoxication and recovery. I. Dose response and the reversibility
of germ cell loss. Toxicol. appl. Pharmacol., 92: 18-27.
BOEKELHEIDE, K. (1988b) Rat testis during 2,5-hexanedione
intoxication and recovery. II. Dynamics of pyrrole reactivity,
tubulin content and microtubule assembly. Toxicol. appl.
Pharmacol., 92: 28-33.
BOEKELHEIDE, K. & EVELETH, J. (1988) The rate of 2,5-hexanedione
intoxication, not total dose, determines the extent of testicular
injury and altered microtubule assembly in the rat. Toxicol. appl.
Pharmacol., 94: 76-83.
BOHLEN, P., SCHLUNEGGER, U.P., & LAUPPI, E. (1973) Uptake and
distribution of hexane in rat tissues. Toxicol. appl. Pharmacol.,
25: 242-249.
BORGATTI, A.R., TRIGARI, G., VENTRELLA, V., & PAGLIARANI, A. (1981)
[Interactions of n-alkanes with respiration and oxidative
phosphorylation of rabbit cardiac mitochondria: n-hexane.] Boll.
Soc. Ital. Biol. Sper., 56: 1569-1575 (in Italian).
BRAIER, L. (1973) A comparative study of isocyclic hydrocarbons in
animals and in man. Haemalogica, 58: 491-500.
BRAVACCIO, F., AMMENDOLA, A., BARRUFFO, L., & CARLOMANGO, S. (1981)
H-reflex behaviour in glue ( n-hexane) neuropathy. Clin. Toxicol.,
18: 1369-1375.
BRINGMANN, G. & KUHN, R. (1982) Results of toxic action of water
pollutants on Daphnia magna tested by an improved standardized
procedure. Z. Wasser Abwasser Forsch., 15: 1-6.
BRUGNONE, F. & PERBELLINI, L. (1980) [Pollution by solvents and
workers' positions in shoe and shoe upper factories.] Med. Lav.,
71: 343-352 (in Italian).
BRUGNONE, F., PERBELLINI, L., GRIGOLINI, L., & APOSTOLI, P. (1978)
Solvent exposure in a shoe upper factory. I. n-Hexane and acetone
concentrations in alveolar and environmental air and in blood. Int.
Arch. occup. environ. Health, 42: 51-62.
BRUGNONE, F., PERBELLINI, L., GAFFURI, E., & APOSTOLI, P. (1980)
Biomonitoring of industrial solvent exposures in workers' alveolar
air. Int. Arch. occup. environ. Health, 47: 245-261.
BRUGNONE, F., PERBELLINI, L., & APOSTOLI, P. (1984) Blood
concentration of solvents in industrial exposure. Collect. Méd.
lég. Toxicol. méd., 125(Part 1): 165-168.
BUITATTI, E., CECCHINI, S., RONCHI, O., DOLARA, P., & BULGARELLI,
G. (1978) Relationship between clinical and electromyographic
findings and exposure to solvents, in shoe and leather workers. Br.
J. ind. Med., 35: 168-173.
BUS, J.S. & TYL, R.W. (1979) Perinatal toxicity of n-hexane in
Fischer 344 rats. Teratology, 19: 22A.
BUS, J.S., WHITE, E.L., TYL, R.W., & BARROW, C.S. (1979) Perinatal
toxicity and metabolism of n-hexane in Fischer-344 rats after
inhalation exposure during gestation. Toxicol. appl. Pharmacol.,
51: 295-302.
BUS, J.S., WHITE, E.L., GILLIES, P.J., & BARROW, C.S. (1981) Tissue
distribution of n-hexane, methyl n-butyl ketone and 2,5-hexanedione
in rats after single or repeated inhalation exposure to n-hexane.
Drug Metab. Dispos., 9: 386-387.
BUS, J.S., DEYO, D., & COX, M. (1982) Dose-dependent disposition of
n-hexane in F-344 rats after inhalation exposure. Fundam. appl.
Toxicol., 2: 226-229.
CCOHS (1985) n-Hexane, Hamilton, Ontario, Canadian Centre for
Occupational Health and Safety (Chemical Hazard Summary No. 11).
CARAPELLA, C. (1977) [Preventive aspects of toxic polyneuropathy in
shoemakers.] Ann. Ist. Super. Sanità, 13: 353-366 (in Italian).
CARLOMAGNO, S., D'ANGELI, A., & MENEGOZZO, M. (1983) [ n-Hexane
polyneuropathy in the shoe making and leather industries.] Med.
Lav., 74: 111-122 (in Italian).
CAVANAGH, J.B. (1982) The pattern of recovery of axons in the
nervous system of rats following 2,5-hexanediol intoxication. A
question of rheology? Neuropathol. appl. Neurobiol., 8: 19-34.
CAVANAGH, J.B. & BENNETTS, R.J. (1981) On the pattern of changes in
the nervous system produced by 2,5-hexanediol. A topographical
study by light microscopy. Brain, 104: 297-318.
CAVENDER, F.L., CASEY, H.W., SALEM, H., GRAHAM, D.G., SWENBERG, J.,
& GRALLA, E.J. (1984) A 13-week vapour inhalation study of n-hexane
in rats with emphasis on neurotoxic effects. Fundam. appl.
Toxicol., 4: 191-201.
CEC (1979) Analysis of organic micropollutants in water, 3rd ed.,
Luxembourg, Commission of the European Communities, Vol. 1, p. 155
(COST 646 bis).
CHANG, Y.C. (1985) Evoked potential study in n-hexane
polyneuropathy. J. Neurol., 22(Suppl.): 200.
CHANG, Y.C. (1987) Neurotoxic effects of n-hexane on the human
central nervous system: evoked potential abnormalities in n-hexane
polyneuropathy. J. Neurol. Neurosurg. Psychiatry, 50: 269-274.
CHANG, Y.C. & YIP, P.K. (1987) n-Hexane-induced electroneurographic
changes and early detection of n-hexane intoxication. J. Formosan
Med. Assoc., 86: 194-200.
CHAPIN, R.E., NORTON, R.M., POPP, J.A., & BUS, J.S. (1982) The
effects of 2,5-hexanedione on reproductive hormones and testicular
enzyme activities in the F-344 rat. Toxicol. appl. Pharmacol., 62:
262-272.
CHAPIN, R.E., MORGAN, K.T., & BUS, J.S. (1983) The morphogenesis of
testicular degeneration induced in rats by orally administered
2,5-hexanedione. Exp. mol. Pathol., 38: 149-169.
CHAUPLANNAZ, G., BADY, B., KOPP, N., LEVRAT, R., & TRILLET, M.
(1982) Neuropathie périphérique au n-hexane chez une toxicomane.
Rev. neurol. (Paris), 138(3): 249-257.
CHIOU, C.T., KILE, D.E., & MALCOLM, R.L. (1988) Sorption of vapours
of some organic liquids on soil humic acid and its relation to
partitioning of organic compounds in soil organic matter. Environ.
Sci. Technol., 22: 298-303.
CIANCHETTI, C., ABBRITTI, G. PERTICONI, G., SIRACUSA, A., &
CURRADI, F. (1976) Toxic polyneuropathy of shoe industry workers.
J. Neurol. Neurosurg. Psychiatry, 39: 1151-1161.
CIIT (1977) n-Hexane, Research Triangle Park, North Carolina,
Chemical Industry Institute of Toxicology (CIIT Current Status
Reports, No. 1).
CLAYTON, G.D. & CLAYTON, F.E. (1981) Patty's industrial hygiene and
toxicology, New York, Chichester, Brisbane, Toronto, John Wiley and
Sons, Vol. 2B.
CFR (1987a) Food additives permitted for direct addition to food
for human consumption. Code fed. Regul., 21(Part 172): 172.560,
172.894.
CFR (1987b) Listing of color additives exempt from certification:
Subpart B - Drugs. Code fed. Regul., 21(Part 73): 256.
COOPER, R., NEWTON, P., & REED, M. (1985) Neurophysiological signs
of brain damage due to glue sniffing. Electroencephalogr. clin.
Neurophysiol., 60: 23-26.
COURI, D. & MILKS, M. (1982) Toxicity and metabolism of the
neurotoxic hexacarbons n-hexane, 2-hexanone and 2,5-hexanedione.
Annu. Rev. Pharmacol. Toxicol., 22: 145-166.
COURI, D., ABDEL-RAHMAN, M.S., & HETLAND, L.B. (1978)
Biotransformation of n-hexane and methyl n-butyl ketone in guinea
pigs and mice. Am. Ind. Hyg. Assoc. J., 39: 295-300.
CREPET, M., GAFFURI, E., & PICOTTI, G. (1968) [Triarylphosphate
disease in the shoe industry.] Minerva med., 59(78): 4073-4075 (in
Italian).
DAHL, A.R., DAMON, E.G., MAUDERLY, J.L., ROTHENBERG, S.J., SEILER,
F.A., & MCCLELLAN, R.O. (1988) Uptake of 19 hydrocarbon vapors
inhaled by F344 rats. Fundam. appl. Toxicol., 10: 262-269.
DALE, G.H. & DREHMAN, L.E. (1985) Hexane. In: Kirk-Othmer
encyclopedia of chemical technology, 3rd ed., New York, Chichester,
Brisbane, Toronto, John Wiley and Sons, pp 926-930.
DECAPRIO, A.P. (1987) n-Hexane neurotoxicity: A mechanism involving
pyrrole adduct formation in axonal cytoskeletal protein.
Neurotoxicology, 8: 199-210.
DECAPRIO, A.P. & O'NEILL, E.A. (1985) Alterations in rat axonal
cytoskeletal proteins induced by in vitro and in vivo
2,5-hexanedione exposure. Toxicol. appl. Pharmacol., 78: 235-247.
DECAPRIO, A.P., OLAJOS, E.J., & WEBER, P. (1982) Covalent binding
of a neurotoxic n-hexane metabolite: conversion of primary amines
to substituted pyrrole adducts by 2,5-hexanedione. Toxicol. appl.
Pharmacol., 65: 440-450.
DECAPRIO, A.P., BRIGSS, R.G., JACKOWSKI, S.J., & KIM, J.C.S. (1988)
Comparative neurotoxicity and pyrrole-forming potential of
2,5-hexanedione and perdenterio-2,5-hexanedione in the rat.
Toxicol. appl. Pharmacol., 92: 75-85.
DE GROOT, D.M.G. & KEPNER, L.A. (1984) The effects of long term
discontinuous exposure to low levels of n-hexane on rats, Zeist,
The Netherlands, Medical Biological Laboratory, TNO (Conducted on
behalf of Shell Internationale Petroleum Maatschappij BV).
DEICHMANN, W.B. & LE BLANC, T.J. (1943) Determination of the
approximate lethal dose with about six animals. J. ind. Hyg.
Toxicol., 25: 415-417.
DELBRUCK, W.R. & KLUGE, A. (1982) [ The effects of n-hexane in man
and animals], Hamburg, German Society for Petroleum Sciences and
Coal Chemistry (Research Report No. 174-2) (in German).
DEMARTINO, C., MALORNI, W., AMANTINI, M.C., SCORZA BARCELLONA, P.,
& FRONTALI, N. (1987) Effects of respiratory treatment with
n-hexane on rat testis morphology. I. A light microscope study.
Exp. mol. Pathol., 46: 199-216.
DE ROSA, E., BARTOLUCCI, G.B., PERBELLINI, L., BRUGNONE, F., &
RAUSA, G. (1988) Environmental and biological monitoring of
exposure to toluene, styrene and n-hexane. Appl. ind. Hyg., 3:
332-337.
DFG (Deutsche Forschungsgemeinschaft) (1988) Maximum concentrations
at the workplace and biological tolerance values for working
materials: Report No. XXIV of the Commission for the Investigation
of Health Hazards of Chemical Compounds in the Work Area, Weinheim,
Verlag Chemie, p. 84.
DI-BOSCO, M. & FONZI, S. (1974) [So-called glue or shoe makers'
polyneuropathy.] Riv. Infort. Mal. prof., 61(Part 1): 165-192 (in
Italian).
DISCALZI, G.L., PERRELLI, G., & PAVAN, I. (1988) [Solvent exposure
risk among homeworkers.] Med. Lav., 79: 234-236 (in Italian).
DIVINCENZO, G.D., KAPLAN, C.J., & DEDINAS, J. (1976)
Characterization of the metabolites of methyl n-butyl ketone,
methyl isobutyl ketone and methyl ethyl ketone in guinea pig serum
and their clearance. Toxicol. appl. Pharmacol., 36: 511-522.
DIVINCENZO, G.D., HAMILTON, M.L., KAPLAN, C.J., & DEDINAS, J.
(1977) Metabolic fate and disposition of 14C-labelled methyl
n-butyl ketone in the rat. Toxicol. appl. Pharmacol., 41: 547-560.
DIVINCENZO, G.D., HAMILTON, M.L., KAPLAN, C.J., KRASAVAGE, W.J., &
O'DONOGHUE, J.L. (1978) Studies on the respiratory uptake and
excretion and the skin absorption of methyl n-butyl ketone in
humans and dogs. Toxicol. appl. Pharmacol., 44: 593-604.
DIVINCENZO, G.D., KRASAVAGE, W.J., & O'DONOGHUE, J.L. (1980) Role
of metabolism in hexacarbon neuropathy. In: Witschi, H., ed. The
scientific basis of toxicity assessment, Amsterdam, Oxford, New
York, Elsevier Science Publishers, pp. 183-200.
DOLARA, P., FRANCONI, F., & BASOSI, D. (1978) Urinary excretion of
some n-hexane metabolites. Pharmacol. Res. Commun., 10(6): 503-510.
DUNNICK, J.K. (1989) Toxicity studies of n-hexane in B6C3F, mice
(inhalation studies), Washington, DC, US Department of Health and
Human Services, National Toxicology Program (Report No. NTPTS 1).
DUNNICK, J.K., GRAHAM, D.G., YANG, R.S.H., HABER, S.B., & BROWN,
H.R. (1989) Thirteen-week toxicity study of n-hexane in B6C3F, mice
after inhalation exposure. Toxicology, 57: 163-172.
DURHAM, H.D., PENA, S.D.J., & CARPENTER, S. (1983) The neurotoxins
2,5-hexanedione and acrylamide promote aggregation in intermediate
filaments in cultured fibroblasts. Muscle Nerve, 6: 631-637.
DURHAM, H.D., PENA, S.D.J., & ECOBICHON, D.J. (1988)
Hexahydrocarbon effects on intermediate filament organization in
human fibroblasts. Muscle Nerve, 11: 160-165.
EBEN, A., FLUCKE, W., MINAIL, F., THYSSEN, J., & KIMMERLE, G.
(1979) Toxicological and metabolic studies of methyl n-butyl
ketone, 2,5-hexanedione and 2,5-hexanediol in male rats.
Ecotoxicol. environ. Saf., 3: 204-217.
EDELFORS, S. & RAVN-JONSEN, A. (1985) Calcium uptake in rat brain
synaptomsomes after short-term exposure to organic solvents: a
pilot study. Acta pharmacol. toxicol., 56: 431-434.
ELKINS, H.B. (1959) The chemistry of industrial toxicology, 2nd
ed., New York, Chichester, Brisbane, Toronto, John Wiley and Sons.
FAWCETT, P.R.W. & BARWICK, D.D. (1981) Studies in nerve conduction.
In: Walton, J.A., ed. Disorders of voluntary muscle, 4th ed., New
York, Churchill Livingstone, pp. 928-951.
FEDTKE, N. & BOLT, H.M. (1986a) Detection of 2,5-hexanedione in the
urine of persons not exposed to n-hexane. Int. Arch. occup.
environ. Health, 57: 143-148.
FEDTKE, N. & BOLT, H.M. (1986b) Methodological investigations on
the determination of n-hexane metabolites in urine. Int. Arch.
occup. environ. Health, 57: 149-158.
FEDTKE, N. & BOLT, H.M. (1987a) 4,5-dihydroxy-2-hexanone: a new
metabolite of n-hexane and of 2,5-hexanedione in rat urine.
Biochem. environ. mass Spectrom., 14: 563-572.
FEDTKE, N. & BOLT, H.M. (1987b) The relevance of 4,5-dihydroxy-2-
hexanone in the excretion kinetics of n-hexane metabolites in rat
and man. Arch. Toxicol., 61: 131-137
FICHERA, G., SPAGNA, C., & TIMPANARO, V. (1968) [Hydrocarbon
intoxication. Note II. Anatomo-pathological findings in
experimental hexane intoxication.] Folia med. (Napoli), 51: 633-640
(in Italian).
FILSER, J.G., PETER, H., BOLT, H.M., & FEDTKE, N. (1987)
Pharmacokinetics of the neurotoxin n-hexane in rat and man. Arch.
Toxicol., 60: 77-80.
FISEROVA-BERGEROVA, V. & PIERCE, J.T. (1989) Biological monitoring
V: Dermal absorption. Appl. ind. Hyg., 8(4): F14-F21.
FRANCHINI, I., CAVATORTA, A., FALZOI, M., LUCERTINI, S., & MUTTI,
A. (1983) Early indicators of renal damage in workers exposed to
organic solvents. Int. Arch. occup. environ. Health, 52: 1-9.
FRANKE, J.P., WIJSBEEK, J., DE ZEEUZ, R.A., MOLLER, M.R., &
NIERMEYER, H. (1988) Systematic analysis of solvents and other
volatile substances by gas chromatography. J. anal. Toxicol., 12:
20-24.
FROMMER, U., ULLRICH, V., & ORRENIUS, S. (1974) Influence of
inducers and inhibitors on the hydroxylation pattern of n-hexane in
rat liver microsomes. FEBS Lett., 41(1): 14-16.
FRONTALI, N., AMANTINI, M.C., SPAGNOLO, A., GUARCINI, A.M., &
SALTARI, M.C. (1981) Experimental neurotoxicity and urinary
metabolites of the C5-C7 alphatic hydrocarbons used as glue
solvents in shoe manufacture. Clin. Toxicol., 18(12): 1357-1367.
FUHNER, H. (1921) [The narcotic effects of gasoline and its
components (pentane, hexane, heptane, octane).] Biochem. Z., 115:
235-261 (in German).
GENTER, M.B., SZAKAL-QUIN, G., ANDERSON, C.W., ANTHONY, D.C., &
GRAHAM, D.G. (1987) Evidence that pyrrole formation is a
pathogenetic step in gamma-diketone neuropathy. Toxicol. appl.
Pharmacol., 87: 351-361.
GENTER ST. CLAIR, M.B., AMARNATH, V., MOODY, A., ANTHONY, D.C.,
ANDERSON, C.W., & GRAHAM, D.G. (1988) Pyrrole oxidation and protein
cross-linking as necessary steps in the development of gamma-
diketone neuropathy. Chem. Res. Toxicol., 1: 179-185.
GENTRY, S.J. & WALSH, P.T. (1987) Eight-hour TWA personal
monitoring using a diffusive sampler and short-term stain tube. Am.
Ind. Hyg. Assoc. J., 48: 287-292.
GERARDE, H.W. & LINDEN, N.J. (1963) Toxicological studies on
hydrocarbons. IX. The aspiration hazard and toxicity of
hydrocarbons and hydrocarbon mixtures. Arch. environ. Health, 6:
329-341.
GHITTORI, S., IMBRIANI, M., PEZZAGNO, G., & CAPODAGLIO, E. (1987)
The urinary concentration of solvents as a biological indicator of
exposure: proposal for the biological equivalent exposure limit for
nine solvents. Am. Ind. Hyg. Assoc. J., 48: 786-790.
GILLIES, P.J., NORTON, R.M., WHITE, E.L., & BUS, J.S. (1980)
Inhibition of sciatic nerve sterologenesis in hexacarbon-induced
distal axonopathy in the rat. Toxicol. appl. Pharmacol., 54:
217-222.
GIULIANO, G., IANNACCONE, A., & ZAPPOLI, R. (1974)
[Electroencephalographic research in shoe industry workers exposed
to the risk of poisoning from adhesive solvents.] Lav. Um., 26(2):
33-42 (in Italian).
GOEL, S.K., RAO, G.S., & PANDYA, K.P. (1982) Toxicity of n-hexane
and n-heptane: some biochemical changes in liver and serum.
Toxicol. Lett., 14: 169-174.
GOEL, S.K., RAO, G.S., & KRISHNAMURTI, C.R. (1987) Toxicity of
n-hexane and 2,5-hexanediol to the haemopoietic system: a
preliminary report. Xenobiotica, 17: 217-222.
GOEL, S.K., RAO, G.S., & PANDYA, K.P. (1988) Hepatotoxic effects
elicited by n-hexane or n-heptane. J. appl. Toxicol., 8: 81-84.
GOMEZ-ARROYO, S., CASTILLO-RUIZ, P., & VILLALOBOS-PIETRINI, R.
(1986) Chromosomal alterations induced in Vicia faba by different
industrial solvents: thinner, toluene, benyene, n-hexane, n-heptane
and ethyl acetate. Cytologia, 51: 133-142.
GONZALEZ, E.G. & DOWNEY, J.A. (1972) Polyneuropathy in a glue
sniffer. Arch. phys. Med. Rehabil., 53: 333-337.
GOTO, I., MATSUMURA, M., INOUE, N., MURAI, Y., SHIDA, K., SANTA,
T., & KUROIWA, Y. (1974) Toxic polyneuropathy due to glue sniffing.
J. Neurol. Neurosurg. Psychiatry, 37: 848-853.
GRAHAM, D.G. & GOTTFREID, M.R. (1984) Cross species extrapolation
in hydrocarbon neuropathy. Neurobehav. Toxicol. Teratol., 6:
433-435.
GRAHAM, D.G., ANTHONY, D.C., & BOEKELHEIDE, K. (1982a) In vitro and
in vivo studies of the molecular pathogenesis of n-hexane
neuropathy. Neurobehav. Toxicol. Teratol., 4: 629-634.
GRAHAM, D.G., ANTHONY, D.C., BOEKELHEIDE, K., MASCHMANN, N.A.,
RICHARDS, R.G., WOLFRAM, J.W., & SHAW, B.R. (1982b) Studies of the
molecular pathogenesis of hexane neuropathy. II. Evidence that
pyrrole derivatization of lysyl residues leads to protein
crosslinking. Toxicol. appl. Pharmacol., 64: 415-422.
GRAHAM, D.G., ANTHONY, D.C., SZAKAL-QUIN, G., GOTTFREID, M.R., &
BOEKELHEIDE, K. (1985) Covalent crosslinking of neurofilaments in
the pathogenesis of n-hexane neuropathy. Neurotoxicology, 6: 55-64.
GRIFFITHS, I.R., KELLY, P.A.T., CARMICHAEL, S., MCCULLOCH, M., &
WATERSON, M. (1981) The relationship of glucose utilization and
morphological change in the visual system in hexacarbon neuropathy.
Brain Res., 222: 447-451.
GRIFFITH, J.F., NIXON, G.A., BRUCE, R.D., REER, P.J., & BANNAN,
E.A. (1980) Dose-response studies with chemical irritants in the
albino rabbit eye as a basis for selecting optimum testing
conditions for predicting hazard to the human eye. Toxicol. appl.
Pharmacol., 55: 501-513.
GOVERNA, M., CALISTI, R., COPPA, G., TAGLIAVENTO, G., COLOMBI, A.,
& TRONI, W. (1987) Urinary excretion of 2,5-hexanedione and
peripheral polyneuropathies in workers exposed to hexane. J.
Toxicol. environ. Health, 20: 219-228.
GROVES, F.R. (1988) Effect of cosolvents on the solubility of
hydrocarbons in water. Environ. Sci. Technol., 22: 282-286.
GUTHRIE, V.B. (1960) Petroleum products handbook, New York, McGraw-
Hill.
HADJIIVANOVA, N.B., SALOVSKI, P.Z., GROSEVA, M.M.,
CHARAKCHIEVA, S.B., & NECHEV, Ch.K. (1987) Early effects of
n-hexane and irradiation on the lung surfactant system. Acta
physiol. pharmacol. Bulg., 13: 25-29.
HAZELTON LABORATORIES (1981) In vivo and in vitro mutagenicity
studies n-hexane (hexane UV), Vienna, Virginia, Hazelton
Laboratories (Project No. 596-114 and 596-115) (Prepared for the
American Petroleum Institute).
HENDERSON, V.E. & SMITH, A.H.R. (1936) Propylene impurities.
Hexenes and hexanes. J. Pharmacol. exp. Ther., 58: 319-327.
HERSKOWITZ, A., ISHII, N., & SCHAUMBERG, H. (1971) n-Hexane
neuropathy. A syndrome occurring as a result of industrial
exposure. New Engl. J. Med., 285(2): 82-85.
HEWITT, W.R., MIYAJIMA, H., COTE, M., & PLAA, G.L. (1980) Acute
alteration of chloroform induced hepato- and nephrotoxicity by
n-hexane, methyl n-butyl ketone and 2,5-hexanedione. Toxicol. appl.
Pharmacol., 53: 230-248.
HINE, C.H. & ZUIDEMA, H.H. (1970) The toxicological properties of
hydrocarbon solvents. Ind. Med., 39(5): 39-44.
HODGSON, A.T., GIRMAN, J.R., & BINENBOYM, J. (1986) A multisorbent
sampler for volatile organic compounds in indoor air. In:
Proceedings of the 79th Annual Meeting, Minneapolis, 22-27 June,
1986, Minneapolis, Minnesota, Air Pollution Control Association,
pp. 1-15 (Report No. 86-37-1).
HOLMBERG, B. & LUNDBERG, P. (1989) Assessment and management of
occupational risks in the Nordic (Scandinavian) countries. Am. J.
ind. Med., 15: 615-626.
HONMA, T. (1983) Changes in acetylcholine metabolism in rat brain
after a short-term exposure to toluene and n-hexane. Toxicol.
Lett., 16: 17-22.
HONMA, T., MIYAGAWA, M., SATO, M., & HASEGAWA, H. (1982) Increase
in glutamine content of rat midbrain induced by short-term exposure
to toluene and hexane. Ind. Health, 20: 109-115.
HOUK, V.S., SCHALKOWSKY, S., & CLAXTON, L.D. (1989) Development and
validation of the spiral Salmonella assay: an automated approach to
bacterial mutagenicity testing. Mutat. Res., 223: 49-64.
HOWD, R.A., BINGHAM, L.R., STEEGER, T.M., REBERT, C.S., & PRYOR,
G.T. (1982) Relation between schedules of exposure to hexane and
plasma levels of 2,5-hexanedione. Neurobehav. Toxicol. Teratol., 4:
87-91.
HOWD, R.A., REBERT, C.S., DICKINSON, J., & PRYOR, G. (1983) A
comparison of the rates of development of functional hexane
neuropathy in weanling and young adult rats. Neurobehav. Toxicol.
Teratol., 5: 63-68.
HOWELL, W.E. (1979) A neurobehavioural evaluation of the pre-natal
toxicity of n-hexane in rats, Cincinnati, Ohio, University of Ohio,
78 pp (Ph.D Thesis) (Dissertation Abstracts Int B. No. 1610B).
HOWELL, W.E. & COOPER, G.P. (1981) Neurophysiological evaluation of
prenatal n-hexane toxicity. Toxicologist, 1: 152 (Abstact 552).
HUANG, C.-C. & CHU, N.-S. (1989) Evoked potentials in chronic
n-hexane intoxication. Clin. Electroencephalogr., 20: 162-168.
HUSMAN, K. & KARLI, P. (1980) Clinical neurological findings among
car painters exposed to a mixture of organic solvents. Scand. J.
Work Environ. Health, 6: 33-39.
ICHIHARA, K., KUSONOSE, E., & KUSUNOSE, M. (1969) Microsomal
hydroxylation of decane. Biochem. Biophys. Acta, 176: 713-719.
IIDA, M. (1982) Neurophysiological studies of n-hexane
polyneuropathy in the sandal factory. Electroencephalogr. clin.
Neurophysiol., 36(Suppl.): 671-681.
IIDA, M. & YAMAMOTO, H. (1973) [Prognosis of n-hexane
polyneuropathy. Follow-up studies of mass outbreak in District F in
Mie Prefecture.] Igaku Ayumi, 84(4): 199-201 (in Japanese).
IIDA, M., YAMAMURA, Y., & SOBUE, I. (1969) Electromyographical
findings and conduction velocity in n-hexane polyneuropathy.
Electromyography, 9: 247-261.
IKEDA, M. & KASAHARA, M. (1986) n-hexane and benzene contents in
gasoline for industrial purpose. Ind. Health, 24: 63-66.
IKEDA, M., KOIZUMI, A., KASAHARA, M., & FUJITA, H. (1986) Combined
effects of n-hexane and toluene on norepinephrine and dopamine
levels in rat brain tissues after long-term exposures. Bull.
environ. Contam. Toxicol., 36: 510-517.
IMBRIANI, M., GHITTORI, S., BORLINI, F., PEZZAGNO, G., &
CAPODAGLIO, E. (1984a) [ n-hexane and toluene in the urine of
occupationally exposed subjects.] Boll. Soc. Ital. Biol. Sper., 60:
1919-1925 (in Italian).
IMBRIANI, M., GHITTORI, S., PEZZAGNO, G., & CAPODAGLIO, E. (1984b)
n-hexane urine elimination and weighted exposure concentration.
Int. Arch. occup. environ. Health, 55: 33-41.
INOUE, T., TAKEUCHI, Y., TAKEUCHI, S., YAMADA, S., SUZUKI, H.,
MATSUSHITA, T., MIYAGAKI, M., MAEDA, K., & MATSUMOTO, T. (1970)
[Industrial health survey of high incidence of n-hexane
intoxication among vinyl sandal manufacturers.] Ind. Health, 12(3):
73-84 (in Japanese).
INOUE, T., YAMADA, S., MIYAGAKI, H., & TAKEUCHI, Y. (1971)
Polyneuritis caused by n-hexane and the maximum allowable
concentration. In: Proceedings of the XVI International Congress on
Occupational Health, 22-27 September 1969, Tokyo, Industrial Safety
Association, pp. 522-524.
IRPTC (1990) IRPTC Legal file, Geneva, International Register of
Potentially Toxic Chemicals, United Nations Environment Programme.
ISHIDATE, M. & SOFUNI, T. (1984) Primary mutagenicity screening of
food additives currently used in Japan. Food chem. Toxicol., 22(8):
623-636.
IWASAKI, K. & TSURUTA, H. (1984) Molecular mechanisms of hexane
neuropathy. Significant differences in pharmacokinetics between
2,3- 2,4-, and 2,5-hexanedione. Ind. Health, 22: 177-187.
IWATA, M., TAKEUCHI, Y., HISANAGA, N., & ONO, Y. (1983a) Changes of
n-hexane metabolites in urine of rats exposed to various
concentrations of n-hexane and to its mixture with toluene or MEK.
Int. Arch. occup. environ. Health, 53: 1-8.
IWATA, M., TAKEUCHI, Y., HISANAGA, N., & ONO, Y. (1983b) A study on
biological monitoring of n-hexane exposure. Int. Arch. occup.
environ. Health, 51: 253-260.
IWATA, M., TAKEUCHI, Y., HISANAGA, N., & ONO, Y. (1984) Changes of
n-hexane neurotoxicity and its urinary metabolites by long-term co-
exposure with MEK or toluene. Int. Arch. occup. environ. Health, 54:
273-281.
JAKOBSON, I., WAHLBERG, J.E., HOLMBERG, B., & JOHANSSON, G. (1982)
Uptake via the blood and elimination of 10 organic solvents
following epicutaneous exposure of anaesthetized guinea pigs.
Toxicol. appl. Pharmacol., 63: 181-187.
JEPPSSON, R. (1975) Parabolic relationship between lipophilicity
and biological activity of aliphatic hydrocarbons, ethers and
ketones after intravenous injection of emulsion formulations into
mice. Acta pharmacol. toxicol., 37: 56-64.
JONES, H.G. & CAVANAGH, J.B. (1982) Recovery from 2,5-hexanediol
intoxication of the retinotectal tract of the rat. An
ultrastructural study. Acta neuropathol. (Berlin), 58: 286-290.
JUHNKE, I. & LUDEMANN, D. (1978) [Results of trials of 200 chemical
compounds for acute toxicity for fish with the golden orfe test.]
Z. Wasser Abwasser Forsch., 11: 161-164 (in German).
KANNAN, K., SINGH, K.P., GOEL, S., & SHANKER, R. (1985) Effects of
2,5-hexanedione on immunocompetence of mice. Environ. Res., 36: 14-25.
KASAHARA, M. & IKEDA, M. (1987) Spontaneous desorption of organic
solvents from carbon cloth. Ind. Health, 25: 73-81
KASAHARA, M., SUZUKI, H., TAKEUCHI, Y., HARA, I., & IKEDA, M.
(1987) n-Hexane, benzene and other aromatic components in petroleum
distillate solvents in Japan. Ind. Health, 25: 205-214.
KATAGIRI, Y. & OHASHI, T. (1975) Photochemical reactions of
hydrocarbons in smog chamber. Annu. Rep. Kanagawa Prefect. Environ.
Cent., 6: 47.
KATZ, G.V., O'DONOGHUE, J.L., DIVINCENZO, G.D., & TERHAAR, C.J.
(1980) Comparative neurotoxicity and metabolism of ethyl n-butyl
ketone and methyl n-butyl ketone in rats. Toxicol. appl.
Pharmacol., 52: 153-158.
KAWACHI, T., YAHAGI, T., KADA, T., TAZIMA, Y., ISHIDATE, M.,
SASAKI, M., & SUGIYAMA, T. (1982) Co-operative programme on short-
term assays for carcinogenicity in Japan. In: Molecular and
cellular aspects of carcinogen screening tests, Lyon, International
Agency for Research on Cancer, pp. 323-330 (IARC Scientific
Publications No. 27).
KEPLINGER, M.L., LANIER, G.E., & DEICHMANN, W.B. (1959) Effects of
environmental temperature on the acute toxicity of a number of
compounds in rats. Toxicol. appl. Pharmacol., 1: 156-161.
KIMURA, E.T., EBERT, D.M., & DODGE, P.W. (1971) Acute toxicity and
limits of solvent residue for sixteen organic solvents. Toxicol.
appl. Pharmacol., 19: 699-704.
KLIGMAN, A.M. (1966) The identification of contact allergens by
human assay. III. The maximization test: a procedure for screening
and rating contact sensitisers. J. invest. Dermatol., 47: 393-409.
KLIMES, I., VISKUPIC, E., ZORAD, S., SVABOVA, E., KISS, A., &
MACHO, L. (1987) Effect of prolonged n-hexane administration on in
vitro insulin degradation in rat liver and blood. Med. Sci. Res.,
15: 1427-1429.
KLOPFFER, W., HAAG, F., KOHL, E.-G., & FRANK, R. (1988) Testing of
the abiotic degradation of chemicals in the atmosphere: The smog
chamber approach. Ecotoxicol. environ. Saf., 15: 298-319.
KOROBKIN, R., ASBURY, A.K., SUMNER, A.J., & NIELSEN, S.L., (1975)
Glue sniffing neuropathy. Arch. Neurol., 32: 158-162.
KRAMER, A., STAUDINGER, H., & ULLRICH, V. (1974) Effect of n-hexane
inhalation on the monooxygenase system in mice liver microsomes.
Chem.-biol. Interact., 8: 11-18.
KRASAVAGE, W.J., O'DONOGHUE, J.L., & TERHAAR, C.J. (1979) Oral
chronic toxicity of methyl n-propyl ketone, methyl n-butyl ketone
and hexane in rats. Toxicol. appl. Pharmacol., 48(Part 2): A205.
KRASAVAGE, W.J., O'DONOGHUE, J.L., DIVINCENZO, G.D., & TERHAAR,
C.J. (1980) The relative neurotoxicity of methyl n-butyl ketone,
n-hexane and their metabolites. Toxicol. appl. Pharmacol., 52: 433-441.
KRONEVI, T., WAHLBERG, J., & HOLMBERG., B. (1979) Histopathology of
skin, liver and kidney after epicutaneous administration of five
industrial solvents to guinea-pigs. Environ. Res., 19: 56-69.
KULIG, B.M. (1983) Chronic behavioural toxicology of hexane and
CS2: Progress report, Zeist, The Netherlands, Medical Biological
Laboratory, TNO (Conducted on behalf of Shell Internationale
Petroleum Maatschappij BV).
KULIG, B.M. & WOLTHIUS, O.L. (1984) The development of neurological
and behavioural impairments in rats exposed subchronically to
n-hexane. In: Abstracts of the International Conference on Organic
Solvent Toxicity, Stockholm, October 1984.
KULIG, B.M., DEMIK, G., & WOLTHIUS, O.L. (1984) 2,5-Hexanedione
concentrations and hexane-induced neuropathy in Wistar rats.
Abstracts of the Second International Conference on Neurotoxicology
of Selected Chemicals, Chicago, 1983. Neurotoxicology, 5(2): 307.
KULIG, B.M., VANWERSCH, R.A.P., & WOLTHUIS, O.L. (1985) The
automated analysis of coordinated hindlimb movement in rats during
acute and prolonged exposure to toxic agents. Toxicol. appl.
Pharmacol., 80: 1-10.
KURITA, H. (1974) [Experimental studies on the effects of n-hexane
on albino rats.] Ind. Health, 9(8): 24-29 (in Japanese).
LADEFOGED, O. & PERBELLINI, L. (1986) Acetone-induced changes in
the toxicokinetics of 2,5-hexanedione in rabbits. Scand. J. Work.
Environ. Health, 12: 627-629.
LADEFOGED, O., HASS, U., & SIMONSEN, L. (1989) Neurophysiological
and bahavioural effects of combined exposure to 2,5-hexanedione and
acetone or ethanol in rats. Pharmacol. Toxicol., 65: 372-375.
LAFFORT, P. & DRAVNIEKS, A. (1973) An approach to a physico-
chemical model of olfactory stimulation in vertebrates by single
compounds. J. theor. Biol., 38: 335-345.
LAGUENY, A., JULIEN, J., VALLAT, J.M., FERRER, X., DELIAC, P., &
DELIAC, M. (1982) Etude électrophysiologique de deux cas de
polynévrite provoquée par une association n-hexane-méthyléthyl-
cétone. Rev. EEG Neurophysiol., 12: 158-166.
LANKAS, G.R., BAXTER, C.S., & CHRISTIAN, R.T. (1978) Effect of
alkane tumour-promoting agents on chemically induced mutagenesis in
cultured V79 Chinese hamster cells. J. Toxicol. environ. Health, 4:
37-41.
LAPADULA, D.M., IRWIN, R., SUWITA, E., & ABOU-DONIA, M.B. (1986)
Cross-linking of neurofilament proteins of rat spinal cord in vivo
after administration of 2,5-hexanedione. J. Neurochem., 46: 1843-1850.
LAZAREV, N.W. (1929) [The toxicity of various hydrocarbon vapours.]
Arch. exp. Pathol. Pharmakol., 143: 223-233 (in German).
LIJINSKY, W. & RAHA, C.R. (1961) Polycyclic aromatic hydrocarbons
in commercial solvents. Toxicol. appl. Pharmacol., 3: 469-473.
LITTON BIONETICS (1979) Teratology study in rats: n-hexane, Litton
Bionetics (LBI Project No. 20698-9) (Final Report submitted to the
American Petroleum Institute).
LITTON BIONETICS (1980) Mutagenicity evaluation of n-hexane in the
mouse dominant lethal assay, Litton Bionetics (LBI Project No.
21141-01) (Final Report submitted to the American Petroleum
Institute).
LODEN, M. (1986) The in vitro permeability of human skin to
benzene, ethylene glycol, formaldehyde and n-hexane. Acta
pharmacol. toxicol., 58: 382-389.
LONNEMAN, W.A., KOPCZYNSKI, S.L., DARLEY, P.E., & SUTTERFIELD, F.D.
(1974) Hydrocarbon composition of urban air pollution. Environ.
Sci. Technol., 8: 229-236.
LUNGARELLA, G., FONZI, L., & CENTINI, F. (1980) Respiratory tract
lesions induced in rabbits by short-term exposure to n-hexane. Res.
Commun. chem. Pathol. Pharmacol., 29(1): 129-139.
LUNGARELLA, G., BARNI-COMPARINI, I., & FONZI, L. (1984) Pulmonary
changes induced in rabbits by long-term exposure to n-hexane. Arch.
Toxicol., 55: 224-228.
MCAULIFFE, C. (1963) Solubility in water of C1-C9 hydrocarbons.
Nature (Lond.), 200: 1092-1093.
MCCARROLL, N.E., PIPER, C.E., & KEECH, B.H., (1980) Bacterial
microsuspension assays with benzene and other organic solvents.
Environ. Mutagen., 2(2): 281-281.
MCCARROLL, N.E., PIPER, C.E., & KEECH, B.H. (1981a) An E. coli
microsuspension assay for the detection of DNA damage induced by
direct-acting agents and promutagens. Environ. Mutagen., 3: 429-444.
MCCARROLL, N.E., KEECH, B.H., & PIPER, C.E. (1981b) A
microsuspension adaptation of the Bacillus subtilis "rec" assay.
Environ. Mutagen., 3: 607-616.
MCNAMARA, B.C. & JEFCOATE, C.R. (1988) Synergistic stimulation of
pregnenolone synthesis in rat adrenal mitochondria by n-hexane and
cardiolipin. Arch. Biochem. Biophys., 260: 780-788.
MAHMOUD, G. & LACHAPELLE, J.M. (1984) Etude de l'efficacité d'un
gel de protection anti-solvants, vis-à-vis de l'irritation cutanée
provoquée par le n-hexane. Arch. Mal. prof. Méd. Trav. Séc. soc.,
45(4): 223-237.
MAHMOUD, G. & LACHAPELLE, J.M. (1985) Evaluation expérimentale de
l'efficacité de crèmes-barrière et de gels antisolvants dans la
prévention de l'irritation cutanée provoquée par des solvants
organiques. Cah. Méd. Trav., 22(2/3): 163-168.
MAIZLISH, N.A., LANGOLF, G.D., WHITEHEAD, L.W., FINE, L.J., ALBERS,
J.W., GOLDBERG, J., & SMITH, P. (1985) Behavioural evaluation of
workers exposed to mixtures of organic solvents. Br. J. ind. Med.,
42: 579-590.
MAIZLISH, N.A., FINE, L.J., ALBERS, J.W., WHITEHEAD, L., & LANGOLF,
G.D. (1987) A neurological evaluation of workers exposed to
mixtures of organic solvents. Br. J. ind. Med., 44: 14-25.
MALORNI, W., DEMARTINO, C., AMANTINI, M., & FRONTALI, N. (1984)
Effects of subchronic treatment with n-hexane on rat testis. A
morphological study. Ultramicroscopy, 12: 148.
MALORNI, W., FORMISANO, G., & DONELLI, G. (1989) Morphologic
changes induced in vitro by 2,5-hexanedione. In vitro cell. dev.
Biol., 25: 82-90.
MARCHISEPPE, I., VALENTINO, M., GOVERNA, M., & STOCCHI, V. (1989)
Determination of total 2,5-hexanedione by reversed-phase high-
performance liquid chromatography. J. Chromatology, 495: 288-294.
MARKS, T.A., FISHER, P.W., & STAPLES, R.E. (1980) Influence of
n-hexane on embryo and fetal development in mice. Drug chem.
Toxicol., 3(4): 393-406.
MELLAN, I., ed. (1977) Industrial solvents handbook, 2nd ed., Park
Ridge, New Jersey, Noyes Data Corporation, p. 14.
MEDINILLA, J., DE & ESPIGARES, M. (1989) [Hexane levels in olive
dreg extraction plants in Granada, Spain.] Ig. mod., 91: 353-361 (in
Spanish).
MISUMI, J. & NAGANO, M. (1984) Neurophysiological studies on the
relation between the structural properties and neurotoxicity of
aliphatic hydrocarbon compounds in rats. Br. J. ind. Med., 41:
526-532.
MISUMI, J. & NAGANO, M. (1985) Experimental study on the
enhancement of the neurotoxicity of methyl butyl ketone by non-
neurotoxic aliphatic monoketones. Br. J. ind. Med., 42: 155-161.
MIYAGAKI, H. (1967) [Electrophysiological studies of the peripheral
neurotoxicity of n-hexane.] Ind. Health, 9: 660-671 (in Japanese).
MORTELMANS, K., HAWORTH, S., LAWLOR, T., SPECK, W., TAINER, B., &
ZEIGER, E. (1986) Salmonella mutagenicity tests. II. Results from
the testing of 270 chemicals. Environ. Mutagen., 8(Suppl. 7): 1-119.
MUTTI, A., LUCERTINI, S., VENTURI, G., CAVATORTA, A., & FRANCHINI,
I. (1980) Organic solvents and glomerulonephritis. Negative
epidemiological findings for hydrocarbons. Abstracts of the 2nd
International Congress on Toxicology, Brussels. Toxicol. Lett.,
Spec. Issue 1: 220.
MUTTI, A., LUCERTINI, S., FALZOI, M., CAVATORTA, A., & FRANCHINI,
I. (1981) Organic solvents and chronic glomerulonephritis. A cross-
sectional study with negative findings for aliphatic and alicyclic
C5-C7 hydrocarbons. J. appl. Toxicol., 1(4): 224-226.
MUTTI, A., CAVATORTA, A., LUCERTINI, S., ARFINI, G., FALZOI, M., &
FRANCHINI, I. (1982a) Neurophysiological changes in workers exposed
to organic solvents in a shoe factory. Scand. J. Work Environ.
Health, 8(Suppl. 1): 136-141.
MUTTI, A., CAVATORTA, A., LOMMI, G., LOTTA, S., & FRANCHINI, I.
(1982b) Neurophysiological effect of long term exposure to
hydrocarbon mixtures. Arch. Toxicol., Suppl. 5: 120-124.
MUTTI, A., FERRI, F., LOMMI, G., LOTTA, S., LUCERTINI, S., &
FRANCHINI, I. (1982c) n-Hexane induced changes in nerve conduction
velocities and somatosensory evoked potentials. Int. Arch. occup.
environ. Health, 51: 45-54.
MUTTI, A., FALZOI, M., LUCERTINI, S., ARFINI, G., ZIGNANI, M.,
LOMBARDI, S., & FRANCHINI, I. (1984) n-Hexane metabolism in
occupationally exposed workers. Br. J. ind. Med., 41: 533-538.
NAKAJIMA, T. & MURAYAMA, N. (1985) Polyneuropathy caused by
n-hexane used under the commercial name of "benzine". Ind.
Health, 27: 340-341.
NASLUND, B.M.A. & HALPERT, J.S. (1984) Selective inhibition by
chloramphenicol of cytochrome P-450 isozymes in rat lung and liver
involved in the hydroxylation of n-hexane. J. Pharmacol. exp.
Ther., 231(1): 16-22.
NELLIGAN, R.E. (1962) Hydrocarbons in the Los Angeles atmosphere.
Arch. environ. Health, 5: 581-591.
NELSON, K.W., EGE, J.F., ROSS, M., WOODMAN, L.E., & SILVERMAN, L.
(1943) Sensory response to certain industrial solvent vapours. J.
ind. Hyg. Toxicol., 25: 282-285.
NICKELSEN, R.L., RODER, M.M., & BERARDINELLI (1986) Permeation of
chemical protective clothing by three binary solvent mixtures. Am.
Ind. Hyg. Assoc. J., 47: 236-240.
NILSEN, O.G., TOFTGARD, R., ENG, L., & GUSTAFSSON, J. (1981) Regio-
specificity of purified forms of rabbit liver microsomal cytochrome
P-450 in the metabolism of benzo( a)pyrene, n-hexane and
7-ethoxyresorufin. Acta pharmacol. toxicol., 48: 369-376.
NIOSH (1977a) Criteria for a recommended standard: Occupational
exposure to alkanes (C5-C8), Cincinnati, Ohio, National Institute
for Occupational Safety and Health (DHEW (NIOSH) Publication No.
77-151).
NIOSH (1977b) NIOSH manual on analytical methods, 2nd ed.,
Cincinnati, Ohio, National Institute for Occupational Safety and
Health, Vol. 1 (DHEW Publication No. (NIOSH) 77-157-A).
NIOSH (1981a) Health hazard evaluation report, Cincinnati, Ohio,
National Institute for Occupational Safety and Health (HHE-80-040-
860).
NIOSH (1981b) Health hazard evaluation report, Cincinnati, Ohio,
National Institute for Occupational Safety and Health (HHE-81-240-
1031).
NIOSH (1983) Health hazard evaluation report, Cincinnati, Ohio,
National Institute for Occupational Safety and Health (HETA-80-256-
1386).
NIOSH (1984) Manual of analytical methods: Hydrocarbons method
1500, Cincinnati, Ohio, National Institute for Occupational Safety
and Health, pp. 1500/1-1500/7.
NIOSH (1988) Testimony to DOL on the Occupational Safety and Health
Administration's proposed rule on air contaminants, Cincinnati,
Ohio, National Institute for Occupational Safety and Health (Pocket
No. H-020).
NOLTING, F., BEHNKE, W., & ZETZSCH, C. (1988) A smog chamber for
studies of the reactions of terpenes and alkanes with ozone and OH.
J. atmos. Chem., 6: 47-59.
NOMEIR, A.A. & ABOU-DONIA, M.B. (1985) Analysis of n-hexane,
2-hexanone, 2,5-hexanedione, and related chemicals by capillary gas
chromatography and high-performance liquid chromatography. Anal.
Biochem., 151: 381-388.
NOMIYAMA, K. & NOMIYAMA, H. (1974a) Respiratory retention, uptake
and excretion of organic solvents in man. Benzene, toluene,
n-hexane, trichloroethylene, acetone, ethylacetate and ethyl
alcohol. Int. Arch. Arbeitsmed., 32: 75-83.
NOMIYAMA, K. & NOMIYAMA, H. (1974b) Respiratory elimination of
organic solvents in man. Benzene, toluene, n-hexane,
trichloroethylene, acetone, ethylacetate and ethyl alcohol. Int.
Arch. Arbeitsmed., 32: 85-91.
NOMIYAMA, K. & NOMIYAMA, H. (1975) [Concerning the cutaneous
absorption of n-hexane in humans.] Jpn. J. Hyg., 30: 140 (in
Japanese).
NOMIYAMA, K., YOSHIDA, T., & YANAGISAWA, H. (1973) [Percutaneous
absorption of n-hexane caused severe polyneuropathy.] In:
[ Proceedings of the 46th Annual Meeting of the Japanese Association
of Industrial Health], Tokyo, Japanese Association of Industrial
Health, pp. 560-561 (in Japanese).
NOTTEN, W.R.F. & HENDERSON, P.T. (1975a) The influence of n-hexane
treatment on the glucuronic acid pathway and activity of some drug
metabolizing enzymes in guinea pig. Biochem. Pharmacol., 24: 127-131.
NOTTEN, W.R.F. & HENDERSON, P.T. (1975b) Action of n-alkanes on
drug-metabolizing enzymes from guinea-pig liver. Biochem.
Pharmacol., 24: 1093-1097.
NYLEN, P., EBENDAL, T., ERIKSDOTTER-NILSSON, M., HANSSON, T.,
HENSCHEN, A., JOHNSON, A.C., KRONEVI, T., KVIST, U., SJOSTRAND,
N.O., HOGLUND, G., & OLSUN, L. (1989) Testicular atrophy and loss
of nerve growth factor - immunoreactive germ cell line in rats
exposed to n-hexane and a protective effect of simultaneous
exposure to toluene or xylene. Arch. Toxicol., 63: 296-307.
O'DONOGHUE, J.L. (1985) Alkanes, alcohols, ketones and ethylene
oxide. In: O'Donoghue, J.L., ed. Neurotoxicology of industrial and
commercial chemicals, Boca Raton, Florida, CRC Press, Vol. 2, pp.
61-97.
O'DONOGHUE, J.L., KRASAVAGE, W.J., & TERHAAR, C.J. (1978a) A
comparative chronic toxicity study of methyl n-propyl ketone,
methyl n-butyl ketone, and hexane by ingestion, Rochester, New
York, Eastman Kodak Company (Unpublished Report No. 104657Y).
O'DONOGHUE, J.L., KRASAVAGE, W.J., & TERHAAR, C.J. (1978b) Toxic
effects of 2,5-hexanedione. Toxicol. appl. Pharmacol., 45: 269
(abstract 115).
OETTEL, H. (1936) [The influence of organic liquids on skin.] Arch.
exp. Pathol. Pharmacol., 38: 641-696 (in German).
OISHI, H. & MINENO, K. (1964) [Polyneuropathy caused by an organic
solvent ( n-hexane).] Saigai-Igaku, 7(4): 218-222 (in Japanese).
OKAMOTO, S., SHIRBAE, T., KURIMARA, T., & ARAKI, S. (1977) [A case
of n-hexane polyneuropathy occurring in an interior decorator and
experimental n-hexane neuropathy in rats.] Clin. Neurol. (Tokyo),
17: 237-242 (in Japanese).
ONO, Y. & TAKEUCHI, Y. (1980) [Polyneuropathy thought to be
petroleum benzine poisoning occurring in a worker engaged in
cleaning gas meters.] Sangyo Igaku, 22: 256-262 (in Japanese).
ONO, Y., TAKEUCHI, Y., & HISANAGA, N. (1979) [Studies on the method
of measuring nerve conduction velocity in rat's tail and on the
comparative toxicity of n-hexane, methyl n-butyl ketone and
2,5-hexanedione.] Jpn. J. ind. Health, 21: 528-538 (in Japanese
with English summary).
ONO, Y., TAKEUCHI, Y., & HISANAGA, N. (1981) A comparative study on
the toxicity of n-hexane and its isomers on the peripheral nerve.
Int. Arch. occup. environ. Health, 48: 289-294.
ONO, Y., TAKEUCHI, Y., HISANAGA, N., IWATA, M., KITOH, J., &
SUGIURA, Y. (1982) Neurotoxicity of petroleum benzine compared with
n-hexane. Int. Arch. occup. environ. Health, 50: 219-229.
ONOSAKA, S., OCHI, Y., MIN, K.-S., FUJITA, Y., & TANAKA, K. (1988)
[Influences of compounds on metallothionein concentration in mouse
tissues: II. Decrease of pancreatic metallothionein concentration
by n-hexane.] Eisei Kagaku, 34: 446-450 (in Japanese).
OSHA (Occupational Safety and Health Administration) (1989) Air
contaminants: Final rule. Fed. Reg., 54(12): 2921, 2939 (29 CFR Part
1910).
PALAO, A. & LAJO, J.L. (1981) [Toxic polyneuropathy as a
consequence of the use of glue and chemical solvents in the shoe
industry.] Arch. Neurobiol., 44(1): 35-48 (in Spanish).
PARLATO, G., INSERRA, A., & TIMPANARO, V. (1968)
Electrocardiographic changes in experimental poisoning with liquid
hydrocarbons (petrol ether, hexane and heptane). Folia med., 51:
799-805.
PASTERNAK, T., FLOOD, D.G., ESKIN, T.A., & MERIGAN, W.H. (1985).
Selective damage to large cells in the cat retinogeniculate pathway
by 2,5-hexanedione. J. Neurosci., 5: 1641-1652.
PATTY, F.A. & YANT, W.P. (1929) Odour intensity and symptoms
produced by commercial propane, butane, pentane, hexane and heptane
vapour, Washington, DC, US Bureau of Mines, 10 pp (Report No. RI
2979).
PAULSON, G.W. & WAYLONIS, G.W. (1976) Polyneuropathy due to
n-hexane. Arch. intern. Med., 136: 880-882.
PELLIN, M.C., VILANOVA, E., & BARRIL, J. (1988) Low non-neuropathic
tri- o-cresyl phosphate (TOCP) doses inhibit neuropathy target
esterase near the neuropathic threshold in n-hexane pretreated
hens. Toxicology, 49:99-105.
PELLIZZARI, E.D., HARTWELL, T.D., HARRIS, B.S.H., WADDELL, R.D.,
WHITAKER, D.A., & ERICKSON, M.D. (1982) Purgeable organic compounds
in mother's milk. Bull. environ. Contamin. Toxicol., 28: 322-328.
PERBELLINI, L. & BRUGNONE, F. (1980) Identification of the
metabolites of n-hexane, cyclohexane and their isomers in men's
urine. Toxicol. appl. Pharmacol., 53: 220-229.
PERBELLINI, L., BRUGNONE, F., PASTORELLO, G., & GRIGOLINI, L.
(1979) Urinary excretion of n-hexane metabolites in rats and
humans. Int. Arch. occup. environ. Health, 42: 349-354.
PERBELLINI, L., BRUGNONE, F., & GAFFURI, E. (1981a) Neurotoxic
metabolites of "commercial hexane" in the urine of shoe factory
workers. Clin. Toxicol., 18(12): 1377-1385.
PERBELLINI, L., BRUGNONE, F., & FAGGIONATO, G. (1981b) Urinary
excretion of the metabolites of n-hexane and its isomers during
occupational exposure. Br. J. ind. Med., 38: 20-26.
PERBELLINI, L., BRUGNONE, F., SILVESTRI, R., & GAFFURI, E. (1981c)
Measurement of the urinary metabolites of n-hexane, cyclohexane and
their isomers by gas chromatography. Int. Arch. occup. environ.
Health, 48: 99-106.
PERBELLINI, L., BRUGNONE, F., & GAFFURI, E. (1981d) [Occupational
health hazard of technical hexane: study of the biotransformation
products extracted from urine.] Ann. Ist. Sup. Sanità., 17: 615-620
(in Italian).
PERBELLINI, L., AMANTINI, M.C., BRUGNONE, F., & FRONTALI, N.
(1982a) Urinary excretion of n-hexane metabolites. A comparative
study in rat, rabbit and monkey. Arch. Toxicol., 50: 203-215.
PERBELLINI, L., LEONE, R., FRACASSO, M.E., BRUGNONE, F., &
VENTURINI, M.S. (1982b) Metabolic interaction between n-hexane and
toluene in vivo and in vitro. Int. Arch. occup. environ. Health,
50: 351-358.
PERBELLINI, L., BARTOLUCCI, G.B. BRUGNONE, F., DE ROSA, E., &
VALENTINI, F. (1985a) [2,5-Hexanedione in the biological monitoring
of occupational exposure to n-hexane.] Med. Lav., 76(1): 35-43 (in
Italian).
PERBELLINI, L., BRUGNONE, F., CARETTA, D., & MARANELLI, G. (1985b)
Partition coefficients of some industrial aliphatic hydrocarbons
(C5-C7) in blood and human tissues. Br. J. ind. Med., 42: 162-167.
PERBELLINI, L., BRUGNONE, F., MOZZO, P., DE ROSA, E., BARTOLUCCI,
B., & FACINNI, G. (1985c) Toxicokinetic aspects of n-hexane
2,5-hexanedione in the biomonitoring of occupational exposure to
n-hexane. Annu. Conf. Ind. Hyg., 12: 357-364.
PERBELLINI, L., MOZZO, P., BRUGNONE, F., & ZEDDE, A. (1986)
Physiologico-mathematical model for studying human exposure to
organic solvents: kinetics of blood/tissue n-hexane concentrations
and of 2,5-hexanedione in urine. Br. J. ind. Med., 43: 760-768.
PEROCCO, P., BOLOGNESI, S., & ALBERGHINI, W. (1983) Toxic activity
of seventeen industrial solvents and halogenated compounds on human
lymphocytes cultured in vitro. Toxicol. Lett., 16: 69-75.
PETERS, M.A., HUDSON, P.M., & DIXON, R.L. (1981) The effect of
totigestational exposure to methyl n-butyl ketone has on postnatal
development and behaviour. Ecotoxicol. environ. Saf., 5: 291-306.
PLANAS, A.M. & CUNNINGHAM, V.J. (1987) Uncoupling of cerebral
glucose supply and utilization after hexane-2,5-dione intoxication
in the rat. J. Neurochem., 48: 816-823.
PRYOR, G.T. & HOWD, R.A. (1982) Developmental long-term effects of
inhalants in animals, Washington, DC, National Institute of Drug
Abuse (NIDA Quarterly Report No. 5).
PRYOR, G.T., BINGHAM, J., DICKINSON, J., REBERT, C.S., & HOWD, R.A.
(1982) Importance of schedule of exposure to hexane in causing
neurotoxicity. Neurobehav. Toxicol. Teratol., 4: 71-78.
PRYOR, G.T., DICKINSON, J., HOWD, R.A., & REBERT, C.S. (1983)
Neurobehavioural effects of subchronic exposure of weanling rats to
toluene or hexane. Neurobehav. Toxicol. Teratol., 5: 47-52.
RABOVSKY, J. & JUDY, D.J. (1989) The in vitro effects of alkanes,
alcohols, and ketones on rat lung cytochrome P450-dependent
alkoxyphenoxazone dealkylase activities. Arch. Toxicol., 63: 13-17.
RABOVSKY, J., JUDY, D.J., & PAILES, W.H. (1986) In vitro effects
of straight-chain alkanes ( n-hexane through n-dodecane) on rat
liver and lung cytochrome P450. J. Toxicol. environ. Health, 18:
409-421.
RAITTA, C., SEPPALAINEN, A.M., & HUUSKONEN, M.S. (1978) n-Hexane
maculopathy in industrial workers. Albrecht Graefes Arch. klin.
exp. Ophthalmol., 209: 99-110.
RAJE, R.R. (1983) In vitro toxicity of n-hexane and
2,5-hexanedione using isolated perfused rabbit heart. J. Toxicol.
environ. Health, 11: 879-884.
RAJE, R.R., GREENING, M., & FINE, M.T. (1984) Blood n-hexane
concentration following acute inhalation exposure in rats. Res.
Commun. chem. Pathol. Pharmacol., 46(2): 297-300.
RALSTON, W.H., HILDERBRAND, R.L., UDDIN, D.E., ANDERSEN, M.E., &
GARDINER, R.W. (1985) Potentiation of 2,5-hexanedione neurotoxicity
by methyl ethyl ketone. Toxicol. appl. Pharmacol., 81: 319-327.
RANADIVE, K.J., GOTHOSKAR, S.V., & TEZABWALA, B.U. (1972)
Carcinogenicity of contaminants in indigenous edible oils. Int. J.
Cancer, 10: 652-666.
RAPPAPORT, S.M., SELVIN, S., & WATERS, M.A. (1987) Exposures to
hydrocarbon components of gasoline in the petroleum industry. Appl.
ind. Hyg., 2: 148-154.
REBERT, C.S. & SORENSON, S.S. (1983) Concentration-related effects
of hexane on evoked responses from brain and peripheral nerve of
the rat. Neurobehav. Toxicol. Teratol., 5: 69-76.
REBERT, C.S., HOUGHTON, P.W., HOWD, R.A., & PRYOR, G.T. (1982)
Effects of hexane on the brainstem auditory response and caudal
nerve action potential. Neurobehav. Toxicol. Teratol., 4: 79-85.
RIZZUTO, N., TERZIAN, H., & GALIAZZO-RIZZUTO, S. (1977) Toxic
polyneuropathies in Italy due to leather cement poisoning in the
shoe industries. A light and electron microscope study. J. neurol.
Sci., 31: 343-354.
RIZZUTO, N., DECRANDIS, D., DITRAPANI, G., & PASTINATO, E. (1980)
n-hexane neuropathy. An occupational disease of shoemakers. Eur.
Neurol., 19(5): 308-315.
ROBERTSON, P., WHITE, E.L., & BUS, J.S. (1982) Effect of methyl
ethyl ketone treatment on hepatic mixed function oxidase activity
and on in vivo metabolism of n-hexane. Toxicologist, 2(2): A273.
ROBERTSON, P., Jr, WHITE, E.L., & BUS, J.S. (1989) Effects of
methyl ethyl ketone pretreatment on hepatic mixed function oxidase
activity and on in vivo metabolism of n-hexane. Xenobiotica, 19:
721-729.
ROSENBERG, C.K., ANTHONY, D.C., SZAKAL-QUIN, G., GENTER, M.B., &
GRAHAM, D.G. (1987a) Hyperbaric oxygen accelerates the
neurotoxicity of 2,5-hexanedione. Toxicol. appl. Pharmacol., 87:
374-379.
ROSENBERG, C.K., GENTER, M.B., SZAKAL-QUIN, G., ANTHONY, D.C., &
GRAHAM, D.G. (1987b) dl- versus meso-3,4-dimethyl-2,5-hexanedione: a
morphometric study of the proximo-distal distribution of axonal
swellings in the anterior root of the rat. Toxicol. appl.
Pharmacol., 87: 363-373.
RUFF, R.L., PETITO, C.K., & ACHESON, L.S. (1981) Neuropathy
associated with chronic low level exposure to n-hexane. Clin.
Toxicol., 18(5): 515-519.
SABRI, M. (1984) In vitro effects of n-hexane and its metabolites
on selected enzymes in glycolysis, pentose phosphate pathway and
citric acid cycle. Brain Res., 297: 145-150.
SABRI, M., MOORE, C.L., & SPENCER, P.S. (1979) Studies on the
biochemical basis of distal axonopathies. I. Inhibition of
glycolysis by neurotoxic hexacarbon compounds. J. Neurochem., 32:
683-689.
SAHU, S.C., LOWTHER, D.E., & JONES, S.L. (1982) Biochemical
response of rat lung to inhaled n-hexane. Toxicol. Lett., 12: 13-17.
SAIDA, K., MENDELL, J.R., & WEISS, H.S. (1976) Peripheral nerve
changes induced by methyl n-butyl ketone and potentiation by methyl
ethyl ketone. J. Neuropathol. exp. Neurol., 35(3): 207-225.
SANAGI, S., SEKI, Y., SUGIMOTO, K., & HIRATA, M. (1980) Peripheral
nervous system functions of workers exposed to n-hexane at a low
level. Int. Arch. occup. environ. Health, 47: 69-79.
SANDMEYER, E.E. (1981) Aliphatic hydrocarbons. In: Clayton, G.D. &
Clayton, F.E., ed. Patty's industrial hygiene and toxicology, 3rd
ed., New York, Chichester, Brisbane, Toronto, John Wiley and Sons,
Vol. 2B, pp. 3175-3220.
SAX, N.I. (1984) Dangerous properties of industrial materials, 6th
ed., New York, Van Nostrand Reinhold Company.
SAYRE, L.M., SHEARSON, C.M., WONGMONGKOLRIT, T., MEDORI, R., &
GAMBETTI, P. (1986) Structural basis of c-diketone neurotoxicity:
non-neurotoxicity of 3,3-dimethyl-2,5-hexanedione, a c-diketone
incapable of pyrolle formation. Toxicol. appl. Pharmacol., 84: 36-44.
SCELSI, R., POGGI, P., FERA, L., & GONELLA, G. (1980) Toxic
polyneuropathy due to n-hexane. J. neurol. Sci., 47: 7-19.
SCELSI, R., POGGI, P., FERA, L., & GONELLA, G. (1981) Industrial
neuropathy due to n-hexane and 2,5-hexanedione. Clinical and
morphological findings in three cases. Clin. Toxicol., 18: 1387-1393.
SCHAUMBURG, H.H. & SPENCER, P.S. (1976) Degeneration in central and
peripheral nervous systems produced by pure n-hexane: an
experimental study. Brain, 99: 183-192.
SCHAUMBURG, H.H. & SPENCER, P.S. (1979) Clinical and experimental
studies of distal axonopathy. A frequent form of brain and nerve
damage produced by environmental chemical hazards. Ann. NY Acad.
Sci., 329: 14-29.
SCHNOY, N., SCHMIDT, R., ALTENKIRCH, H., & WAGNER, H.M. (1982)
Ultrastructural alteration of alveolar epithelium after exposure
to organic solvents. Respiration, 43: 221-231.
SCHMIDT, R., SCHNOY, N., ALTENKIRCH, H., & WAGNER, H.M. (1984)
Ultrastructural alteration of intrapulmonary nerves after exposure
to organic solvents. Respiration, 46: 362-369.
SELKOE, D.J., LUCKENBILL-EDDS, L., & SHELANSKI, M.L. (1978) Effects
of neurotoxic industrial solvents on cultured neuroblastoma cells:
methyl n-methyl ketone, n-hexane and derivatives. J. Neuropathol.
exp. Neurol., 37: 768-789.
SEPPALAINEN, A. & RAITTA, C. (1981) Neurotoxic properties of
n-hexane among occupationally exposed workers. In: Proceedings of
the 2nd Finnish-Estonian Symposium on Early Effects of Toxic
Substances, Helsinki, June 1981, Helsinki, Institute of
Occupational Health, pp. 180-184.
SEPPALAINEN, A., RAITTA, C., & HUUSKONEN, M. (1979) n-Hexane
induced changes in visual evoked potentials and electroretinograms
of industrial workers. Electroencephalogr. clin. Neurophysiol., 47:
492-498.
SHIRBAE, T., TSUDA, T., TERAO, A., & ARAKI, S. (1974) Toxic
polyneuropathy due to glue sniffing. J. neurol. Sci., 21: 101-113.
SICE, J. (1966) Tumour-promoting activity of n-alkanes and
1-alkanols. Toxicol. appl. Pharmacol., 9: 70-74.
SINGH, D. & JAIN, D.C. (1987) Relative toxicity of various organic
solvents generally used in screening plant products for
insecticidal activity against the house fly Musca domestica L.
Indian J. exp. Biol., 25: 569-570.
SINGH, K.P., KANNAN, K., GOEL, S.K., PANDYA, K.P., & SHANKER, R.
(1983) 2,5-Hexanediol induced thymic atrophy and lymphocytotoxicity
in rats. Ind. Health, 21: 235-242.
SINGH, K.P., UPRETI, R.K., SAXENA, A.K., & SHANKER, R. (1984)
Thymic toxicity produced by 2,5-hexanediol and 2,5-hexanedione in
rats. In: Proceedings of the International Conference on Organic
Solvents Toxicity, Stockholm, October 1984, p. 144 (Abstract 550).
SOBUE, I. & YAMAMURA, Y. (1968) [ n-Hexane polyneuropathy. Outbreak
among vinyl sandal manufacturers.] Rinsho Shinkei, 8: 393-403 (in
Japanese).
SOBUE, I., IIDA, M., YAMAMURA, Y., & TAKAYANAGUI, T. (1978)
n-Hexane polyneuropathy. Int. J. Neurol., 11(4): 317-330.
SPAGNA, C., PARLATO, G., & TIMPANARO, V. (1967) [Behaviour of blood
picture in experimental hexane intoxication.] Folia med. (Napoli),
50: 977-985 (in Italian).
SPENCER, P. (1981) Morphological study of n-hexane intoxication:
Final report, Washington, DC, American Petroleum Institute
(Attachment to API Medical Research Publication No. 28-30077).
SPENCER, P. (1982) Neuropathic potential of n-hexane in the
presence of other hexane isomers, Washington, DC, American
Petroleum Institute (API Medical Research Publication No.
30-30226).
SPENCER, P.S. & SCHAUMBURG, H.H. (1977a) Ultrastructural studies of
the dying-back process. III. The evolution of experimental
peripheral giant axonal degeneration. J. Neuropathol. exp. Neurol.,
36: 276-299.
SPENCER, P.S. & SCHAUMBURG, H.H. (1977b) Ultrastructural studies of
the dying-back process. IV. Differential vulnerability of PNS and
CNS fibres in experimental central-peripheral distal axonopathies.
J. Neuropathol. exp. Neurol., 36: 300-320.
SPENCER, P.S., SCHAUMBURG, H.H., SABRI, M.I., & VERONES, B. (1980)
The enlarging view of hexacarbon neurotoxicity. CRC crit. Rev.
Toxicol., 17(4): 279-356.
STOLTENBURG-DIDINGER, G., WAGNER, H.M., & ALTENKIRCH, H.
(1984) Embryo and fetotoxicity of n-hexane and methyl ethyl ketone.
In: Proceedings of the International Conference on Organic Solvent
Toxicity, Stockholm, October 1984, p. 125 (Abstract 405).
STRATTON, G.W. & SMITH, T.M. (1988) Interaction of organic solvents
with the green alga Chlorella pyrenoidosa. Bull. environ. Contam.
Toxicol., 40: 736-742.
SUZUKI, T., SHIMBO, S., & NISHITANI, H. (1974) Muscular atrophy due
to glue sniffing. Int. Arch. Arbeitsmed., 33: 115-123.
SVABOVA, E., ZORAD, S., MITKOVA, A., KLIMES, I., & MACHO, L. (1987)
[Increase of insulin binding and degradation on isolated human
erythrocytes by n-heptane and n-hexane; effects of trypsin and
dithiotreitol.] Bratisl. lek. Listy., 88(6): 615-624 (in Slovak).
SWANN, H.E., KWON, B.K., HOGAN, G.K., & SNELLINGS, W.M. (1974)
Acute inhalation toxicology of volatile hydrocarbons. Am. Ind. Hyg.
Assoc. J., 35: 511-518.
TAIRA, M. (1975) [Experimental studies on pathomorphological
changes of lung produced by aliphatic and aromatic hydrocarbons and
gasoline.] J. Tokyo med. Univ., 33(4): 597-607 (in Japanese).
TAKAHASHI, M., TAKEUCHI, H., KYO, S., YORIFUJI, S., SANAGI, SL.,
SEKI, Y., & HARA, I. (1977) n-Hexane polyneuropathy: a case report
with review of literature. Med. J. Osaka Univ., 28(1): 77-85.
TAKEUCHI, Y., MABUCHI, C., & TAKAGI, S. (1975) Polyneuropathy
caused by petroleum benzine. Int. Arch. Arbeitsmed., 34: 185-197.
TAKEUCHI, Y., ONO, Y., HISANAGA, N., KITOH, J., & SUGIURA, Y.
(1980) A comparative study on the neurotoxicity of n-pentane,
n-hexane and n-heptane in the rat. Br. J. ind. Med., 37: 241-247.
TAKEUCHI, Y., ONO, Y., & HISANAGA, N. (1981) An experimental study
on the combined effects of n-hexane and toluene on the peripheral
nerve of the rat. Br. J. ind. Med., 38: 14-19.
TAKEUCHI, Y., ONO, Y., HISANAGA, N., IWATA, M., AOYAMA, M., KITOH,
J., & SUGIURA, Y. (1983) An experimental study of the combined
effects of n-hexane and methyl ethyl ketone. Br. J. ind. Med., 40:
199-203.
THAM, R., BUNNFORS, I., ERIKSSON, B., LARSBY, B., LINDGREN, S., &
ODKVIST. L. (1984) Vestibulo-ocular disturbances in rats exposed to
organic solvents. Acta pharmacol. toxicol., 54: 58-63.
TIL, H.P., FALKE, H.E., BEEMS, R.B., BRUIJNTJES, J.P., & DE VOGEL,
N. (1989) Subchronic (90-day) oral toxicity study in rats,
including metaphase chromosomal analysis of bone marrow cells, with
light petroleum solvent (technical hexane) for oil seed extraction:
Final report, Zeist, The Netherlands, TNO-CIVO Institutes, 166 pp
(Project No. B/88-0541) (Report No. V/89.089).
TOFTGARD, R., NILSEN, O.G., & GUSTAFSSON, J.A. (1981) Changes in
rat liver microsomal cytochrome P-450 and enzymatic activities
after the inhalation of n-hexane, xylene, methyl ethyl ketone and
methyl chloroform for four weeks. Scand. J. Work Environ. Health,
7: 31-37.
TOFTGARD, R., NILSEN, O.G., GLAUMANN, H., & GUSTAFSSON, J.A. (1983)
Induction of cytochrome P-450 in the rat liver after exposure to
xylenes. Dose response relationship and dependence on endocrine
factors. Toxicology, 27: 119-137.
TOFTGARD, R., HAAPARANTA, T., ENG, L., & HALPERT, J. (1984) Rat
lung and liver microsomal cytochrome P-450 isozymes involved in the
hydroxylation of n-hexane and m-xylene. In: Abstracts of the
International Conference on Organic Solvent Toxicity, Stockholm,
October 1984, p. 17 (Abstract 111).
TOFTGARD, R., HAAPARANTA, T., ENG, L., & HALPERT, J. (1986) Rat
lung and liver microsomal cytochrome P-450 isozymes involved in the
hydroxylation of n-hexane. Biochem. Pharmacol., 35: 3733-3738.
TOWFIGHI, J., NICHOLAS, K., GONATAS, M.D., PLEASURE, D., COOPER,
H.S., & MCCREE, L. (1976) Glue sniffer's neuropathy. Neurology, 26:
238-243.
TOXIGENICS (1982) 90-Day vapour inhalation toxicity study on
n-hexane in albino rats (Report submitted to Chemical Industry
Institute of Toxicology, North Carolina) (CIIT Pocket No. 10242).
TSURUTA, H. (1977) Percutaneous absorption of organic solvents. II.
A method for measuring the penetration rate of chlorinated solvents
through excised rat skin. Ind. Health, 15: 131-139.
TSURUTA, H. (1980) Microdetermination of n-hexane in biological
samples by gas chromatography - mass spectroscopy. Ind. Health, 18:
113-114.
TSURUTA, H. (1982) Percutaneous absorption of organic solvents.
III. On the penetration rates of hydrophobic solvents through the
excised rat skin. Ind. Health, 20: 335-345.
UPRETI, R.K., SINGH, K.P., SAXENA, A.K., & SHANKER, R. (1986)
Effect of 2,5-hexanedione on lymphoid organs in rats: a preliminary
report. Environ. Res., 39: 188-198.
USITC (1988) Synthetic organic chemicals. United States Production
and Sales 1987, Washington, DC, United States International Trade
Commission, pp. 2-7 (USITC Publication No. 2009).
VAINIO, H. (1974) Activation and inactivation of membrane-bound
UDP-glucuronosyltransferase by organic solvents in vitro. Acta
pharmacol. toxicol., 34: 152-156.
VDI (Verband Deutscher Ingenieure) (1984) Handbook of clean air,
Berlin, Benth Verlag Gmbh, Vol. 5 (Guideline 3482).
VERONESI, B., PETERSON, E.R., DI VINCENCO, G., & SPENCER, P.S.
(1978) A tissue culture model of distal (dying-back) axonopathy:
its use in determining primary neurotoxic hexacarbon compounds. J.
Neuropathol. exp. Neurol., 33: 703.
VERONESI, B., PETERSON, E.R., & SPENCER, P.S. (1980) Reproduction
and analysis of MnBK neurotoxicity in organotypic tissue culture.
In: Spencer, P.S. & Schaumburg, H.H., ed. Experimental and clinical
neurotoxicology, Baltimore, Maryland, Williams and Wilkins, pp.
863-871.
VERONESI, B., PETERSON, E.R., BORNSTEIN, M., & SPENCER, P.S. (1983)
Ultrastructural studies of the dying-back process. VI. Examination
of nerve fibres undergoing giant axonal degeneration in organotypic
culture. J. Neuropathol. exp. Neurol., 24(2): 153-165.
VERONESI, B., LINGTON, A.W., & SPENCER, P.S. (1984) A tissue
culture model of methyl ethyl ketone's potentiation of n-hexane
neurotoxicity. Neurotoxicology, 5(2): 43-52.
VEULEMANS, H., VAN VLEM, E., JANSSENS, H., MASSCHELEIN, R., &
LEPLAT, A. (1982) Experimental human exposure to n-hexane. Study of
the respiratory uptake and elimination, and of n-hexane
concentrations in peripheral venous blood. Int. Arch. occup.
environ. Health, 49: 251-263.
VEULEMANS, H., GROESENEKEN, D., MASSCHELEIN, R., & VLEM, E.V.
(1987) Survey of ethylene glycol ether exposures in Belgian
industries and workshops. Am. Ind. Hyg. Assoc. J., 48: 671-676.
VICEDO, J.L., PELLIN, M., & VILLANOVA, E. (1985) Phthalates and
organophosphorus compounds as cholinesterase inhibitors in
fractions of industrial hexane impurities. Arch. Toxicol., 57: 46-52.
VILANOVA, E. & VICEDO, J.L. (1983) Serum cholinesterase inhibitors
in the commercial hexane impurities. Arch. Toxicol., 53: 59-69.
WADA, Y., OKAMOTO, S., & TAKAGI, S. (1965) [Intoxication
polyneuropathy following exposure to n-hexane.] Rinsho Shinkeigaku,
10(5): 591-597 (in Japanese).
WAGNER, H.M., STEPPART, R., & ALTENKIRCH, H. (1984) [Animal
experiments on the neurotoxicity of organic solvents in rats
challenged with heavy metals.] Schriftenr. Ver. Wasser Boden
Lufthyg., 59: 191-200 (in German).
WAHLBERG, J.E. (1984a) Erythema-inducing effects of solvents
following epicutaneous administration to man: studied by laser
Doppler flowmetry. Scand. J. Work Environ. Health, 10: 159-162.
WAHLBERG, J.E. (1984b) Edema inducing effects of solvents following
topical administration. Derm. Beruf Umwelt, 32(3): 91-94.
WAHLBERG, J.E. & BOMAN, A. (1979) Comparative percutaneous toxicity
of ten industrial solvents in the guinea-pig. Scand. J. Work
Environ. Health, 5: 345-351.
WALSETH, F. & NILSEN, O.G. (1984) Phthalate esters. II. Effects of
inhaled dibutylphthalate on cytochrome P-450 mediated metabolism in
rat liver and lung. Arch. Toxicol., 55: 132-136.
WALSETH, F., TOFTGARD, R., & NILSEN, O.G. (1982) Phthalate esters.
I. Effects on cytochrome P-450 mediated metabolism in rat liver and
lung, serum enzymatic activities and serum protein levels. Arch.
Toxicol., 50: 1-10.
WANG, J.D., CHANG, Y.C., KAO, K.P., HUANG, C.C., LIN, C.C., & YEH,
W.Y. (1986) An outbreak of n-hexane induced polyneuropathy among
press proofing workers in Taipei. Am. J. ind. Med., 10: 111-118.
WEAST, R.C. (1981) CRC handbook of chemistry and physics, 6th ed.,
Boca Raton, Florida, CRC Press.
WHITE, E.L. & BUS, J.S. (1980) Determination of n-hexane, methyl
ethyl ketone and their metabolites in sciatic nerves of rats by gas
chromatography-mass spectrometry. In: Abstracts of the Pittsburgh
Conference on Analytical Chemistry and Applied Spectroscopy, 10-14
March, 1980, Pittsburgh, Pennsylvania, US Society of Analytical
Chemistry (Abstract No. 525).
WHITE, E.L., BUS, J.S., & HECK, H. (1979) Simultaneous
determination of n-hexane, 2-hexanone and 2,5-hexanedione in
biological tissues by gas chromatography mass spectrometry. Biomed.
mass Spectrom. 6(4): 169-172.
WHO (1971) Evaluation of food additives. Fourteenth Report of the
Joint FAO/WHO Expert Committee on Food Additives, Geneva, World
Health Organization, pp. 21-24 (Technical Report Series No. 462).
YAMADA, N. (1964) [Polyneuritis in workers handling n-hexane.]
Sangyo Igaku, 6: 192 (in Japanese).
YAMADA, S. (1967) [Intoxication polyneuritis in workers exposed to
n-hexane.] Jpn. J. ind. Health, 9: 651-659 (in Japanese).
YAMAKAGE, A. & ISHIKAWA, H. (1982) Generalised morphea-like
scleroderma occurring in people exposed to organic solvents.
Dermatologia, 165: 186-193.
YAMAMURA, Y. (1969) n-Hexane polyneuropathy. Folia psychiatr.
neurol. Jpn., 23(1): 45-57.
YOSHIDA, T., YANAGISAWA, H., MUNEYUKI, T., & SHIGIYA, R. (1974) [An
electrophysiological study of n-hexane polyneuropathy.] Rinsho
Shinkeigaku, 14(5): 454-461 (in Japanese).
ZAHLSEN, K., AARSTAD, K., & NILSEN, O.G. (1985) Inhalation of
isopropanol. Induction of activating and deactivating enzymes in
rat kidney and liver. Increased microsomal metabolism of n-hexane.
Toxicology, 34: 57-66.
RESUME
Le n-hexane (hexane normal) est un liquide volatil incolore.
L'hexane du commerce se présente essentiellement sous la forme d'un
mélange de divers isomères de l'hexane et de dérivés apparentés à
six atomes de carbone; sa teneur en n-hexane varie entre 20 et 80%.
La chromatographie en phase gazeuse avec détection par ionisation
de flamme ou couplage à un spectrographe de masse constitue une
bonne technique pour le dosage du n-hexane. Les limites
d'exposition professionnelle varient de 100 à 1800 mg/m3 (moyenne
pondérée par rapport au temps, TWA) et de 400 à 1500 mg/m3 (valeur
plafond, CLV) selon les pays.
On peut isoler le n-hexane du gaz naturel et du pétrole brut.
On l'utilise dans les industries de transformation alimentaire,
notamment pour l'extraction des huiles végétales ou comme solvant
dans différents produits et procédés.
Une fois libéré dans l'environnement, le n-hexane y subsiste
essentiellement en phase vapeur. Sa demi-vie dans l'atmosphère est
estimée à environ deux jours, si l'on se base uniquement sur sa
réactivité vis-à-vis du radical OH. Les valeurs de la CL50 pour les
organismes aquatiques sont rares et dispersées; elles ont été
obtenues dans des conditions peu satisfaisantes. Dans ces
conditions il n'est pas possible d'apprécier les effets toxiques du
n-hexane dans l'environnement. Du fait de sa faible solubilité
dans l'eau et de sa forte volatilité il est peu probable que les
organismes aquatiques courent un risque d'exposition, sauf en cas
de décharge sauvage dans les eaux superficielles.
Chez les mammifères, le n-hexane est rapidement résorbé au
niveau des poumons et se distribue largement dans l'organisme de
l'adulte ainsi que dans les tissus du foetus. Au niveau cutané, la
résorption est faible. Le n-hexane subit une métabolisation
oxydative en un certain nombre de dérivés, notamment l'hexane-2,5-
dione dont on pense qu'elle constitue l'agent neurotoxique final.
Dans le nerf sciatique de rats d'expérience on a pu trouver des
concentrations particulièrement élevées de n-hexane et d'hexane-
2,5-dione. La majeure partie du n-hexane est excrétée sans
modification dans l'air exhalé; une certaine quantité est également
excrétée sous forme de métabolites dans l'air expiré et dans les
urines.
Le n-hexane présente une faible toxicité aiguë pour le rat
adulte après administration par voie orale ou inhalation. On a noté
des valeurs de la DL50 par voie orale allant de 15 à 30 g/kg; quant
à la CL50 par inhalation, elle serait de 271 040 mg/m3 (77 000 ppm)
pour une exposition d'une heure. Lorsque la teneur en vapeurs est
élevée, on constate chez l'animal de l'ataxie, des convulsions et
les signes d'une dépression du système nerveux central.
Il semble que chez le rat, les principaux effets d'une
exposition répétée au n-hexane consistent en lésions testiculaires
et signes de neurotoxicité. C'est ainsi que l'inhalation de
n-hexane et l'ingestion d'hexane-2,5-dione ont produit de graves
lésions testiculaires chez les animaux d'expérience. Ces effets ont
été attribués à la désagrégation du cytosquelette des cellules de
Sertoli. On note des effets secondaires sur les cellules germinales
des stades postérieurs aux spermatogonies, qui disparaissent des
tubules endommagés. Après une seule exposition de 24 heures à
17 600 mg/m3 de n-hexane (5000 ppm) les effets testiculaires
étaient réversibles mais ils devenaient irréversibles après deux
semaines d'exposition à la même concentration, 16 heures par jour,
six jours par semaine. A la concentration de 1% dans l'eau de
boisson, l'hexane-2,5-dione a produit des lésions testiculaires
réversibles du même type au bout de deux à trois semaines
d'administration et des effets irréversibles (en l'espace de 17
semaines) au bout de cinq semaines d'administration.
L'effet neurotoxique se caractérise cliniquement par une
faiblesse des pattes arrières qui peut évoluer vers la paralysie.
Il se produit un gonflement de l'axone aux niveaux central et
périphérique avec parfois des lésions plus sévères (dégénérescence
et disparition de l'axone), notamment au niveau des troncs nerveux
de plus grande longueur et de plus gros diamètre. Lors d'études
comportant principalement l'inhalation d'hexane pendant six mois,
on a noté la présence de lésions du système nerveux central et
périphérique aux doses de 1760 mg/m3 (500 ppm) ou davantage; en
revanche il n'y avait pas d'effets cliniques ou
anatomopathologiques à la dose de 400 mg/m3 (125 ppm). Quinze à 22
semaines après l'arrêt de l'exposition, cinq jours par semaine
pendant 11 semaines à des vapeurs de n-hexane à la concentration de
3520 mg/m3 (1000 ppm), on a noté que l'amplitude des potentiels
évoqués auditifs du tronc cérébral (PEAP) n'était que partiellement
restaurée. L'exposition discontinue de rats à une concentration de
3168 mg/m3 (900 ppm) pendant 72 semaines n'a pas provoqué de
lésions apparentes aux niveaux périphérique ou central mais
certains effets électrophysiologiques étaient perceptibles au
niveau des nerfs périphériques.
La neurotoxicité induite par le n-hexane peut être
potentialisée en présence de méthyléthylcétone, de
méthylisobutylcétone et d'acétate de plomb et réduite par
exposition simultanée au toluène. La perturbation des
concentrations de dopamine est également synergisée par une
exposition simultanée au toluène et au n-hexane.
Des lésions microscopiques graves ont été observées sur la peau
après application de n-hexane et pose d'un pansement occlusif
après une brève période. L'exposition prolongée à des vapeurs de
n-hexane à une concentration de 1560 mg/m3 (3000 ppm) peut
provoquer chez le rat une irritation des conjonctives et une forte
irritation oculaire chez le lapin. On ne dispose d'aucune donnée
sur la sensibilisation cutanée provenant d'études sur l'animal.
Des lésions chromosomiques (polyploïdie dans le cas d'une
étude, et aberrations de structure dans le cas d'une autre) ont été
signalées tant in vitro qu' in vivo. On n'a pas observé
d'augmentation de la fréquence des mutations ponctuelles ni de
signes de lésion de l'ADN.
Une étude de cancérogénicité a été effectuée sur des souris
(badigeonnage de la peau avec du n-hexane): elle n'a révélé aucun
signe de cancérogénicité.
Les effets du n-hexane sur la fonction de reproduction n'ont
pas été suffisamment étudiés. Aucun signe important
d'embryotoxicité ou de tératogénicité n'a été observé chez des rats
après inhalation de n-hexane à des concentrations relativement
basses ni chez des souris après administration par voie orale. On a
constaté un retard transitoire du développement postnatal chez des
ratons après exposition des femelles gravides à des vapeurs de
n-hexane à des concentrations de 3520 mg/m3 (1000 ppm).
On ne possède guère de données sur la toxicité aiguë du
n-hexane pour l'être humain. La plupart des études portent sur
l'exposition professionnelle à des mélanges de solvants. D'après
les données disponibles, il semble que le n-hexane n'ait qu'une
faible toxicité aiguë. On a signalé des signes de dépression du
système nerveux central, tels que somnolence, vertiges, sensation
ébrieuse, après exposition à de l'hexane du commerce pendant 10 à
60 minutes à des concentrations allant de 3520 à 17 600 mg/m3
(1000 - 5000 ppm).
Le n-hexane est légèrement irritant et provoque un érythème
passager chez l'homme après un court contact cutané. Des effets
plus graves (érythème et phlyctènes) ont été enregistrés après un
contact de cinq heures avec du n-hexane de qualité commerciale, la
zone de contact étant recouverte. Aucun cas de sensibilisation
cutanée n'a été signalé chez les ouvriers exposés et un test de
sensibilisation cutanée maximale effectué avec du n-hexane a donné
des résultats négatifs.
En cas d'exposition répétée, le n-hexane entraîne une
polynévrite sensitivo-motrice périphérique. De nombreuses études
sur la prévalence des effets neurotoxiques dus au n-hexane ont été
publiées; toutefois on manque bien souvent de données
satisfaisantes sur l'exposition. Des cas de neuropathie ont été
attribués à l'exposition à des concentrations atmosphériques de
n-hexane allant de 106 à 8800 mg/m3 (30 - 2500 ppm). Des cas de
neuropathie périphérique marquée ont été signalés parmi des
ouvriers japonais travaillant à la fabrication de sandales ainsi
que chez des opérateurs taïwanais de presse à épreuves exposés à
des teneurs en n-hexane d'environ 76 et 352 mg/m3 (50 et 100 ppm),
respectivement, pendant des périodes supérieures à huit heures par
jour. Dans la plupart des cas, les mesures d'exposition étaient
récentes et ne reflétaient probablement pas très exactement les
expositions antérieures susceptibles d'être à l'origine de la
neuropathie en question.
Plusieurs études transversales ont fait état, indépendamment
les unes des autres, d'effets infracliniques légers (par exemple
des altérations électrophysiologiques au niveau des nerfs
périphériques) chez des travailleurs exposés à des concentrations
de 70 - 352 mg/m3 (20 - 100 ppm). Aucun cas bien défini de
neurophathie périphérique cliniquement manifeste n'a pu être relevé
dans l'une quelconque de ces études à des niveaux d'exposition
inférieurs à 352 mg/m3 (100 ppm).
Seuls quelques travaux ont été consacrés à l'étude des effets
du n-hexane sur le système nerveux central. On pense que les
modifications observées dans les potentiels évoqués
somatosensoriels d'ouvriers exposés à du n-hexane proviennent d'un
bloc de la conduction nerveuse centrale. On a également noté une
altération des potentiels évoqués visuels et du tracé électro-
encéphalographique. Ces résultats incitent à penser que le
n-hexane pourrait entraîner des troubles du système nerveux
central, mais les données disponibles ne donnent aucune indication
sur les niveaux d'exposition en cause.
EVALUATION DES RISQUES POUR LA SANTE HUMAINE ET DES EFFETS SUR
L'ENVIRONNEMENT
1. Etudes sur l'animal
1.1 Exposition à court et à long terme
Le n-hexane présente une faible toxicité aiguë puisque l'on
fait état de valeurs de la DL50 comprises entre 15 et 30 g/kg et
d'une valeur de la CL50 à une heure de 271 040 mg.
Lors d'études consistant essentiellement à faire inhaler du
n-hexane à des rats de manière continue, on a observé une
mortalité chez ces animaux après exposition à une concentration de
3520 mg/m3 (1000 ppm) pendant des durées allant jusqu'à 11
semaines, ainsi qu'une diminution du gain de poids après trois
semaines d'exposition à 1760 mg/m3 (500 ppm). Lors d'études
comportant des expositions quotidiennes de 12 heures à du
n-hexane, on a enregistré une mortalité chez les animaux à la
concentration de 10 560 mg/m3 (3000 ppm) au bout de 15 semaines et
une légère réduction du gain de poids corporel à la concentration
de 740 mg/m3 (200 ppm) au bout de 24 semaines. Aucun effet sur le
poids corporel des rats n'a été observé après une exposition
pratiquement continue à 443 mg/m3 (126 ppm) pendant 34 semaines.
Il semble que les lésions testiculaires et les effets
neurotoxiques aux niveaux périphérique et central soient les
principaux effets non mortels d'une exposition répétée au n-hexane
chez le rat. Après inhalation de n-hexane et ingestion de hexane-
2,5-dione on a observé de graves lésions testiculaires. Ces effets
ont été attribués à la désagrégation du cytosquelette des cellules
de Sertoli. On observe également des effets secondaires sur les
cellules germinales des stades qui suivent la spermatogonie et qui
disparaissent des tubules endommagés. Après une seule exposition de
24 heures à 17 600 mg/m3 (5000 ppm), les effets testiculaires
étaient réversibles mais ils devenaient irréversibles après deux
semaines d'exposition à la même concentration 16 heures par jour,
six jours par semaine. De l'hexane-2,5-dione administrée dans l'eau
de boisson à la concentration de 1% a produit des lésions
testiculaires réversibles au bout de deux à trois semaines
d'administration et des effets irréversibles (en l'espace de 17
semaines) au bout de cinq semaines d'administration.
Du point de vue clinique, la neurotoxicité se caractérise par
une faiblesse des pattes arrières, qui peut évoluer vers la
paralysie et aboutir à une quadriplégie. Ces signes cliniques ont
été observés après dix semaines d'exposition discontinue à 10 560
mg/m3 (3000 ppm) (12 heures par jour et sept jours par semaine) et
au bout de six à dix semaines d'exposition continue à des doses
allant de 1048 - 2112 mg/m3 (400 à 600 ppm).
Avant que ne se manifeste la faiblesse des pattes arrières on a
observé des lésions (gonflement de l'axone) au niveau des nerfs des
pattes arrières, du cerveau et de la moëlle épinière. Des effets
plus graves (dégénérescence et disparition de l'axone) ont été
notés en particulier au niveau des troncs nerveux de plus grande
longueur et plus grand diamètre au bout de six mois d'exposition à
1760 mg/m3 (500 ppm). Au total, il est difficile d'établir la
valeur de la dose sans effet nocif observable dans le cas des
effets neurotoxiques provoqués par le n-hexane chez le rat, en
raison de la diversité des concentrations et des modalités
d'exposition expérimentées ainsi que de la variété des effets
étudiés. Lors d'études de six mois consistant essentiellement à
faire inhaler en continu des vapeurs de n-hexane, on a observé des
lésions nerveuses à des concentrations de 1760 mg/m3 (500 ppm ou
davantage), toutefois aucun effet clinique ni anatomopathologique
n'a été constaté à la dose de 440 mg/m3 (125 ppm). En mode
discontinu l'exposition de rats à la dose de 3168 mg/m3 (900 ppm)
pendant 72 semaines n'a pas provoqué de lésions nerveuses
évidentes, mais on a pu constater les premiers signes d'effets sur
la force de préhension.
Après arrêt de l'exposition il peut y avoir résolution
(partielle ou complète) des signes cliniques de toxicité, de
l'amoindrissement de la force de préhension et des effets
électrophysiologiques. Dans les 15 à 22 semaines qui suivent
l'arrêt d'une exposition répétée à la concentration de 3520 mg/m3
(1000 ppm) de n-hexane pendant 11 semaines, on constate que
l'amplitude de l'onde cinq des potentiels évoqués auditifs du tronc
cérébral (qui est supposée refléter l'activité du système nerveux
central) ainsi que le potentiel d'action du nerf caudal, ne sont
pas complètement rétablis. On n'a pas encore étudié
systématiquement la résolution des lésions anatomopathologiques
consécutives à une exposition par inhalation, mais on a toutefois
observé que lorsqu'on cesse d'exposer les animaux à des
concentrations allant de 1408 à 2112 mg/m3 (400 à 600 ppm), il y a
remyélinisation et régénération limitées des axones.
L'inhalation en discontinu pendant sept semaines de n-hexane à
la concentration de 1760 mg/m3 (500 ppm) peut provoquer de petites
lésions au niveau des poumons chez le rat. Chez le lapin, on a
observé de graves lésions de l'arbre respiratoire après exposition
à une concentration de 10 560 mg/m3 (3000 ppm), huit heures par
jour pendant huit jours, et on a noté une prolifération papillaire
des cellules broncholiaires non ciliées au bout de 24 semaines
d'exposition.
On a fait état d'effets sur le rein et le foie à la suite de
quelques études d'inhalation sur des rats mais seulement aux doses
les plus fortes. Quant à la plupart des autres lésions et des
autres effets, on a estimé qu'ils étaient d'une importance
toxicologique mineure ou constaté qu'ils n'étaient pas
reproductibles. Cependant, on a observé de graves lésions
hépatiques chez des rats après administration intrapéritonéale
répétée de n-hexane.
Des études portant sur l'exposition à plusieurs solvants ont
montré que les effets neurotoxiques de la vapeur de n-hexane
pouvaient être accrus chez des poules par la présence de méthyl-
isobutylcétone et chez des rats par la présence simultanée de
méthyl-isobutylcétone et d'acétate de plomb; toutefois l'effet
était au contraire réduit en présence de toluène. On a également
indiqué que les lésions pulmonaires observées chez les rats
augmentaient en cas d'exposition simultanée à la méthyléthylcétone
et à l'hexane.
Peu d'études ont été consacrées aux effets de l'ingestion de
n-hexane ou à ceux de l'exposition cutanée à cette substance.
L'administration quotidienne par voie orale de 500 mg/kg de
n-hexane pendant quatre semaines à des rats a entraîné une baisse
de la vitesse de conduction du nerf caudal.
1.2 Génotoxicité
Le n-hexane a fait l'objet de tests de mutagénicité limités.
On a fait état de lésions chromosomiques (polyploïdie dans le cas
d'une étude et aberrations structurales dans le cas d'une autre)
lors d'essais in vitro au cours desquels une lignée cellulaire
provenant de hamsters chinois a été exposée à du n-hexane liquide
non dilué en l'absence de fraction métabolisante. Dans le cas de
cellules mammaliennes en culture ou de bactéries, on n'a pas
observé d'augmentation significative du taux de mutations
ponctuelles. L'exposition au n-hexane n'augmente pas les lésions
de l'ADN comme on a pu en juger par des mesures de toxicité
bactérienne différentielle. De même il n'y a pas d'augmentation de
la synthèse non programmée d'ADN dans les lymphocytes humains in
vitro. Bien que les tests portant sur les mutations ponctuelles
aient donné des résultats négatifs, il semble, à certains signes,
que le n-hexane soit capable de produire des aberrations
chromosomiques in vitro. Des lésions chromosomiques (rupture des
chromatides) ont été signalées à l'occasion d'une étude in vivo
sur des cellules de moëlle osseuse de rat.
1.3 Cancérogénicité
Le pouvoir cancérogène du n-hexane n'a pas été suffisamment
étudié. Une étude de portée limitée a montré que l'application
cutanée de n-hexane ne semblait pas accroître la formation de
tumeurs cutanées chez la souris après exposition à un initiateur
tumoral (diméthyl-7,12 benzanthracène) ou après exposition
simultanée à un agent promoteur (huile de croton).
1.4 Effets sur la reproduction
Les effets toxiques du n-hexane sur la fonction de
reproduction n'ont pas été convenablement étudiés. Il n'existe pas
de preuves substantielles que le n-hexane entraîne des effets
embryotoxiques ou tératogènes après inhalation chez le rat ou
ingestion chez la souris. Chez des rats exposés à une dose de
10 560 mg/m3 (3000 ppm), on a observé une légère augmentation de
l'incidence de certaines anomalies, toutefois en raison
d'insuffisances dans la manière dont l'étude a été conduite (par
exemple concentration probablement trop faible) les conclusions
qu'on peut en tirer demeurent limitées. Des signes de toxicité
foetale (réduction du poids des souriceaux) ont été observés mais
seulement à des doses qui entraînaient une mortalité importante
chez les femelles gravides. Une des études effectuées a mis en
évidence un retard passager dans le développement postnatal de
ratons exposés à la concentration de 10 560 mg/m3 (3000 ppm). Aucun
signe de neuropathie n'a été signalé chez ces animaux mais on n'est
pas en mesure, sur la base des résultats de cette étude, de se
prononcer sur la possibilité que l'exposition in utero au
n-hexane induise effectivement une neuropathie, car les animaux
n'ont été exposés à cette substance que durant une courte fraction
de la période de gestation. Selon d'autres rapports, l'exposition
in utero au n-hexane retarderait la genèse du tissu cérébelleux
ainsi que la maturation des neurones et produirait des anomalies
électrophysiologiques au niveau central chez les rats nouveaux-nés.
Cependant on ne possède pas suffisamment de détails sur ces
travaux pour en apprécier toute la portée.
On est fondé à penser que le n-hexane entraîne une stérilité
irréversible chez le rat. Une étude limitée de dominance létale
chez la souris n'a pas révélé d'effets sur la fertilité des mâles.
Aucune étude n'a été publiée sur la fertilité des femelles de
divers animaux ou sur les effets portant sur plusieurs générations.
2. Etudes sur l'homme
2.1 Effets sur le système nerveux
Selon de nombreuses études un certain nombre de cas humains de
neuropathie périphérique seraient imputables à une exposition
professionnelle au n-hexane. Toutefois les concentrations
atmosphériques sont rarement mentionnées et quand elles le sont,
elles peuvent très bien être inexactes, sans compter qu'il n'est
pas toujours facile de savoir s'il s'agit d'hexane ou de n-hexane.
En outre, l'exposition au n-hexane en milieu industriel se produit
généralement lors de l'utilisation de produits contenant divers
solvants, de sorte que les ouvriers sont exposés en fait à un
mélange de composés volatils (par exemple de l'acétate d'éthyle, de
la méthyléthylcétone, du toluène, de l'acétone et des hydrocarbures
aliphatiques autres que le n-hexane). La concentration
atmosphérique des autres solvants présents est rarement précisée.
Dans la présente mise au point, on ne prend en considération que
celles des études qui donnent les concentrations atmosphériques.
Les résultats de l'étude électrophysiologique des nerfs
périphériques peuvent varier considérablement selon la technique
utilisée, la température du laboratoire dans lequel les mesures
sont effectuées, la portion du nerf étudiée et l'âge du sujet.
Aussi est-il difficile d'apprécier la portée des études qui
reposent sur ces méthodes, en l'absence de renseignements sur tous
ces facteurs.
L'étude de 93 cas de neuropathie a permis de caractériser le
tableau clinique de la neuropathie due au n-hexane et a montré que
les données électrophysiologiques étaient en bonne corrélation avec
la gravité de la maladie. On a en particulier constaté que plus
l'exposition était importante plus la maladie était grave.
Toutefois il n'a pas été possible d'établir une relation bien
définie à partir des données limitées dont on disposait sur
l'exposition effective.
On a observé de légers signes infracliniques de neuropathie
chez des ouvriers d'une fabrique de chaussures exposés à une
concentration de 70 mg/m3 (20 ppm). L'importance des anomalies
électrophysiologiques augmentait avec la durée de l'exposition.
Toutefois aucun cas de neuropathie déclarée n'a été observé même
chez les ouvriers qui avaient été exposés pendant plus de cinq ans.
On a observé des signes subjectifs et électrophysiologiques de
neuropathie périphérique infraclinique dans un groupe d'ouvriers
exposés à une dose d'environ 317 mg/m3 (90 ppm). Toutefois, on ne
peut pas attribuer entièrement ces effets à l'exposition au
n-hexane étant donné que l'on a probablement sous-estimé les
expositions antérieures à divers solvants et que les ouvriers
avaient probablement introduit eux-mêmes un biais sélectif. En
outre, il peut y avoir eu exposition simultanée à de la
méthyléthylcétone, dont on sait qu'elle accroît les effets
neurotoxiques du n-hexane.
L'exposition à un mélange de solvants contenant du n-hexane à
la concentration d'environ 690 mg/m3 (196 ppm) a entraîné une
réduction de la vitesse de conduction nerveuse périphérique,
anomalie qui s'est maintenue même après une réduction importante de
l'exposition. En outre, on pense que les variations observées dans
les potentiels évoqués somatosensoriels s'expliquent par l'effet du
n-hexane sur le système nerveux central.
Des signes subjectifs et électrophysiologiques d'une
neuropathie infraclinique légère ont été observés dans un petit
groupe de travailleurs exposés à une concentration moyenne de
n-hexane dans l'atmosphère de 208 mg/m3 (59 ppm). Aucun signe
clinique de neuropathie n'était perceptible. L'exposition à
d'autres solvants était limitée et on a pu procéder à des études de
conduction nerveuse dans des conditions bien définies. Toutefois,
le faible effectif des ouvriers, l'absence de données
rétrospectives sur l'exposition et le fait que les groupes témoins
n'étaient pas très bien appariés au groupe exposé limitent la
portée des conclusions qu'on peut en tirer. Néanmoins, les
résultats sont en bon accord général avec les données fournies par
les études antérieures dont il a été question plus haut. On a
attribué le déclenchement d'une neuropathie périphérique
cliniquement manifeste à l'exposition pendant des périodes
supérieures à huit heures par jour à une concentration de n-hexane
de 669 mg/m3 (190 ppm). Au bout d'un certain nombre d'années, on
assiste à une lente restauration - encore que partielle - de la
fonction nerveuse. L'exposition à une concentration inférieure à
352 mg/m3 (100 ppm) n'a pas entraîné de neuropathie mais l'étude
électrophysiologique a révélé la présence d'effets infracliniques.
Chez un petit nombre de travailleurs exposés de par leur
profession à toute une gamme de concentrations atmosphériques de
n-hexane, on a observé une perturbation de la perception des
couleurs et des signes de maculopathie. La mesure des potentiels
évoqués visuels et l'électrorétinogramme ont révélé chez ces sujets
la présence d'anomalies par comparaison avec un groupe témoin non
exposé. On a également signalé l'existence d'anomalies dans les
potentiels évoqués visuels chez des renifleurs de colle exposés à
du n-hexane. On pense qu'il pourrait se produire une dégénérescence
axonale au niveau des voies optiques et un bloc de conduction au
niveau central. On est également fondé à croire que les signes de
maculopathie pourraient être corrélés avec les signes
électrophysiologiques de neuropathie périphérique. Certains des
sujets étudiés présentaient également un électroencéphalogramme
anormal.
2.2 Autres effets
Aucune étude n'a été publiée au sujet d'effets éventuels sur
les testicules et la fertilité des mâles. On ne dispose que de très
peu d'informations sur la toxicité aiguë du n-hexane pour l'homme.
La plupart des études toxicologiques portaient sur l'exposition à
des mélanges de solvants.
A la lumière des données disponibles, il semble que le
n-hexane n'ait qu'une faible toxicité aiguë.
Très peu d'études portant sur des volontaires ont été publiées
à propos du pouvoir irritant de n-hexane pour la peau et les yeux.
Le n-hexane est légèrement irritant et il provoque l'apparition
d'un érythème passager après un bref contact avec la peau. Des
effets plus graves (érythème et formation de phlyctènes) ont été
observés après un contact cutané de cinq heures avec de l'hexane de
qualité commerciale, la zone touchée étant recouverte. On n'a pas
suffisamment de données pour se prononcer sur le pouvoir irritant
de l'hexane liquide pour l'oeil humain. Lors d'une étude au cours
de laquelle des volontaires ont été exposés à de la vapeur d'hexane
à une concentration de 1760 mg/m3 (500 ppm) pendant trois à cinq
minutes, on a relevé aucun signe d'irritation oculaire. Il n'a pas
non plus été fait état de sensibilisation cutanée chez des ouvriers
exposés, ni lors d'un test de sensibilisation maximale effectué
avec ce produit.
3. Effets sur l'environnement
Le n-hexane est peu soluble dans l'eau et extrêmement volatil.
Les organismes aquatiques sont donc peu exposés à cette substance;
une exposition passagère peut cependant se produire en cas de
décharge sauvage dans les eaux superficielles. Les données dont on
dispose sur la toxicité du n-hexane pour les organismes aquatiques
sont très peu nombreuses; elles sont très diverses et ont été
obtenues dans des conditions peu satisfaisantes; dans ces
conditions, elles ne permettent pas d'évaluer convenablement le
risque pour l'environnement.
RECOMMANDATIONS
1. Protection de la santé humaine
Les concentrations de n-hexane auxquelles la population est
exposée dans son ensemble, ne représentent pour elle aucun danger.
Dans les ambiances de travail, il convient de maintenir les teneurs
dans l'atmosphère endessous des limites recommandées pour
l'exposition professionnelle en recourant à des procédés
convenablement étudiés et en utilisant certains dispositifs
mécaniques, notamment la ventilation. Dans les milieux confinés, en
cas d'urgence et lors de certaines opérations d'entretien, il est
souhaitable que des vêtements spéciaux et une protection
respiratoire appropriée soient mis à la disposition du personnel.
Il est recommandé d'utiliser un appareil respiratoire autonome à
débit continu pour des concentrations de n-hexane allant jusqu'à
8800 mg par m3 (2500 ppm). Entre 8800 mg/m3 (2500 ppm) et 17 600
mg/m3 (5000 ppm), l'appareil respiratoire doit comporter un masque
facial complet. Les gants protecteurs résistent, semble-t-il, à la
pénétration du n-hexane mais certains mélanges de n-hexane avec
d'autres solvants tels que la méthyléthylcétone pourraient
faciliter la pénétration du n-hexane à travers le tissu des gants
ou des vêtements protecteurs. C'est un facteur à prendre en compte
lorsqu'on utilise du n-hexane en mélange avec d'autres solvants.
2. Protection de l'environnement
Il est peu probable que le n-hexane présente un danger pour
l'environnement, sauf en cas de décharge ou de déversement très
importants, auquel cas des effets passagers peuvent se produire.
RECHERCHES A EFFECTUER
a) Il faudrait étudier la fonction testiculaire au moyen d'un
certain nombre d'indices parmi des groupes de travailleurs
exposés de par leur profession, afin d'établir si l'un ou la
totalité des effets observés chez l'animal se produisent
également chez l'homme.
b) Il conviendrait d'étudier plus à fond in vivo les effets
génotoxiques du n-hexane afin de compléter les données
limitées obtenues in vitro.
c) Il est nécessaire de procéder à une étude comportant une
exposition répétée au n-hexane afin d'établir quelle est sa
toxicité par voie orale. C'est à partir des résultats obtenus
qu'on pourra déterminer la dose sans effet nocif observable qui
permettra de fixer une DJA.
d) Le mécanisme de la neurotoxicité reste obscur au niveau
moléculaire. L'élucidation de ce mécanisme est d'un intérêt
direct en vue d'une estimation valable du risque encouru. Des
études in vivo et in vitro sont nécessaires pour étudier
plus à fond et clarifier ces mécanismes.
d) Les études de tératogénicité existantes ne permettent pas
d'évaluer le risque. D'autres études sont à entre-prendre pour
que l'on puisse disposer de données utilisables.
f) On n'a pas pu établir de relations dose-réponse nettes en ce
qui concerne les effets neurotoxiques du n-hexane chez l'homme
car les données relatives à l'exposition sont soit incomplètes
soit peu fiables. En outre, même si l'on a amplement démontré
l'existence d'effets neurotoxiques périphériques, les effets
sur le système nerveux central n'ont pas été suffisamment
étudiés. Il importe d'entreprendre d'autres études
épidémiologiques, notamment des études longitudinales
prospectives, afin de combler les lacunes des travaux
antérieurs.
RESUMEN
El n-hexano (hexano normal) es un líquido volátil e incoloro.
El hexano comercial está formado principalmente por una mezcla de
isómeros del hexano y de compuestos afines de seis átomos de
carbono; su contenido de n-hexano oscila entre el 20 y el 80%. Un
sistema adecuado para la medida del n-hexano es la combinación de
la cromatografía de gases con la detección de ionización por
conductor o la espectroscopía de masa. Los límites de la exposición
profesional varían entre 100 - 1800 mg/m3 (promedio ponderado en el
tiempo) y 400 - 1500 mg/m3 (valor máximo) en diversos países.
El n-hexano se puede aislar del gas natural y del petróleo
crudo. Se utiliza en la elaboración de alimentos, inclusive en la
extracción de aceite vegetal, y como disolvente en varios productos
y procesos.
Una vez liberado en el medio ambiente, el n-hexano se encuentra
fundamentalmente en fase de vapor. Se estima que su semivida en la
atmósfera es de unos 2 días, de acuerdo con su reactividad con el
radical OH solo. Los valores descritos de la CL50 para los
organismos acuáticos son pocos y variables, y se han determinado en
condiciones inadecuadas; por consiguiente, no es posible valorar
los efectos tóxicos del n-hexano en el medio ambiente. Su baja
solubilidad en agua y su elevada volatilidad hacen improbable la
exposición de los organismos acuáticos, salvo en el caso de vertido
incontrolado en aguas de superficie.
En los mamíferos, el n-hexano se absorbe rápidamente a través
de los pulmones y se distribuye por todo el cuerpo adulto, así como
en los tejidos fetales. La absorción cutánea es limitada. El
n-hexano sufre un metabolismo oxidativo y da diversos compuestos,
entre los que se encuentra la 2,5-hexanodiona, considerada el
agente neurotóxico final. En el nervio ciático de las ratas se
pueden encontrar niveles particularmente elevados de n-hexano y de
2,5-hexanodiona. La mayor parte del n-hexano se elimina inalterado
en el aire exhalado; otra parte se excreta metabolizado en el aire
exhalado y la orina.
El n-hexano administrado a ratas adultas por vía oral o
respiratoria tiene una toxicidad aguda baja. Para una hora de
exposición se han registrado valores de DL50 de 15 - 30 g/kg por
vía oral y de 271 040 mg/m3 (77 000 ppm) por vía respiratoria. En
concentraciones altas de vapor, los animales muestran ataxia,
convulsiones y signos de depresión del sistema nervioso central.
Parece ser que el efecto principal de la exposición repetida al
n-hexano en ratas es la aparición de lesiones testiculares y
neurotoxicidad. La exposición por vía respiratoria al n-hexano y
por vía oral a la 2,5-hexanodiona produjeron lesiones testiculares
graves. Estos efectos se han atribuido a la alteración del
citoesqueleto de las células de Sertoli. Se producen efectos
secundarios en las células germinales postespermatogónicas, que
desaparecen de los túbulos afectados. Los efectos testiculares
fueron reversibles tras la exposición única durante 24 h a 17 600
mg/m3 (5000 ppm) pero irreversibles tras exposiciónes de 16 h/día y
6 días/semana durante dos semanas a la misma concentración. La
ingestión de agua de bebida con un 1% de 2,5-hexanodiona durante
dos o tres semanas produjo lesiones testiculares reversibles
análogas y efectos irreversibles (en un plazo de 17 semanas) tras
cinco semanas de tratamiento.
El efecto neurotóxico se caracteriza clínicamente por debilidad
en las extremidades posteriores, que puede llegar a la parálisis.
En el sistema nervioso central y periférico se produce inflamación
axonal; se pueden presentar lesiones más graves (degeneración y
pérdida axonal) sobre todo en los nervios más largos y de diámetro
mayor. En estudios de inhalación prácticamente continua durante
seis meses con dosis de 1760 mg/m3 (500 ppm) o superiores se
produjeron lesiones en el sistema nervioso periférico y central,
pero no se observaron efectos clínicos o patológicos a
concentraciones de 440 mg/m3 (125 ppm). De 15 a 22 semanas después
de cesar la exposición constante a una concentración de vapor de
3520 mg por m3 (1000 ppm), cinco días a la semana durante 11
semanas, sólo se había producido una recuperación limitada de la
amplitud de la respuesta evocada por un estímulo auditivo del
quinto par craneal (que se considera que indica actividad del
sistema nervioso central) y del potencial de acción del nervio de
la cola. La exposición discontinua de ratas a una concentración de
3168 mg/m3 (900 ppm) durante 72 semanas no causó ninguna lesión
evidente en el sistema nervioso periférico o central, pero se
observaron indicios de efectos electrofisiológicos en los nervios
periféricos.
La neurotoxicidad inducida por el n-hexano puede aumentar con
la exposición simultánea a metiletilcetona, metilisobutilcetona y
acetato de plomo y disminuir con la exposición simultánea al
tolueno. El tolueno y el n-hexano ejercen también un efecto
sinérgico en la alteración de los niveles de dopamina.
Tras la aplicación cutánea de n-hexano en condiciones
oclusivas durante períodos cortos se observó en la piel la
aparición de lesiones microscópicas graves. La exposición
prolongada a una concentración de vapor de n-hexano de 10 560
mg/m3 (3000 ppm) puede ocasionar irritación de la conjuntiva en la
rata y pronunciada irritación ocular en el conejo. No se dispone de
datos sobre sensibilización cutánea a partir de estudios en
animales.
Se ha informado de la aparición de lesiones cromosómicas
(poliploidía en un estudio y aberraciones estructurales en otro)
en experimentos tanto in vivo como in vitro. En las pruebas
realizadas para determinar las lesiones en el ADN no se ha
observado aumento de la frecuencia de las mutaciones puntuales ni
de los efectos en ellas.
Se ha realizado un estudio de la carcinogenicidad del n-hexano
(pintando la piel de ratones), que resultó negativo.
No se ha estudiado lo bastante la toxicidad del n-hexano para
la reproducción. No se observaron indicios claros de que su
inhalación tenga en las ratas efectos embriotóxicos o
teratogénicos, aunque las concentraciones utilizadas fueron
relativamente bajas, y tampoco en ratones después de la
administración oral. Cuando, en un estudio con ratas, se expusieron
a las madres a una concentración de vapor de n-hexano de 3520
mg/m3 (1000 ppm), se retrasó de forma pasajera el desarrollo
postnatal de la descendencia.
Se dispone de muy poca información acerca de la toxicidad
aguda del n-hexano en la especie humana. La mayor parte de los
estudios se han centrado en la exposición profesional a mezclas de
disolventes. Los datos disponibles indican que el n-hexano tiene
una toxicidad aguda escasa. Se han señalado manifestaciones de
depresión del sistema nervioso central, como somnolencia, vértigo y
mareos, tras la exposición a n-hexano comercial en concentraciones
que van de 3520 a 17 600 mg/m3 (1000 - 5000 ppm) durante 10 - 60
minutos.
El n-hexano es una sustancia ligeramente irritante que al
entrar en contacto con la piel del hombre durante períodos cortos
produce un eritema pasajero. Se ha informado de efectos más graves
(eritema y vesículas) tras un contacto cutáneo oclusivo de cinco
horas con un hexano de tipo comercial. No se han notificado casos
de sensibilización cutánea en trabajadores expuestos, y tampoco se
observó en un ensayo de maximización con n-hexano.
Con exposiciones repetidas, el n-hexano es neurotóx-ico e
induce un tipo de neuropatía periférica sensomotora. Se han
publicado muchos estudios sobre la prevalencia de la neurotoxicidad
inducida por el n-hexano; sin embargo, a menudo se carece de datos
adecuados sobre la exposición. La exposición a concentraciones de
n-hexano en el aire que oscilan entre 106 y 8800 mg/m3 (30 - 2500
ppm) se ha relacionado con neuropatías. Se han descrito casos de
acusada neuropatía periférica entre trabajadores japoneses de la
industria de las sandalias y en tipógrafos de Taiwán expuestos a
niveles de n-hexano de unos 176 y 352 mg/m3 (50 y 100 ppm),
respectivamente, durante períodos superiores a 8 h diarias. En
muchos casos las mediciones de la exposición eran recientes y no
reflejaban de manera exacta las exposiciones anteriores causantes
de neuropatía.
En varios estudios cruzados se han observado por separado
efectos subclínicos ligeros (por ejemplo, cambios
electrofisiológicos en los nervios periféricos) en trabajadores
expuestos a 70 - 352 mg/m3 (20 - 100 ppm). En ninguno de esos
estudios se identificaron casos clínicamente evidentes de
neuropatía periférica con niveles de exposición inferiores a 352
mg/m3 (100 ppm).
Sólo en un pequeño número de estudios se han investigado los
efectos del n-hexano en el sistema nervioso central. Se ha
indicado que los cambios del potencial inducido por estímulos
somatosensoriales en trabajadores expuestos al n-hexano podrían
deberse a un bloqueo de la conducción del sistema nervioso central.
También se ha observado una alteración del potencial inducido por
estímulos visuales, así como modificaciones
electroencefalográficas. Estos resultados parecen indicar que el
n-hexano puede producir una disfunción del sistema nervioso
central. Sin embargo, los datos disponibles no aportan información
relativa a los niveles de exposición correspondientes.
EVALUACION DE LOS RIESGOS PARA LA SALUD HUMANA Y DE LOS EFECTOS EN
EL MEDIO AMBIENTE
1. Estudios en animales
1.1 Exposición a corto y largo plazo
El n-hexano tiene una toxicidad aguda baja; se han comunicado
valores de DL50 del orden de 15 - 30 g/kg y una CL50 (1 hora) de
271 040 mg/m3.
En los estudios de inhalación prácticamente continua en ratas
se produjo la muerte tras una exposición a 3520 mg/m3 (1000 ppm) de
n-hexano durante un período máximo de 11 semanas y se redujo la
ganancia de peso corporal después de tres semanas de exposición a
1760 mg/m3 (500 ppm). En estudios con una exposición diaria de 12
horas al n-hexano, se produjo la muerte con una concentración de
10 560 mg/m3 (3000 ppm) al cabro de 15 semanas, y una ligera
disminución de la ganancia de peso corporal con 740 mg/m3 (200 ppm)
después de 24 semanas. No se observaron efectos en el peso corporal
en ratas expuestas a 443 mg/m3 (126 ppm) de manera prácticamente
continua durante 34 semanas.
Los principales efectos no letales de la exposición repetida de
ratas al n-hexano parecen ser la toxicidad testicular y la
neurotoxicidad, tanto periférica como central. La exposición
respiratoria al n-hexano y la oral a la 2,5-hexanodiona producen
lesiones testiculares graves. Los efectos se han atribuido a la
alteración del citoesqueleto de las células de Sertoli. Hay efectos
secundarios en las células germinales postespermatogónicas, que
desaparecen de los túbulos afectados. Los efectos testiculares
fueron reversibles tras una exposición única de 24 horas a 17 600
mg/m3 (5000 ppm), pero se hacían irreversibles después de dos
semanas de exposición a la misma concentración durante 16 horas/día
y 6 días/semana. El agua de bebida con un 1% de 2,5-hexanodiona
produjo lesiones testiculares reversibles después de 2 - 3 semanas
de administración, y efectos irreversibles (en un plazo de 17
semanas) tras 5 semanas de tratamiento.
Clínicamente, la neurotoxicidad se caracteriza por debilidad en
las extremidades posteriores, que puede evolucioner hasta parálisis
y por último tetraplejía. Se han observado manifestaciones clínicas
tras 10 semanas de exposición discontinua a 10 560 mg/m3 (3000 ppm)
(12 horas/día, 7 días/semana) y después de 6 - 10 semanas de
exposición continua a una concentración de 1408-2112 mg/m3 (400 -
600 ppm).
Antes de la aparición de debilidad en las extremidades
posteriores, se observaron lesiones (inflamación axonal) en los
nervios de dichas extremidades, en el cerebro y en la médula
espinal. Tras seis meses de exposición a 1760 mg/m3 (500 ppm) se
apreciaron efectos más graves (degeneración y pérdida axonal),
principalmente en los nervios más largos y de diámetro mayor. En
conjunto, resulta difícil determinar el nivel de no observación de
efectos adversos correspondiente a la neurotoxicidad inducida por
el n-hexano en ratas, a causa de la variedad de concentraciones de
vapor y de tipos de exposición en las pruebas, y de la diversidad
de efectos investigados. En estudios de exposición por inhalación
prácticamente continua durante seis meses, se observaron lesiones
nerviosas con concentraciones de n-hexano de 1760 mg por m3 (500
ppm) o más, pero no se manifestaron efectos clínicos o patológicos
a 440 mg/m3 (125 ppm). La exposición discontinua de ratas a una
concentración de 3168 mg/m3 (900 ppm) durante 72 semanas no causó
ninguna lesión nerviosa manifiesta, pero hubo indicios preliminares
de un efecto en la fuerza prensil.
Cuando se interrumpe la exposición pueden desaparecer (parcial
o totalmente) los signos clínicos de toxicidad, la disminución de
la fuerza prensil y los efectos electrofisiológicos. De 15 a 22
semanas después de cesar la exposición repetida durante 11 semanas
a una concentración de 3520 mg/m3 (1000 ppm), sólo se observó una
recuperación limitada de la amplitud de la respuesta inducida por
los estímulos auditivos en el quinto componente del tronco
encefálico (que se considera que refleja la actividad del sistema
nervioso central) y del potencial de acción del nervio caudal. No
se ha investigado la desaparición de las lesiones patológicas
después de la exposición por vía respiratoria, pero se ha observado
remielinización y regeneración axonal limitada al interrumpirse la
exposición a concentraciónes de 1408 a 2112 mg/m3 (400 - 600 ppm).
El tratamiento discontinuo durante siete semanas por vía
respiratoria con 1760 mg/m3 (500 ppm) puede causar ligeras lesiones
pulmonares en la rata. En conejos se apreciaron, tras la exposición
a 10 560 mg/m3 (3000 ppm) 8 horas/día durante 8 días, lesiones
graves en el tracto respiratorio y, después de 24 semanas de
exposición, proliferaciones papilares de células bronquiolares no
ciliadas.
En algunos estudios de inhalación en ratas se han señalado
efectos en el riñón y el hígado, pero solo con niveles de
exposición más elevados. La mayor parte de las lesiones y otros
efectos se consideraron de escasa importancia toxicológica y/o no
se reprodujeron en estudios repetidos. Sin embargo, tras la
administración intraperitoneal repetida de n-hexano a ratas se han
observado lesiones hepáticas graves.
Los estudios de exposición combinada han puesto de manifiesto
que los efectos neurotóxicos del vapor de n-hexano en las gallinas
pueden aumentar con la presencia de metilisobutilcetona y en las
ratas con la de metilisobutilcetona y acetato de plomo, mientras
que el tolueno los hace disminuir. También se ha señalado que la
exposición de ratas a la combinación con metiletilcetona aumenta
las lesiones pulmonares.
Se han realizado algunos estudios sobre los efectos de la
exposición oral o cutánea al n-hexano. La administración diaria
por vía oral de 500 mg/kg durante cuatro semanas produjo en las
ratas una disminución de la velocidad de conducción en el nervio
caudal.
1.2 Genotoxicidad
Se han realizado pocas pruebas de mutagenicidad. Se ha
informado de la aparición de lesiones cromosómicas (poliploidía en
un estudio, y aberraciones estructurales en otro) en ensayos in
vitro, en los que se expuso una línea celular de hámster chino a
n-hexano líquido no diluido en ausencia de fracción metabolizante.
Tras la exposición de células cultivadas de mamíferos o de
bacterias a n-hexano no se produjo un aumento significativo del
índice de mutaciones puntuales. Las lesiones en el ADN, medidas en
ensayos de toxicidad diferencial en bacterias, no aumentaron por la
exposición al n-hexano. Tampoco observó aumento de la síntesis no
programada del ADN en linfocitos humanos in vitro. Aunque el
n-hexano ha dado resultados negativos en los ensayos para detectar
mutaciones puntuales, hay algunas pruebas de que puede producir
aberraciones cromosómicas in vitro. En un estudio in vivo con
células de médula ósea de rata se informó de lesiones cromosómicas
(roturas cromatídicas).
1.3 Carcinogenicidad
No se ha estudiado de manera adecuada el potencial
carcinogénico del n-hexano. En un estudio limitado de aplicación
en la piel a ratones, no pareció que aumentase la formación de
tumores cutáneos tras la exposición a un iniciador cancerígeno
(7,12-dimetilbenzantraceno) o junto con un agente promotor (aceite
de crotón).
1.4 Efectos en la reproducción
No se ha investigado debidamente la toxicidad del n-hexano
para la reproducción. No hay pruebas sólidas de embriotoxicidad o
teratogenicidad en las ratas tras su inhalación, ni tampoco en los
ratones después de la administración oral. En ratas expuestas a una
concentración de 10 560 mg/m3 (3000 ppm) se observó un ligero
aumento de la incidencia de algunas anomalías, pero las
deficiencias de método del estudio (por ejemplo, es probable que
las concentraciones ensayadas no fueran lo bastante altas) limita
el valor de las conclusiones. En ratones se detectaron signos de
fetotoxicidad (menor peso de las crías), pero sólo con dosis que
provocaron una letalidad considerable en las madres. En un estudio
con crías de ratas expuestas a una concentración de 10 560 mg/m3
(3000 ppm) se informó de un retraso pasajero del desarrollo
postnatal. Aunque no se describieron signos de neuropatía en esos
animales, no es posible evaluar a partir de ese estudio si la
podría inducir la exposición in utero, puesto que los animales sólo
estuvieron expuestos al n-hexano durante una pequeña parte del
período de gestación. En otros informes se ha afirmado que la
exposición in utero al n-hexano retarda la histogénesis cerebelar y
la maduración neuronal e induce anomalías electrofisiológicas en el
sistema nervioso central de ratas recién nacidas. Sin embargo, este
trabajo no se ha expuesto con suficiente detalle para hacer una
evaluación completa.
Existen pruebas de que el n-hexano causa esterilidad
irreversible en las ratas. En un estudio limitado de la letalidad
dominante en ratones no se registraron efectos en la fecundidad del
macho. No se han comunicado investigaciones acerca de la fertilidad
de las hembras ni estudios multigeneracionales.
2. Estudios en la especie humana
2.1 Efectos en el sistema nervioso
En muchos estudios se ha relacionado la exposición laboral al
n-hexano con la frecuencia de la neuropatía periférica en la
especie humana. Sin embargo, pocos de esos estudios informaban
acerca de las concentraciones en el aire, los datos comunicados
pueden ser inexactos y a menudo no está claro si esas
concentraciones eran de hexano o de n-hexano. Además, la
exposición industrial al n-hexano suele darse al utilizar productos
a base de disolventes, de manera que los trabajadores están
expuestos a una mezcla de compuestos volátiles (por ejemplo,
acetato de etilo, metiletilcetona, tolueno, acetona y otros
hidrocarburos alifáticos). Raramente se ha señalado la
concentración en el aire de los otros disolventes presentes. En el
presente examen sólo se han incluido los estudios que contenían
algunos datos de las concentraciones en el aire.
Los resultados de la electrofisiología de los nervios
periféricos pueden variar de manera considerable en función del
tipo de técnica utilizada, la temperatura ambiente a la que se
efectúan las mediciones, el segmento del nervio estudiado y la edad
del individuo. Así pues, sin información sobre esos factores es
difícil evaluar los estudios que se basan en la electrofisiología
de los nervios periféricos.
Mediante el estudio de 93 casos de neuropatía se definieron las
características clínicas de la producida por el n-hexano y se puso
de manifiesto que existe una correlación clara entre las medidas
electrofisiológicas y la gravedad de trastorno. Se encontró una
relación entre el aumento de la gravedad y el de la exposición. Sin
embargo, no fue posible establecer una relación definitiva a partir
de los escasos detalles notificados sobre la exposición.
En los trabajadores del calzado expuestos a una concentración
de 70 mg/m3 (20 ppm) se encontraron signos subclínicos leves de
neuropatía. El grado de anomalía electrofisiológica aumentaba con
la duración de la exposición. Sin embargo, no se produjeron casos
de neuropatía manifiesta, ni siquiera en trabajadores con una
exposición de más de cinco años de duración.
Se observaron signos subjetivos y electrofisiológicos de
neuropatía periférica subclínica en un grupo de trabajadores
expuestos a 317 mg/m3 (90 ppm). Sin embargo, esos efectos no se
pueden atribuir exclusivamente a la exposición a esta concentración
de n-hexano, puesto que probablemente se infravaloró la exposición
anterior a disolventes y la autoselección de los trabajadores puede
haber dado lugar a cierto sesgo. Además, se puede haber producido
una exposición simultánea a la metiletilcetona, que, como se ha
demostrado, aumenta los efectos neurotóxicos del n-hexano.
La exposición a una mezcla de disolventes con una concentración
aproximada de n-hexano de 690 mg/m3 (196 ppm) causó una reducción
de la velocidad de conducción de los nervios periféricos, que se
mantuvo tras disminuir la exposición de manera significativa. Se ha
indicado asimismo que la causa de algunos de los cambios de
potencial inducidos por estímulos somatosensoriales podría ser un
efecto del n-hexano en el sistema nervioso central.
En un pequeño grupo de trabajadores expuestos a una
concentración media de n-hexano en el aire de 208 mg/m3 (59 ppm)
se produjeron signos subjetivos y electrofisiológicos de una
neuropatía subclínica leve. No se detectó neuropatía clínica
manifiesta. En este estudio, la exposición a otros disolventes fue
limitada y los estudios de conducción nerviosa se realizaron en
condiciones definidas. Sin embargo, el pequeño número de individuos
estudiados, la ausencia de historial de exposición y la escasa
correspondencia entre los grupos de exposición y testigo limitan el
valor de las conclusiones alcanzadas. No obstante, los resultados
en general coinciden con los datos obtenidos en anteriores estudios
de los que se informa más arriba. La exposición a 669 mg/m3 (190
ppm) durante períodos superiores a 8 h/día se asoció con la
aparición de neuropatía periférica clínicamente manifiesta. Puede
producirse una lenta recuperación parcial de la función nerviosa a
lo largo de varios años. La exposición a concentraciones inferiores
a 352 mg/m3 (100 ppm) no produjo neuropatía, pero aparecieron
pruebas electrofisiológicas de efectos subclínicos.
En un pequeño grupo de trabajadores profesionalmente expuestos
a niveles muy diversos de n-hexano en el aire se han descrito
defectos en cuanto a la distinción de los colores y signos de
maculopatía. La medición del potencial inducido por estímulos
visuales y el electrorretinograma de esos individuos mostró
anomalías en comparación con un grupo testigo de trabajadores no
expuestos. También se ha informado de anomalías en el trazado del
potencial causado por estímulos visuales en sujetos inhaladores de
pegamento expuestos a n-hexano. Se ha indicado que puede
producirse degeneración axonal en la vía óptica y que el n-hexano
puede inducir un bloqueo de la conducción en el sistema nervioso
central. También hay algunas pruebas de que existe cierta
correlación entre los signos de maculopatía y los signos
electrofisiológicos de neuropatía periférica. Se encontraron
asimismo anomalías en el trazado electroencefalografico de algunos
de los individuos estudiados.
2.2 Otros efectos
No se conoce ningún estudio sobre la fertilidad o los posibles
efectos testiculares en los varones. Es muy escasa la información
sobre la toxicidad aguda del n-hexano en la especie humana. La
mayor parte de los estudios han consistido en la exposición a
mezclas de disolventes. Los datos disponibles parecen indicar que
el n-hexano tiene una toxicidad aguda baja.
Se han publicado muy pocos estudios sobre la irritación cutánea
y ocular en voluntarios. El n-hexano es un agente ligeramente
irritante que cuando entra en contacto durante un corto período con
la piel del hombre causa un eritema pasajero. Se han descrito
efectos más graves (eritema y vesículas) tras un contacto cutáneo
oclusivo de cinco horas con n-hexano de tipo comercial. Se carece
de datos suficientes para evaluar si el líquido puede causar en el
hombre irritación ocular. No se observaron signos de irritación de
los ojos en un estudio en que se expuso a un grupo de voluntarios a
vapor de hexano con una concentración de 1760 mg/m3 (500 ppm)
durante 3 - 5 minutos. No se han comunicado casos de
sensibilización cutánea en trabajadores expuestos y tampoco se ha
observado en una prueba de maximización con n-hexano.
3. Efectos en el medio ambiente
El n-hexano es poco soluble en agua y muy volátil. Por
consiguiente, la exposición de los organismos acuáticos es
probablemente baja y pasajera, excepto en el caso de vertidos
incontrolados en aguas de superficie. Los datos acerca de la
toxicidad del n-hexano para los organismos acuáticos son escasos,
variables, obtenidos en condiciones inapropiadas e insuficientes
para evaluar del riesgo para el medio ambiente.
RECOMENDACIONES
1. Protección de la salud humana
La población general está expuesta a unas concentraciones de
n-hexano a las que es improbable que corra algún riesgo. En
relación con la salud en el trabajo, los niveles atmosféricos se
deben mantener por debajo de los límites de exposición profesional
recomendados, mediante el diseño de procesos de trabajo adecuados y
por medio de controles técnicos, incluido el de la ventilación. En
los espacios cerrados, en casos de emergencia y en determinadas
actividades de mantenimiento se debe disponer de ropas protectoras
adecuadas y protección de la respiración. Se recomienda el uso de
respiradores con admisión de aire puro o respiradores autónomos con
flujo continuo de oxígeno para concentraciones de n-hexano de
hasta 8800 mg/m3 (2500 ppm). En el caso de concentraciones entre
8800 mg/m3 (2500 ppm) y 17 600 mg/m3 (5000 ppm), el respirador debe
llevar adaptada una protección facial completa. Aunque los guantes
pueden impedir la penetración del n-hexano, las mezclas de éste
con otros disolventes, como la metiletilcetona, pueden permitir al
n-hexano pasar a través de los guantes y de otras prendas
protectoras. Hay que tener en cuenta este factor cuando se utilicen
mezclas de n-hexano.
2. Protección del medio ambiente
No es probable que el n-hexano represente un peligro para el
medio ambiente, excepto en el caso de derrames o vertidos
importantes, que podrían provocar efectos locales pasajeros.
NUEVAS INVESTIGACIONES
a) Se debería hacer un estudio sobre los índices de la función
testicular en uno o varios grupos de trabajadores expuestos
para establecer si alguno o todos los efectos cuya presencia
se ha descrito en los testículos de los animales se manifiestan
también en el hombre.
b) Conviene seguir investigando in vivo los efectos genotóxicos
del n-hexano, a fin de complementar la limitada información
que se posee sobre estos efectos in vitro.
c) Para establecer la toxicidad del n-hexano administrado por vía
oral es necesario realizar un estudio de dosis repetidas. Es
preciso hacer esto para determinar el nivel sin efecto
observado, a fin de definir una ingesta diaria admisible (IDA).
d) No está claro el mecanismo de la neurotoxicidad a nivel
molecular. Su conocimiento contribuirá directamente a la
validez de la evaluación del riesgo. Se necesitar estudios
in vivo e in vitro para seguir investigando y aclarar esos
mecanismos.
e) Los estudios de teratogenia disponibles no bastan para evaluar
el riesgo. Conviene realizar estudios que proporcionen datos
utilizables.
f) No se conoce la relación dosis-respuesta en cuanto a los
efectos neurotóxicos del n-hexano en la especie humana, debido
a que los datos de exposición son incompletos o no fidedignos.
Además, aunque se ha demostrado ampliamente la neurotoxicidad
periférica del n-hexano, los efectos en el sistema nervioso
central sólo se han examinado de forma limitada. Deben
realizarse más investigaciones epidemiológicas, como por
ejemplo estudios prospectivos de seguimiento, para subsanar la
escasez de datos de las anteriores investigaciones.
Monoclonal antibodies to neurofilament proteins have been used
to demonstrate protein cross-linking in the spinal cords of rats
treated with 2,5-hexanedione (Lapadula et al., 1986). In male
Sprague-Dawley rats given 2,5-hexanedione in drinking-water at
concentrations of up to 1% (v/v) for 70 days, there was dose-
dependent protein cross-linking, with decreased protein
phosphorylation of neurofilament proteins and microtubulin-
associated protein-2 (MAP-2) (Abou-Donia et al., 1988). The use of
monoclonal antibodies to neurofilament proteins has permitted the
demonstration of protein cross-linking in the spinal cords of rats
treated with 2,5-hexanedione (Lapadula et al., 1986). It was also
found that protein phosphorylation was decreased in association
with protein cross-linking in 2,5-hexanedione-treated rats (Abou-
Donia et al., 1988). It is not known if the decrease in protein
phosphorylation is a primary event or a consequence of cross-
linking. Exposure of three cell lines derived from human tumours
or fetal mouse brain to 2,5-hexanedione in vitro (0.04 - 0.16%)
resulted in reduced proliferation and an increase in cell
protrusions or dendritic-like processes (Malorni et al., 1989),
indicating that in vitro toxicity is not restricted to the neuronal
system.
2,5-Hexanedione causes aggregation of intermediate filaments of
the vimentin type in cultured human skin fibroblasts. Exposure to
2.8 mmol 2,5-hexanedione/litre for 1 or 2 weeks induced aggregation
in 50% and 80% of the cells, respectively; a concentration of 2
mmol/litre required 2 and 3 weeks to produce corresponding effects
(Durham et al., 1983). The non-neurotoxic analogues,
1,6-hexanedione and 2,4-hexanedione, had no effect on filament
distribution in fibroblasts, although in the latter case
cytotoxicity prevented the study of effects at concentrations
equivalent to those required for 2,5-hexanedione-induced
aggregation (Durham et al., 1988).
The neurotoxicity of 2,5-hexanedione was increased by
co-exposure to methyl ethyl ketone (Ralston et al., 1985). Exposure
to other apparently non-neurotoxic aliphatic ketones (2-pentanone,
2-heptanone, and 2-octanone) increased the neurotoxicity of the
n-hexane metabolite 2-hexanone; 2-octanone had a greater effect
than methyl ethyl ketone (Misumi & Nagano, 1985). Ladefoged et al.
(1989) dosed rats with 2,5-hexanedione (0.5% in the drinking-water)
for six weeks and compared the effects obtained with or without
administration of 0.5% acetone or 5.0% ethanol. Nerve conduction
velocity and behavioural rotarod performance were reduced after 3
weeks of dosing with 2,5-hexanedione. Acetone, but not ethanol,
together with 2,5-hexanedione, had an additional significant effect
on these parameters from the fourth week of dosing.
9. EFFECTS ON MAN
9.1. Single Exposures
In studies on human volunteers, inhalation of 17 600 mg
hexane/m3 (5000 ppm) for 10 min resulted in vertigo and giddiness
but there were no symptoms with exposure to 7040 mg/m3 (2000 ppm)
for the same duration (Patty & Yant, 1929). Occupational exposures
to hexane concentrations of 3520 to 89 760 mg/m3 (1000 to 25 500
ppm) for periods of 30 - 60 min caused drowsiness (Yamada, 1967).
9.2. Skin and Eye Irritation; Sensitization
9.2.1. Skin irritation
Prolonged occlusive skin contact for 1 - 5 h with liquid hexane
caused erythema and, after 5 h, blistering (Oettel, 1936).
Application of 1.5 ml n-hexane (analytical grade) to 3.1 cm2 of
forearm skin for 5 min resulted in a stinging and burning sensation
and transient erythema (Wahlberg, 1984a). 0.1 ml n-hexane rubbed
gently into the forearm skin for 18 days did not produce erythema
or oedema (Wahlberg, 1984b).
Operators at a soybean hexane-extraction facility had a higher
incidence of dry or irritated skin than maintenance workers (65%
versus 20%) (NIOSH, 1983).
9.2.2. Eye irritation
Eye irritation did not occur in volunteers exposed to hexane
vapour at a level of 1760 mg/m3 (500 ppm) for 3 - 5 min (Nelson et
al., 1943).
9.2.3. Skin sensitization
A maximization test on 25 subjects with n-hexane applied
undiluted for induction and as a 25% solution for challenge did not
induce skin sensitization (Kligman, 1966).
9.3. Short- and Long-Term Exposures
9.3.1. Effects on the nervous system
9.3.1.1 Peripheral neuropathy
Peripheral neuropathy has been reported in humans exposed
industrially to n-hexane or through solvent abuse of glues or
solvents containing n-hexane. The majority of occupational cases
have occurred in small industrial concerns such as shoe manufacture
in Italy and vinyl sandal production in Japan (Sobue & Yamamura,
1968; Yamamura, 1969; Inoue et al., 1970; Abbritti et al., 1976;
Carapella, 1977; Palao & Lajo, 1981; Nakajima & Murayama, 1985).
Workers were exposed to many volatile compounds, not only
n-hexane, in badly ventilated rooms, sometimes for periods in
excess of 8 h/day under poor hygienic conditions (Yamamura, 1969;
Inoue et al., 1970, 1971; Battistini et al., 1974; Abbritti et al.,
1976; Carlomagno et al., 1983).
The first signs of n-hexane neuropathy are symmetrical
paraesthesia and weakness. The lower extremities are normally
affected first. A number of other symptoms such as headache,
anorexia, and dizziness may precede or coincide with the
neuropathy. A "glove and stocking" sensory impairment to touch,
pain, vibration, and temperature develops, with weakness and
atrophy affecting proximal muscles of the extremities. On clinical
examination, most patients show reduced body weight and diminished
or absent reflexes. There is a marked reduction in conduction
velocity in sensory and motor nerves. Electromyography (EMG) shows
a neuropathic pattern including fibrillation potentials, positive
sharp waves, polyphasic potentials, and reduced interference
patterns. Axonal lesions of the large myelinated fibres (identical
to those produced in experimental animals) with axonal swelling
followed by myelin retraction at the node of Ranvier are found on
nerve biopsy. There is only a slight to moderate reduction in the
number of fibres, mainly of large myelinated fibres (Goto et al.,
1974; Shirbae et al., 1974; Okamoto et al., 1977; Rizzuto et al.,
1977, 1980; Scelsi et al., 1980; Chauplannaz et al., 1982; Lagueny
et al., 1982). There are masses of 9- to 10-nm long neurofilaments
in the axons. Lesions are rarely found in the small myelinated and
unmyelinated fibres (Shirbae et al., 1974; Altenkirch et al., 1977;
Rizzuto et al., 1977, 1980; Scelsi et al., 1980; Ruff et al., 1981;
Chauplannaz et al., 1982). Muscle biopsies show signs of
myofibrillar atrophy (Yamamura, 1969; Gonzalez & Downey, 1972;
Shirbae et al., 1974; Altenkirch et al., 1977). Recovery is always
gradual and slow and signs of residual neuropathy may persist in
severe cases for 3 - 4 years (Yamamura, 1969; Iida & Yamamoto,
1973; Altenkirch et al., 1977; Sobue et al., 1978; Rizzuto et al.,
1980; Carlomagno et al., 1983). Signs of pyramidal tract defects
such as residual hyper-reflexia and spasiticity and autonomic
defects have been reported in a few severe cases (Korobkin et al.,
1975; Altenkirch et al., 1977; Altenkirch & Schultz, 1979;
Schaumburg & Spencer, 1979). Electrophysiological evidence of
central nervous system dysfunction (abnormal evoked potentials) has
occasionally been reported in individual cases of peripheral
neuropathy (Yoshida et al., 1974; Lagueny et al., 1982).
Neuropathy has occurred following exposure to a wide range of
n-hexane levels in air from 97 to 8125 mg/m3 (30 - 2500 ppm) (Oishi
& Mineno, 1964; Yamada, 1964, 1967; Wada & Okamoto, 1965;
Herskowitz et al., 1971; Nomiyama et al., 1973; Yoshida et al.,
1974; Takeuchi et al., 1975; Paulson & Waylonis, 1976; Takahashi et
al., 1977; Brugnone & Perbellini, 1980; Ono & Takeuchi, 1980; Ruff
et al., 1981; Carlomagno et al., 1983). Exposure durations ranging
from 2 months to 5 years may precede the onset of overt neuropathy,
and a mild recurrent neuropathy with n-hexane exposure may go
undetected (Wada & Okamoto, 1965; Herskowitz et al., 1971; Nomiyama
et al., 1973; Takeuchi et al., 1975; Takahashi et al., 1977; Ruff
et al., 1981).
In the Italian shoe industry, n-hexane (a component of the
glues used) has been implicated as the cause of peripheral
neuropathies (Di-Bosco & Fonzi, 1974; Abbritti et al., 1976;
Cianchetti et al., 1976; Carapella, 1977; Buitatti et al., 1978;
Aiello et al., 1980; Sanagi et al., 1980; Palao & Lajo, 1981; Mutti
et al., 1982a,b,c).
Towfighi et al. (1976) reported the development of neuropathy
in two patients who sniffed glue containing n-hexane. Complete
clinical recovery occurred with one patient who changed to a glue
not containing n-hexane. Sural nerve biopsies of the affected
individuals indicated loss of axons. Some remaining axons had
accumulation of filaments, widened nodes of Ranvier, and focal
enlargements. Muscle biopsy revealed neurogenic atrophy. Three
individuals in a laboratory developed neuropathy that was
attributed to n-hexane in glue (80.4% n-hexane). In addition to
axonal changes, the main findings were breakdown of nerve fibres
and swelling and degeneration of myelin. In addition, a
denervation muscle atrophy, combined with focal myopathic changes,
occurred (Scelsi et al., 1981).
a) Japanese sandal workers
Health surveillance studies were carried out in Japan following
a mass outbreak of polyneuropathy in 1967 (Sobue & Yamamura, 1968;
Iida et al., 1969, Yamamura, 1969; Inoue et al., 1970, 1971; Iida &
Yamamoto, 1973). Through the use of questionnaires and medical
examinations, 93 cases of polyneuropathy were identified out of
1662 vinyl sandal workers who were using glues based on solvents
containing more than 70% commercial grade hexane. Their work was
carried out for periods of up to 14 h/day in small, badly
ventilated rooms with estimated hexane air concentrations of
between 1625 and 8125 mg/m3 (500 and 2500 ppm). Air concentrations
of the other volatile solvents in the glue were not described.
The workrooms were often used also as dwelling places so that
continuous exposure for up to 24 h/day was possible. Clinically,
the cases were classified into three groups, according to the
severity of the disease:
Group I: sensory neuropathy (53 cases);
Group II: sensorimotor neuropathy (32 cases); and
Group III: motor neuropathy with amyotrophy (8 cases).
Single exposure estimations at the homes/workrooms of some of
the patients indicated a positive relationship between the air
concentration of hexane, the duration of the working day and the
number of sandals made, and the severity of the polyneuropathy.
n-Hexane concentrations of 1625 - 3250 mg/m3 (500 - 1000 ppm)
during an 8-h day were sufficient to cause severe peripheral
neuropathy (Inoue et al., 1971).
In 89% of the cases, the onset was insidious. Initial symptoms
were sensory impairment (88%), muscular weakness (14%), blurred
vision (14%), coldness of extremities (13%), headache (9%), loss of
body weight (9%), lassitude (3%), skin eruptions (2%), anorexia
(2%), and dizziness (1%). On examination, the most common finding
was a symmetrical sensorimotor disorder of the extremities.
Hypoaesthesia was found in all cases and dysaesthesia was found in
the early stages of the disease in 23% of patients. Other finding
were muscular weakness of the limbs (43%), diminished tendon
reflexes (39%), and exaggerated tendon reflexes (11%). Numbness of
the face was described in five cases. Of 39 cases examined
ophthalmologically, eight had abnormal findings, seven had
restriction of the visual field, two had optic nerve atrophy, and
one had optic neuritis. There was no correlation between the
severity of neuropathy and the severity of optic effects.
Electromyography (EMG) was performed on 44 patients (11 in
Group I, 25 in Group II, and 8 in Group III) (Iida et al., 1969;
Yamamura, 1969). Signs of denervation, including fibrillation and
positive sharp waves, were found in 70% of cases in Groups I and II
and in all cases in Group III. A reduction in the number of motor
units under voluntary control (reduced interference pattern) was
found in 50% of cases in Group I and 70% of cases in Group III.
High amplitude and polyphasic action potentials were more common in
Group III than in Groups I and II. The severity of the neuropathy
correlated with the type and magnitude of electromyographic
abnormalities.
Motor nerve conduction velocities and evoked potentials (mixed
nerve conduction velocity) were measured in the ulnar, median,
peroneal, and tibial nerves in the same 44 cases. Sensory nerve
conduction velocities were measured in the ulnar and median nerves.
The number of cases with reduced motor nerve and sensory nerve
conduction velocities correlated with the severity of the
neuropathy. Reduced mixed nerve conduction velocity correlated
only with the severity of neuropathy in measurements obtained from
peroneal and tibial nerves.
Muscle and nerve biopsies were taken from a small number of
cases. Sections of muscle showed neurogenic atrophy. Sections of
nerve showed axonal and myelin lesions, and demyelination was
considered to be the primary lesion (data obtained from animal
studies indicate that the primary lesion in n-hexane neuropathy is
axonal).
Polyneuropathy progressed in all severe cases (Group III) for
up to 4 months after cessation of exposure. A gradual, slow
recovery then ensued. Of 36 cases that were followed-up, 50%,
including six severe cases (Group III), had completely or nearly
completely recovered within 3 - 18 months. Two cases in Group III
displayed residual atrophy and weakness. There were no deaths
(Sobue & Yamamura, 1968; Yamamura, 1969; Inoue et al., 1970).
The clinical recovery of 90 cases was followed for
approximately 5 years. Approximately 3 years after the outbreak,
51 cases had recovered completely. The remaining 39 cases were
reclassified into Groups I and II (34 and 5 cases, respectively).
After 5 years, 82 cases had recovered completely, while the
remainder had residual sensory symptoms (Iida & Yamamoto, 1973;
Sobue et al., 1978). A further study reported a significant
improvement during the 5-year follow-up period in the EMG and nerve
conduction velocities of some patients initially placed in Groups I
and II (Iida, 1982). Patients in Group III also showed
improvements in the EMG. Denervation activity and polyphasic
potentials disappeared and interference patterns and high-voltage
action potentials reappeared. In some patients, nerve conduction
velocities were depressed for over a year after the onset of the
neuropathy.
In 1981, 21 new cases of mild neuropathy were identified in the
same district (Iida, 1982). Air measurements at the workplaces of
the sandal makers indicated that n-hexane air levels were below 176
mg/m3 (50 ppm). These cases were divided by clinical examination
into 20 cases in Group I and one case in Group II. Signs of
neuropathy included numbness (twelve cases), dysaesthesia (six
cases), slight muscle weakness (six cases), diminished tendon
reflexes (ten cases), and exaggerated tendon reflexes (three
cases). Skin changes (coldness, redness, and roughness) were
observed in four cases. A slight floating sensation on change of
posture was reported by five patients.
EMG did not reveal any denervation in resting muscles. EMG
abnormalities, more severe in the lower extremities, including
reduced interference pattern, large amplitude, and polyphasic
potentials, were recorded during voluntary muscle contraction in
approximately 75% of cases. Significant reductions in motor or
sensory nerve conduction velocities were reported in nearly half
the patients.
b) Italian shoe industry
A study of exposure to organic solvents was carried out in an
Italian shoe factory (Mutti et al., 1982a). The exposed group
consisted of 95 workers (24 males, 71 females) with an age range of
16 - 62 years (mean, 30.9 ± 11.7 years), and the exposure duration
ranged from 1 to 25 years (mean, 9.1 ± 8 years). The approximate
mean air concentrations in the breathing zone, over a 2-year
period, for a number of solvents were: n-hexane, 317 mg/m3 (90
ppm); cyclohexane, 315 mg/m3 (92 ppm); methyl ethyl ketone, 115
mg/m3 (39 ppm); and ethyl acetate, 205 mg/m3 (57 ppm). A
concurrent reference group from the same factory of 52 non-exposed
workers (12 males, 40 females) with an age range of 16 - 58 years
(mean, 29.6 ± 12.3 years) and an employment duration ranging from 4
months to 29 years (mean, 10.2 months ± 9.7 years) was selected.
Both exposed and reference workers underwent detailed physical
examination and electrophysiological studies of the median, ulnar,
and peroneal nerves. An exposure score (work time in years
multiplied by an estimate of total solvent exposure) was calculated
for each of the exposed workers.
Acute and chronic neurological signs and symptoms that were
more frequently reported by, or found in, the exposed workers
included sleepiness, dizziness, weakness, paraesthesia, and
hypoaesthesia. Other neurological symptoms, such as headache,
muscular cramps, neurasthenic syndrome, and sleep disturbances,
were found more often in exposed workers, but the differences in
incidence between the exposed and reference group were not
statistically significant. Some parameters of electrophysiological
measurements indicated subclinical neuropathy among exposed
workers. The mean motor nerve conduction velocity was
significantly reduced in the median and peroneal nerves but not in
the ulnar nerve. The amplitude of the motor action potential (MAP)
was significantly reduced in all nerves and its duration was
increased in the ulnar nerve. There were no significant effects on
the distal latency. A significantly higher number of abnormal
action potentials was observed in the median and peroneal nerves of
exposed workers. There was a correlation between the reduction in
motor conduction velocity and the exposure score, but not with age
of the individual. Comparison of the electrophysiological
parameters between two exposed sub-groups and the reference group
indicated an exposure-related trend for reduced motor conduction
velocity in the median and peroneal nerves, and an increased motor
action potential amplitude duration.
When the 95 exposed workers were divided into two sub-groups
based on n-hexane exposure, 65 workers having low exposure (243
mg/m3, 69 ppm) and 30 having high exposure (475 mg/m3, 135 ppm),
there was an exposure-related trend in the reduction of median
nerve motor conduction velocity and increase in the duration of the
ulnar nerve motor action potential (Mutti et al., 1982b).
In a follow-up study, electrophysiological measurements
including somatosensory evoked potentials (SEPs) were recorded from
a group of 15 women shoe factory workers aged 19 - 53 years (mean
age, 26.6 ± 11.4 years) with an exposure duration to solvents of
2 - 8 years (mean, 4.5 ± 2.3 years) (Mutti et al., 1982c). The
mean air concentrations for various solvents in the breathing zone
of the workers were: n-hexane, 690 mg/m3 (196 ppm); cyclohexane,
585 mg/m3 (170 ppm); methyl ethyl ketone, 177 mg/m3 (60 ppm); and
ethyl acetate, 360 mg/m3 (100 ppm). However, total solvent levels
occasionally exceeded 5000 - 7000 mg/m3 when the ventilation system
failed. The study started 3 months after hygiene improvements
successfully controlled air levels of solvents. A control group of
15 age- and health-matched women was used.
Electrophysiological measurements in peripheral nerves showed
significant reductions in maximal motor and distal sensory nerve
conduction velocities in the median and ulnar nerves and reduced
maximal motor nerve conduction velocity in the peroneal nerve. The
latency of the sensory peak action potential was significantly
increased in the median and ulnar nerves. The amplitude of all
peripheral nerve action potentials was slightly reduced but this
was not statistically significant.
There were changes in the somatosensory evoked potentials
(SEPs) of exposed workers with significant increases in the latency
of some early component peaks. The amplitude of some of the early
peaks was also increased, but the amplitude of the later peaks was
significantly reduced. Comparison of SEPs from exposed and non-
exposed workers showed that later components in exposed workers
were flatter.
An electrophysiological study was carried out on 25 workers (13
women, 12 men), placed into two groups according to job
specification, out of 31 workers at an Italian shoe factory (Aiello
et al., 1980). Group A contained 11 subjects (seven women and four
men), aged 21 - 50 years with 0.3 to 20 years of exposure, who
worked in direct contact with solvents. Group B contained 14
subjects (six women and eight men), aged 20 - 49 years and exposed
for 0.25 - 20 years, who had no direct contact with solvents but
were exposed by inhalation. Peak values for air concentrations of
various solvents in the factory were: n-hexane, 70 mg/m3 (20 ppm);
ethyl acetate, 1.8 mg/m3 (0.5 ppm); cyclohexane, 620 mg/m3 (40
ppm); and trichloroethylene, 2.7 mg/m3 (0.5 ppm).
Motor nerve conduction velocity was measured in the median and
peroneal nerves, and sensory nerve conduction velocity in the
median and ulnar nerves. The distal latency and amplitude of
action potentials were measured in motor and in sensory fibres.
Statistically significant electrophysiological changes were
found in seven workers from Group A. An increased distal latency
in sensory or motor fibres was found in six of these workers and
reduced motor nerve conduction velocity in the peroneal nerve in
one worker. In three workers from Group B, there was a significant
increase in the distal latency in sensory nerves of the upper limb.
In one of these, there was reduced motor nerve conduction velocity.
EMG abnormalities were detected in two workers, one from each
group. Seven out of 14 workers exposed for more than 5 years
exhibited one or more electrophysiological abnormalities. In three
out of eight workers exposed for between 1 and 5 years, there were
abnormal findings. There were no abnormalities in the three
workers with less than one year's exposure.
c) Taiwan press proofing workers
In a cross-sectional survey among Taiwanese press proofers,
using solvents containing n-hexane to clean machinery, workers from
16 factories who had been employed in the industry for more than 2
months underwent detailed neurological examination (Wang et al.,
1986). A total of 59 workers (57 males, and 2 females) with a mean
age of 25.8 (± 10.2) years and a mean employment duration of 5.8
years (range, 2 months to 25 years) was examined. Nerve conduction
velocity studies were undertaken in 54 workers, and a diagnosis of
peripheral neuropathy was made on the basis of objective clinical
signs compatible with peripheral neuropathy and at least two
abnormally slow conduction velocities (45 m/sec in the upper
extremities and 40 m/sec in the lower extremities). Mean nerve
conduction velocities for the exposed group were compared with a
control group of 150 individuals (50 in each of the age ranges
10 -35, 36 - 50, and 51 - 80 years). The sex ratio of the control
group was not provided and exposed and control groups were not
matched for age.
n-Hexane was present in samples of the commercial solvents used
to clean the machines from all the factories at concentrations of
between 3 and 65%. Post-event measurements of n-hexane air
concentrations (14 factories) varied from 0 to 669 mg/m3 (190
ppm). Prior to the discovery of neuropathy, standards of hygiene
were described as poor. Groups of 3 - 9 workers cleaned press
proof machines in small poorly ventilated rooms for more than 8
h/day.
Fifteen cases of overt peripheral neuropathy were diagnosed
among exposed workers (14 males, and 1 female; employment range, 6
months to 5 years; median, 1.5 years). Sural nerve biopsies from
three cases were consistent with n-hexane peripheral neuropathy.
The reported n-hexane air concentrations for two factories where
eight of these people worked were 669 mg/m3 (190 ppm) (six cases)
and 74 mg/m3 (21 ppm) (two cases). No exposure data were available
for the remaining seven cases employed in one factory. Twelve of
the 15 cases regularly slept in the factory and were exposed to
n-hexane for daily periods in excess of 8 h.
Exposure to n-hexane air concentrations of less than 352 mg/m3
(100 ppm) was not associated with clinical peripheral neuropathy.
There were significant reductions in the median, ulnar, and
peroneal nerve mean motor nerve conduction velocities in the
n-hexane-exposed workers without diagnosed peripheral neuropathy,
compared with the control group, but an exposure-related trend was
not found.
In a subsequent neurological investigation of workers at these
factories, there was evidence of central nervous system
malfunction, indicated by somatosensory and visual evoked
responses, in individuals with peripheral neuropathy and also in
other workers (Chang, 1985). Twenty-two of the polyneuropathy
cases, five subclinical cases and seven unaffected workers were
investigated. The absolute and interpeak latencies of patterned
visual evoked potential in affected cases were longer than in
normal controls, and the evoked potential interpeak amplitude was
decreased in individuals with polyneuropathy (Chang, 1987).
Effects on the brainstem auditory evoked response (BAER) were also
reported (Chang, 1985). Wave I latency between factory workers and
normal controls did not differ, but prolongation of the wave I-V
interpeak latencies was noted in workers and corresponded to the
severity of polyneuropathy. Five factory workers exposed to
n-hexane for 5 - 30 months had a reduction in nerve conduction
velocity that was correlated with clinical signs (Huang & Chu,
1989). Neurological symptoms consisted of muscle cramp and
weakness, distal numbness, and, in one case, blurred vision and
tinnitus. The tibial-central conduction in the somatosensory evoked
potentials was delayed and the central conduction time of the BAER
was also prolonged, indicating primary defects in the spinal cord
and brainstem.
d) Tungsten carbide milling
The effect of exposure to n-hexane was investigated in a
factory producing tungsten carbide alloys where powders of tungsten
carbide and other metals and carbides were mixed with n-hexane or
acetone by ball mills (Sanagi et al., 1980). Fourteen workers
(concurrently exposed), aged less than 50 with exposure durations
of 1 - 12 years (mean, 6.2 years), who were engaged in the ball
milling and drying of tungsten carbide alloys were studied. A
further group (previously exposed) of five workers, aged less than
50 with exposure durations of 1 - 16 years (mean, 5.2 years), who
had worked in ball milling in the past were also investigated.
Workers exhibiting any evidence of neurological disorders were
excluded from this study. A concurrent control group of 14 workers
not exposed to n-hexane was selected from the sintering workshop.
The mean 8-h time-weighted average concentration of n-hexane over a
2-year period was 204 mg/m3 (58 ppm). The average concentrations
for acetone were in the range 52 - 164 mg per m3 (22 - 69 ppm).
The medical history of each worker was assessed by interview and
questionnaire. Neurological status was determined by detailed
clinical examination and electrophysiological studies.
Headaches, hyperaesthesia in the limbs, and muscle weakness
tended to be reported more often by workers in the exposed group.
Neurological examination did not reveal any abnormalities of
cranial nerves, the pyramidal tract, or peripheral motor nerves.
Paraesthesia was noted in three exposed subjects and one control
subject. There was a slightly higher prevalence of diminished
bicipital and patellar tendon reflexes in the exposed group. There
were significant effects on muscle strength (jump test on one foot)
and vibration sensation (tuning fork) when mean values for the
exposed groups were compared with the control group.
Electrophysiological measurements showed no signs of neuropathy in
any worker, but in the concurrently exposed group mean maximal
motor conduction velocities in the posterior tibial nerve were
significantly reduced compared with those of the control group. In
addition, the mean distal latency in the posterior tibial nerve was
significantly increased in both the concurrently and previously
exposed groups.
e) Extraction facilities
Two workers at a hexane extraction facility reported transient
paraesthesia following excessive acute exposure to hexane. The
predominant complaint at the facility was of temporary episodes of
light-headedness and dizziness. The maximum time-weighted average
(8 h) hexane concentration at this site was 92 mg/m3 (26 ppm)
(NIOSH, 1981a). The most frequently reported symptoms in workers
associated with a soybean extraction facility were dizziness,
giddiness, and light-headedness (each reported by 74% of
participants). Headache and weakness were also reported by 56 and
52%, respectively, of cases. Operators had a higher incidence of
sleepiness (41%) than did maintenance workers (0%). Exposure
concentrations in personal breathing zones ranged from 15.5 to 46.5
mg/m3 (4.4 - 13.2 ppm), but leaks from some process equipment may
have led to higher levels of acute exposure (NIOSH, 1983).
f) Maizlish study
Workers with long-term exposure to mixtures of organic solvents
have been reported to experience mild, clinically detectable,
sensory or sensorimotor polyneuropathies (Husman & Karli, 1980).
In a study by Maizlish et al. (1987), there was no association
between impaired neurological function and solvent concentration in
mixed exposure to solvents (isopropanol, naphtha, hexane) in
workers occupationally exposed for an average of 6 years. Of 240
workers exposed to mixtures of these solvents, with hexane at
breathing-zone concentrations of 3.5 - 137 mg/m3 (1 - 39 ppm), 16%
had presumed, but not clinically significant, signs of peripheral
neuropathy (diminished pain response, vibratory sensation, and
ankle reflex). The signs were not related to solvent exposure
concentration. Overall, the relative risk (RR = 1.4) of the
presumed signs of peripheral neuropathy was not statistically
significant.
9.3.1.2 Effects on vision and optic nerves
Blurred vision has been reported in some cases of peripheral
neuropathy attributed to n-hexane exposure (Wada & Okamoto, 1965;
Sobue & Yamamura, 1968; Yamamura, 1969). No effects on visual
acuity or on the optic fundus have been reported in case histories
of peripheral neuropathy where information on ophthalmic
examinations was provided (Wada et al., 1965; Yamamura, 1969;
Shirbae et al., 1974). Constriction of the visual field, optic
neuritis, and optic atrophy were reported in one study on Japanese
vinyl sandal makers with peripheral neuropathy (Rizzuto et al.,
1980).
In a study of the ocular effects of n-hexane, 15 workers (11
males, 4 females) making adhesive bandages or extracting vegetable
oils, aged 30 - 65 (mean age, 46 years) with exposure durations of
5 - 21 years (mean exposure, 12 years), were examined
ophthalmologically and given colour discrimination tests and
fluorescein angiography (Raitta et al., 1978). Eight workers were
from adhesive bandage factories where peak n-hexane air
concentrations were 1489, 2126, and 3992 mg/m3 (423, 604, and 1134
ppm) around a glue-spreading machine, 1866 and 4506 mg/m3 (530 and
1280 ppm) at an opened mixing machine, and 11 440 mg/m3 (3250 ppm)
at floor level near a tape machine. Air levels of n-hexane in the
breathing zone of workers were lower than these values and
n-hexane air levels in the factories were generally below 1760
mg/m3 (500 ppm). Seven individuals worked in the extraction of
vegetable oils at a factory where the n-hexane air concentrations
in the processing room were between 35 and 176 mg/m3 (10 and 50
ppm), although air levels as high as 7040 - 10 560 mg/m3 (2000 -
3000 ppm) could be reached when processing was interrupted. There
was no control group.
In three individuals (two of whom had pre-existing defects)
there were disturbances in visual acuity or field of vision.
Colour vision defects, mainly blue-yellow, were found in 12
workers. There were no cases of optic atrophy. In nine workers
the maculae showed slight pigment dystrophy, in three there were
yellowish dots, and in one the foveal reflex was absent.
Fluorescein angiography showed patchy choroidal underfilling,
capillary widening of the perifoveal arcade, and loss of retinal
pigment around the macula. Colour discrimination defects and
maculopathy appeared to be related.
In a subsequent study, visual evoked potentials and
electroretinograms (ERGs) were recorded in the same 15 workers and
compared with control measurements obtained from 10 healthy
subjects aged 21 - 57 (mean age, 34.9 years) who were not
occupationally exposed to solvents (Seppalainen et al., 1979).
There were significant reductions in peak-to-peak amplitudes for a
number of visual evoked potential components, possibly due to
conduction block in intracerebral axons. The latency of several
components was significantly increased. This was attributed to
partial axonal degeneration in the visual pathways. A significant
decrease in the amplitude of the electroretinogram (ERG) was
reported, with a reduced latency in the "b" wave. The effects on
the ERG in the exposed workers could not be correlated directly
with the effects on retinal pigments and colour discrimination
reported by Raitta et al. (1978). Seppalainen & Raitta (1981)
found electroneuromyographic signs of peripheral neuropathy,
including reduced nerve conduction velocity and/or fibrillation or
long duration motor unit potentials, in 11 of the 15 workers.
Symptoms included "drunkenness" (8), headache (7), tiredness (5),
and paraesthesia in the limbs (5). All workers with paraesthesia
had at least one electrophysiological sign of neuropathy. Abnormal
EEG findings (slow wave abnormalities) were found in 9 of the
workers.
Signs of maculopathy and colour discrimination defects were
noted in 10 out of the 11 workers with abnormal electrophysiological
signs or peripheral neuropathy. Colour discrimination defects were
present in 8 of the 9 workers with EEG abnormalities. There were
altered EMG and EEG traces in 6 of the 12 workers with colour
discrimination defects. The authors suggested that there was a
correlation between maculopathy and peripheral neuropathy.
Abnormal visual evoked potentials were reported in a study of
glue sniffers exposed to n-hexane (Cooper et al., 1985). Abnormal
electrophysiological signs persisted in two individuals who were
retested 6 months after they had stopped inhaling solvents.
However, these individuals had been exposed to a wide range of
organic solvents and exposure information was inadequate.
In somatosensory evoked potentials, both the absolute latencies
and central conduction time were prolonged in affected workers
(Chang, 1987). In a study of 25 patients, 5 subclinical cases, and
45 apparently healthy workers, Chang & Yip (1987) reported various
electrophysiological abnormalities that reflected, in
polyneuropathy patients, pathological changes of primary giant
axonal degeneration with secondary myelinopathy. This was based on
the observation of a marked fall in nerve conduction velocities,
profound amplitude reduction of muscle action potentials and
sensory action potentials and, prolongation of latencies. Early
electroneurographic changes in the apparently healthy individuals
exposed to n-hexane were reduction in the nerve conduction
velocities, prolongation of motor distal latency, and attenuation
of sensory action potential amplitude.
9.3.2. Effects on the kidney
An investigation of kidney function was conducted on a group of
64 workers (age and sex not reported) exposed for 1 - 24 years to
hydrocarbon mixtures at a shoe factory and an undefined control
group of 80 subjects (Nakajima & Murayama, 1985). Mean air
concentrations for solvents in the factory over a period of one
year were: n-hexane, 250 mg/m3 (71 ppm); cyclohexane, 203 mg/m3
(59 ppm); methyl ethyl ketone, 44 mg/m3 (15 ppm); and ethyl
acetate, 180 mg/m3 (50 ppm). No effects on mean total urinary
protein, albumin, beta-glucuronidase, or muramidase levels were
reported.
A study of kidney function was also undertaken on a group of
182 Italian shoe workers selected from 185 workers employed at four
factories (59 males, 123 females; mean age, 39.4 years; mean
exposure duration, 10.2 years) (Mutti et al., 1981; Franchini et
al., 1983). Breathing-zone concentrations were estimated for a
number of solvents (hexane, ethyl acetate, cyclohexane, methyl
ethyl ketone, and acetone), and concentrations of hexane ( n-hexane
and 2- and 3-methylpentane) were reported to be significantly
higher than 100 ppm (352 mg n-hexane per m3) on a number of
occasions. Eighty healthy subjects (30 males, 50 females; mean
age, 37.9 years) without a history of renal disease or exposure to
organic solvents were used as a control group. A further control
group of 30 unexposed workers (10 males, 20 females; mean age, 41.6
years) was randomly selected from 124 workers who had left the
factories during the previous 5 years (mean exposure duration, 8.8
years).
Spot samples of urine were analysed for total protein, albumin,
and creatinine levels and for beta-glucuronidase activity. Urinary
lysozyme activity was estimated in a few workers. Blood samples
were analysed for creatinine levels. The mean total urinary
protein level was significantly higher in the exposed workers than
in either control or historically exposed workers. Seven workers
(4 males, 3 females) had abnormally high levels (greater than two
standard deviations of the mean control value). Four of these
workers also had abnormally high urinary lysozyme activity, and two
had increased beta-glucuronidase activity. There were no effects
on urinary albumin or serum creatinine levels.
9.3.3. Effects on other organs
Health surveillance studies were carried out in Japan following
a mass outbreak of polyneuropathy in 1967 (Sobue & Yamamura, 1968,
Iida et al., 1969, Yamamura, 1969; Inoue et al., 1970, 1971; Iida &
Yamamoto, 1973).
No effects on the respiratory system, kidney, liver, or
haematopoietic organs were reported.
Laboratory examinations were inconsistent. Urinalysis showed
positive urobilinogen in 15 cases and raised coproporphyrin levels
in four cases. The cephalin cholesterol flocculation test was
positive in 9 cases and a slight to moderately raised serum lactate
dehydrogenase level was found in 17 cases. Depressed cholinesterase
levels were reported in 26 out of 42 cases examined. There was no
correlation between the depressed cholinesterase levels and severity
of neuropathy. The cerebrospinal fluid was examined in 7 cases and
was normal, with no significant effect on pressure or protein
levels. Plethysmography was performed in 11 cases and weak
vasomotor responses were found in two cases.
10. EVALUATION OF HUMAN HEALTH RISKS AND EFFECTS ON THE ENVIRONMENT
10.1. Animal Studies
10.1.1. Short- and long-term exposure
n-Hexane is of low acute toxicity, LD50 values in the range
15 - 30 g/kg and an LC50 (1 h) of 271 040 mg per m3 having been
reported.
In essentially continuous inhalation studies in rats, deaths
occurred following n-hexane exposure to 3520 mg per m3 (1000 ppm)
for up to 11 weeks, and reduced body weight gain after 3 weeks of
exposure to 1760 mg/m3 (500 ppm). In studies involving daily 12-h
exposures to n-hexane, deaths were recorded at 10 560 mg/m3 (3000
ppm) after 15 weeks and slightly reduced body weight gain at 740 mg
per m3 (200 ppm) after 24 weeks. No effects on body weight were
noted in rats exposed to 443 mg/m3 (126 ppm) virtually continuously
for 34 weeks.
Testicular toxicity and peripheral and central neurotoxicity
appear to be the principal non-lethal effects of repeated n-hexane
exposure in rats. Severe testicular lesions have resulted from
inhalation exposure to n-hexane and oral exposure to
2,5-hexanedione. Effects have been attributed to disruption of the
cytoskeleton of Sertoli cells. There are secondary effects on
post-spermatogonial germ cells, which disappear from affected
tubules. Testicular effects were reversible after a single
exposure for 24 h to 17 600 mg/m3 (5000 ppm) but irreversible after
a 2-weeks exposure to the same concentration for 16 h/day, 6
days/week. 2,5-Hexanedione at 1% in drinking water produced
reversible testicular lesions after 2 to 3 weeks of dosing and
irreversible effects (within 17 weeks) after 5 weeks of dosing.
Clinically, neurotoxicity is characterized by hindlimb
weakness, which can progress to paralysis and eventually to
quadriplegia. Clinical signs have been seen after 10 weeks of
discontinuous exposure to 10 560 mg/m3 (3000 ppm) (12 h/day, 7
days/week) and after 6 - 10 weeks of continuous exposure to 1408 to
2112 mg/m3 (400 to 600 ppm).
Lesions (axonal swellings) in hindlimb nerves, brain, and
spinal cord have been observed prior to the appearance of hindlimb
weakness. More severe effects (axonal degeneration and loss) were
noted, particularly in the longest nerves with the largest
diameter, after 6 months of exposure to 1760 mg/m3 (500 ppm).
Overall, no-observed-adverse-effect levels for n-hexane-induced
neurotoxicity in rats are difficult to establish because of the
range of vapour concentrations and exposure regimes tested, and the
variety of effects investigated. In essentially continuous 6-month
inhalation studies, nerve lesions were seen at n-hexane
concentrations of 1760 mg per m3 (500 ppm) or more, but no clinical
or pathological effects were noted at 440 mg/m3 (125 ppm).
Discontinuous exposure of rats to 3168 mg/m3 (900 ppm) for 72 weeks
did not cause any apparent nerve lesions, but there was some
preliminary evidence for an effect on grip strength.
Resolution (partial or complete) of clinical signs of toxicity,
reduced grip strength, and electrophysiological effects can occur
following cessation of exposure. Only limited recovery of
amplitude of the fifth component of the brainstem auditory-evoked
response component (believed to reflect central nervous system
activity) and tail nerve action potential was recorded 15 to 22
weeks after the end of repeated exposure to 3520 mg/m3 (1000 ppm)
for 11 weeks. Resolution of pathological lesions has not been
systematically investigated following inhalation exposure, but
remyelination and limited axonal regeneration have been observed
following cessation of exposure to 1408 to 2112 mg/m3 (400 to 600
ppm).
Inhalation of 1760 mg/m3 (500 ppm) discontinuously for 7 weeks
can cause minor damage to the lungs of rats. In rabbits, severe
respiratory tract lesions were seen following exposure to 10 560
mg/m3 (3000 ppm), 8 h/day for 8 days, and papillary proliferations
of non-ciliated bronchiolar cells were noted after 24 weeks of
exposure.
Effects on the kidney and liver have been reported in a few
inhalation studies on rats but only at higher exposure levels.
Most lesions and other effects were considered to be of minor
toxicological significance and/or were not reproducible in repeated
studies. However, severe liver lesions have been noted in rats
following repeated intraperitoneal administration of n-hexane.
Combined exposure studies have shown that the neurotoxic
effects of n-hexane vapour can be enhanced in hens by methyl
isobutyl ketone (MIBK) and in rats by both methyl ethyl ketone and
lead acetate, but decreased by toluene. Pulmonary lesions in rats
have also been reported to be enhanced by co-exposure to methyl
ethyl ketone.
There have been few studies on the effects of oral or dermal
exposure to n-hexane. Daily oral administration of 500 mg/kg per
day for 4 weeks resulted in reduced tail nerve conduction velocity
in rats.
10.1.2. Genotoxicity
Only limited mutagenicity testing has been conducted.
Chromosomal damage (polyploidy in one study, structural aberrations
in a second study) was reported in in vitro assays in which a
Chinese hamster cell line was exposed to undiluted liquid n-hexane
in the absence of a metabolizing fraction. No significant increase
in point mutation rate occurred in cultured mammalian cells or
bacteria following exposure to n-hexane. DNA damage, as measured
in differential bacterial toxicity assays, was not increased by
exposure to n-hexane. Similarly, no increase in unscheduled DNA
synthesis was noted in human lymphocytes in vitro. Although
n-hexane has given negative results in assays for point mutations
there is some evidence that it can produce chromosomal aberrations
in vitro. Chromosomal damage (chromatid breaks) in rat bone marrow
cells was reported in an in vivo study.
10.1.3. Carcinogenicity
The carcinogenic potential of n-hexane has not been adequately
studied. In a limited study, dermally applied n-hexane did not
appear to enhance the formation of skin tumours in mice following
exposure to a tumour initiator (7,12-dimethylbenzanthracene) or
co-exposure with a promoting agent (croton oil).
10.1.4. Reproductive effects
The reproductive toxicity of n-hexane has not been adequately
investigated. There is no substantial evidence of embryotoxicity
or teratogenicity in rats following inhalation or in mice after
oral dosing. Slight increases in the incidence of some anomalies
were noted in rats exposed to 10 560 mg/m3 (3000 ppm), but
inadequacies in the conduct of the study (e.g., the concentration
tested was probably not high enough) limit the conclusions that can
be reached. Signs of fetotoxicity (reduced pup weight) were noted
in mice, but only at doses that resulted in significant maternal
mortality. A transient delay in postnatal development was reported
in one study in the pups of rats exposed to 10 560 mg/m3 (3000
ppm). Although no signs of neuropathy were reported in these
animals, it is not possible to assess from this study whether in
utero exposure could induce neuropathy, as the animals were only
exposed to n-hexane during a small part of the gestation period.
Other reports have claimed that in utero exposure to n-hexane
retards cerebellar histogenesis and neuronal maturation and induces
electrophysiological abnormalities of the central nervous system in
neonatal rats. However, this work has not been reported in
sufficient detail for a full assessment to be made.
There is evidence that n-hexane causes irreversible sterility
in the rat. No effects on male fertility were reported in a
limited dominant lethal study on the mouse. No investigations of
fertility in female animals or multi-generation studies have been
reported.
10.2. Human Studies
10.2.1. Effects on the nervous system
Many studies have linked occupational n-hexane exposure to the
incidence of peripheral neuropathy in humans. However, few of
these studies reported air concentrations, those reported may be
inaccurate, and it is often not clear whether these concentrations
are of hexane or n-hexane. In addition, industrial exposure to
n-hexane usually occurs during the use of solvent-based products,
so workers are exposed to a mixture of volatile compounds (e.g.,
ethyl acetate, methyl ethyl ketone, toluene, acetone, and other
aliphatic hydrocarbons). Air concentrations for the other solvents
present have rarely been documented. Only those studies that
present some air concentration data have been included in this
review.
The results of peripheral nerve electrophysiology may vary
considerably depending on the type of technique used, the ambient
temperature at which measurements are taken, the segment of the
nerve studied, and the age of the individual. Thus, the assessment
of studies that rely on peripheral nerve electrophysiology is
difficult in the absence of information on these factors.
Studies on 93 cases of neuropathy characterized the clinical
features of n-hexane neuropathy and showed that electro-
physiological measurements correlated well with the severity of the
condition. An association between increasing severity of the
disease and increasing exposure was found. However, from the
limited details of exposure reported, it was not possible to
establish any definite relationship.
Mild subclinical signs of neuropathy were found in shoe workers
exposed to 70 mg/m3 (20 ppm). The degree of electrophysiological
abnormalities increased with duration of exposure. However, no
cases of overt neuropathy occurred, even in workers with an
exposure duration longer than 5 years.
Subjective and electrophysiological signs of subclinical
peripheral neuropathy were found in a group of workers exposed to
approximately 317 mg/m3 (90 ppm). However, these effects cannot be
wholly ascribed to exposure to this level of n-hexane, since
previous exposure to solvents was probably underestimated and
worker self-selection may have introduced bias. In addition, there
may have been co-exposure to methyl ethyl ketone, which has been
demonstrated to enhance the neurotoxic effects of n-hexane.
Exposure to a solvent mixture containing an n-hexane level of
approximately 690 mg/m3 (196 ppm) resulted in reduced nerve
conduction velocity along peripheral nerves, which persisted after
exposure had been significantly reduced. In addition, an effect of
n-hexane on the central nervous system has been suggested to be the
cause of some of the somatosensory evoked potential changes noted.
Subjective and electrophysiological signs of mild subclinical
neuropathy occurred in a small group of workers exposed to mean
n-hexane concentrations in air of 208 mg/m3 (59 ppm). Clinical
overt neuropathy was not detected. In this study exposure to other
solvents was limited and the nerve conduction studies were
performed under defined conditions. However, the small number of
subjects studied, lack of historical exposure data, and the poor
matching of the exposed and control groups limit the value of the
conclusions reached. Nevertheless, the results are in general
agreement with the data obtained in the earlier studies reported
above. Exposure to 669 mg/m3 (190 ppm) for periods in excess of 8
h/day was associated with the onset of clinically overt peripheral
neuropathy. Partial recovery of nerve function can occur slowly
over several years. Exposure to less than 352 mg/m3 (100 ppm) did
not result in neuropathy but showed electrophysiological evidence
of subclinical effects.
Colour discrimination defects and signs of maculopathy have
been described in a small number of workers occupationally exposed
to a wide range of n-hexane levels in air. Visual-evoked potential
and electroretinogram measurements of these individuals showed
abnormalities compared with a control group of non-exposed workers.
Abnormal visual-evoked potential traces have also been reported in
some glue sniffers exposed to n-hexane. It has been suggested that
axonal degeneration may occur in the visual pathway and that
n-hexane may induce a central nerve conduction block. There is
also some evidence that signs of maculopathy may be correlated with
electrophysiological signs of peripheral neuropathy. Abnormal EEG
traces were also found in some of the individuals studied.
10.2.2. Other effects
No studies have been reported on fertility or possible
testicular effects in males. Very little information is available
on the acute toxicity of n-hexane to humans. Most studies have
involved exposure to solvent mixtures. The available data suggest
that n-hexane is of low acute toxicity.
Very few studies of skin or eye irritancy involving volunteers
have been published. n-Hexane is a mild irritant causing transient
erythema when in contact with human skin for short periods. More
severe effects (erythema and blistering) were documented after
occlusive skin contact for 5 h to commercial grade hexane.
Insufficient data are available to assess whether the liquid can
cause eye irritation in human beings. No signs of eye irritation
were noted in a study in which volunteers were exposed to a hexane
vapour concentration of 1760 mg/m3 (500 ppm) for 3 - 5 min. There
have been no reports of skin sensitization in exposed workers and
no skin sensitization was noted in a maximization test with
n-hexane.
10.3. Environmental Effects
n-Hexane has low solubility in water and is highly volatile.
Exposure of aquatic organisms is, therefore, likely to be low and
transient except for uncontrolled discharges into surface waters.
Data on the toxicity of n-hexane to aquatic organisms are few,
variable, conducted under inappropriate conditions and inadequate
to make an assessment of this environmental risk.
11. RECOMMENDATIONS
11.1. Human Health Protection
At the levels of n-hexane to which the general population is
exposed, there is unlikely to be any hazard. In the occupational
health context, atmospheric levels should be kept below the
recommended occupational exposure limits by suitably designed work
processes and engineering controls including ventilation. Suitable
protective clothing and respiratory protection should be readily
available for use in enclosed spaces, in emergencies, and for
certain maintenance operations. The use of a supplied-air
respirator or a self-contained breathing apparatus in continuous
flow mode for n-hexane concentrations up to 8800 mg/m3 (2500 ppm)
is recommended. It should be fitted with a full facepiece for
concentrations between 8800 mg/m3 (2500 ppm) and 17 600 mg/m3 (5000
ppm). Although protective gloves may resist penetration by
n-hexane, mixtures of n-hexane with other solvents, such as methyl
ethyl ketone, may permit the n-hexane to penetrate gloves and other
protective clothing. This factor should be considered when using
n-hexane mixtures.
11.2. Environmental Protection
n-Hexane is unlikely to present a hazard to the environment,
except in the case of major spills or discharges where there could
be transient local effects.
12. FURTHER RESEARCH
a) A study of indices of testicular function should be made in an
occupationally exposed group(s) to establish whether any or all
of the effects reported to occur in the testes of animals are
also manifested in man.
b) The genotoxic effects of n-hexane should be further
investigated in vivo in order to supplement the limited in
vitro information on genotoxicity.
c) A repeat dose study is required to establish the toxicity of
n-hexane given by the oral route. This is needed to provide a
NOEL in order to establish an ADI.
d) The mechanism of neurotoxicity at the molecular level is not
clear. Understanding the mechanism will contribute directly to
the validity of risk assessment. In vivo and in vitro studies
are required to investigate further and clarify these mechanisms.
e) Available teratogenicity studies are inadequate for an
evaluation of risk. Studies should be undertaken to provide
usable data.
f) Dose-response relationships concerning neurotoxic effects of
n-hexane in humans are unclear because of incomplete or
unreliable exposure data. Moreover, although peripheral
neurotoxicity by n-hexane has been amply demonstrated, effects
on the central nervous system have been explored only to a
limited extent. Further epidemiological investigations such as
prospective follow-up studies should be undertaken in order to
overcome the shortcomings of previous investigations.
REFERENCES
ABBRITTI, G., SIRACUSA, A., CIANCHETTI, C., COLI, C.A., CURRADI,
F., PERTICONI, G.F., & DE ROSA, F. (1976) Shoe makers'
polyneuropathy in Italy: the aetiological problem. Br. J. ind.
Med., 33: 92-99.
ABDEL-RAHMAN, M.S., HETLAND, L.B., & COURI, D. (1976) Toxicity and
metabolism of methyl n-butyl ketone. Am. Ind. Hyg. Assoc. J., 37:
95-102.
ABE, K., MISUMI, J., KAWAKAMI, M., & NOMURA, S. (1980) Effects of
n-hexane, methyl n-butyl ketone, and 2,5-hexanedione on the
excitability of sweat glands in rats to mecholyl. Jpn. J. ind.
Health, 22: 380-381.
ABOU-DONIA, M.B., LAPADULA, D.M., CAMPBELL, G., & TIMMONS, P.R.
(1985a) The synergism of n-hexane-induced neurotoxicity by methyl
isobutyl ketone following subchronic (90 days) inhalation in hens:
Induction of hepatic microsomal cytochrome P-450. Toxicol. appl.
Pharmacol., 81: 1-16.
ABOU-DONIA, M.B., MAKKAWY, H.M., & CAMPBELL, G.M. (1985b) Pattern
of neurotoxicity of n-hexane, methyl n-butyl ketone,
2,5-hexanediol, and 2,5-hexanedione alone and in combination with
0-ethyl 0-4-nitrophenyl phenylphosphonothioate in hens. J. Toxicol.
environ. Health, 16: 85-100.
ABOU-DONIA, M.B., LAPADULA, D.M., & SUWITA, E. (1988) Cytoskeletal
proteins as targets for organophosphorus compound and aliphatic
hexacarbon-induced neurotoxicity. Toxicology, 49: 469-477.
ACGIH (1986) Documentation of the threshold limit values, 5th ed.,
Cincinnati, Ohio, American Conference of Governmental Industrial
Hygienists.
AHONEN, I. & SCHIMBERG, R.W. (1988) 2,5-hexanedione excretion after
occupational exposure to n-hexane. Br. J. ind. Med., 45: 133-136.
AIELLO, I., ROSATI, G., SERRA, G., & MANCA, M. (1980) Subclinical
neuropathic disorders and precautionary measures in the shoe
industry. An electrophysiological investigation. Acta neurol.
(Napoli), 35: 285-292.
ALTENKIRCH, H. & SCHULTZ, H. (1979) [Glue sniffing and neuropathy.
Neurological findings and social data from 40 cases.] Nervenarzt,
50: 21-27 (in German).
ALTENKIRCH, H., MAGER, J., STOLTENBURG, G., & HELMBRECHT, J. (1977)
Toxic polyneuropathies after sniffing a glue thinner. J. Neurol.,
214: 137-152.
ALTENKIRCH, H., STOLTENBURG-DIDINGER, G., & WAGNER, H.M. (1978)
Experimental studies on hydrocarbon neuropathies induced by methyl
ethyl ketone (MEK). J. Neurol., 219: 159-170.
ALTENKIRCH, H., STOLTENBURG-DIDINGER, G., & WAGNER, H.M. (1979)
Experimental data on the neurotoxicity of methyl ethyl ketone
(MEK). Experientia (Basel), 35: 503-504.
ALTENKIRCH, H., WAGNER, H.M., STOLTENBURG, G., & SPENCER, P.S.
(1982) Nervous system responses of rats to subchronic inhalation of
n-hexane and n-hexane plus methyl ethyl ketone mixtures. J. neurol.
Sci., 57: 209-219.
ANDERSON, R.J. & DUNHAM, C.B. (1984) Electrophysiologic deficits in
peripheral nerve as a discriminator of early hexacarbon
neurotoxicity. J. Toxicol. environ. Health, 13: 835-843.
ANGELO, M.J. (1977) The pharmacokinetics of the neurotoxin
2,5-hexanedione: distribution, elimination and model simulations,
Newark, University of Delaware (Ph.D Thesis).
ANGELO, M.J. & BISCHOFF, K.B. (1982) A physiologically based
pharmacokinetic model for 2,5-hexanedione. In: Proceedings of the
12th Conference on Environmental Toxicology, 1981, Dayton, Ohio,
Aerospace Medical Research Laboratory, pp. 250-286 (Paper No. 16)
(AFAMRL-TR-81-149).
ANTHONY, D.C., BOEKELHEIDE, K., & GRAHAM, D.G. (1983) The effect of
3,4-dimethyl substitution on the neurotoxicity of 2,5-hexanedione.
I. Accelerated clinical neuropathy is accompanied by more proximal
axonal swellings. Toxicol. appl. Pharmacol., 71: 362-371.
API (1978) 26 week inhalation toxicity study of n-hexane in the
rat, Washington, DC, American Petroleum Institute (API Medical
Research Report No. 28-30077).
API (1983a) Six month continuous inhalation exposures of rats to
hexane mixtures - phase I, Washington, DC, American Petroleum
Institute (API Medical Research Publication No. 30-32858).
API (1983b) Six month continuous inhalation exposures of rats to
hexane mixtures - phase II, Washington, DC, American Petroleum
Institute (API Medical Research Publication No. 30-32846).
ATKINSON, R., ASCHMANN, S.M., CARTER, W.P.L., WINER, A.M., & PITTS,
J.N., Jr (1982) Kinetics of the reactions of OH radicals with
n-alkanes at 299 ± 2 K. Int. J. chem. Kinet., 14: 781-788.
BABANOV, G.P. & BABANOV, A.G. (1981) [Permeability of tissues to
alkanes and their relationship with molecular weight, parity of
carbon atoms and branching of hydrocarbon chains.] Gig. Tr. prof.
Zabol., 12: 48-51 (in Russian).
BABANOV, G.P., ABRAMYAN, G.G., ARTEMYEVA, G.N., VERKHOVSKI, L.G., &
SHAINA, V.K. (1988) [Elevated temperature effect on animals'
sensitivity to n-pentane, n-hexane, n-heptane, n-octane and toluene
and their distribution in the tissues.] Gig. Tr. prof. Zabol., 10:
23-25 (in Russian).
BABICH, H., DAVIS, D.L., & ADLER, R. (1982) Updating federal
standards for toxicants: n-hexane as the model. Environ. Monit.
Assess., 2: 287-299.
BAKER, T.S. & RICKERT, D.E. (1981) Dose-dependent uptake,
distribution and elimination of inhaled n-hexane in the Fischer-344
rat. Toxicol. appl. Pharmacol., 61: 414-422.
BAKER, T.S., WHITE, E.L., & RICKERT, D.E. (1980) Differences in the
metabolism of n-hexane in Fischer 344 rats and C57B1 mice.
Pharmacologist, 22: 279.
BARLOTTA, F., MALFITANO, D., GORGONE, G., & INSERRA, A. (1972)
[Electroretinographic study in experimental hexane intoxication.]
Boll. Soc. Ital. Biol. Sper., 48: 115-117 (in Italian).
BARNI-COMPARINI, I., LUNGARELLA, G., & FONZI, L. (1982) Responses
of lung enzyme activities in rabbits following short-term exposure
to n-hexane: correlation between morphological and biochemical
changes. Agents Actions, 12: 737-742.
BARTOLUCCI, G.B., MAZZOTTA, M., MANNO, M., & DEROSA, E. (1977)
[Industrial exposure to TOCP and n-hexane: behaviour of serum
cholinesterase.] Med. Lav., 68: 222-231 (in Italian).
BASTONE, A., FRONTALI, N., MALLOZZI, C., SBRACCIA, M., & SETTIMI,
L. (1987) Cholinesterases in blood plasma and tissues of rats
treated with n-hexane or with its neurotoxic metabolite
2,4-hexanedione. Arch. Toxicol., 61: 138-144.
BATTISTINI, N., LENZI, G.L., ZANETTE, E., FIESCHI, C., BATTISTA,
F., FRANZINELLI, A., & SARTORELLI, E. (1974) [Observations of a
group of cases of glue polyneuropathy (Clinical electromyographic
and experimental contribution).] Riv. Patol. nerv. ment., 95:
871-885 (in Italian).
BATTISTINI, N., CIONI, R., LENZI, G.L., & ZANETTE, E., (1975)
[Study of the H-reflex in glue neuropathy.] Riv. Neurol., 45: 74-79
(in Italian).
BEHNKE, W., HOLLANDER, W., KOCH, W., NOLTING, F., & ZETZSCH, C.
(1988) A smog chamber for studies of the photochemical degradation
of chemicals in the presence of aerosols. Atmos. Environ., 22:
1113-1120.
BERGER, M.L. & SOZERI, T. (1987) Rapid halogenated hydrocarbon
toxicity in isolated hepatocytes is mediated by direct solvent
effects. Toxicology, 45: 319-330.
BERNARD, A.M., DE RUSSIS, R., NORMAND, J.-C., & LAUWERYS, R.R.
(1989) Evaluation of the subacute nephrotoxicity of cyclohexane and
other industrial solvents in the female Sprague-Dawley rat.
Toxicol. Lett., 45: 271-280.
BOBRA, A.M., WAN, Y.S., & MACKAY, D. (1983) A predictive
correlation for the acute toxicity of hydrocarbons and chlorinated
hydrocarbons to the water flea (Daphnia magna). Chemosphere, 12:
1121-1129.
BOCK, F.G. & CHORTYK, O.T. (1970) Tumor promoting activity of
aliphatic hydrocarbons. Proc. Am. Assoc. Cancer Res., 11: 9.
BOEKELHEIDE, K. (1987) 2,5-Hexanedione alters microtubule assembly.
1. Testicular atrophy, not nervous system toxicity, correlates with
enhanced tubulin polymerization. Toxicol. appl. Pharmacol., 88:
379-382.
BOEKELHEIDE, K. (1988a) Rat testis during 2,5-hexanedione
intoxication and recovery. I. Dose response and the reversibility
of germ cell loss. Toxicol. appl. Pharmacol., 92: 18-27.
BOEKELHEIDE, K. (1988b) Rat testis during 2,5-hexanedione
intoxication and recovery. II. Dynamics of pyrrole reactivity,
tubulin content and microtubule assembly. Toxicol. appl.
Pharmacol., 92: 28-33.
BOEKELHEIDE, K. & EVELETH, J. (1988) The rate of 2,5-hexanedione
intoxication, not total dose, determines the extent of testicular
injury and altered microtubule assembly in the rat. Toxicol. appl.
Pharmacol., 94: 76-83.
BOHLEN, P., SCHLUNEGGER, U.P., & LAUPPI, E. (1973) Uptake and
distribution of hexane in rat tissues. Toxicol. appl. Pharmacol.,
25: 242-249.
BORGATTI, A.R., TRIGARI, G., VENTRELLA, V., & PAGLIARANI, A. (1981)
[Interactions of n-alkanes with respiration and oxidative
phosphorylation of rabbit cardiac mitochondria: n-hexane.] Boll.
Soc. Ital. Biol. Sper., 56: 1569-1575 (in Italian).
BRAIER, L. (1973) A comparative study of isocyclic hydrocarbons in
animals and in man. Haemalogica, 58: 491-500.
BRAVACCIO, F., AMMENDOLA, A., BARRUFFO, L., & CARLOMANGO, S. (1981)
H-reflex behaviour in glue ( n-hexane) neuropathy. Clin. Toxicol.,
18: 1369-1375.
BRINGMANN, G. & KUHN, R. (1982) Results of toxic action of water
pollutants on Daphnia magna tested by an improved standardized
procedure. Z. Wasser Abwasser Forsch., 15: 1-6.
BRUGNONE, F. & PERBELLINI, L. (1980) [Pollution by solvents and
workers' positions in shoe and shoe upper factories.] Med. Lav.,
71: 343-352 (in Italian).
BRUGNONE, F., PERBELLINI, L., GRIGOLINI, L., & APOSTOLI, P. (1978)
Solvent exposure in a shoe upper factory. I. n-Hexane and acetone
concentrations in alveolar and environmental air and in blood. Int.
Arch. occup. environ. Health, 42: 51-62.
BRUGNONE, F., PERBELLINI, L., GAFFURI, E., & APOSTOLI, P. (1980)
Biomonitoring of industrial solvent exposures in workers' alveolar
air. Int. Arch. occup. environ. Health, 47: 245-261.
BRUGNONE, F., PERBELLINI, L., & APOSTOLI, P. (1984) Blood
concentration of solvents in industrial exposure. Collect. Méd.
lég. Toxicol. méd., 125(Part 1): 165-168.
BUITATTI, E., CECCHINI, S., RONCHI, O., DOLARA, P., & BULGARELLI,
G. (1978) Relationship between clinical and electromyographic
findings and exposure to solvents, in shoe and leather workers. Br.
J. ind. Med., 35: 168-173.
BUS, J.S. & TYL, R.W. (1979) Perinatal toxicity of n-hexane in
Fischer 344 rats. Teratology, 19: 22A.
BUS, J.S., WHITE, E.L., TYL, R.W., & BARROW, C.S. (1979) Perinatal
toxicity and metabolism of n-hexane in Fischer-344 rats after
inhalation exposure during gestation. Toxicol. appl. Pharmacol.,
51: 295-302.
BUS, J.S., WHITE, E.L., GILLIES, P.J., & BARROW, C.S. (1981) Tissue
distribution of n-hexane, methyl n-butyl ketone and 2,5-hexanedione
in rats after single or repeated inhalation exposure to n-hexane.
Drug Metab. Dispos., 9: 386-387.
BUS, J.S., DEYO, D., & COX, M. (1982) Dose-dependent disposition of
n-hexane in F-344 rats after inhalation exposure. Fundam. appl.
Toxicol., 2: 226-229.
CCOHS (1985) n-Hexane, Hamilton, Ontario, Canadian Centre for
Occupational Health and Safety (Chemical Hazard Summary No. 11).
CARAPELLA, C. (1977) [Preventive aspects of toxic polyneuropathy in
shoemakers.] Ann. Ist. Super. Sanità, 13: 353-366 (in Italian).
CARLOMAGNO, S., D'ANGELI, A., & MENEGOZZO, M. (1983) [ n-Hexane
polyneuropathy in the shoe making and leather industries.] Med.
Lav., 74: 111-122 (in Italian).
CAVANAGH, J.B. (1982) The pattern of recovery of axons in the
nervous system of rats following 2,5-hexanediol intoxication. A
question of rheology? Neuropathol. appl. Neurobiol., 8: 19-34.
CAVANAGH, J.B. & BENNETTS, R.J. (1981) On the pattern of changes in
the nervous system produced by 2,5-hexanediol. A topographical
study by light microscopy. Brain, 104: 297-318.
CAVENDER, F.L., CASEY, H.W., SALEM, H., GRAHAM, D.G., SWENBERG, J.,
& GRALLA, E.J. (1984) A 13-week vapour inhalation study of n-hexane
in rats with emphasis on neurotoxic effects. Fundam. appl.
Toxicol., 4: 191-201.
CEC (1979) Analysis of organic micropollutants in water, 3rd ed.,
Luxembourg, Commission of the European Communities, Vol. 1, p. 155
(COST 646 bis).
CHANG, Y.C. (1985) Evoked potential study in n-hexane
polyneuropathy. J. Neurol., 22(Suppl.): 200.
CHANG, Y.C. (1987) Neurotoxic effects of n-hexane on the human
central nervous system: evoked potential abnormalities in n-hexane
polyneuropathy. J. Neurol. Neurosurg. Psychiatry, 50: 269-274.
CHANG, Y.C. & YIP, P.K. (1987) n-Hexane-induced electroneurographic
changes and early detection of n-hexane intoxication. J. Formosan
Med. Assoc., 86: 194-200.
CHAPIN, R.E., NORTON, R.M., POPP, J.A., & BUS, J.S. (1982) The
effects of 2,5-hexanedione on reproductive hormones and testicular
enzyme activities in the F-344 rat. Toxicol. appl. Pharmacol., 62:
262-272.
CHAPIN, R.E., MORGAN, K.T., & BUS, J.S. (1983) The morphogenesis of
testicular degeneration induced in rats by orally administered
2,5-hexanedione. Exp. mol. Pathol., 38: 149-169.
CHAUPLANNAZ, G., BADY, B., KOPP, N., LEVRAT, R., & TRILLET, M.
(1982) Neuropathie périphérique au n-hexane chez une toxicomane.
Rev. neurol. (Paris), 138(3): 249-257.
CHIOU, C.T., KILE, D.E., & MALCOLM, R.L. (1988) Sorption of vapours
of some organic liquids on soil humic acid and its relation to
partitioning of organic compounds in soil organic matter. Environ.
Sci. Technol., 22: 298-303.
CIANCHETTI, C., ABBRITTI, G. PERTICONI, G., SIRACUSA, A., &
CURRADI, F. (1976) Toxic polyneuropathy of shoe industry workers.
J. Neurol. Neurosurg. Psychiatry, 39: 1151-1161.
CIIT (1977) n-Hexane, Research Triangle Park, North Carolina,
Chemical Industry Institute of Toxicology (CIIT Current Status
Reports, No. 1).
CLAYTON, G.D. & CLAYTON, F.E. (1981) Patty's industrial hygiene and
toxicology, New York, Chichester, Brisbane, Toronto, John Wiley and
Sons, Vol. 2B.
CFR (1987a) Food additives permitted for direct addition to food
for human consumption. Code fed. Regul., 21(Part 172): 172.560,
172.894.
CFR (1987b) Listing of color additives exempt from certification:
Subpart B - Drugs. Code fed. Regul., 21(Part 73): 256.
COOPER, R., NEWTON, P., & REED, M. (1985) Neurophysiological signs
of brain damage due to glue sniffing. Electroencephalogr. clin.
Neurophysiol., 60: 23-26.
COURI, D. & MILKS, M. (1982) Toxicity and metabolism of the
neurotoxic hexacarbons n-hexane, 2-hexanone and 2,5-hexanedione.
Annu. Rev. Pharmacol. Toxicol., 22: 145-166.
COURI, D., ABDEL-RAHMAN, M.S., & HETLAND, L.B. (1978)
Biotransformation of n-hexane and methyl n-butyl ketone in guinea
pigs and mice. Am. Ind. Hyg. Assoc. J., 39: 295-300.
CREPET, M., GAFFURI, E., & PICOTTI, G. (1968) [Triarylphosphate
disease in the shoe industry.] Minerva med., 59(78): 4073-4075 (in
Italian).
DAHL, A.R., DAMON, E.G., MAUDERLY, J.L., ROTHENBERG, S.J., SEILER,
F.A., & MCCLELLAN, R.O. (1988) Uptake of 19 hydrocarbon vapors
inhaled by F344 rats. Fundam. appl. Toxicol., 10: 262-269.
DALE, G.H. & DREHMAN, L.E. (1985) Hexane. In: Kirk-Othmer
encyclopedia of chemical technology, 3rd ed., New York, Chichester,
Brisbane, Toronto, John Wiley and Sons, pp 926-930.
DECAPRIO, A.P. (1987) n-Hexane neurotoxicity: A mechanism involving
pyrrole adduct formation in axonal cytoskeletal protein.
Neurotoxicology, 8: 199-210.
DECAPRIO, A.P. & O'NEILL, E.A. (1985) Alterations in rat axonal
cytoskeletal proteins induced by in vitro and in vivo
2,5-hexanedione exposure. Toxicol. appl. Pharmacol., 78: 235-247.
DECAPRIO, A.P., OLAJOS, E.J., & WEBER, P. (1982) Covalent binding
of a neurotoxic n-hexane metabolite: conversion of primary amines
to substituted pyrrole adducts by 2,5-hexanedione. Toxicol. appl.
Pharmacol., 65: 440-450.
DECAPRIO, A.P., BRIGSS, R.G., JACKOWSKI, S.J., & KIM, J.C.S. (1988)
Comparative neurotoxicity and pyrrole-forming potential of
2,5-hexanedione and perdenterio-2,5-hexanedione in the rat.
Toxicol. appl. Pharmacol., 92: 75-85.
DE GROOT, D.M.G. & KEPNER, L.A. (1984) The effects of long term
discontinuous exposure to low levels of n-hexane on rats, Zeist,
The Netherlands, Medical Biological Laboratory, TNO (Conducted on
behalf of Shell Internationale Petroleum Maatschappij BV).
DEICHMANN, W.B. & LE BLANC, T.J. (1943) Determination of the
approximate lethal dose with about six animals. J. ind. Hyg.
Toxicol., 25: 415-417.
DELBRUCK, W.R. & KLUGE, A. (1982) [ The effects of n-hexane in man
and animals], Hamburg, German Society for Petroleum Sciences and
Coal Chemistry (Research Report No. 174-2) (in German).
DEMARTINO, C., MALORNI, W., AMANTINI, M.C., SCORZA BARCELLONA, P.,
& FRONTALI, N. (1987) Effects of respiratory treatment with
n-hexane on rat testis morphology. I. A light microscope study.
Exp. mol. Pathol., 46: 199-216.
DE ROSA, E., BARTOLUCCI, G.B., PERBELLINI, L., BRUGNONE, F., &
RAUSA, G. (1988) Environmental and biological monitoring of
exposure to toluene, styrene and n-hexane. Appl. ind. Hyg., 3:
332-337.
DFG (Deutsche Forschungsgemeinschaft) (1988) Maximum concentrations
at the workplace and biological tolerance values for working
materials: Report No. XXIV of the Commission for the Investigation
of Health Hazards of Chemical Compounds in the Work Area, Weinheim,
Verlag Chemie, p. 84.
DI-BOSCO, M. & FONZI, S. (1974) [So-called glue or shoe makers'
polyneuropathy.] Riv. Infort. Mal. prof., 61(Part 1): 165-192 (in
Italian).
DISCALZI, G.L., PERRELLI, G., & PAVAN, I. (1988) [Solvent exposure
risk among homeworkers.] Med. Lav., 79: 234-236 (in Italian).
DIVINCENZO, G.D., KAPLAN, C.J., & DEDINAS, J. (1976)
Characterization of the metabolites of methyl n-butyl ketone,
methyl isobutyl ketone and methyl ethyl ketone in guinea pig serum
and their clearance. Toxicol. appl. Pharmacol., 36: 511-522.
DIVINCENZO, G.D., HAMILTON, M.L., KAPLAN, C.J., & DEDINAS, J.
(1977) Metabolic fate and disposition of 14C-labelled methyl
n-butyl ketone in the rat. Toxicol. appl. Pharmacol., 41: 547-560.
DIVINCENZO, G.D., HAMILTON, M.L., KAPLAN, C.J., KRASAVAGE, W.J., &
O'DONOGHUE, J.L. (1978) Studies on the respiratory uptake and
excretion and the skin absorption of methyl n-butyl ketone in
humans and dogs. Toxicol. appl. Pharmacol., 44: 593-604.
DIVINCENZO, G.D., KRASAVAGE, W.J., & O'DONOGHUE, J.L. (1980) Role
of metabolism in hexacarbon neuropathy. In: Witschi, H., ed. The
scientific basis of toxicity assessment, Amsterdam, Oxford, New
York, Elsevier Science Publishers, pp. 183-200.
DOLARA, P., FRANCONI, F., & BASOSI, D. (1978) Urinary excretion of
some n-hexane metabolites. Pharmacol. Res. Commun., 10(6): 503-510.
DUNNICK, J.K. (1989) Toxicity studies of n-hexane in B6C3F, mice
(inhalation studies), Washington, DC, US Department of Health and
Human Services, National Toxicology Program (Report No. NTPTS 1).
DUNNICK, J.K., GRAHAM, D.G., YANG, R.S.H., HABER, S.B., & BROWN,
H.R. (1989) Thirteen-week toxicity study of n-hexane in B6C3F, mice
after inhalation exposure. Toxicology, 57: 163-172.
DURHAM, H.D., PENA, S.D.J., & CARPENTER, S. (1983) The neurotoxins
2,5-hexanedione and acrylamide promote aggregation in intermediate
filaments in cultured fibroblasts. Muscle Nerve, 6: 631-637.
DURHAM, H.D., PENA, S.D.J., & ECOBICHON, D.J. (1988)
Hexahydrocarbon effects on intermediate filament organization in
human fibroblasts. Muscle Nerve, 11: 160-165.
EBEN, A., FLUCKE, W., MINAIL, F., THYSSEN, J., & KIMMERLE, G.
(1979) Toxicological and metabolic studies of methyl n-butyl
ketone, 2,5-hexanedione and 2,5-hexanediol in male rats.
Ecotoxicol. environ. Saf., 3: 204-217.
EDELFORS, S. & RAVN-JONSEN, A. (1985) Calcium uptake in rat brain
synaptomsomes after short-term exposure to organic solvents: a
pilot study. Acta pharmacol. toxicol., 56: 431-434.
ELKINS, H.B. (1959) The chemistry of industrial toxicology, 2nd
ed., New York, Chichester, Brisbane, Toronto, John Wiley and Sons.
FAWCETT, P.R.W. & BARWICK, D.D. (1981) Studies in nerve conduction.
In: Walton, J.A., ed. Disorders of voluntary muscle, 4th ed., New
York, Churchill Livingstone, pp. 928-951.
FEDTKE, N. & BOLT, H.M. (1986a) Detection of 2,5-hexanedione in the
urine of persons not exposed to n-hexane. Int. Arch. occup.
environ. Health, 57: 143-148.
FEDTKE, N. & BOLT, H.M. (1986b) Methodological investigations on
the determination of n-hexane metabolites in urine. Int. Arch.
occup. environ. Health, 57: 149-158.
FEDTKE, N. & BOLT, H.M. (1987a) 4,5-dihydroxy-2-hexanone: a new
metabolite of n-hexane and of 2,5-hexanedione in rat urine.
Biochem. environ. mass Spectrom., 14: 563-572.
FEDTKE, N. & BOLT, H.M. (1987b) The relevance of 4,5-dihydroxy-2-
hexanone in the excretion kinetics of n-hexane metabolites in rat
and man. Arch. Toxicol., 61: 131-137
FICHERA, G., SPAGNA, C., & TIMPANARO, V. (1968) [Hydrocarbon
intoxication. Note II. Anatomo-pathological findings in
experimental hexane intoxication.] Folia med. (Napoli), 51: 633-640
(in Italian).
FILSER, J.G., PETER, H., BOLT, H.M., & FEDTKE, N. (1987)
Pharmacokinetics of the neurotoxin n-hexane in rat and man. Arch.
Toxicol., 60: 77-80.
FISEROVA-BERGEROVA, V. & PIERCE, J.T. (1989) Biological monitoring
V: Dermal absorption. Appl. ind. Hyg., 8(4): F14-F21.
FRANCHINI, I., CAVATORTA, A., FALZOI, M., LUCERTINI, S., & MUTTI,
A. (1983) Early indicators of renal damage in workers exposed to
organic solvents. Int. Arch. occup. environ. Health, 52: 1-9.
FRANKE, J.P., WIJSBEEK, J., DE ZEEUZ, R.A., MOLLER, M.R., &
NIERMEYER, H. (1988) Systematic analysis of solvents and other
volatile substances by gas chromatography. J. anal. Toxicol., 12:
20-24.
FROMMER, U., ULLRICH, V., & ORRENIUS, S. (1974) Influence of
inducers and inhibitors on the hydroxylation pattern of n-hexane in
rat liver microsomes. FEBS Lett., 41(1): 14-16.
FRONTALI, N., AMANTINI, M.C., SPAGNOLO, A., GUARCINI, A.M., &
SALTARI, M.C. (1981) Experimental neurotoxicity and urinary
metabolites of the C5-C7 alphatic hydrocarbons used as glue
solvents in shoe manufacture. Clin. Toxicol., 18(12): 1357-1367.
FUHNER, H. (1921) [The narcotic effects of gasoline and its
components (pentane, hexane, heptane, octane).] Biochem. Z., 115:
235-261 (in German).
GENTER, M.B., SZAKAL-QUIN, G., ANDERSON, C.W., ANTHONY, D.C., &
GRAHAM, D.G. (1987) Evidence that pyrrole formation is a
pathogenetic step in gamma-diketone neuropathy. Toxicol. appl.
Pharmacol., 87: 351-361.
GENTER ST. CLAIR, M.B., AMARNATH, V., MOODY, A., ANTHONY, D.C.,
ANDERSON, C.W., & GRAHAM, D.G. (1988) Pyrrole oxidation and protein
cross-linking as necessary steps in the development of gamma-
diketone neuropathy. Chem. Res. Toxicol., 1: 179-185.
GENTRY, S.J. & WALSH, P.T. (1987) Eight-hour TWA personal
monitoring using a diffusive sampler and short-term stain tube. Am.
Ind. Hyg. Assoc. J., 48: 287-292.
GERARDE, H.W. & LINDEN, N.J. (1963) Toxicological studies on
hydrocarbons. IX. The aspiration hazard and toxicity of
hydrocarbons and hydrocarbon mixtures. Arch. environ. Health, 6:
329-341.
GHITTORI, S., IMBRIANI, M., PEZZAGNO, G., & CAPODAGLIO, E. (1987)
The urinary concentration of solvents as a biological indicator of
exposure: proposal for the biological equivalent exposure limit for
nine solvents. Am. Ind. Hyg. Assoc. J., 48: 786-790.
GILLIES, P.J., NORTON, R.M., WHITE, E.L., & BUS, J.S. (1980)
Inhibition of sciatic nerve sterologenesis in hexacarbon-induced
distal axonopathy in the rat. Toxicol. appl. Pharmacol., 54:
217-222.
GIULIANO, G., IANNACCONE, A., & ZAPPOLI, R. (1974)
[Electroencephalographic research in shoe industry workers exposed
to the risk of poisoning from adhesive solvents.] Lav. Um., 26(2):
33-42 (in Italian).
GOEL, S.K., RAO, G.S., & PANDYA, K.P. (1982) Toxicity of n-hexane
and n-heptane: some biochemical changes in liver and serum.
Toxicol. Lett., 14: 169-174.
GOEL, S.K., RAO, G.S., & KRISHNAMURTI, C.R. (1987) Toxicity of
n-hexane and 2,5-hexanediol to the haemopoietic system: a
preliminary report. Xenobiotica, 17: 217-222.
GOEL, S.K., RAO, G.S., & PANDYA, K.P. (1988) Hepatotoxic effects
elicited by n-hexane or n-heptane. J. appl. Toxicol., 8: 81-84.
GOMEZ-ARROYO, S., CASTILLO-RUIZ, P., & VILLALOBOS-PIETRINI, R.
(1986) Chromosomal alterations induced in Vicia faba by different
industrial solvents: thinner, toluene, benyene, n-hexane, n-heptane
and ethyl acetate. Cytologia, 51: 133-142.
GONZALEZ, E.G. & DOWNEY, J.A. (1972) Polyneuropathy in a glue
sniffer. Arch. phys. Med. Rehabil., 53: 333-337.
GOTO, I., MATSUMURA, M., INOUE, N., MURAI, Y., SHIDA, K., SANTA,
T., & KUROIWA, Y. (1974) Toxic polyneuropathy due to glue sniffing.
J. Neurol. Neurosurg. Psychiatry, 37: 848-853.
GRAHAM, D.G. & GOTTFREID, M.R. (1984) Cross species extrapolation
in hydrocarbon neuropathy. Neurobehav. Toxicol. Teratol., 6:
433-435.
GRAHAM, D.G., ANTHONY, D.C., & BOEKELHEIDE, K. (1982a) In vitro and
in vivo studies of the molecular pathogenesis of n-hexane
neuropathy. Neurobehav. Toxicol. Teratol., 4: 629-634.
GRAHAM, D.G., ANTHONY, D.C., BOEKELHEIDE, K., MASCHMANN, N.A.,
RICHARDS, R.G., WOLFRAM, J.W., & SHAW, B.R. (1982b) Studies of the
molecular pathogenesis of hexane neuropathy. II. Evidence that
pyrrole derivatization of lysyl residues leads to protein
crosslinking. Toxicol. appl. Pharmacol., 64: 415-422.
GRAHAM, D.G., ANTHONY, D.C., SZAKAL-QUIN, G., GOTTFREID, M.R., &
BOEKELHEIDE, K. (1985) Covalent crosslinking of neurofilaments in
the pathogenesis of n-hexane neuropathy. Neurotoxicology, 6: 55-64.
GRIFFITHS, I.R., KELLY, P.A.T., CARMICHAEL, S., MCCULLOCH, M., &
WATERSON, M. (1981) The relationship of glucose utilization and
morphological change in the visual system in hexacarbon neuropathy.
Brain Res., 222: 447-451.
GRIFFITH, J.F., NIXON, G.A., BRUCE, R.D., REER, P.J., & BANNAN,
E.A. (1980) Dose-response studies with chemical irritants in the
albino rabbit eye as a basis for selecting optimum testing
conditions for predicting hazard to the human eye. Toxicol. appl.
Pharmacol., 55: 501-513.
GOVERNA, M., CALISTI, R., COPPA, G., TAGLIAVENTO, G., COLOMBI, A.,
& TRONI, W. (1987) Urinary excretion of 2,5-hexanedione and
peripheral polyneuropathies in workers exposed to hexane. J.
Toxicol. environ. Health, 20: 219-228.
GROVES, F.R. (1988) Effect of cosolvents on the solubility of
hydrocarbons in water. Environ. Sci. Technol., 22: 282-286.
GUTHRIE, V.B. (1960) Petroleum products handbook, New York, McGraw-
Hill.
HADJIIVANOVA, N.B., SALOVSKI, P.Z., GROSEVA, M.M.,
CHARAKCHIEVA, S.B., & NECHEV, Ch.K. (1987) Early effects of
n-hexane and irradiation on the lung surfactant system. Acta
physiol. pharmacol. Bulg., 13: 25-29.
HAZELTON LABORATORIES (1981) In vivo and in vitro mutagenicity
studies n-hexane (hexane UV), Vienna, Virginia, Hazelton
Laboratories (Project No. 596-114 and 596-115) (Prepared for the
American Petroleum Institute).
HENDERSON, V.E. & SMITH, A.H.R. (1936) Propylene impurities.
Hexenes and hexanes. J. Pharmacol. exp. Ther., 58: 319-327.
HERSKOWITZ, A., ISHII, N., & SCHAUMBERG, H. (1971) n-Hexane
neuropathy. A syndrome occurring as a result of industrial
exposure. New Engl. J. Med., 285(2): 82-85.
HEWITT, W.R., MIYAJIMA, H., COTE, M., & PLAA, G.L. (1980) Acute
alteration of chloroform induced hepato- and nephrotoxicity by
n-hexane, methyl n-butyl ketone and 2,5-hexanedione. Toxicol. appl.
Pharmacol., 53: 230-248.
HINE, C.H. & ZUIDEMA, H.H. (1970) The toxicological properties of
hydrocarbon solvents. Ind. Med., 39(5): 39-44.
HODGSON, A.T., GIRMAN, J.R., & BINENBOYM, J. (1986) A multisorbent
sampler for volatile organic compounds in indoor air. In:
Proceedings of the 79th Annual Meeting, Minneapolis, 22-27 June,
1986, Minneapolis, Minnesota, Air Pollution Control Association,
pp. 1-15 (Report No. 86-37-1).
HOLMBERG, B. & LUNDBERG, P. (1989) Assessment and management of
occupational risks in the Nordic (Scandinavian) countries. Am. J.
ind. Med., 15: 615-626.
HONMA, T. (1983) Changes in acetylcholine metabolism in rat brain
after a short-term exposure to toluene and n-hexane. Toxicol.
Lett., 16: 17-22.
HONMA, T., MIYAGAWA, M., SATO, M., & HASEGAWA, H. (1982) Increase
in glutamine content of rat midbrain induced by short-term exposure
to toluene and hexane. Ind. Health, 20: 109-115.
HOUK, V.S., SCHALKOWSKY, S., & CLAXTON, L.D. (1989) Development and
validation of the spiral Salmonella assay: an automated approach to
bacterial mutagenicity testing. Mutat. Res., 223: 49-64.
HOWD, R.A., BINGHAM, L.R., STEEGER, T.M., REBERT, C.S., & PRYOR,
G.T. (1982) Relation between schedules of exposure to hexane and
plasma levels of 2,5-hexanedione. Neurobehav. Toxicol. Teratol., 4:
87-91.
HOWD, R.A., REBERT, C.S., DICKINSON, J., & PRYOR, G. (1983) A
comparison of the rates of development of functional hexane
neuropathy in weanling and young adult rats. Neurobehav. Toxicol.
Teratol., 5: 63-68.
HOWELL, W.E. (1979) A neurobehavioural evaluation of the pre-natal
toxicity of n-hexane in rats, Cincinnati, Ohio, University of Ohio,
78 pp (Ph.D Thesis) (Dissertation Abstracts Int B. No. 1610B).
HOWELL, W.E. & COOPER, G.P. (1981) Neurophysiological evaluation of
prenatal n-hexane toxicity. Toxicologist, 1: 152 (Abstact 552).
HUANG, C.-C. & CHU, N.-S. (1989) Evoked potentials in chronic
n-hexane intoxication. Clin. Electroencephalogr., 20: 162-168.
HUSMAN, K. & KARLI, P. (1980) Clinical neurological findings among
car painters exposed to a mixture of organic solvents. Scand. J.
Work Environ. Health, 6: 33-39.
ICHIHARA, K., KUSONOSE, E., & KUSUNOSE, M. (1969) Microsomal
hydroxylation of decane. Biochem. Biophys. Acta, 176: 713-719.
IIDA, M. (1982) Neurophysiological studies of n-hexane
polyneuropathy in the sandal factory. Electroencephalogr. clin.
Neurophysiol., 36(Suppl.): 671-681.
IIDA, M. & YAMAMOTO, H. (1973) [Prognosis of n-hexane
polyneuropathy. Follow-up studies of mass outbreak in District F in
Mie Prefecture.] Igaku Ayumi, 84(4): 199-201 (in Japanese).
IIDA, M., YAMAMURA, Y., & SOBUE, I. (1969) Electromyographical
findings and conduction velocity in n-hexane polyneuropathy.
Electromyography, 9: 247-261.
IKEDA, M. & KASAHARA, M. (1986) n-hexane and benzene contents in
gasoline for industrial purpose. Ind. Health, 24: 63-66.
IKEDA, M., KOIZUMI, A., KASAHARA, M., & FUJITA, H. (1986) Combined
effects of n-hexane and toluene on norepinephrine and dopamine
levels in rat brain tissues after long-term exposures. Bull.
environ. Contam. Toxicol., 36: 510-517.
IMBRIANI, M., GHITTORI, S., BORLINI, F., PEZZAGNO, G., &
CAPODAGLIO, E. (1984a) [ n-hexane and toluene in the urine of
occupationally exposed subjects.] Boll. Soc. Ital. Biol. Sper., 60:
1919-1925 (in Italian).
IMBRIANI, M., GHITTORI, S., PEZZAGNO, G., & CAPODAGLIO, E. (1984b)
n-hexane urine elimination and weighted exposure concentration.
Int. Arch. occup. environ. Health, 55: 33-41.
INOUE, T., TAKEUCHI, Y., TAKEUCHI, S., YAMADA, S., SUZUKI, H.,
MATSUSHITA, T., MIYAGAKI, M., MAEDA, K., & MATSUMOTO, T. (1970)
[Industrial health survey of high incidence of n-hexane
intoxication among vinyl sandal manufacturers.] Ind. Health, 12(3):
73-84 (in Japanese).
INOUE, T., YAMADA, S., MIYAGAKI, H., & TAKEUCHI, Y. (1971)
Polyneuritis caused by n-hexane and the maximum allowable
concentration. In: Proceedings of the XVI International Congress on
Occupational Health, 22-27 September 1969, Tokyo, Industrial Safety
Association, pp. 522-524.
IRPTC (1990) IRPTC Legal file, Geneva, International Register of
Potentially Toxic Chemicals, United Nations Environment Programme.
ISHIDATE, M. & SOFUNI, T. (1984) Primary mutagenicity screening of
food additives currently used in Japan. Food chem. Toxicol., 22(8):
623-636.
IWASAKI, K. & TSURUTA, H. (1984) Molecular mechanisms of hexane
neuropathy. Significant differences in pharmacokinetics between
2,3- 2,4-, and 2,5-hexanedione. Ind. Health, 22: 177-187.
IWATA, M., TAKEUCHI, Y., HISANAGA, N., & ONO, Y. (1983a) Changes of
n-hexane metabolites in urine of rats exposed to various
concentrations of n-hexane and to its mixture with toluene or MEK.
Int. Arch. occup. environ. Health, 53: 1-8.
IWATA, M., TAKEUCHI, Y., HISANAGA, N., & ONO, Y. (1983b) A study on
biological monitoring of n-hexane exposure. Int. Arch. occup.
environ. Health, 51: 253-260.
IWATA, M., TAKEUCHI, Y., HISANAGA, N., & ONO, Y. (1984) Changes of
n-hexane neurotoxicity and its urinary metabolites by long-term co-
exposure with MEK or toluene. Int. Arch. occup. environ. Health, 54:
273-281.
JAKOBSON, I., WAHLBERG, J.E., HOLMBERG, B., & JOHANSSON, G. (1982)
Uptake via the blood and elimination of 10 organic solvents
following epicutaneous exposure of anaesthetized guinea pigs.
Toxicol. appl. Pharmacol., 63: 181-187.
JEPPSSON, R. (1975) Parabolic relationship between lipophilicity
and biological activity of aliphatic hydrocarbons, ethers and
ketones after intravenous injection of emulsion formulations into
mice. Acta pharmacol. toxicol., 37: 56-64.
JONES, H.G. & CAVANAGH, J.B. (1982) Recovery from 2,5-hexanediol
intoxication of the retinotectal tract of the rat. An
ultrastructural study. Acta neuropathol. (Berlin), 58: 286-290.
JUHNKE, I. & LUDEMANN, D. (1978) [Results of trials of 200 chemical
compounds for acute toxicity for fish with the golden orfe test.]
Z. Wasser Abwasser Forsch., 11: 161-164 (in German).
KANNAN, K., SINGH, K.P., GOEL, S., & SHANKER, R. (1985) Effects of
2,5-hexanedione on immunocompetence of mice. Environ. Res., 36: 14-25.
KASAHARA, M. & IKEDA, M. (1987) Spontaneous desorption of organic
solvents from carbon cloth. Ind. Health, 25: 73-81
KASAHARA, M., SUZUKI, H., TAKEUCHI, Y., HARA, I., & IKEDA, M.
(1987) n-Hexane, benzene and other aromatic components in petroleum
distillate solvents in Japan. Ind. Health, 25: 205-214.
KATAGIRI, Y. & OHASHI, T. (1975) Photochemical reactions of
hydrocarbons in smog chamber. Annu. Rep. Kanagawa Prefect. Environ.
Cent., 6: 47.
KATZ, G.V., O'DONOGHUE, J.L., DIVINCENZO, G.D., & TERHAAR, C.J.
(1980) Comparative neurotoxicity and metabolism of ethyl n-butyl
ketone and methyl n-butyl ketone in rats. Toxicol. appl.
Pharmacol., 52: 153-158.
KAWACHI, T., YAHAGI, T., KADA, T., TAZIMA, Y., ISHIDATE, M.,
SASAKI, M., & SUGIYAMA, T. (1982) Co-operative programme on short-
term assays for carcinogenicity in Japan. In: Molecular and
cellular aspects of carcinogen screening tests, Lyon, International
Agency for Research on Cancer, pp. 323-330 (IARC Scientific
Publications No. 27).
KEPLINGER, M.L., LANIER, G.E., & DEICHMANN, W.B. (1959) Effects of
environmental temperature on the acute toxicity of a number of
compounds in rats. Toxicol. appl. Pharmacol., 1: 156-161.
KIMURA, E.T., EBERT, D.M., & DODGE, P.W. (1971) Acute toxicity and
limits of solvent residue for sixteen organic solvents. Toxicol.
appl. Pharmacol., 19: 699-704.
KLIGMAN, A.M. (1966) The identification of contact allergens by
human assay. III. The maximization test: a procedure for screening
and rating contact sensitisers. J. invest. Dermatol., 47: 393-409.
KLIMES, I., VISKUPIC, E., ZORAD, S., SVABOVA, E., KISS, A., &
MACHO, L. (1987) Effect of prolonged n-hexane administration on in
vitro insulin degradation in rat liver and blood. Med. Sci. Res.,
15: 1427-1429.
KLOPFFER, W., HAAG, F., KOHL, E.-G., & FRANK, R. (1988) Testing of
the abiotic degradation of chemicals in the atmosphere: The smog
chamber approach. Ecotoxicol. environ. Saf., 15: 298-319.
KOROBKIN, R., ASBURY, A.K., SUMNER, A.J., & NIELSEN, S.L., (1975)
Glue sniffing neuropathy. Arch. Neurol., 32: 158-162.
KRAMER, A., STAUDINGER, H., & ULLRICH, V. (1974) Effect of n-hexane
inhalation on the monooxygenase system in mice liver microsomes.
Chem.-biol. Interact., 8: 11-18.
KRASAVAGE, W.J., O'DONOGHUE, J.L., & TERHAAR, C.J. (1979) Oral
chronic toxicity of methyl n-propyl ketone, methyl n-butyl ketone
and hexane in rats. Toxicol. appl. Pharmacol., 48(Part 2): A205.
KRASAVAGE, W.J., O'DONOGHUE, J.L., DIVINCENZO, G.D., & TERHAAR,
C.J. (1980) The relative neurotoxicity of methyl n-butyl ketone,
n-hexane and their metabolites. Toxicol. appl. Pharmacol., 52: 433-441.
KRONEVI, T., WAHLBERG, J., & HOLMBERG., B. (1979) Histopathology of
skin, liver and kidney after epicutaneous administration of five
industrial solvents to guinea-pigs. Environ. Res., 19: 56-69.
KULIG, B.M. (1983) Chronic behavioural toxicology of hexane and
CS2: Progress report, Zeist, The Netherlands, Medical Biological
Laboratory, TNO (Conducted on behalf of Shell Internationale
Petroleum Maatschappij BV).
KULIG, B.M. & WOLTHIUS, O.L. (1984) The development of neurological
and behavioural impairments in rats exposed subchronically to
n-hexane. In: Abstracts of the International Conference on Organic
Solvent Toxicity, Stockholm, October 1984.
KULIG, B.M., DEMIK, G., & WOLTHIUS, O.L. (1984) 2,5-Hexanedione
concentrations and hexane-induced neuropathy in Wistar rats.
Abstracts of the Second International Conference on Neurotoxicology
of Selected Chemicals, Chicago, 1983. Neurotoxicology, 5(2): 307.
KULIG, B.M., VANWERSCH, R.A.P., & WOLTHUIS, O.L. (1985) The
automated analysis of coordinated hindlimb movement in rats during
acute and prolonged exposure to toxic agents. Toxicol. appl.
Pharmacol., 80: 1-10.
KURITA, H. (1974) [Experimental studies on the effects of n-hexane
on albino rats.] Ind. Health, 9(8): 24-29 (in Japanese).
LADEFOGED, O. & PERBELLINI, L. (1986) Acetone-induced changes in
the toxicokinetics of 2,5-hexanedione in rabbits. Scand. J. Work.
Environ. Health, 12: 627-629.
LADEFOGED, O., HASS, U., & SIMONSEN, L. (1989) Neurophysiological
and bahavioural effects of combined exposure to 2,5-hexanedione and
acetone or ethanol in rats. Pharmacol. Toxicol., 65: 372-375.
LAFFORT, P. & DRAVNIEKS, A. (1973) An approach to a physico-
chemical model of olfactory stimulation in vertebrates by single
compounds. J. theor. Biol., 38: 335-345.
LAGUENY, A., JULIEN, J., VALLAT, J.M., FERRER, X., DELIAC, P., &
DELIAC, M. (1982) Etude électrophysiologique de deux cas de
polynévrite provoquée par une association n-hexane-méthyléthyl-
cétone. Rev. EEG Neurophysiol., 12: 158-166.
LANKAS, G.R., BAXTER, C.S., & CHRISTIAN, R.T. (1978) Effect of
alkane tumour-promoting agents on chemically induced mutagenesis in
cultured V79 Chinese hamster cells. J. Toxicol. environ. Health, 4:
37-41.
LAPADULA, D.M., IRWIN, R., SUWITA, E., & ABOU-DONIA, M.B. (1986)
Cross-linking of neurofilament proteins of rat spinal cord in vivo
after administration of 2,5-hexanedione. J. Neurochem., 46: 1843-1850.
LAZAREV, N.W. (1929) [The toxicity of various hydrocarbon vapours.]
Arch. exp. Pathol. Pharmakol., 143: 223-233 (in German).
LIJINSKY, W. & RAHA, C.R. (1961) Polycyclic aromatic hydrocarbons
in commercial solvents. Toxicol. appl. Pharmacol., 3: 469-473.
LITTON BIONETICS (1979) Teratology study in rats: n-hexane, Litton
Bionetics (LBI Project No. 20698-9) (Final Report submitted to the
American Petroleum Institute).
LITTON BIONETICS (1980) Mutagenicity evaluation of n-hexane in the
mouse dominant lethal assay, Litton Bionetics (LBI Project No.
21141-01) (Final Report submitted to the American Petroleum
Institute).
LODEN, M. (1986) The in vitro permeability of human skin to
benzene, ethylene glycol, formaldehyde and n-hexane. Acta
pharmacol. toxicol., 58: 382-389.
LONNEMAN, W.A., KOPCZYNSKI, S.L., DARLEY, P.E., & SUTTERFIELD, F.D.
(1974) Hydrocarbon composition of urban air pollution. Environ.
Sci. Technol., 8: 229-236.
LUNGARELLA, G., FONZI, L., & CENTINI, F. (1980) Respiratory tract
lesions induced in rabbits by short-term exposure to n-hexane. Res.
Commun. chem. Pathol. Pharmacol., 29(1): 129-139.
LUNGARELLA, G., BARNI-COMPARINI, I., & FONZI, L. (1984) Pulmonary
changes induced in rabbits by long-term exposure to n-hexane. Arch.
Toxicol., 55: 224-228.
MCAULIFFE, C. (1963) Solubility in water of C1-C9 hydrocarbons.
Nature (Lond.), 200: 1092-1093.
MCCARROLL, N.E., PIPER, C.E., & KEECH, B.H., (1980) Bacterial
microsuspension assays with benzene and other organic solvents.
Environ. Mutagen., 2(2): 281-281.
MCCARROLL, N.E., PIPER, C.E., & KEECH, B.H. (1981a) An E. coli
microsuspension assay for the detection of DNA damage induced by
direct-acting agents and promutagens. Environ. Mutagen., 3: 429-444.
MCCARROLL, N.E., KEECH, B.H., & PIPER, C.E. (1981b) A
microsuspension adaptation of the Bacillus subtilis "rec" assay.
Environ. Mutagen., 3: 607-616.
MCNAMARA, B.C. & JEFCOATE, C.R. (1988) Synergistic stimulation of
pregnenolone synthesis in rat adrenal mitochondria by n-hexane and
cardiolipin. Arch. Biochem. Biophys., 260: 780-788.
MAHMOUD, G. & LACHAPELLE, J.M. (1984) Etude de l'efficacité d'un
gel de protection anti-solvants, vis-à-vis de l'irritation cutanée
provoquée par le n-hexane. Arch. Mal. prof. Méd. Trav. Séc. soc.,
45(4): 223-237.
MAHMOUD, G. & LACHAPELLE, J.M. (1985) Evaluation expérimentale de
l'efficacité de crèmes-barrière et de gels antisolvants dans la
prévention de l'irritation cutanée provoquée par des solvants
organiques. Cah. Méd. Trav., 22(2/3): 163-168.
MAIZLISH, N.A., LANGOLF, G.D., WHITEHEAD, L.W., FINE, L.J., ALBERS,
J.W., GOLDBERG, J., & SMITH, P. (1985) Behavioural evaluation of
workers exposed to mixtures of organic solvents. Br. J. ind. Med.,
42: 579-590.
MAIZLISH, N.A., FINE, L.J., ALBERS, J.W., WHITEHEAD, L., & LANGOLF,
G.D. (1987) A neurological evaluation of workers exposed to
mixtures of organic solvents. Br. J. ind. Med., 44: 14-25.
MALORNI, W., DEMARTINO, C., AMANTINI, M., & FRONTALI, N. (1984)
Effects of subchronic treatment with n-hexane on rat testis. A
morphological study. Ultramicroscopy, 12: 148.
MALORNI, W., FORMISANO, G., & DONELLI, G. (1989) Morphologic
changes induced in vitro by 2,5-hexanedione. In vitro cell. dev.
Biol., 25: 82-90.
MARCHISEPPE, I., VALENTINO, M., GOVERNA, M., & STOCCHI, V. (1989)
Determination of total 2,5-hexanedione by reversed-phase high-
performance liquid chromatography. J. Chromatology, 495: 288-294.
MARKS, T.A., FISHER, P.W., & STAPLES, R.E. (1980) Influence of
n-hexane on embryo and fetal development in mice. Drug chem.
Toxicol., 3(4): 393-406.
MELLAN, I., ed. (1977) Industrial solvents handbook, 2nd ed., Park
Ridge, New Jersey, Noyes Data Corporation, p. 14.
MEDINILLA, J., DE & ESPIGARES, M. (1989) [Hexane levels in olive
dreg extraction plants in Granada, Spain.] Ig. mod., 91: 353-361 (in
Spanish).
MISUMI, J. & NAGANO, M. (1984) Neurophysiological studies on the
relation between the structural properties and neurotoxicity of
aliphatic hydrocarbon compounds in rats. Br. J. ind. Med., 41:
526-532.
MISUMI, J. & NAGANO, M. (1985) Experimental study on the
enhancement of the neurotoxicity of methyl butyl ketone by non-
neurotoxic aliphatic monoketones. Br. J. ind. Med., 42: 155-161.
MIYAGAKI, H. (1967) [Electrophysiological studies of the peripheral
neurotoxicity of n-hexane.] Ind. Health, 9: 660-671 (in Japanese).
MORTELMANS, K., HAWORTH, S., LAWLOR, T., SPECK, W., TAINER, B., &
ZEIGER, E. (1986) Salmonella mutagenicity tests. II. Results from
the testing of 270 chemicals. Environ. Mutagen., 8(Suppl. 7): 1-119.
MUTTI, A., LUCERTINI, S., VENTURI, G., CAVATORTA, A., & FRANCHINI,
I. (1980) Organic solvents and glomerulonephritis. Negative
epidemiological findings for hydrocarbons. Abstracts of the 2nd
International Congress on Toxicology, Brussels. Toxicol. Lett.,
Spec. Issue 1: 220.
MUTTI, A., LUCERTINI, S., FALZOI, M., CAVATORTA, A., & FRANCHINI,
I. (1981) Organic solvents and chronic glomerulonephritis. A cross-
sectional study with negative findings for aliphatic and alicyclic
C5-C7 hydrocarbons. J. appl. Toxicol., 1(4): 224-226.
MUTTI, A., CAVATORTA, A., LUCERTINI, S., ARFINI, G., FALZOI, M., &
FRANCHINI, I. (1982a) Neurophysiological changes in workers exposed
to organic solvents in a shoe factory. Scand. J. Work Environ.
Health, 8(Suppl. 1): 136-141.
MUTTI, A., CAVATORTA, A., LOMMI, G., LOTTA, S., & FRANCHINI, I.
(1982b) Neurophysiological effect of long term exposure to
hydrocarbon mixtures. Arch. Toxicol., Suppl. 5: 120-124.
MUTTI, A., FERRI, F., LOMMI, G., LOTTA, S., LUCERTINI, S., &
FRANCHINI, I. (1982c) n-Hexane induced changes in nerve conduction
velocities and somatosensory evoked potentials. Int. Arch. occup.
environ. Health, 51: 45-54.
MUTTI, A., FALZOI, M., LUCERTINI, S., ARFINI, G., ZIGNANI, M.,
LOMBARDI, S., & FRANCHINI, I. (1984) n-Hexane metabolism in
occupationally exposed workers. Br. J. ind. Med., 41: 533-538.
NAKAJIMA, T. & MURAYAMA, N. (1985) Polyneuropathy caused by
n-hexane used under the commercial name of "benzine". Ind.
Health, 27: 340-341.
NASLUND, B.M.A. & HALPERT, J.S. (1984) Selective inhibition by
chloramphenicol of cytochrome P-450 isozymes in rat lung and liver
involved in the hydroxylation of n-hexane. J. Pharmacol. exp.
Ther., 231(1): 16-22.
NELLIGAN, R.E. (1962) Hydrocarbons in the Los Angeles atmosphere.
Arch. environ. Health, 5: 581-591.
NELSON, K.W., EGE, J.F., ROSS, M., WOODMAN, L.E., & SILVERMAN, L.
(1943) Sensory response to certain industrial solvent vapours. J.
ind. Hyg. Toxicol., 25: 282-285.
NICKELSEN, R.L., RODER, M.M., & BERARDINELLI (1986) Permeation of
chemical protective clothing by three binary solvent mixtures. Am.
Ind. Hyg. Assoc. J., 47: 236-240.
NILSEN, O.G., TOFTGARD, R., ENG, L., & GUSTAFSSON, J. (1981) Regio-
specificity of purified forms of rabbit liver microsomal cytochrome
P-450 in the metabolism of benzo( a)pyrene, n-hexane and
7-ethoxyresorufin. Acta pharmacol. toxicol., 48: 369-376.
NIOSH (1977a) Criteria for a recommended standard: Occupational
exposure to alkanes (C5-C8), Cincinnati, Ohio, National Institute
for Occupational Safety and Health (DHEW (NIOSH) Publication No.
77-151).
NIOSH (1977b) NIOSH manual on analytical methods, 2nd ed.,
Cincinnati, Ohio, National Institute for Occupational Safety and
Health, Vol. 1 (DHEW Publication No. (NIOSH) 77-157-A).
NIOSH (1981a) Health hazard evaluation report, Cincinnati, Ohio,
National Institute for Occupational Safety and Health (HHE-80-040-
860).
NIOSH (1981b) Health hazard evaluation report, Cincinnati, Ohio,
National Institute for Occupational Safety and Health (HHE-81-240-
1031).
NIOSH (1983) Health hazard evaluation report, Cincinnati, Ohio,
National Institute for Occupational Safety and Health (HETA-80-256-
1386).
NIOSH (1984) Manual of analytical methods: Hydrocarbons method
1500, Cincinnati, Ohio, National Institute for Occupational Safety
and Health, pp. 1500/1-1500/7.
NIOSH (1988) Testimony to DOL on the Occupational Safety and Health
Administration's proposed rule on air contaminants, Cincinnati,
Ohio, National Institute for Occupational Safety and Health (Pocket
No. H-020).
NOLTING, F., BEHNKE, W., & ZETZSCH, C. (1988) A smog chamber for
studies of the reactions of terpenes and alkanes with ozone and OH.
J. atmos. Chem., 6: 47-59.
NOMEIR, A.A. & ABOU-DONIA, M.B. (1985) Analysis of n-hexane,
2-hexanone, 2,5-hexanedione, and related chemicals by capillary gas
chromatography and high-performance liquid chromatography. Anal.
Biochem., 151: 381-388.
NOMIYAMA, K. & NOMIYAMA, H. (1974a) Respiratory retention, uptake
and excretion of organic solvents in man. Benzene, toluene,
n-hexane, trichloroethylene, acetone, ethylacetate and ethyl
alcohol. Int. Arch. Arbeitsmed., 32: 75-83.
NOMIYAMA, K. & NOMIYAMA, H. (1974b) Respiratory elimination of
organic solvents in man. Benzene, toluene, n-hexane,
trichloroethylene, acetone, ethylacetate and ethyl alcohol. Int.
Arch. Arbeitsmed., 32: 85-91.
NOMIYAMA, K. & NOMIYAMA, H. (1975) [Concerning the cutaneous
absorption of n-hexane in humans.] Jpn. J. Hyg., 30: 140 (in
Japanese).
NOMIYAMA, K., YOSHIDA, T., & YANAGISAWA, H. (1973) [Percutaneous
absorption of n-hexane caused severe polyneuropathy.] In:
[ Proceedings of the 46th Annual Meeting of the Japanese Association
of Industrial Health], Tokyo, Japanese Association of Industrial
Health, pp. 560-561 (in Japanese).
NOTTEN, W.R.F. & HENDERSON, P.T. (1975a) The influence of n-hexane
treatment on the glucuronic acid pathway and activity of some drug
metabolizing enzymes in guinea pig. Biochem. Pharmacol., 24: 127-131.
NOTTEN, W.R.F. & HENDERSON, P.T. (1975b) Action of n-alkanes on
drug-metabolizing enzymes from guinea-pig liver. Biochem.
Pharmacol., 24: 1093-1097.
NYLEN, P., EBENDAL, T., ERIKSDOTTER-NILSSON, M., HANSSON, T.,
HENSCHEN, A., JOHNSON, A.C., KRONEVI, T., KVIST, U., SJOSTRAND,
N.O., HOGLUND, G., & OLSUN, L. (1989) Testicular atrophy and loss
of nerve growth factor - immunoreactive germ cell line in rats
exposed to n-hexane and a protective effect of simultaneous
exposure to toluene or xylene. Arch. Toxicol., 63: 296-307.
O'DONOGHUE, J.L. (1985) Alkanes, alcohols, ketones and ethylene
oxide. In: O'Donoghue, J.L., ed. Neurotoxicology of industrial and
commercial chemicals, Boca Raton, Florida, CRC Press, Vol. 2, pp.
61-97.
O'DONOGHUE, J.L., KRASAVAGE, W.J., & TERHAAR, C.J. (1978a) A
comparative chronic toxicity study of methyl n-propyl ketone,
methyl n-butyl ketone, and hexane by ingestion, Rochester, New
York, Eastman Kodak Company (Unpublished Report No. 104657Y).
O'DONOGHUE, J.L., KRASAVAGE, W.J., & TERHAAR, C.J. (1978b) Toxic
effects of 2,5-hexanedione. Toxicol. appl. Pharmacol., 45: 269
(abstract 115).
OETTEL, H. (1936) [The influence of organic liquids on skin.] Arch.
exp. Pathol. Pharmacol., 38: 641-696 (in German).
OISHI, H. & MINENO, K. (1964) [Polyneuropathy caused by an organic
solvent ( n-hexane).] Saigai-Igaku, 7(4): 218-222 (in Japanese).
OKAMOTO, S., SHIRBAE, T., KURIMARA, T., & ARAKI, S. (1977) [A case
of n-hexane polyneuropathy occurring in an interior decorator and
experimental n-hexane neuropathy in rats.] Clin. Neurol. (Tokyo),
17: 237-242 (in Japanese).
ONO, Y. & TAKEUCHI, Y. (1980) [Polyneuropathy thought to be
petroleum benzine poisoning occurring in a worker engaged in
cleaning gas meters.] Sangyo Igaku, 22: 256-262 (in Japanese).
ONO, Y., TAKEUCHI, Y., & HISANAGA, N. (1979) [Studies on the method
of measuring nerve conduction velocity in rat's tail and on the
comparative toxicity of n-hexane, methyl n-butyl ketone and
2,5-hexanedione.] Jpn. J. ind. Health, 21: 528-538 (in Japanese
with English summary).
ONO, Y., TAKEUCHI, Y., & HISANAGA, N. (1981) A comparative study on
the toxicity of n-hexane and its isomers on the peripheral nerve.
Int. Arch. occup. environ. Health, 48: 289-294.
ONO, Y., TAKEUCHI, Y., HISANAGA, N., IWATA, M., KITOH, J., &
SUGIURA, Y. (1982) Neurotoxicity of petroleum benzine compared with
n-hexane. Int. Arch. occup. environ. Health, 50: 219-229.
ONOSAKA, S., OCHI, Y., MIN, K.-S., FUJITA, Y., & TANAKA, K. (1988)
[Influences of compounds on metallothionein concentration in mouse
tissues: II. Decrease of pancreatic metallothionein concentration
by n-hexane.] Eisei Kagaku, 34: 446-450 (in Japanese).
OSHA (Occupational Safety and Health Administration) (1989) Air
contaminants: Final rule. Fed. Reg., 54(12): 2921, 2939 (29 CFR Part
1910).
PALAO, A. & LAJO, J.L. (1981) [Toxic polyneuropathy as a
consequence of the use of glue and chemical solvents in the shoe
industry.] Arch. Neurobiol., 44(1): 35-48 (in Spanish).
PARLATO, G., INSERRA, A., & TIMPANARO, V. (1968)
Electrocardiographic changes in experimental poisoning with liquid
hydrocarbons (petrol ether, hexane and heptane). Folia med., 51:
799-805.
PASTERNAK, T., FLOOD, D.G., ESKIN, T.A., & MERIGAN, W.H. (1985).
Selective damage to large cells in the cat retinogeniculate pathway
by 2,5-hexanedione. J. Neurosci., 5: 1641-1652.
PATTY, F.A. & YANT, W.P. (1929) Odour intensity and symptoms
produced by commercial propane, butane, pentane, hexane and heptane
vapour, Washington, DC, US Bureau of Mines, 10 pp (Report No. RI
2979).
PAULSON, G.W. & WAYLONIS, G.W. (1976) Polyneuropathy due to
n-hexane. Arch. intern. Med., 136: 880-882.
PELLIN, M.C., VILANOVA, E., & BARRIL, J. (1988) Low non-neuropathic
tri- o-cresyl phosphate (TOCP) doses inhibit neuropathy target
esterase near the neuropathic threshold in n-hexane pretreated
hens. Toxicology, 49:99-105.
PELLIZZARI, E.D., HARTWELL, T.D., HARRIS, B.S.H., WADDELL, R.D.,
WHITAKER, D.A., & ERICKSON, M.D. (1982) Purgeable organic compounds
in mother's milk. Bull. environ. Contamin. Toxicol., 28: 322-328.
PERBELLINI, L. & BRUGNONE, F. (1980) Identification of the
metabolites of n-hexane, cyclohexane and their isomers in men's
urine. Toxicol. appl. Pharmacol., 53: 220-229.
PERBELLINI, L., BRUGNONE, F., PASTORELLO, G., & GRIGOLINI, L.
(1979) Urinary excretion of n-hexane metabolites in rats and
humans. Int. Arch. occup. environ. Health, 42: 349-354.
PERBELLINI, L., BRUGNONE, F., & GAFFURI, E. (1981a) Neurotoxic
metabolites of "commercial hexane" in the urine of shoe factory
workers. Clin. Toxicol., 18(12): 1377-1385.
PERBELLINI, L., BRUGNONE, F., & FAGGIONATO, G. (1981b) Urinary
excretion of the metabolites of n-hexane and its isomers during
occupational exposure. Br. J. ind. Med., 38: 20-26.
PERBELLINI, L., BRUGNONE, F., SILVESTRI, R., & GAFFURI, E. (1981c)
Measurement of the urinary metabolites of n-hexane, cyclohexane and
their isomers by gas chromatography. Int. Arch. occup. environ.
Health, 48: 99-106.
PERBELLINI, L., BRUGNONE, F., & GAFFURI, E. (1981d) [Occupational
health hazard of technical hexane: study of the biotransformation
products extracted from urine.] Ann. Ist. Sup. Sanità., 17: 615-620
(in Italian).
PERBELLINI, L., AMANTINI, M.C., BRUGNONE, F., & FRONTALI, N.
(1982a) Urinary excretion of n-hexane metabolites. A comparative
study in rat, rabbit and monkey. Arch. Toxicol., 50: 203-215.
PERBELLINI, L., LEONE, R., FRACASSO, M.E., BRUGNONE, F., &
VENTURINI, M.S. (1982b) Metabolic interaction between n-hexane and
toluene in vivo and in vitro. Int. Arch. occup. environ. Health,
50: 351-358.
PERBELLINI, L., BARTOLUCCI, G.B. BRUGNONE, F., DE ROSA, E., &
VALENTINI, F. (1985a) [2,5-Hexanedione in the biological monitoring
of occupational exposure to n-hexane.] Med. Lav., 76(1): 35-43 (in
Italian).
PERBELLINI, L., BRUGNONE, F., CARETTA, D., & MARANELLI, G. (1985b)
Partition coefficients of some industrial aliphatic hydrocarbons
(C5-C7) in blood and human tissues. Br. J. ind. Med., 42: 162-167.
PERBELLINI, L., BRUGNONE, F., MOZZO, P., DE ROSA, E., BARTOLUCCI,
B., & FACINNI, G. (1985c) Toxicokinetic aspects of n-hexane
2,5-hexanedione in the biomonitoring of occupational exposure to
n-hexane. Annu. Conf. Ind. Hyg., 12: 357-364.
PERBELLINI, L., MOZZO, P., BRUGNONE, F., & ZEDDE, A. (1986)
Physiologico-mathematical model for studying human exposure to
organic solvents: kinetics of blood/tissue n-hexane concentrations
and of 2,5-hexanedione in urine. Br. J. ind. Med., 43: 760-768.
PEROCCO, P., BOLOGNESI, S., & ALBERGHINI, W. (1983) Toxic activity
of seventeen industrial solvents and halogenated compounds on human
lymphocytes cultured in vitro. Toxicol. Lett., 16: 69-75.
PETERS, M.A., HUDSON, P.M., & DIXON, R.L. (1981) The effect of
totigestational exposure to methyl n-butyl ketone has on postnatal
development and behaviour. Ecotoxicol. environ. Saf., 5: 291-306.
PLANAS, A.M. & CUNNINGHAM, V.J. (1987) Uncoupling of cerebral
glucose supply and utilization after hexane-2,5-dione intoxication
in the rat. J. Neurochem., 48: 816-823.
PRYOR, G.T. & HOWD, R.A. (1982) Developmental long-term effects of
inhalants in animals, Washington, DC, National Institute of Drug
Abuse (NIDA Quarterly Report No. 5).
PRYOR, G.T., BINGHAM, J., DICKINSON, J., REBERT, C.S., & HOWD, R.A.
(1982) Importance of schedule of exposure to hexane in causing
neurotoxicity. Neurobehav. Toxicol. Teratol., 4: 71-78.
PRYOR, G.T., DICKINSON, J., HOWD, R.A., & REBERT, C.S. (1983)
Neurobehavioural effects of subchronic exposure of weanling rats to
toluene or hexane. Neurobehav. Toxicol. Teratol., 5: 47-52.
RABOVSKY, J. & JUDY, D.J. (1989) The in vitro effects of alkanes,
alcohols, and ketones on rat lung cytochrome P450-dependent
alkoxyphenoxazone dealkylase activities. Arch. Toxicol., 63: 13-17.
RABOVSKY, J., JUDY, D.J., & PAILES, W.H. (1986) In vitro effects
of straight-chain alkanes ( n-hexane through n-dodecane) on rat
liver and lung cytochrome P450. J. Toxicol. environ. Health, 18:
409-421.
RAITTA, C., SEPPALAINEN, A.M., & HUUSKONEN, M.S. (1978) n-Hexane
maculopathy in industrial workers. Albrecht Graefes Arch. klin.
exp. Ophthalmol., 209: 99-110.
RAJE, R.R. (1983) In vitro toxicity of n-hexane and
2,5-hexanedione using isolated perfused rabbit heart. J. Toxicol.
environ. Health, 11: 879-884.
RAJE, R.R., GREENING, M., & FINE, M.T. (1984) Blood n-hexane
concentration following acute inhalation exposure in rats. Res.
Commun. chem. Pathol. Pharmacol., 46(2): 297-300.
RALSTON, W.H., HILDERBRAND, R.L., UDDIN, D.E., ANDERSEN, M.E., &
GARDINER, R.W. (1985) Potentiation of 2,5-hexanedione neurotoxicity
by methyl ethyl ketone. Toxicol. appl. Pharmacol., 81: 319-327.
RANADIVE, K.J., GOTHOSKAR, S.V., & TEZABWALA, B.U. (1972)
Carcinogenicity of contaminants in indigenous edible oils. Int. J.
Cancer, 10: 652-666.
RAPPAPORT, S.M., SELVIN, S., & WATERS, M.A. (1987) Exposures to
hydrocarbon components of gasoline in the petroleum industry. Appl.
ind. Hyg., 2: 148-154.
REBERT, C.S. & SORENSON, S.S. (1983) Concentration-related effects
of hexane on evoked responses from brain and peripheral nerve of
the rat. Neurobehav. Toxicol. Teratol., 5: 69-76.
REBERT, C.S., HOUGHTON, P.W., HOWD, R.A., & PRYOR, G.T. (1982)
Effects of hexane on the brainstem auditory response and caudal
nerve action potential. Neurobehav. Toxicol. Teratol., 4: 79-85.
RIZZUTO, N., TERZIAN, H., & GALIAZZO-RIZZUTO, S. (1977) Toxic
polyneuropathies in Italy due to leather cement poisoning in the
shoe industries. A light and electron microscope study. J. neurol.
Sci., 31: 343-354.
RIZZUTO, N., DECRANDIS, D., DITRAPANI, G., & PASTINATO, E. (1980)
n-hexane neuropathy. An occupational disease of shoemakers. Eur.
Neurol., 19(5): 308-315.
ROBERTSON, P., WHITE, E.L., & BUS, J.S. (1982) Effect of methyl
ethyl ketone treatment on hepatic mixed function oxidase activity
and on in vivo metabolism of n-hexane. Toxicologist, 2(2): A273.
ROBERTSON, P., Jr, WHITE, E.L., & BUS, J.S. (1989) Effects of
methyl ethyl ketone pretreatment on hepatic mixed function oxidase
activity and on in vivo metabolism of n-hexane. Xenobiotica, 19:
721-729.
ROSENBERG, C.K., ANTHONY, D.C., SZAKAL-QUIN, G., GENTER, M.B., &
GRAHAM, D.G. (1987a) Hyperbaric oxygen accelerates the
neurotoxicity of 2,5-hexanedione. Toxicol. appl. Pharmacol., 87:
374-379.
ROSENBERG, C.K., GENTER, M.B., SZAKAL-QUIN, G., ANTHONY, D.C., &
GRAHAM, D.G. (1987b) dl- versus meso-3,4-dimethyl-2,5-hexanedione: a
morphometric study of the proximo-distal distribution of axonal
swellings in the anterior root of the rat. Toxicol. appl.
Pharmacol., 87: 363-373.
RUFF, R.L., PETITO, C.K., & ACHESON, L.S. (1981) Neuropathy
associated with chronic low level exposure to n-hexane. Clin.
Toxicol., 18(5): 515-519.
SABRI, M. (1984) In vitro effects of n-hexane and its metabolites
on selected enzymes in glycolysis, pentose phosphate pathway and
citric acid cycle. Brain Res., 297: 145-150.
SABRI, M., MOORE, C.L., & SPENCER, P.S. (1979) Studies on the
biochemical basis of distal axonopathies. I. Inhibition of
glycolysis by neurotoxic hexacarbon compounds. J. Neurochem., 32:
683-689.
SAHU, S.C., LOWTHER, D.E., & JONES, S.L. (1982) Biochemical
response of rat lung to inhaled n-hexane. Toxicol. Lett., 12: 13-17.
SAIDA, K., MENDELL, J.R., & WEISS, H.S. (1976) Peripheral nerve
changes induced by methyl n-butyl ketone and potentiation by methyl
ethyl ketone. J. Neuropathol. exp. Neurol., 35(3): 207-225.
SANAGI, S., SEKI, Y., SUGIMOTO, K., & HIRATA, M. (1980) Peripheral
nervous system functions of workers exposed to n-hexane at a low
level. Int. Arch. occup. environ. Health, 47: 69-79.
SANDMEYER, E.E. (1981) Aliphatic hydrocarbons. In: Clayton, G.D. &
Clayton, F.E., ed. Patty's industrial hygiene and toxicology, 3rd
ed., New York, Chichester, Brisbane, Toronto, John Wiley and Sons,
Vol. 2B, pp. 3175-3220.
SAX, N.I. (1984) Dangerous properties of industrial materials, 6th
ed., New York, Van Nostrand Reinhold Company.
SAYRE, L.M., SHEARSON, C.M., WONGMONGKOLRIT, T., MEDORI, R., &
GAMBETTI, P. (1986) Structural basis of c-diketone neurotoxicity:
non-neurotoxicity of 3,3-dimethyl-2,5-hexanedione, a c-diketone
incapable of pyrolle formation. Toxicol. appl. Pharmacol., 84: 36-44.
SCELSI, R., POGGI, P., FERA, L., & GONELLA, G. (1980) Toxic
polyneuropathy due to n-hexane. J. neurol. Sci., 47: 7-19.
SCELSI, R., POGGI, P., FERA, L., & GONELLA, G. (1981) Industrial
neuropathy due to n-hexane and 2,5-hexanedione. Clinical and
morphological findings in three cases. Clin. Toxicol., 18: 1387-1393.
SCHAUMBURG, H.H. & SPENCER, P.S. (1976) Degeneration in central and
peripheral nervous systems produced by pure n-hexane: an
experimental study. Brain, 99: 183-192.
SCHAUMBURG, H.H. & SPENCER, P.S. (1979) Clinical and experimental
studies of distal axonopathy. A frequent form of brain and nerve
damage produced by environmental chemical hazards. Ann. NY Acad.
Sci., 329: 14-29.
SCHNOY, N., SCHMIDT, R., ALTENKIRCH, H., & WAGNER, H.M. (1982)
Ultrastructural alteration of alveolar epithelium after exposure
to organic solvents. Respiration, 43: 221-231.
SCHMIDT, R., SCHNOY, N., ALTENKIRCH, H., & WAGNER, H.M. (1984)
Ultrastructural alteration of intrapulmonary nerves after exposure
to organic solvents. Respiration, 46: 362-369.
SELKOE, D.J., LUCKENBILL-EDDS, L., & SHELANSKI, M.L. (1978) Effects
of neurotoxic industrial solvents on cultured neuroblastoma cells:
methyl n-methyl ketone, n-hexane and derivatives. J. Neuropathol.
exp. Neurol., 37: 768-789.
SEPPALAINEN, A. & RAITTA, C. (1981) Neurotoxic properties of
n-hexane among occupationally exposed workers. In: Proceedings of
the 2nd Finnish-Estonian Symposium on Early Effects of Toxic
Substances, Helsinki, June 1981, Helsinki, Institute of
Occupational Health, pp. 180-184.
SEPPALAINEN, A., RAITTA, C., & HUUSKONEN, M. (1979) n-Hexane
induced changes in visual evoked potentials and electroretinograms
of industrial workers. Electroencephalogr. clin. Neurophysiol., 47:
492-498.
SHIRBAE, T., TSUDA, T., TERAO, A., & ARAKI, S. (1974) Toxic
polyneuropathy due to glue sniffing. J. neurol. Sci., 21: 101-113.
SICE, J. (1966) Tumour-promoting activity of n-alkanes and
1-alkanols. Toxicol. appl. Pharmacol., 9: 70-74.
SINGH, D. & JAIN, D.C. (1987) Relative toxicity of various organic
solvents generally used in screening plant products for
insecticidal activity against the house fly Musca domestica L.
Indian J. exp. Biol., 25: 569-570.
SINGH, K.P., KANNAN, K., GOEL, S.K., PANDYA, K.P., & SHANKER, R.
(1983) 2,5-Hexanediol induced thymic atrophy and lymphocytotoxicity
in rats. Ind. Health, 21: 235-242.
SINGH, K.P., UPRETI, R.K., SAXENA, A.K., & SHANKER, R. (1984)
Thymic toxicity produced by 2,5-hexanediol and 2,5-hexanedione in
rats. In: Proceedings of the International Conference on Organic
Solvents Toxicity, Stockholm, October 1984, p. 144 (Abstract 550).
SOBUE, I. & YAMAMURA, Y. (1968) [ n-Hexane polyneuropathy. Outbreak
among vinyl sandal manufacturers.] Rinsho Shinkei, 8: 393-403 (in
Japanese).
SOBUE, I., IIDA, M., YAMAMURA, Y., & TAKAYANAGUI, T. (1978)
n-Hexane polyneuropathy. Int. J. Neurol., 11(4): 317-330.
SPAGNA, C., PARLATO, G., & TIMPANARO, V. (1967) [Behaviour of blood
picture in experimental hexane intoxication.] Folia med. (Napoli),
50: 977-985 (in Italian).
SPENCER, P. (1981) Morphological study of n-hexane intoxication:
Final report, Washington, DC, American Petroleum Institute
(Attachment to API Medical Research Publication No. 28-30077).
SPENCER, P. (1982) Neuropathic potential of n-hexane in the
presence of other hexane isomers, Washington, DC, American
Petroleum Institute (API Medical Research Publication No.
30-30226).
SPENCER, P.S. & SCHAUMBURG, H.H. (1977a) Ultrastructural studies of
the dying-back process. III. The evolution of experimental
peripheral giant axonal degeneration. J. Neuropathol. exp. Neurol.,
36: 276-299.
SPENCER, P.S. & SCHAUMBURG, H.H. (1977b) Ultrastructural studies of
the dying-back process. IV. Differential vulnerability of PNS and
CNS fibres in experimental central-peripheral distal axonopathies.
J. Neuropathol. exp. Neurol., 36: 300-320.
SPENCER, P.S., SCHAUMBURG, H.H., SABRI, M.I., & VERONES, B. (1980)
The enlarging view of hexacarbon neurotoxicity. CRC crit. Rev.
Toxicol., 17(4): 279-356.
STOLTENBURG-DIDINGER, G., WAGNER, H.M., & ALTENKIRCH, H.
(1984) Embryo and fetotoxicity of n-hexane and methyl ethyl ketone.
In: Proceedings of the International Conference on Organic Solvent
Toxicity, Stockholm, October 1984, p. 125 (Abstract 405).
STRATTON, G.W. & SMITH, T.M. (1988) Interaction of organic solvents
with the green alga Chlorella pyrenoidosa. Bull. environ. Contam.
Toxicol., 40: 736-742.
SUZUKI, T., SHIMBO, S., & NISHITANI, H. (1974) Muscular atrophy due
to glue sniffing. Int. Arch. Arbeitsmed., 33: 115-123.
SVABOVA, E., ZORAD, S., MITKOVA, A., KLIMES, I., & MACHO, L. (1987)
[Increase of insulin binding and degradation on isolated human
erythrocytes by n-heptane and n-hexane; effects of trypsin and
dithiotreitol.] Bratisl. lek. Listy., 88(6): 615-624 (in Slovak).
SWANN, H.E., KWON, B.K., HOGAN, G.K., & SNELLINGS, W.M. (1974)
Acute inhalation toxicology of volatile hydrocarbons. Am. Ind. Hyg.
Assoc. J., 35: 511-518.
TAIRA, M. (1975) [Experimental studies on pathomorphological
changes of lung produced by aliphatic and aromatic hydrocarbons and
gasoline.] J. Tokyo med. Univ., 33(4): 597-607 (in Japanese).
TAKAHASHI, M., TAKEUCHI, H., KYO, S., YORIFUJI, S., SANAGI, SL.,
SEKI, Y., & HARA, I. (1977) n-Hexane polyneuropathy: a case report
with review of literature. Med. J. Osaka Univ., 28(1): 77-85.
TAKEUCHI, Y., MABUCHI, C., & TAKAGI, S. (1975) Polyneuropathy
caused by petroleum benzine. Int. Arch. Arbeitsmed., 34: 185-197.
TAKEUCHI, Y., ONO, Y., HISANAGA, N., KITOH, J., & SUGIURA, Y.
(1980) A comparative study on the neurotoxicity of n-pentane,
n-hexane and n-heptane in the rat. Br. J. ind. Med., 37: 241-247.
TAKEUCHI, Y., ONO, Y., & HISANAGA, N. (1981) An experimental study
on the combined effects of n-hexane and toluene on the peripheral
nerve of the rat. Br. J. ind. Med., 38: 14-19.
TAKEUCHI, Y., ONO, Y., HISANAGA, N., IWATA, M., AOYAMA, M., KITOH,
J., & SUGIURA, Y. (1983) An experimental study of the combined
effects of n-hexane and methyl ethyl ketone. Br. J. ind. Med., 40:
199-203.
THAM, R., BUNNFORS, I., ERIKSSON, B., LARSBY, B., LINDGREN, S., &
ODKVIST. L. (1984) Vestibulo-ocular disturbances in rats exposed to
organic solvents. Acta pharmacol. toxicol., 54: 58-63.
TIL, H.P., FALKE, H.E., BEEMS, R.B., BRUIJNTJES, J.P., & DE VOGEL,
N. (1989) Subchronic (90-day) oral toxicity study in rats,
including metaphase chromosomal analysis of bone marrow cells, with
light petroleum solvent (technical hexane) for oil seed extraction:
Final report, Zeist, The Netherlands, TNO-CIVO Institutes, 166 pp
(Project No. B/88-0541) (Report No. V/89.089).
TOFTGARD, R., NILSEN, O.G., & GUSTAFSSON, J.A. (1981) Changes in
rat liver microsomal cytochrome P-450 and enzymatic activities
after the inhalation of n-hexane, xylene, methyl ethyl ketone and
methyl chloroform for four weeks. Scand. J. Work Environ. Health,
7: 31-37.
TOFTGARD, R., NILSEN, O.G., GLAUMANN, H., & GUSTAFSSON, J.A. (1983)
Induction of cytochrome P-450 in the rat liver after exposure to
xylenes. Dose response relationship and dependence on endocrine
factors. Toxicology, 27: 119-137.
TOFTGARD, R., HAAPARANTA, T., ENG, L., & HALPERT, J. (1984) Rat
lung and liver microsomal cytochrome P-450 isozymes involved in the
hydroxylation of n-hexane and m-xylene. In: Abstracts of the
International Conference on Organic Solvent Toxicity, Stockholm,
October 1984, p. 17 (Abstract 111).
TOFTGARD, R., HAAPARANTA, T., ENG, L., & HALPERT, J. (1986) Rat
lung and liver microsomal cytochrome P-450 isozymes involved in the
hydroxylation of n-hexane. Biochem. Pharmacol., 35: 3733-3738.
TOWFIGHI, J., NICHOLAS, K., GONATAS, M.D., PLEASURE, D., COOPER,
H.S., & MCCREE, L. (1976) Glue sniffer's neuropathy. Neurology, 26:
238-243.
TOXIGENICS (1982) 90-Day vapour inhalation toxicity study on
n-hexane in albino rats (Report submitted to Chemical Industry
Institute of Toxicology, North Carolina) (CIIT Pocket No. 10242).
TSURUTA, H. (1977) Percutaneous absorption of organic solvents. II.
A method for measuring the penetration rate of chlorinated solvents
through excised rat skin. Ind. Health, 15: 131-139.
TSURUTA, H. (1980) Microdetermination of n-hexane in biological
samples by gas chromatography - mass spectroscopy. Ind. Health, 18:
113-114.
TSURUTA, H. (1982) Percutaneous absorption of organic solvents.
III. On the penetration rates of hydrophobic solvents through the
excised rat skin. Ind. Health, 20: 335-345.
UPRETI, R.K., SINGH, K.P., SAXENA, A.K., & SHANKER, R. (1986)
Effect of 2,5-hexanedione on lymphoid organs in rats: a preliminary
report. Environ. Res., 39: 188-198.
USITC (1988) Synthetic organic chemicals. United States Production
and Sales 1987, Washington, DC, United States International Trade
Commission, pp. 2-7 (USITC Publication No. 2009).
VAINIO, H. (1974) Activation and inactivation of membrane-bound
UDP-glucuronosyltransferase by organic solvents in vitro. Acta
pharmacol. toxicol., 34: 152-156.
VDI (Verband Deutscher Ingenieure) (1984) Handbook of clean air,
Berlin, Benth Verlag Gmbh, Vol. 5 (Guideline 3482).
VERONESI, B., PETERSON, E.R., DI VINCENCO, G., & SPENCER, P.S.
(1978) A tissue culture model of distal (dying-back) axonopathy:
its use in determining primary neurotoxic hexacarbon compounds. J.
Neuropathol. exp. Neurol., 33: 703.
VERONESI, B., PETERSON, E.R., & SPENCER, P.S. (1980) Reproduction
and analysis of MnBK neurotoxicity in organotypic tissue culture.
In: Spencer, P.S. & Schaumburg, H.H., ed. Experimental and clinical
neurotoxicology, Baltimore, Maryland, Williams and Wilkins, pp.
863-871.
VERONESI, B., PETERSON, E.R., BORNSTEIN, M., & SPENCER, P.S. (1983)
Ultrastructural studies of the dying-back process. VI. Examination
of nerve fibres undergoing giant axonal degeneration in organotypic
culture. J. Neuropathol. exp. Neurol., 24(2): 153-165.
VERONESI, B., LINGTON, A.W., & SPENCER, P.S. (1984) A tissue
culture model of methyl ethyl ketone's potentiation of n-hexane
neurotoxicity. Neurotoxicology, 5(2): 43-52.
VEULEMANS, H., VAN VLEM, E., JANSSENS, H., MASSCHELEIN, R., &
LEPLAT, A. (1982) Experimental human exposure to n-hexane. Study of
the respiratory uptake and elimination, and of n-hexane
concentrations in peripheral venous blood. Int. Arch. occup.
environ. Health, 49: 251-263.
VEULEMANS, H., GROESENEKEN, D., MASSCHELEIN, R., & VLEM, E.V.
(1987) Survey of ethylene glycol ether exposures in Belgian
industries and workshops. Am. Ind. Hyg. Assoc. J., 48: 671-676.
VICEDO, J.L., PELLIN, M., & VILLANOVA, E. (1985) Phthalates and
organophosphorus compounds as cholinesterase inhibitors in
fractions of industrial hexane impurities. Arch. Toxicol., 57: 46-52.
VILANOVA, E. & VICEDO, J.L. (1983) Serum cholinesterase inhibitors
in the commercial hexane impurities. Arch. Toxicol., 53: 59-69.
WADA, Y., OKAMOTO, S., & TAKAGI, S. (1965) [Intoxication
polyneuropathy following exposure to n-hexane.] Rinsho Shinkeigaku,
10(5): 591-597 (in Japanese).
WAGNER, H.M., STEPPART, R., & ALTENKIRCH, H. (1984) [Animal
experiments on the neurotoxicity of organic solvents in rats
challenged with heavy metals.] Schriftenr. Ver. Wasser Boden
Lufthyg., 59: 191-200 (in German).
WAHLBERG, J.E. (1984a) Erythema-inducing effects of solvents
following epicutaneous administration to man: studied by laser
Doppler flowmetry. Scand. J. Work Environ. Health, 10: 159-162.
WAHLBERG, J.E. (1984b) Edema inducing effects of solvents following
topical administration. Derm. Beruf Umwelt, 32(3): 91-94.
WAHLBERG, J.E. & BOMAN, A. (1979) Comparative percutaneous toxicity
of ten industrial solvents in the guinea-pig. Scand. J. Work
Environ. Health, 5: 345-351.
WALSETH, F. & NILSEN, O.G. (1984) Phthalate esters. II. Effects of
inhaled dibutylphthalate on cytochrome P-450 mediated metabolism in
rat liver and lung. Arch. Toxicol., 55: 132-136.
WALSETH, F., TOFTGARD, R., & NILSEN, O.G. (1982) Phthalate esters.
I. Effects on cytochrome P-450 mediated metabolism in rat liver and
lung, serum enzymatic activities and serum protein levels. Arch.
Toxicol., 50: 1-10.
WANG, J.D., CHANG, Y.C., KAO, K.P., HUANG, C.C., LIN, C.C., & YEH,
W.Y. (1986) An outbreak of n-hexane induced polyneuropathy among
press proofing workers in Taipei. Am. J. ind. Med., 10: 111-118.
WEAST, R.C. (1981) CRC handbook of chemistry and physics, 6th ed.,
Boca Raton, Florida, CRC Press.
WHITE, E.L. & BUS, J.S. (1980) Determination of n-hexane, methyl
ethyl ketone and their metabolites in sciatic nerves of rats by gas
chromatography-mass spectrometry. In: Abstracts of the Pittsburgh
Conference on Analytical Chemistry and Applied Spectroscopy, 10-14
March, 1980, Pittsburgh, Pennsylvania, US Society of Analytical
Chemistry (Abstract No. 525).
WHITE, E.L., BUS, J.S., & HECK, H. (1979) Simultaneous
determination of n-hexane, 2-hexanone and 2,5-hexanedione in
biological tissues by gas chromatography mass spectrometry. Biomed.
mass Spectrom. 6(4): 169-172.
WHO (1971) Evaluation of food additives. Fourteenth Report of the
Joint FAO/WHO Expert Committee on Food Additives, Geneva, World
Health Organization, pp. 21-24 (Technical Report Series No. 462).
YAMADA, N. (1964) [Polyneuritis in workers handling n-hexane.]
Sangyo Igaku, 6: 192 (in Japanese).
YAMADA, S. (1967) [Intoxication polyneuritis in workers exposed to
n-hexane.] Jpn. J. ind. Health, 9: 651-659 (in Japanese).
YAMAKAGE, A. & ISHIKAWA, H. (1982) Generalised morphea-like
scleroderma occurring in people exposed to organic solvents.
Dermatologia, 165: 186-193.
YAMAMURA, Y. (1969) n-Hexane polyneuropathy. Folia psychiatr.
neurol. Jpn., 23(1): 45-57.
YOSHIDA, T., YANAGISAWA, H., MUNEYUKI, T., & SHIGIYA, R. (1974) [An
electrophysiological study of n-hexane polyneuropathy.] Rinsho
Shinkeigaku, 14(5): 454-461 (in Japanese).
ZAHLSEN, K., AARSTAD, K., & NILSEN, O.G. (1985) Inhalation of
isopropanol. Induction of activating and deactivating enzymes in
rat kidney and liver. Increased microsomal metabolism of n-hexane.
Toxicology, 34: 57-66.
RESUME
Le n-hexane (hexane normal) est un liquide volatil incolore.
L'hexane du commerce se présente essentiellement sous la forme d'un
mélange de divers isomères de l'hexane et de dérivés apparentés à
six atomes de carbone; sa teneur en n-hexane varie entre 20 et 80%.
La chromatographie en phase gazeuse avec détection par ionisation
de flamme ou couplage à un spectrographe de masse constitue une
bonne technique pour le dosage du n-hexane. Les limites
d'exposition professionnelle varient de 100 à 1800 mg/m3 (moyenne
pondérée par rapport au temps, TWA) et de 400 à 1500 mg/m3 (valeur
plafond, CLV) selon les pays.
On peut isoler le n-hexane du gaz naturel et du pétrole brut.
On l'utilise dans les industries de transformation alimentaire,
notamment pour l'extraction des huiles végétales ou comme solvant
dans différents produits et procédés.
Une fois libéré dans l'environnement, le n-hexane y subsiste
essentiellement en phase vapeur. Sa demi-vie dans l'atmosphère est
estimée à environ deux jours, si l'on se base uniquement sur sa
réactivité vis-à-vis du radical OH. Les valeurs de la CL50 pour les
organismes aquatiques sont rares et dispersées; elles ont été
obtenues dans des conditions peu satisfaisantes. Dans ces
conditions il n'est pas possible d'apprécier les effets toxiques du
n-hexane dans l'environnement. Du fait de sa faible solubilité
dans l'eau et de sa forte volatilité il est peu probable que les
organismes aquatiques courent un risque d'exposition, sauf en cas
de décharge sauvage dans les eaux superficielles.
Chez les mammifères, le n-hexane est rapidement résorbé au
niveau des poumons et se distribue largement dans l'organisme de
l'adulte ainsi que dans les tissus du foetus. Au niveau cutané, la
résorption est faible. Le n-hexane subit une métabolisation
oxydative en un certain nombre de dérivés, notamment l'hexane-2,5-
dione dont on pense qu'elle constitue l'agent neurotoxique final.
Dans le nerf sciatique de rats d'expérience on a pu trouver des
concentrations particulièrement élevées de n-hexane et d'hexane-
2,5-dione. La majeure partie du n-hexane est excrétée sans
modification dans l'air exhalé; une certaine quantité est également
excrétée sous forme de métabolites dans l'air expiré et dans les
urines.
Le n-hexane présente une faible toxicité aiguë pour le rat
adulte après administration par voie orale ou inhalation. On a noté
des valeurs de la DL50 par voie orale allant de 15 à 30 g/kg; quant
à la CL50 par inhalation, elle serait de 271 040 mg/m3 (77 000 ppm)
pour une exposition d'une heure. Lorsque la teneur en vapeurs est
élevée, on constate chez l'animal de l'ataxie, des convulsions et
les signes d'une dépression du système nerveux central.
Il semble que chez le rat, les principaux effets d'une
exposition répétée au n-hexane consistent en lésions testiculaires
et signes de neurotoxicité. C'est ainsi que l'inhalation de
n-hexane et l'ingestion d'hexane-2,5-dione ont produit de graves
lésions testiculaires chez les animaux d'expérience. Ces effets ont
été attribués à la désagrégation du cytosquelette des cellules de
Sertoli. On note des effets secondaires sur les cellules germinales
des stades postérieurs aux spermatogonies, qui disparaissent des
tubules endommagés. Après une seule exposition de 24 heures à
17 600 mg/m3 de n-hexane (5000 ppm) les effets testiculaires
étaient réversibles mais ils devenaient irréversibles après deux
semaines d'exposition à la même concentration, 16 heures par jour,
six jours par semaine. A la concentration de 1% dans l'eau de
boisson, l'hexane-2,5-dione a produit des lésions testiculaires
réversibles du même type au bout de deux à trois semaines
d'administration et des effets irréversibles (en l'espace de 17
semaines) au bout de cinq semaines d'administration.
L'effet neurotoxique se caractérise cliniquement par une
faiblesse des pattes arrières qui peut évoluer vers la paralysie.
Il se produit un gonflement de l'axone aux niveaux central et
périphérique avec parfois des lésions plus sévères (dégénérescence
et disparition de l'axone), notamment au niveau des troncs nerveux
de plus grande longueur et de plus gros diamètre. Lors d'études
comportant principalement l'inhalation d'hexane pendant six mois,
on a noté la présence de lésions du système nerveux central et
périphérique aux doses de 1760 mg/m3 (500 ppm) ou davantage; en
revanche il n'y avait pas d'effets cliniques ou
anatomopathologiques à la dose de 400 mg/m3 (125 ppm). Quinze à 22
semaines après l'arrêt de l'exposition, cinq jours par semaine
pendant 11 semaines à des vapeurs de n-hexane à la concentration de
3520 mg/m3 (1000 ppm), on a noté que l'amplitude des potentiels
évoqués auditifs du tronc cérébral (PEAP) n'était que partiellement
restaurée. L'exposition discontinue de rats à une concentration de
3168 mg/m3 (900 ppm) pendant 72 semaines n'a pas provoqué de
lésions apparentes aux niveaux périphérique ou central mais
certains effets électrophysiologiques étaient perceptibles au
niveau des nerfs périphériques.
La neurotoxicité induite par le n-hexane peut être
potentialisée en présence de méthyléthylcétone, de
méthylisobutylcétone et d'acétate de plomb et réduite par
exposition simultanée au toluène. La perturbation des
concentrations de dopamine est également synergisée par une
exposition simultanée au toluène et au n-hexane.
Des lésions microscopiques graves ont été observées sur la peau
après application de n-hexane et pose d'un pansement occlusif
après une brève période. L'exposition prolongée à des vapeurs de
n-hexane à une concentration de 1560 mg/m3 (3000 ppm) peut
provoquer chez le rat une irritation des conjonctives et une forte
irritation oculaire chez le lapin. On ne dispose d'aucune donnée
sur la sensibilisation cutanée provenant d'études sur l'animal.
Des lésions chromosomiques (polyploïdie dans le cas d'une
étude, et aberrations de structure dans le cas d'une autre) ont été
signalées tant in vitro qu' in vivo. On n'a pas observé
d'augmentation de la fréquence des mutations ponctuelles ni de
signes de lésion de l'ADN.
Une étude de cancérogénicité a été effectuée sur des souris
(badigeonnage de la peau avec du n-hexane): elle n'a révélé aucun
signe de cancérogénicité.
Les effets du n-hexane sur la fonction de reproduction n'ont
pas été suffisamment étudiés. Aucun signe important
d'embryotoxicité ou de tératogénicité n'a été observé chez des rats
après inhalation de n-hexane à des concentrations relativement
basses ni chez des souris après administration par voie orale. On a
constaté un retard transitoire du développement postnatal chez des
ratons après exposition des femelles gravides à des vapeurs de
n-hexane à des concentrations de 3520 mg/m3 (1000 ppm).
On ne possède guère de données sur la toxicité aiguë du
n-hexane pour l'être humain. La plupart des études portent sur
l'exposition professionnelle à des mélanges de solvants. D'après
les données disponibles, il semble que le n-hexane n'ait qu'une
faible toxicité aiguë. On a signalé des signes de dépression du
système nerveux central, tels que somnolence, vertiges, sensation
ébrieuse, après exposition à de l'hexane du commerce pendant 10 à
60 minutes à des concentrations allant de 3520 à 17 600 mg/m3
(1000 - 5000 ppm).
Le n-hexane est légèrement irritant et provoque un érythème
passager chez l'homme après un court contact cutané. Des effets
plus graves (érythème et phlyctènes) ont été enregistrés après un
contact de cinq heures avec du n-hexane de qualité commerciale, la
zone de contact étant recouverte. Aucun cas de sensibilisation
cutanée n'a été signalé chez les ouvriers exposés et un test de
sensibilisation cutanée maximale effectué avec du n-hexane a donné
des résultats négatifs.
En cas d'exposition répétée, le n-hexane entraîne une
polynévrite sensitivo-motrice périphérique. De nombreuses études
sur la prévalence des effets neurotoxiques dus au n-hexane ont été
publiées; toutefois on manque bien souvent de données
satisfaisantes sur l'exposition. Des cas de neuropathie ont été
attribués à l'exposition à des concentrations atmosphériques de
n-hexane allant de 106 à 8800 mg/m3 (30 - 2500 ppm). Des cas de
neuropathie périphérique marquée ont été signalés parmi des
ouvriers japonais travaillant à la fabrication de sandales ainsi
que chez des opérateurs taïwanais de presse à épreuves exposés à
des teneurs en n-hexane d'environ 76 et 352 mg/m3 (50 et 100 ppm),
respectivement, pendant des périodes supérieures à huit heures par
jour. Dans la plupart des cas, les mesures d'exposition étaient
récentes et ne reflétaient probablement pas très exactement les
expositions antérieures susceptibles d'être à l'origine de la
neuropathie en question.
Plusieurs études transversales ont fait état, indépendamment
les unes des autres, d'effets infracliniques légers (par exemple
des altérations électrophysiologiques au niveau des nerfs
périphériques) chez des travailleurs exposés à des concentrations
de 70 - 352 mg/m3 (20 - 100 ppm). Aucun cas bien défini de
neurophathie périphérique cliniquement manifeste n'a pu être relevé
dans l'une quelconque de ces études à des niveaux d'exposition
inférieurs à 352 mg/m3 (100 ppm).
Seuls quelques travaux ont été consacrés à l'étude des effets
du n-hexane sur le système nerveux central. On pense que les
modifications observées dans les potentiels évoqués
somatosensoriels d'ouvriers exposés à du n-hexane proviennent d'un
bloc de la conduction nerveuse centrale. On a également noté une
altération des potentiels évoqués visuels et du tracé électro-
encéphalographique. Ces résultats incitent à penser que le
n-hexane pourrait entraîner des troubles du système nerveux
central, mais les données disponibles ne donnent aucune indication
sur les niveaux d'exposition en cause.
EVALUATION DES RISQUES POUR LA SANTE HUMAINE ET DES EFFETS SUR
L'ENVIRONNEMENT
1. Etudes sur l'animal
1.1 Exposition à court et à long terme
Le n-hexane présente une faible toxicité aiguë puisque l'on
fait état de valeurs de la DL50 comprises entre 15 et 30 g/kg et
d'une valeur de la CL50 à une heure de 271 040 mg.
Lors d'études consistant essentiellement à faire inhaler du
n-hexane à des rats de manière continue, on a observé une
mortalité chez ces animaux après exposition à une concentration de
3520 mg/m3 (1000 ppm) pendant des durées allant jusqu'à 11
semaines, ainsi qu'une diminution du gain de poids après trois
semaines d'exposition à 1760 mg/m3 (500 ppm). Lors d'études
comportant des expositions quotidiennes de 12 heures à du
n-hexane, on a enregistré une mortalité chez les animaux à la
concentration de 10 560 mg/m3 (3000 ppm) au bout de 15 semaines et
une légère réduction du gain de poids corporel à la concentration
de 740 mg/m3 (200 ppm) au bout de 24 semaines. Aucun effet sur le
poids corporel des rats n'a été observé après une exposition
pratiquement continue à 443 mg/m3 (126 ppm) pendant 34 semaines.
Il semble que les lésions testiculaires et les effets
neurotoxiques aux niveaux périphérique et central soient les
principaux effets non mortels d'une exposition répétée au n-hexane
chez le rat. Après inhalation de n-hexane et ingestion de hexane-
2,5-dione on a observé de graves lésions testiculaires. Ces effets
ont été attribués à la désagrégation du cytosquelette des cellules
de Sertoli. On observe également des effets secondaires sur les
cellules germinales des stades qui suivent la spermatogonie et qui
disparaissent des tubules endommagés. Après une seule exposition de
24 heures à 17 600 mg/m3 (5000 ppm), les effets testiculaires
étaient réversibles mais ils devenaient irréversibles après deux
semaines d'exposition à la même concentration 16 heures par jour,
six jours par semaine. De l'hexane-2,5-dione administrée dans l'eau
de boisson à la concentration de 1% a produit des lésions
testiculaires réversibles au bout de deux à trois semaines
d'administration et des effets irréversibles (en l'espace de 17
semaines) au bout de cinq semaines d'administration.
Du point de vue clinique, la neurotoxicité se caractérise par
une faiblesse des pattes arrières, qui peut évoluer vers la
paralysie et aboutir à une quadriplégie. Ces signes cliniques ont
été observés après dix semaines d'exposition discontinue à 10 560
mg/m3 (3000 ppm) (12 heures par jour et sept jours par semaine) et
au bout de six à dix semaines d'exposition continue à des doses
allant de 1048 - 2112 mg/m3 (400 à 600 ppm).
Avant que ne se manifeste la faiblesse des pattes arrières on a
observé des lésions (gonflement de l'axone) au niveau des nerfs des
pattes arrières, du cerveau et de la moëlle épinière. Des effets
plus graves (dégénérescence et disparition de l'axone) ont été
notés en particulier au niveau des troncs nerveux de plus grande
longueur et plus grand diamètre au bout de six mois d'exposition à
1760 mg/m3 (500 ppm). Au total, il est difficile d'établir la
valeur de la dose sans effet nocif observable dans le cas des
effets neurotoxiques provoqués par le n-hexane chez le rat, en
raison de la diversité des concentrations et des modalités
d'exposition expérimentées ainsi que de la variété des effets
étudiés. Lors d'études de six mois consistant essentiellement à
faire inhaler en continu des vapeurs de n-hexane, on a observé des
lésions nerveuses à des concentrations de 1760 mg/m3 (500 ppm ou
davantage), toutefois aucun effet clinique ni anatomopathologique
n'a été constaté à la dose de 440 mg/m3 (125 ppm). En mode
discontinu l'exposition de rats à la dose de 3168 mg/m3 (900 ppm)
pendant 72 semaines n'a pas provoqué de lésions nerveuses
évidentes, mais on a pu constater les premiers signes d'effets sur
la force de préhension.
Après arrêt de l'exposition il peut y avoir résolution
(partielle ou complète) des signes cliniques de toxicité, de
l'amoindrissement de la force de préhension et des effets
électrophysiologiques. Dans les 15 à 22 semaines qui suivent
l'arrêt d'une exposition répétée à la concentration de 3520 mg/m3
(1000 ppm) de n-hexane pendant 11 semaines, on constate que
l'amplitude de l'onde cinq des potentiels évoqués auditifs du tronc
cérébral (qui est supposée refléter l'activité du système nerveux
central) ainsi que le potentiel d'action du nerf caudal, ne sont
pas complètement rétablis. On n'a pas encore étudié
systématiquement la résolution des lésions anatomopathologiques
consécutives à une exposition par inhalation, mais on a toutefois
observé que lorsqu'on cesse d'exposer les animaux à des
concentrations allant de 1408 à 2112 mg/m3 (400 à 600 ppm), il y a
remyélinisation et régénération limitées des axones.
L'inhalation en discontinu pendant sept semaines de n-hexane à
la concentration de 1760 mg/m3 (500 ppm) peut provoquer de petites
lésions au niveau des poumons chez le rat. Chez le lapin, on a
observé de graves lésions de l'arbre respiratoire après exposition
à une concentration de 10 560 mg/m3 (3000 ppm), huit heures par
jour pendant huit jours, et on a noté une prolifération papillaire
des cellules broncholiaires non ciliées au bout de 24 semaines
d'exposition.
On a fait état d'effets sur le rein et le foie à la suite de
quelques études d'inhalation sur des rats mais seulement aux doses
les plus fortes. Quant à la plupart des autres lésions et des
autres effets, on a estimé qu'ils étaient d'une importance
toxicologique mineure ou constaté qu'ils n'étaient pas
reproductibles. Cependant, on a observé de graves lésions
hépatiques chez des rats après administration intrapéritonéale
répétée de n-hexane.
Des études portant sur l'exposition à plusieurs solvants ont
montré que les effets neurotoxiques de la vapeur de n-hexane
pouvaient être accrus chez des poules par la présence de méthyl-
isobutylcétone et chez des rats par la présence simultanée de
méthyl-isobutylcétone et d'acétate de plomb; toutefois l'effet
était au contraire réduit en présence de toluène. On a également
indiqué que les lésions pulmonaires observées chez les rats
augmentaient en cas d'exposition simultanée à la méthyléthylcétone
et à l'hexane.
Peu d'études ont été consacrées aux effets de l'ingestion de
n-hexane ou à ceux de l'exposition cutanée à cette substance.
L'administration quotidienne par voie orale de 500 mg/kg de
n-hexane pendant quatre semaines à des rats a entraîné une baisse
de la vitesse de conduction du nerf caudal.
1.2 Génotoxicité
Le n-hexane a fait l'objet de tests de mutagénicité limités.
On a fait état de lésions chromosomiques (polyploïdie dans le cas
d'une étude et aberrations structurales dans le cas d'une autre)
lors d'essais in vitro au cours desquels une lignée cellulaire
provenant de hamsters chinois a été exposée à du n-hexane liquide
non dilué en l'absence de fraction métabolisante. Dans le cas de
cellules mammaliennes en culture ou de bactéries, on n'a pas
observé d'augmentation significative du taux de mutations
ponctuelles. L'exposition au n-hexane n'augmente pas les lésions
de l'ADN comme on a pu en juger par des mesures de toxicité
bactérienne différentielle. De même il n'y a pas d'augmentation de
la synthèse non programmée d'ADN dans les lymphocytes humains in
vitro. Bien que les tests portant sur les mutations ponctuelles
aient donné des résultats négatifs, il semble, à certains signes,
que le n-hexane soit capable de produire des aberrations
chromosomiques in vitro. Des lésions chromosomiques (rupture des
chromatides) ont été signalées à l'occasion d'une étude in vivo
sur des cellules de moëlle osseuse de rat.
1.3 Cancérogénicité
Le pouvoir cancérogène du n-hexane n'a pas été suffisamment
étudié. Une étude de portée limitée a montré que l'application
cutanée de n-hexane ne semblait pas accroître la formation de
tumeurs cutanées chez la souris après exposition à un initiateur
tumoral (diméthyl-7,12 benzanthracène) ou après exposition
simultanée à un agent promoteur (huile de croton).
1.4 Effets sur la reproduction
Les effets toxiques du n-hexane sur la fonction de
reproduction n'ont pas été convenablement étudiés. Il n'existe pas
de preuves substantielles que le n-hexane entraîne des effets
embryotoxiques ou tératogènes après inhalation chez le rat ou
ingestion chez la souris. Chez des rats exposés à une dose de
10 560 mg/m3 (3000 ppm), on a observé une légère augmentation de
l'incidence de certaines anomalies, toutefois en raison
d'insuffisances dans la manière dont l'étude a été conduite (par
exemple concentration probablement trop faible) les conclusions
qu'on peut en tirer demeurent limitées. Des signes de toxicité
foetale (réduction du poids des souriceaux) ont été observés mais
seulement à des doses qui entraînaient une mortalité importante
chez les femelles gravides. Une des études effectuées a mis en
évidence un retard passager dans le développement postnatal de
ratons exposés à la concentration de 10 560 mg/m3 (3000 ppm). Aucun
signe de neuropathie n'a été signalé chez ces animaux mais on n'est
pas en mesure, sur la base des résultats de cette étude, de se
prononcer sur la possibilité que l'exposition in utero au
n-hexane induise effectivement une neuropathie, car les animaux
n'ont été exposés à cette substance que durant une courte fraction
de la période de gestation. Selon d'autres rapports, l'exposition
in utero au n-hexane retarderait la genèse du tissu cérébelleux
ainsi que la maturation des neurones et produirait des anomalies
électrophysiologiques au niveau central chez les rats nouveaux-nés.
Cependant on ne possède pas suffisamment de détails sur ces
travaux pour en apprécier toute la portée.
On est fondé à penser que le n-hexane entraîne une stérilité
irréversible chez le rat. Une étude limitée de dominance létale
chez la souris n'a pas révélé d'effets sur la fertilité des mâles.
Aucune étude n'a été publiée sur la fertilité des femelles de
divers animaux ou sur les effets portant sur plusieurs générations.
2. Etudes sur l'homme
2.1 Effets sur le système nerveux
Selon de nombreuses études un certain nombre de cas humains de
neuropathie périphérique seraient imputables à une exposition
professionnelle au n-hexane. Toutefois les concentrations
atmosphériques sont rarement mentionnées et quand elles le sont,
elles peuvent très bien être inexactes, sans compter qu'il n'est
pas toujours facile de savoir s'il s'agit d'hexane ou de n-hexane.
En outre, l'exposition au n-hexane en milieu industriel se produit
généralement lors de l'utilisation de produits contenant divers
solvants, de sorte que les ouvriers sont exposés en fait à un
mélange de composés volatils (par exemple de l'acétate d'éthyle, de
la méthyléthylcétone, du toluène, de l'acétone et des hydrocarbures
aliphatiques autres que le n-hexane). La concentration
atmosphérique des autres solvants présents est rarement précisée.
Dans la présente mise au point, on ne prend en considération que
celles des études qui donnent les concentrations atmosphériques.
Les résultats de l'étude électrophysiologique des nerfs
périphériques peuvent varier considérablement selon la technique
utilisée, la température du laboratoire dans lequel les mesures
sont effectuées, la portion du nerf étudiée et l'âge du sujet.
Aussi est-il difficile d'apprécier la portée des études qui
reposent sur ces méthodes, en l'absence de renseignements sur tous
ces facteurs.
L'étude de 93 cas de neuropathie a permis de caractériser le
tableau clinique de la neuropathie due au n-hexane et a montré que
les données électrophysiologiques étaient en bonne corrélation avec
la gravité de la maladie. On a en particulier constaté que plus
l'exposition était importante plus la maladie était grave.
Toutefois il n'a pas été possible d'établir une relation bien
définie à partir des données limitées dont on disposait sur
l'exposition effective.
On a observé de légers signes infracliniques de neuropathie
chez des ouvriers d'une fabrique de chaussures exposés à une
concentration de 70 mg/m3 (20 ppm). L'importance des anomalies
électrophysiologiques augmentait avec la durée de l'exposition.
Toutefois aucun cas de neuropathie déclarée n'a été observé même
chez les ouvriers qui avaient été exposés pendant plus de cinq ans.
On a observé des signes subjectifs et électrophysiologiques de
neuropathie périphérique infraclinique dans un groupe d'ouvriers
exposés à une dose d'environ 317 mg/m3 (90 ppm). Toutefois, on ne
peut pas attribuer entièrement ces effets à l'exposition au
n-hexane étant donné que l'on a probablement sous-estimé les
expositions antérieures à divers solvants et que les ouvriers
avaient probablement introduit eux-mêmes un biais sélectif. En
outre, il peut y avoir eu exposition simultanée à de la
méthyléthylcétone, dont on sait qu'elle accroît les effets
neurotoxiques du n-hexane.
L'exposition à un mélange de solvants contenant du n-hexane à
la concentration d'environ 690 mg/m3 (196 ppm) a entraîné une
réduction de la vitesse de conduction nerveuse périphérique,
anomalie qui s'est maintenue même après une réduction importante de
l'exposition. En outre, on pense que les variations observées dans
les potentiels évoqués somatosensoriels s'expliquent par l'effet du
n-hexane sur le système nerveux central.
Des signes subjectifs et électrophysiologiques d'une
neuropathie infraclinique légère ont été observés dans un petit
groupe de travailleurs exposés à une concentration moyenne de
n-hexane dans l'atmosphère de 208 mg/m3 (59 ppm). Aucun signe
clinique de neuropathie n'était perceptible. L'exposition à
d'autres solvants était limitée et on a pu procéder à des études de
conduction nerveuse dans des conditions bien définies. Toutefois,
le faible effectif des ouvriers, l'absence de données
rétrospectives sur l'exposition et le fait que les groupes témoins
n'étaient pas très bien appariés au groupe exposé limitent la
portée des conclusions qu'on peut en tirer. Néanmoins, les
résultats sont en bon accord général avec les données fournies par
les études antérieures dont il a été question plus haut. On a
attribué le déclenchement d'une neuropathie périphérique
cliniquement manifeste à l'exposition pendant des périodes
supérieures à huit heures par jour à une concentration de n-hexane
de 669 mg/m3 (190 ppm). Au bout d'un certain nombre d'années, on
assiste à une lente restauration - encore que partielle - de la
fonction nerveuse. L'exposition à une concentration inférieure à
352 mg/m3 (100 ppm) n'a pas entraîné de neuropathie mais l'étude
électrophysiologique a révélé la présence d'effets infracliniques.
Chez un petit nombre de travailleurs exposés de par leur
profession à toute une gamme de concentrations atmosphériques de
n-hexane, on a observé une perturbation de la perception des
couleurs et des signes de maculopathie. La mesure des potentiels
évoqués visuels et l'électrorétinogramme ont révélé chez ces sujets
la présence d'anomalies par comparaison avec un groupe témoin non
exposé. On a également signalé l'existence d'anomalies dans les
potentiels évoqués visuels chez des renifleurs de colle exposés à
du n-hexane. On pense qu'il pourrait se produire une dégénérescence
axonale au niveau des voies optiques et un bloc de conduction au
niveau central. On est également fondé à croire que les signes de
maculopathie pourraient être corrélés avec les signes
électrophysiologiques de neuropathie périphérique. Certains des
sujets étudiés présentaient également un électroencéphalogramme
anormal.
2.2 Autres effets
Aucune étude n'a été publiée au sujet d'effets éventuels sur
les testicules et la fertilité des mâles. On ne dispose que de très
peu d'informations sur la toxicité aiguë du n-hexane pour l'homme.
La plupart des études toxicologiques portaient sur l'exposition à
des mélanges de solvants.
A la lumière des données disponibles, il semble que le
n-hexane n'ait qu'une faible toxicité aiguë.
Très peu d'études portant sur des volontaires ont été publiées
à propos du pouvoir irritant de n-hexane pour la peau et les yeux.
Le n-hexane est légèrement irritant et il provoque l'apparition
d'un érythème passager après un bref contact avec la peau. Des
effets plus graves (érythème et formation de phlyctènes) ont été
observés après un contact cutané de cinq heures avec de l'hexane de
qualité commerciale, la zone touchée étant recouverte. On n'a pas
suffisamment de données pour se prononcer sur le pouvoir irritant
de l'hexane liquide pour l'oeil humain. Lors d'une étude au cours
de laquelle des volontaires ont été exposés à de la vapeur d'hexane
à une concentration de 1760 mg/m3 (500 ppm) pendant trois à cinq
minutes, on a relevé aucun signe d'irritation oculaire. Il n'a pas
non plus été fait état de sensibilisation cutanée chez des ouvriers
exposés, ni lors d'un test de sensibilisation maximale effectué
avec ce produit.
3. Effets sur l'environnement
Le n-hexane est peu soluble dans l'eau et extrêmement volatil.
Les organismes aquatiques sont donc peu exposés à cette substance;
une exposition passagère peut cependant se produire en cas de
décharge sauvage dans les eaux superficielles. Les données dont on
dispose sur la toxicité du n-hexane pour les organismes aquatiques
sont très peu nombreuses; elles sont très diverses et ont été
obtenues dans des conditions peu satisfaisantes; dans ces
conditions, elles ne permettent pas d'évaluer convenablement le
risque pour l'environnement.
RECOMMANDATIONS
1. Protection de la santé humaine
Les concentrations de n-hexane auxquelles la population est
exposée dans son ensemble, ne représentent pour elle aucun danger.
Dans les ambiances de travail, il convient de maintenir les teneurs
dans l'atmosphère endessous des limites recommandées pour
l'exposition professionnelle en recourant à des procédés
convenablement étudiés et en utilisant certains dispositifs
mécaniques, notamment la ventilation. Dans les milieux confinés, en
cas d'urgence et lors de certaines opérations d'entretien, il est
souhaitable que des vêtements spéciaux et une protection
respiratoire appropriée soient mis à la disposition du personnel.
Il est recommandé d'utiliser un appareil respiratoire autonome à
débit continu pour des concentrations de n-hexane allant jusqu'à
8800 mg par m3 (2500 ppm). Entre 8800 mg/m3 (2500 ppm) et 17 600
mg/m3 (5000 ppm), l'appareil respiratoire doit comporter un masque
facial complet. Les gants protecteurs résistent, semble-t-il, à la
pénétration du n-hexane mais certains mélanges de n-hexane avec
d'autres solvants tels que la méthyléthylcétone pourraient
faciliter la pénétration du n-hexane à travers le tissu des gants
ou des vêtements protecteurs. C'est un facteur à prendre en compte
lorsqu'on utilise du n-hexane en mélange avec d'autres solvants.
2. Protection de l'environnement
Il est peu probable que le n-hexane présente un danger pour
l'environnement, sauf en cas de décharge ou de déversement très
importants, auquel cas des effets passagers peuvent se produire.
RECHERCHES A EFFECTUER
a) Il faudrait étudier la fonction testiculaire au moyen d'un
certain nombre d'indices parmi des groupes de travailleurs
exposés de par leur profession, afin d'établir si l'un ou la
totalité des effets observés chez l'animal se produisent
également chez l'homme.
b) Il conviendrait d'étudier plus à fond in vivo les effets
génotoxiques du n-hexane afin de compléter les données
limitées obtenues in vitro.
c) Il est nécessaire de procéder à une étude comportant une
exposition répétée au n-hexane afin d'établir quelle est sa
toxicité par voie orale. C'est à partir des résultats obtenus
qu'on pourra déterminer la dose sans effet nocif observable qui
permettra de fixer une DJA.
d) Le mécanisme de la neurotoxicité reste obscur au niveau
moléculaire. L'élucidation de ce mécanisme est d'un intérêt
direct en vue d'une estimation valable du risque encouru. Des
études in vivo et in vitro sont nécessaires pour étudier
plus à fond et clarifier ces mécanismes.
d) Les études de tératogénicité existantes ne permettent pas
d'évaluer le risque. D'autres études sont à entre-prendre pour
que l'on puisse disposer de données utilisables.
f) On n'a pas pu établir de relations dose-réponse nettes en ce
qui concerne les effets neurotoxiques du n-hexane chez l'homme
car les données relatives à l'exposition sont soit incomplètes
soit peu fiables. En outre, même si l'on a amplement démontré
l'existence d'effets neurotoxiques périphériques, les effets
sur le système nerveux central n'ont pas été suffisamment
étudiés. Il importe d'entreprendre d'autres études
épidémiologiques, notamment des études longitudinales
prospectives, afin de combler les lacunes des travaux
antérieurs.
RESUMEN
El n-hexano (hexano normal) es un líquido volátil e incoloro.
El hexano comercial está formado principalmente por una mezcla de
isómeros del hexano y de compuestos afines de seis átomos de
carbono; su contenido de n-hexano oscila entre el 20 y el 80%. Un
sistema adecuado para la medida del n-hexano es la combinación de
la cromatografía de gases con la detección de ionización por
conductor o la espectroscopía de masa. Los límites de la exposición
profesional varían entre 100 - 1800 mg/m3 (promedio ponderado en el
tiempo) y 400 - 1500 mg/m3 (valor máximo) en diversos países.
El n-hexano se puede aislar del gas natural y del petróleo
crudo. Se utiliza en la elaboración de alimentos, inclusive en la
extracción de aceite vegetal, y como disolvente en varios productos
y procesos.
Una vez liberado en el medio ambiente, el n-hexano se encuentra
fundamentalmente en fase de vapor. Se estima que su semivida en la
atmósfera es de unos 2 días, de acuerdo con su reactividad con el
radical OH solo. Los valores descritos de la CL50 para los
organismos acuáticos son pocos y variables, y se han determinado en
condiciones inadecuadas; por consiguiente, no es posible valorar
los efectos tóxicos del n-hexano en el medio ambiente. Su baja
solubilidad en agua y su elevada volatilidad hacen improbable la
exposición de los organismos acuáticos, salvo en el caso de vertido
incontrolado en aguas de superficie.
En los mamíferos, el n-hexano se absorbe rápidamente a través
de los pulmones y se distribuye por todo el cuerpo adulto, así como
en los tejidos fetales. La absorción cutánea es limitada. El
n-hexano sufre un metabolismo oxidativo y da diversos compuestos,
entre los que se encuentra la 2,5-hexanodiona, considerada el
agente neurotóxico final. En el nervio ciático de las ratas se
pueden encontrar niveles particularmente elevados de n-hexano y de
2,5-hexanodiona. La mayor parte del n-hexano se elimina inalterado
en el aire exhalado; otra parte se excreta metabolizado en el aire
exhalado y la orina.
El n-hexano administrado a ratas adultas por vía oral o
respiratoria tiene una toxicidad aguda baja. Para una hora de
exposición se han registrado valores de DL50 de 15 - 30 g/kg por
vía oral y de 271 040 mg/m3 (77 000 ppm) por vía respiratoria. En
concentraciones altas de vapor, los animales muestran ataxia,
convulsiones y signos de depresión del sistema nervioso central.
Parece ser que el efecto principal de la exposición repetida al
n-hexano en ratas es la aparición de lesiones testiculares y
neurotoxicidad. La exposición por vía respiratoria al n-hexano y
por vía oral a la 2,5-hexanodiona produjeron lesiones testiculares
graves. Estos efectos se han atribuido a la alteración del
citoesqueleto de las células de Sertoli. Se producen efectos
secundarios en las células germinales postespermatogónicas, que
desaparecen de los túbulos afectados. Los efectos testiculares
fueron reversibles tras la exposición única durante 24 h a 17 600
mg/m3 (5000 ppm) pero irreversibles tras exposiciónes de 16 h/día y
6 días/semana durante dos semanas a la misma concentración. La
ingestión de agua de bebida con un 1% de 2,5-hexanodiona durante
dos o tres semanas produjo lesiones testiculares reversibles
análogas y efectos irreversibles (en un plazo de 17 semanas) tras
cinco semanas de tratamiento.
El efecto neurotóxico se caracteriza clínicamente por debilidad
en las extremidades posteriores, que puede llegar a la parálisis.
En el sistema nervioso central y periférico se produce inflamación
axonal; se pueden presentar lesiones más graves (degeneración y
pérdida axonal) sobre todo en los nervios más largos y de diámetro
mayor. En estudios de inhalación prácticamente continua durante
seis meses con dosis de 1760 mg/m3 (500 ppm) o superiores se
produjeron lesiones en el sistema nervioso periférico y central,
pero no se observaron efectos clínicos o patológicos a
concentraciones de 440 mg/m3 (125 ppm). De 15 a 22 semanas después
de cesar la exposición constante a una concentración de vapor de
3520 mg por m3 (1000 ppm), cinco días a la semana durante 11
semanas, sólo se había producido una recuperación limitada de la
amplitud de la respuesta evocada por un estímulo auditivo del
quinto par craneal (que se considera que indica actividad del
sistema nervioso central) y del potencial de acción del nervio de
la cola. La exposición discontinua de ratas a una concentración de
3168 mg/m3 (900 ppm) durante 72 semanas no causó ninguna lesión
evidente en el sistema nervioso periférico o central, pero se
observaron indicios de efectos electrofisiológicos en los nervios
periféricos.
La neurotoxicidad inducida por el n-hexano puede aumentar con
la exposición simultánea a metiletilcetona, metilisobutilcetona y
acetato de plomo y disminuir con la exposición simultánea al
tolueno. El tolueno y el n-hexano ejercen también un efecto
sinérgico en la alteración de los niveles de dopamina.
Tras la aplicación cutánea de n-hexano en condiciones
oclusivas durante períodos cortos se observó en la piel la
aparición de lesiones microscópicas graves. La exposición
prolongada a una concentración de vapor de n-hexano de 10 560
mg/m3 (3000 ppm) puede ocasionar irritación de la conjuntiva en la
rata y pronunciada irritación ocular en el conejo. No se dispone de
datos sobre sensibilización cutánea a partir de estudios en
animales.
Se ha informado de la aparición de lesiones cromosómicas
(poliploidía en un estudio y aberraciones estructurales en otro)
en experimentos tanto in vivo como in vitro. En las pruebas
realizadas para determinar las lesiones en el ADN no se ha
observado aumento de la frecuencia de las mutaciones puntuales ni
de los efectos en ellas.
Se ha realizado un estudio de la carcinogenicidad del n-hexano
(pintando la piel de ratones), que resultó negativo.
No se ha estudiado lo bastante la toxicidad del n-hexano para
la reproducción. No se observaron indicios claros de que su
inhalación tenga en las ratas efectos embriotóxicos o
teratogénicos, aunque las concentraciones utilizadas fueron
relativamente bajas, y tampoco en ratones después de la
administración oral. Cuando, en un estudio con ratas, se expusieron
a las madres a una concentración de vapor de n-hexano de 3520
mg/m3 (1000 ppm), se retrasó de forma pasajera el desarrollo
postnatal de la descendencia.
Se dispone de muy poca información acerca de la toxicidad
aguda del n-hexano en la especie humana. La mayor parte de los
estudios se han centrado en la exposición profesional a mezclas de
disolventes. Los datos disponibles indican que el n-hexano tiene
una toxicidad aguda escasa. Se han señalado manifestaciones de
depresión del sistema nervioso central, como somnolencia, vértigo y
mareos, tras la exposición a n-hexano comercial en concentraciones
que van de 3520 a 17 600 mg/m3 (1000 - 5000 ppm) durante 10 - 60
minutos.
El n-hexano es una sustancia ligeramente irritante que al
entrar en contacto con la piel del hombre durante períodos cortos
produce un eritema pasajero. Se ha informado de efectos más graves
(eritema y vesículas) tras un contacto cutáneo oclusivo de cinco
horas con un hexano de tipo comercial. No se han notificado casos
de sensibilización cutánea en trabajadores expuestos, y tampoco se
observó en un ensayo de maximización con n-hexano.
Con exposiciones repetidas, el n-hexano es neurotóx-ico e
induce un tipo de neuropatía periférica sensomotora. Se han
publicado muchos estudios sobre la prevalencia de la neurotoxicidad
inducida por el n-hexano; sin embargo, a menudo se carece de datos
adecuados sobre la exposición. La exposición a concentraciones de
n-hexano en el aire que oscilan entre 106 y 8800 mg/m3 (30 - 2500
ppm) se ha relacionado con neuropatías. Se han descrito casos de
acusada neuropatía periférica entre trabajadores japoneses de la
industria de las sandalias y en tipógrafos de Taiwán expuestos a
niveles de n-hexano de unos 176 y 352 mg/m3 (50 y 100 ppm),
respectivamente, durante períodos superiores a 8 h diarias. En
muchos casos las mediciones de la exposición eran recientes y no
reflejaban de manera exacta las exposiciones anteriores causantes
de neuropatía.
En varios estudios cruzados se han observado por separado
efectos subclínicos ligeros (por ejemplo, cambios
electrofisiológicos en los nervios periféricos) en trabajadores
expuestos a 70 - 352 mg/m3 (20 - 100 ppm). En ninguno de esos
estudios se identificaron casos clínicamente evidentes de
neuropatía periférica con niveles de exposición inferiores a 352
mg/m3 (100 ppm).
Sólo en un pequeño número de estudios se han investigado los
efectos del n-hexano en el sistema nervioso central. Se ha
indicado que los cambios del potencial inducido por estímulos
somatosensoriales en trabajadores expuestos al n-hexano podrían
deberse a un bloqueo de la conducción del sistema nervioso central.
También se ha observado una alteración del potencial inducido por
estímulos visuales, así como modificaciones
electroencefalográficas. Estos resultados parecen indicar que el
n-hexano puede producir una disfunción del sistema nervioso
central. Sin embargo, los datos disponibles no aportan información
relativa a los niveles de exposición correspondientes.
EVALUACION DE LOS RIESGOS PARA LA SALUD HUMANA Y DE LOS EFECTOS EN
EL MEDIO AMBIENTE
1. Estudios en animales
1.1 Exposición a corto y largo plazo
El n-hexano tiene una toxicidad aguda baja; se han comunicado
valores de DL50 del orden de 15 - 30 g/kg y una CL50 (1 hora) de
271 040 mg/m3.
En los estudios de inhalación prácticamente continua en ratas
se produjo la muerte tras una exposición a 3520 mg/m3 (1000 ppm) de
n-hexano durante un período máximo de 11 semanas y se redujo la
ganancia de peso corporal después de tres semanas de exposición a
1760 mg/m3 (500 ppm). En estudios con una exposición diaria de 12
horas al n-hexano, se produjo la muerte con una concentración de
10 560 mg/m3 (3000 ppm) al cabro de 15 semanas, y una ligera
disminución de la ganancia de peso corporal con 740 mg/m3 (200 ppm)
después de 24 semanas. No se observaron efectos en el peso corporal
en ratas expuestas a 443 mg/m3 (126 ppm) de manera prácticamente
continua durante 34 semanas.
Los principales efectos no letales de la exposición repetida de
ratas al n-hexano parecen ser la toxicidad testicular y la
neurotoxicidad, tanto periférica como central. La exposición
respiratoria al n-hexano y la oral a la 2,5-hexanodiona producen
lesiones testiculares graves. Los efectos se han atribuido a la
alteración del citoesqueleto de las células de Sertoli. Hay efectos
secundarios en las células germinales postespermatogónicas, que
desaparecen de los túbulos afectados. Los efectos testiculares
fueron reversibles tras una exposición única de 24 horas a 17 600
mg/m3 (5000 ppm), pero se hacían irreversibles después de dos
semanas de exposición a la misma concentración durante 16 horas/día
y 6 días/semana. El agua de bebida con un 1% de 2,5-hexanodiona
produjo lesiones testiculares reversibles después de 2 - 3 semanas
de administración, y efectos irreversibles (en un plazo de 17
semanas) tras 5 semanas de tratamiento.
Clínicamente, la neurotoxicidad se caracteriza por debilidad en
las extremidades posteriores, que puede evolucioner hasta parálisis
y por último tetraplejía. Se han observado manifestaciones clínicas
tras 10 semanas de exposición discontinua a 10 560 mg/m3 (3000 ppm)
(12 horas/día, 7 días/semana) y después de 6 - 10 semanas de
exposición continua a una concentración de 1408-2112 mg/m3 (400 -
600 ppm).
Antes de la aparición de debilidad en las extremidades
posteriores, se observaron lesiones (inflamación axonal) en los
nervios de dichas extremidades, en el cerebro y en la médula
espinal. Tras seis meses de exposición a 1760 mg/m3 (500 ppm) se
apreciaron efectos más graves (degeneración y pérdida axonal),
principalmente en los nervios más largos y de diámetro mayor. En
conjunto, resulta difícil determinar el nivel de no observación de
efectos adversos correspondiente a la neurotoxicidad inducida por
el n-hexano en ratas, a causa de la variedad de concentraciones de
vapor y de tipos de exposición en las pruebas, y de la diversidad
de efectos investigados. En estudios de exposición por inhalación
prácticamente continua durante seis meses, se observaron lesiones
nerviosas con concentraciones de n-hexano de 1760 mg por m3 (500
ppm) o más, pero no se manifestaron efectos clínicos o patológicos
a 440 mg/m3 (125 ppm). La exposición discontinua de ratas a una
concentración de 3168 mg/m3 (900 ppm) durante 72 semanas no causó
ninguna lesión nerviosa manifiesta, pero hubo indicios preliminares
de un efecto en la fuerza prensil.
Cuando se interrumpe la exposición pueden desaparecer (parcial
o totalmente) los signos clínicos de toxicidad, la disminución de
la fuerza prensil y los efectos electrofisiológicos. De 15 a 22
semanas después de cesar la exposición repetida durante 11 semanas
a una concentración de 3520 mg/m3 (1000 ppm), sólo se observó una
recuperación limitada de la amplitud de la respuesta inducida por
los estímulos auditivos en el quinto componente del tronco
encefálico (que se considera que refleja la actividad del sistema
nervioso central) y del potencial de acción del nervio caudal. No
se ha investigado la desaparición de las lesiones patológicas
después de la exposición por vía respiratoria, pero se ha observado
remielinización y regeneración axonal limitada al interrumpirse la
exposición a concentraciónes de 1408 a 2112 mg/m3 (400 - 600 ppm).
El tratamiento discontinuo durante siete semanas por vía
respiratoria con 1760 mg/m3 (500 ppm) puede causar ligeras lesiones
pulmonares en la rata. En conejos se apreciaron, tras la exposición
a 10 560 mg/m3 (3000 ppm) 8 horas/día durante 8 días, lesiones
graves en el tracto respiratorio y, después de 24 semanas de
exposición, proliferaciones papilares de células bronquiolares no
ciliadas.
En algunos estudios de inhalación en ratas se han señalado
efectos en el riñón y el hígado, pero solo con niveles de
exposición más elevados. La mayor parte de las lesiones y otros
efectos se consideraron de escasa importancia toxicológica y/o no
se reprodujeron en estudios repetidos. Sin embargo, tras la
administración intraperitoneal repetida de n-hexano a ratas se han
observado lesiones hepáticas graves.
Los estudios de exposición combinada han puesto de manifiesto
que los efectos neurotóxicos del vapor de n-hexano en las gallinas
pueden aumentar con la presencia de metilisobutilcetona y en las
ratas con la de metilisobutilcetona y acetato de plomo, mientras
que el tolueno los hace disminuir. También se ha señalado que la
exposición de ratas a la combinación con metiletilcetona aumenta
las lesiones pulmonares.
Se han realizado algunos estudios sobre los efectos de la
exposición oral o cutánea al n-hexano. La administración diaria
por vía oral de 500 mg/kg durante cuatro semanas produjo en las
ratas una disminución de la velocidad de conducción en el nervio
caudal.
1.2 Genotoxicidad
Se han realizado pocas pruebas de mutagenicidad. Se ha
informado de la aparición de lesiones cromosómicas (poliploidía en
un estudio, y aberraciones estructurales en otro) en ensayos in
vitro, en los que se expuso una línea celular de hámster chino a
n-hexano líquido no diluido en ausencia de fracción metabolizante.
Tras la exposición de células cultivadas de mamíferos o de
bacterias a n-hexano no se produjo un aumento significativo del
índice de mutaciones puntuales. Las lesiones en el ADN, medidas en
ensayos de toxicidad diferencial en bacterias, no aumentaron por la
exposición al n-hexano. Tampoco observó aumento de la síntesis no
programada del ADN en linfocitos humanos in vitro. Aunque el
n-hexano ha dado resultados negativos en los ensayos para detectar
mutaciones puntuales, hay algunas pruebas de que puede producir
aberraciones cromosómicas in vitro. En un estudio in vivo con
células de médula ósea de rata se informó de lesiones cromosómicas
(roturas cromatídicas).
1.3 Carcinogenicidad
No se ha estudiado de manera adecuada el potencial
carcinogénico del n-hexano. En un estudio limitado de aplicación
en la piel a ratones, no pareció que aumentase la formación de
tumores cutáneos tras la exposición a un iniciador cancerígeno
(7,12-dimetilbenzantraceno) o junto con un agente promotor (aceite
de crotón).
1.4 Efectos en la reproducción
No se ha investigado debidamente la toxicidad del n-hexano
para la reproducción. No hay pruebas sólidas de embriotoxicidad o
teratogenicidad en las ratas tras su inhalación, ni tampoco en los
ratones después de la administración oral. En ratas expuestas a una
concentración de 10 560 mg/m3 (3000 ppm) se observó un ligero
aumento de la incidencia de algunas anomalías, pero las
deficiencias de método del estudio (por ejemplo, es probable que
las concentraciones ensayadas no fueran lo bastante altas) limita
el valor de las conclusiones. En ratones se detectaron signos de
fetotoxicidad (menor peso de las crías), pero sólo con dosis que
provocaron una letalidad considerable en las madres. En un estudio
con crías de ratas expuestas a una concentración de 10 560 mg/m3
(3000 ppm) se informó de un retraso pasajero del desarrollo
postnatal. Aunque no se describieron signos de neuropatía en esos
animales, no es posible evaluar a partir de ese estudio si la
podría inducir la exposición in utero, puesto que los animales sólo
estuvieron expuestos al n-hexano durante una pequeña parte del
período de gestación. En otros informes se ha afirmado que la
exposición in utero al n-hexano retarda la histogénesis cerebelar y
la maduración neuronal e induce anomalías electrofisiológicas en el
sistema nervioso central de ratas recién nacidas. Sin embargo, este
trabajo no se ha expuesto con suficiente detalle para hacer una
evaluación completa.
Existen pruebas de que el n-hexano causa esterilidad
irreversible en las ratas. En un estudio limitado de la letalidad
dominante en ratones no se registraron efectos en la fecundidad del
macho. No se han comunicado investigaciones acerca de la fertilidad
de las hembras ni estudios multigeneracionales.
2. Estudios en la especie humana
2.1 Efectos en el sistema nervioso
En muchos estudios se ha relacionado la exposición laboral al
n-hexano con la frecuencia de la neuropatía periférica en la
especie humana. Sin embargo, pocos de esos estudios informaban
acerca de las concentraciones en el aire, los datos comunicados
pueden ser inexactos y a menudo no está claro si esas
concentraciones eran de hexano o de n-hexano. Además, la
exposición industrial al n-hexano suele darse al utilizar productos
a base de disolventes, de manera que los trabajadores están
expuestos a una mezcla de compuestos volátiles (por ejemplo,
acetato de etilo, metiletilcetona, tolueno, acetona y otros
hidrocarburos alifáticos). Raramente se ha señalado la
concentración en el aire de los otros disolventes presentes. En el
presente examen sólo se han incluido los estudios que contenían
algunos datos de las concentraciones en el aire.
Los resultados de la electrofisiología de los nervios
periféricos pueden variar de manera considerable en función del
tipo de técnica utilizada, la temperatura ambiente a la que se
efectúan las mediciones, el segmento del nervio estudiado y la edad
del individuo. Así pues, sin información sobre esos factores es
difícil evaluar los estudios que se basan en la electrofisiología
de los nervios periféricos.
Mediante el estudio de 93 casos de neuropatía se definieron las
características clínicas de la producida por el n-hexano y se puso
de manifiesto que existe una correlación clara entre las medidas
electrofisiológicas y la gravedad de trastorno. Se encontró una
relación entre el aumento de la gravedad y el de la exposición. Sin
embargo, no fue posible establecer una relación definitiva a partir
de los escasos detalles notificados sobre la exposición.
En los trabajadores del calzado expuestos a una concentración
de 70 mg/m3 (20 ppm) se encontraron signos subclínicos leves de
neuropatía. El grado de anomalía electrofisiológica aumentaba con
la duración de la exposición. Sin embargo, no se produjeron casos
de neuropatía manifiesta, ni siquiera en trabajadores con una
exposición de más de cinco años de duración.
Se observaron signos subjetivos y electrofisiológicos de
neuropatía periférica subclínica en un grupo de trabajadores
expuestos a 317 mg/m3 (90 ppm). Sin embargo, esos efectos no se
pueden atribuir exclusivamente a la exposición a esta concentración
de n-hexano, puesto que probablemente se infravaloró la exposición
anterior a disolventes y la autoselección de los trabajadores puede
haber dado lugar a cierto sesgo. Además, se puede haber producido
una exposición simultánea a la metiletilcetona, que, como se ha
demostrado, aumenta los efectos neurotóxicos del n-hexano.
La exposición a una mezcla de disolventes con una concentración
aproximada de n-hexano de 690 mg/m3 (196 ppm) causó una reducción
de la velocidad de conducción de los nervios periféricos, que se
mantuvo tras disminuir la exposición de manera significativa. Se ha
indicado asimismo que la causa de algunos de los cambios de
potencial inducidos por estímulos somatosensoriales podría ser un
efecto del n-hexano en el sistema nervioso central.
En un pequeño grupo de trabajadores expuestos a una
concentración media de n-hexano en el aire de 208 mg/m3 (59 ppm)
se produjeron signos subjetivos y electrofisiológicos de una
neuropatía subclínica leve. No se detectó neuropatía clínica
manifiesta. En este estudio, la exposición a otros disolventes fue
limitada y los estudios de conducción nerviosa se realizaron en
condiciones definidas. Sin embargo, el pequeño número de individuos
estudiados, la ausencia de historial de exposición y la escasa
correspondencia entre los grupos de exposición y testigo limitan el
valor de las conclusiones alcanzadas. No obstante, los resultados
en general coinciden con los datos obtenidos en anteriores estudios
de los que se informa más arriba. La exposición a 669 mg/m3 (190
ppm) durante períodos superiores a 8 h/día se asoció con la
aparición de neuropatía periférica clínicamente manifiesta. Puede
producirse una lenta recuperación parcial de la función nerviosa a
lo largo de varios años. La exposición a concentraciones inferiores
a 352 mg/m3 (100 ppm) no produjo neuropatía, pero aparecieron
pruebas electrofisiológicas de efectos subclínicos.
En un pequeño grupo de trabajadores profesionalmente expuestos
a niveles muy diversos de n-hexano en el aire se han descrito
defectos en cuanto a la distinción de los colores y signos de
maculopatía. La medición del potencial inducido por estímulos
visuales y el electrorretinograma de esos individuos mostró
anomalías en comparación con un grupo testigo de trabajadores no
expuestos. También se ha informado de anomalías en el trazado del
potencial causado por estímulos visuales en sujetos inhaladores de
pegamento expuestos a n-hexano. Se ha indicado que puede
producirse degeneración axonal en la vía óptica y que el n-hexano
puede inducir un bloqueo de la conducción en el sistema nervioso
central. También hay algunas pruebas de que existe cierta
correlación entre los signos de maculopatía y los signos
electrofisiológicos de neuropatía periférica. Se encontraron
asimismo anomalías en el trazado electroencefalografico de algunos
de los individuos estudiados.
2.2 Otros efectos
No se conoce ningún estudio sobre la fertilidad o los posibles
efectos testiculares en los varones. Es muy escasa la información
sobre la toxicidad aguda del n-hexano en la especie humana. La
mayor parte de los estudios han consistido en la exposición a
mezclas de disolventes. Los datos disponibles parecen indicar que
el n-hexano tiene una toxicidad aguda baja.
Se han publicado muy pocos estudios sobre la irritación cutánea
y ocular en voluntarios. El n-hexano es un agente ligeramente
irritante que cuando entra en contacto durante un corto período con
la piel del hombre causa un eritema pasajero. Se han descrito
efectos más graves (eritema y vesículas) tras un contacto cutáneo
oclusivo de cinco horas con n-hexano de tipo comercial. Se carece
de datos suficientes para evaluar si el líquido puede causar en el
hombre irritación ocular. No se observaron signos de irritación de
los ojos en un estudio en que se expuso a un grupo de voluntarios a
vapor de hexano con una concentración de 1760 mg/m3 (500 ppm)
durante 3 - 5 minutos. No se han comunicado casos de
sensibilización cutánea en trabajadores expuestos y tampoco se ha
observado en una prueba de maximización con n-hexano.
3. Efectos en el medio ambiente
El n-hexano es poco soluble en agua y muy volátil. Por
consiguiente, la exposición de los organismos acuáticos es
probablemente baja y pasajera, excepto en el caso de vertidos
incontrolados en aguas de superficie. Los datos acerca de la
toxicidad del n-hexano para los organismos acuáticos son escasos,
variables, obtenidos en condiciones inapropiadas e insuficientes
para evaluar del riesgo para el medio ambiente.
RECOMENDACIONES
1. Protección de la salud humana
La población general está expuesta a unas concentraciones de
n-hexano a las que es improbable que corra algún riesgo. En
relación con la salud en el trabajo, los niveles atmosféricos se
deben mantener por debajo de los límites de exposición profesional
recomendados, mediante el diseño de procesos de trabajo adecuados y
por medio de controles técnicos, incluido el de la ventilación. En
los espacios cerrados, en casos de emergencia y en determinadas
actividades de mantenimiento se debe disponer de ropas protectoras
adecuadas y protección de la respiración. Se recomienda el uso de
respiradores con admisión de aire puro o respiradores autónomos con
flujo continuo de oxígeno para concentraciones de n-hexano de
hasta 8800 mg/m3 (2500 ppm). En el caso de concentraciones entre
8800 mg/m3 (2500 ppm) y 17 600 mg/m3 (5000 ppm), el respirador debe
llevar adaptada una protección facial completa. Aunque los guantes
pueden impedir la penetración del n-hexano, las mezclas de éste
con otros disolventes, como la metiletilcetona, pueden permitir al
n-hexano pasar a través de los guantes y de otras prendas
protectoras. Hay que tener en cuenta este factor cuando se utilicen
mezclas de n-hexano.
2. Protección del medio ambiente
No es probable que el n-hexano represente un peligro para el
medio ambiente, excepto en el caso de derrames o vertidos
importantes, que podrían provocar efectos locales pasajeros.
NUEVAS INVESTIGACIONES
a) Se debería hacer un estudio sobre los índices de la función
testicular en uno o varios grupos de trabajadores expuestos
para establecer si alguno o todos los efectos cuya presencia
se ha descrito en los testículos de los animales se manifiestan
también en el hombre.
b) Conviene seguir investigando in vivo los efectos genotóxicos
del n-hexano, a fin de complementar la limitada información
que se posee sobre estos efectos in vitro.
c) Para establecer la toxicidad del n-hexano administrado por vía
oral es necesario realizar un estudio de dosis repetidas. Es
preciso hacer esto para determinar el nivel sin efecto
observado, a fin de definir una ingesta diaria admisible (IDA).
d) No está claro el mecanismo de la neurotoxicidad a nivel
molecular. Su conocimiento contribuirá directamente a la
validez de la evaluación del riesgo. Se necesitar estudios
in vivo e in vitro para seguir investigando y aclarar esos
mecanismos.
e) Los estudios de teratogenia disponibles no bastan para evaluar
el riesgo. Conviene realizar estudios que proporcionen datos
utilizables.
f) No se conoce la relación dosis-respuesta en cuanto a los
efectos neurotóxicos del n-hexano en la especie humana, debido
a que los datos de exposición son incompletos o no fidedignos.
Además, aunque se ha demostrado ampliamente la neurotoxicidad
periférica del n-hexano, los efectos en el sistema nervioso
central sólo se han examinado de forma limitada. Deben
realizarse más investigaciones epidemiológicas, como por
ejemplo estudios prospectivos de seguimiento, para subsanar la
escasez de datos de las anteriores investigaciones.